
Impacts of Drought and Climatic Factors on Vegetation Dynamics in the Yellow River Basin and Yangtze River Basin, China


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
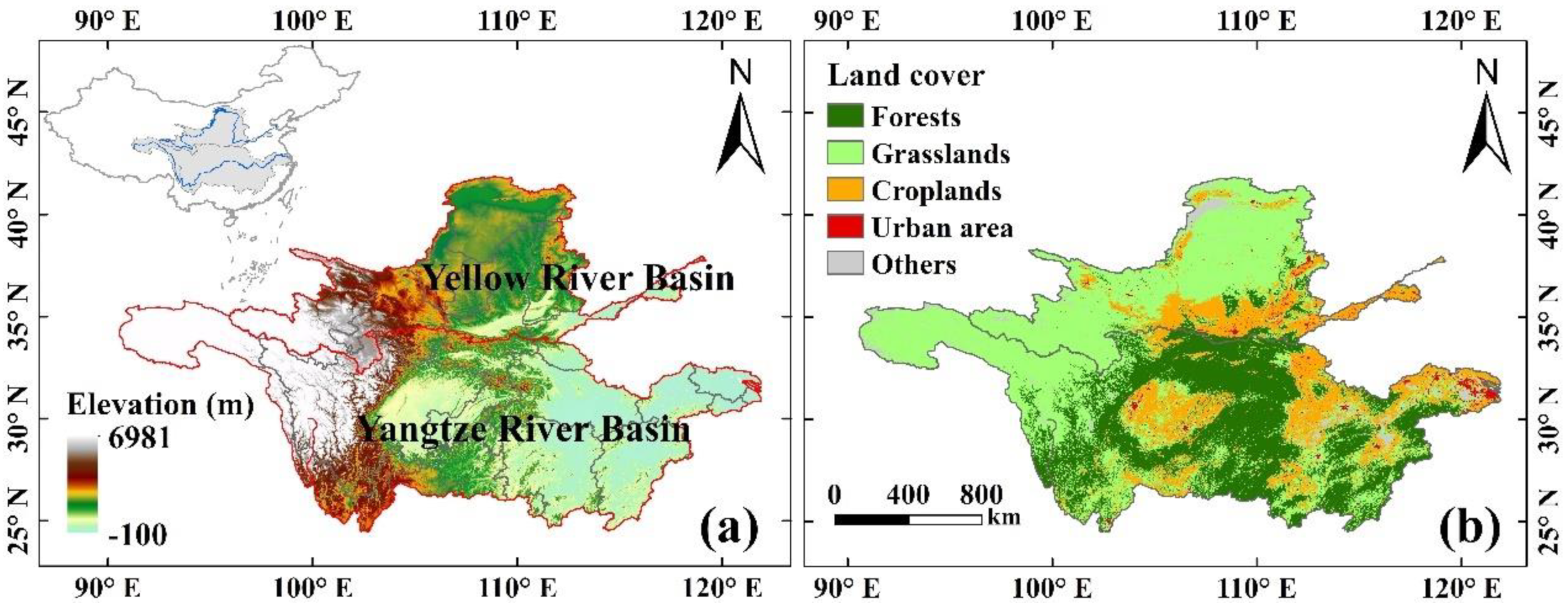
2.1. Study Area
2.2. Datasets
2.3. Methods
2.3.1. Theil–Sen Median Trend and Mann–Kendall Test
2.3.2. Pearson Correlation Analysis
2.3.3. Temporal Effects of Climatic Factors on NDVI
2.3.4. Multiple Linear Regression
2.3.5. Residual Trend Method
3. Results
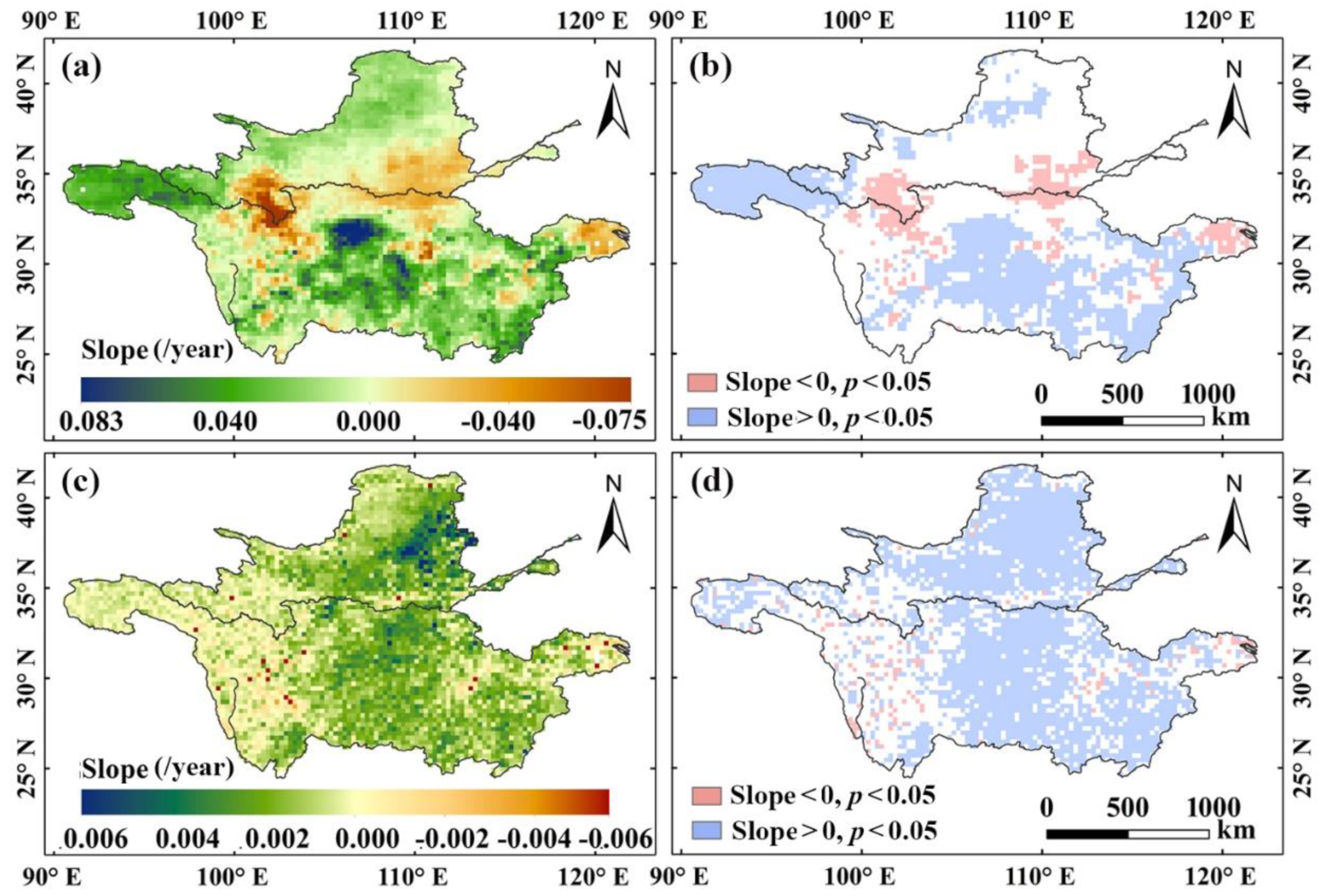
3.1. Spatiotemporal Variabilities of Drought, Climatic Factors, and Vegetation Coverage
3.2. Impacts of Drought on Vegetation Change
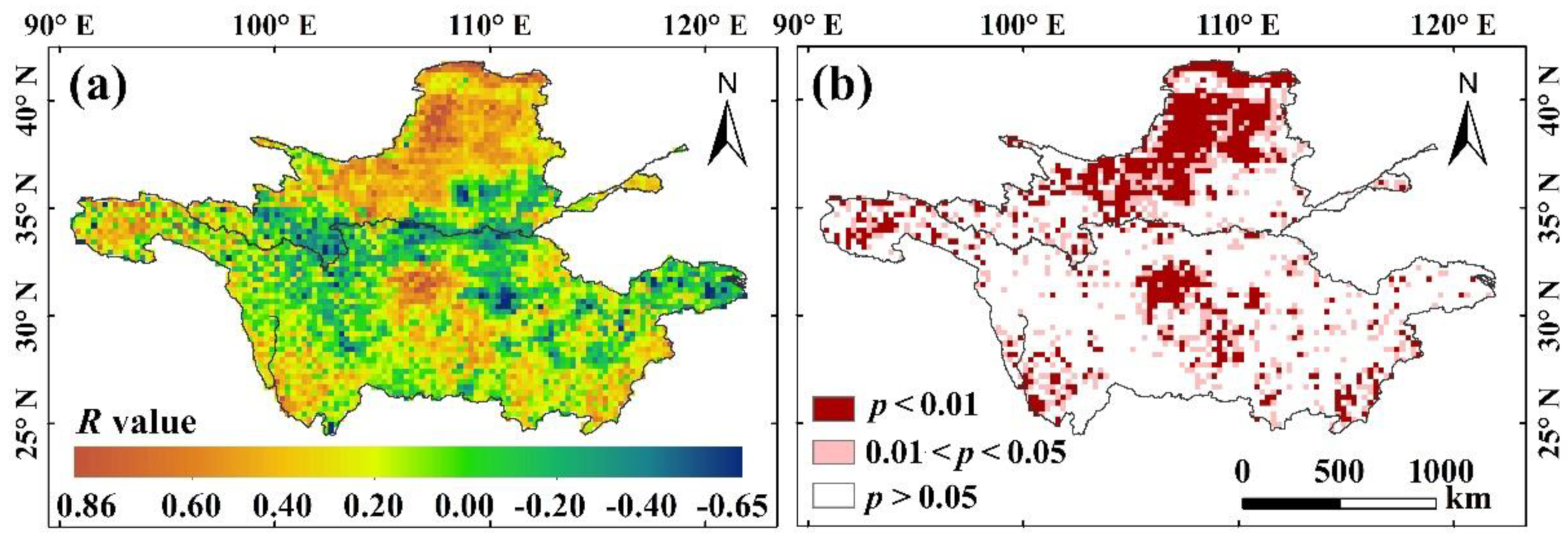
3.3. Impacts of Climatic Factors on Vegetation Changes
3.3.1. Correlation Analysis between Climatic Factors and Vegetation Changes
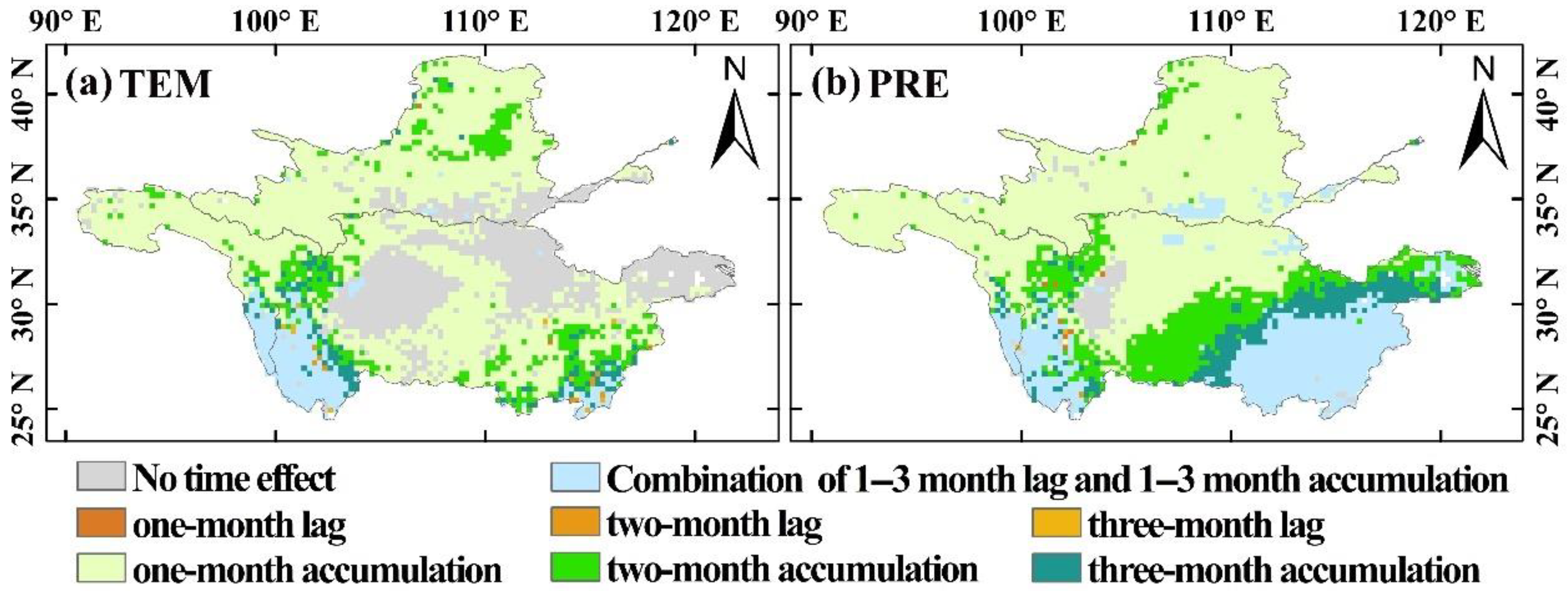
3.3.2. Temporal Effects of Climatic Factors on Vegetation Changes
4. Discussion
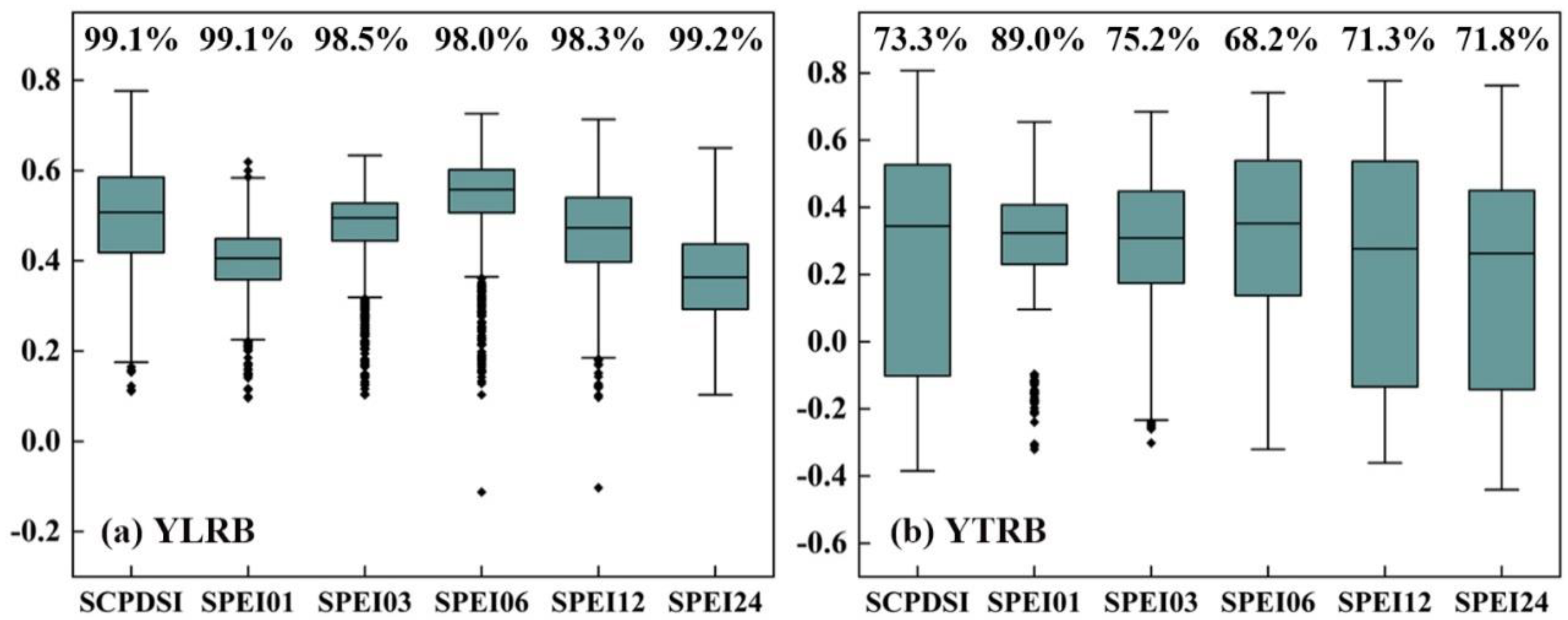
4.1. Comparison of SEDI with Other Drought Indices
4.2. Explanation of Vegetation Variation by Climatic Factors
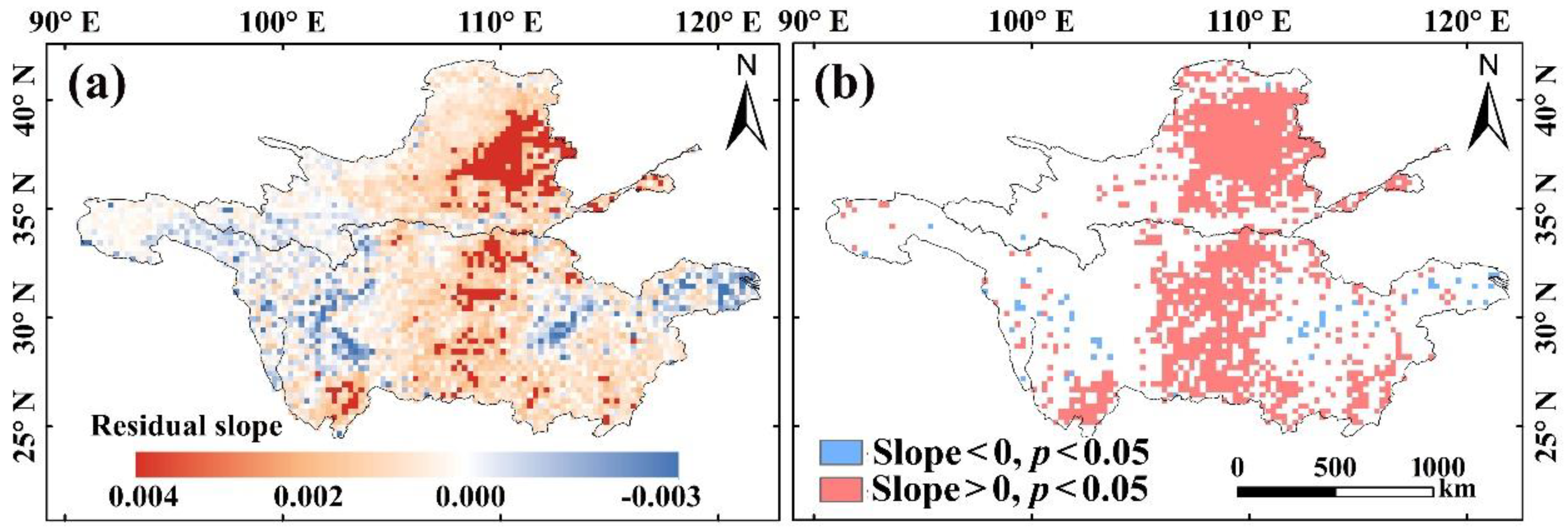
4.3. Contributions of Anthropogenic Factors to Vegetation Variation
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eekhout, J.P.C.; Boix-Fayos, C.; Pérez-Cutillas, P.; de Vente, J. The impact of reservoir construction and changes in land use and climate on ecosystem services in a large Mediterranean catchment. J. Hydrol. 2020, 590, 125208. [Google Scholar] [CrossRef]
- Wen, Z.; Wu, S.; Chen, J.; Lu, M. NDVI indicated long-term interannual changes in vegetation activities and their responses to climatic and anthropogenic factors in the Three Gorges Reservoir Region, China. Sci. Total Environ. 2017, 574, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zheng, X.; Ma, L.; Wang, H.; Huang, Q.; Leng, G.; Meng, E.; Guo, Y. Quantitative contribution of climate change and human activities to vegetation cover variations based on GA-SVM model. J. Hydrol. 2020, 584, 124687. [Google Scholar] [CrossRef]
- Jiang, L.; Guli, J.; Bao, A.; Guo, H.; Ndayisaba, F. Vegetation dynamics and responses to climate change and human activities in Central Asia. Sci. Total Environ. 2017, 599–600, 967–980. [Google Scholar] [CrossRef]
- Piao, S.; Zhang, X.; Chen, A.; Liu, Q.; Lian, X.; Wang, X.; Peng, S.; Wu, X. The impacts of climate extremes on the terrestrial carbon cycle: A review. Sci. China Earth Sci. 2019, 62, 1551–1563. [Google Scholar] [CrossRef]
- Wang, Z.; Zhong, R.; Lai, C.; Zeng, Z.; Lian, Y.; Bai, X. Climate change enhances the severity and variability of drought in the Pearl River Basin in South China in the 21st century. Agric. For. Meteorol. 2018, 249, 149–162. [Google Scholar] [CrossRef]
- Zhang, L.; Ameca, E.I.; Cowlishaw, G.; Pettorelli, N.; Foden, W.; Mace, G.M. Global assessment of primate vulnerability to extreme climatic events. Nat. Clim. Chang. 2019, 9, 554–561. [Google Scholar] [CrossRef]
- Zhao, A.; Zhang, A.; Liu, J.; Feng, L.; Zhao, Y. Assessing the effects of drought and “Grain for Green” Program on vegetation dynamics in China’s Loess Plateau from 2000 to 2014. Catena 2019, 175, 446–455. [Google Scholar] [CrossRef]
- Ding, Y.; Xu, J.; Wang, X.; Peng, X.; Cai, H. Spatial and temporal effects of drought on Chinese vegetation under different coverage levels. Sci. Total Environ. 2020, 716, 137166. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, L.; Feng, L.; Zhang, M.; Yao, R. Drought characteristics and its impact on changes in surface vegetation from 1981 to 2015 in the Yangtze River Basin, China. Int. J. Climatol. 2019, 40, 3380–3397. [Google Scholar] [CrossRef]
- Zhao, A.; Yu, Q.; Feng, L.; Zhang, A.; Pei, T. Evaluating the cumulative and time-lag effects of drought on grassland vegetation: A case study in the Chinese Loess Plateau. J. Environ. Manag. 2020, 261, 110214. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Lai, C. Severe drought events inducing large decrease of net primary productivity in mainland China during 1982–2015. Sci. Total Environ. 2020, 703, 135541. [Google Scholar] [CrossRef]
- Easterling, D.R.; Wallis, T.W.R.; Lawrimore, J.H.; Heim, R.R. Effects of temperature and precipitation trends on U.S. drought. Geophys. Res. Lett. 2007, 34, L20709. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Apps, D.; Arcand, S.; Xu, H.; Pan, M.; Hoerling, M. Contribution of temperature and precipitation anomalies to the California drought during 2012–2015. Geophys. Res. Lett. 2017, 44, 3184–3192. [Google Scholar] [CrossRef]
- Yang, M.; Mou, Y.; Meng, Y.; Liu, S.; Peng, C.; Zhou, X. Modeling the effects of precipitation and temperature patterns on agricultural drought in China from 1949 to 2015. Sci. Total Environ. 2020, 711, 135139. [Google Scholar] [CrossRef]
- Sun, C.; Ma, Y. Effects of non-linear temperature and precipitation trends on Loess Plateau droughts. Quat. Int. 2015, 372, 175–179. [Google Scholar] [CrossRef]
- Wang, D.; Alimohammadi, N. Responses of annual runoff, evaporation, and storage change to climate variability at the watershed scale. Water Resour. Res. 2012, 48, W05546. [Google Scholar] [CrossRef] [Green Version]
- Kong, D.; Miao, C.; Wu, J.; Zheng, H.; Wu, S. Time lag of vegetation growth on the Loess Plateau in response to climate factors, Estimation, distribution, and influence. Sci. Total Environ. 2020, 744, 140726. [Google Scholar] [CrossRef]
- Li, P.; Zhu, D.; Wang, Y.; Liu, D. Elevation dependence of drought legacy effects on vegetation greenness over the Tibetan Plateau. Agric. For. Meteorol. 2020, 295, 108190. [Google Scholar] [CrossRef]
- Zhao, J.; Huang, S.; Huang, Q.; Wang, H.; Leng, G.; Fang, W. Time-lagged response of vegetation dynamics to climatic and teleconnection factors. Catena 2020, 189, 104474. [Google Scholar] [CrossRef]
- Gu, Z.; Duan, X.; Shi, Y.; Li, Y.; Pan, X. Spatiotemporal variation in vegetation coverage and its response to climatic factors in the Red River Basin, China. Ecol. Indic. 2018, 93, 54–64. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Climate 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Sun, J.; Liu, M.; Xiong, J.; Zong, N.; Hu, J.; Huang, Y.; Duan, X.; Tsunekawa, A. Concurrent and Lagged Effects of Extreme Drought Induce Net Reduction in Vegetation Carbon Uptake on Tibetan Plateau. Remote Sens. 2020, 12, 2347. [Google Scholar] [CrossRef]
- Guo, L.; Cheng, J.; Luedeling, E.; Koerner, S.E.; He, J.-S.; Xu, J.; Gang, C.; Li, W.; Luo, R.; Peng, C. Critical climate periods for grassland productivity on China’s Loess Plateau. Agric. For. Meteorol. 2017, 233, 101–109. [Google Scholar] [CrossRef]
- Guo, L.; Wang, J.; Li, M.; Liu, L.; Xu, J.; Cheng, J.; Gang, C.; Yu, Q.; Chen, J.; Peng, C.; et al. Distribution margins as natural laboratories to infer species’ flowering responses to climate warming and implications for frost risk. Agric. For. Meteorol. 2019, 268, 299–307. [Google Scholar] [CrossRef]
- Zuo, D.; Han, Y.; Xu, Z.; Li, P.; Ban, C.; Sun, W.; Pang, B.; Peng, D.; Kan, G.; Zhang, R.; et al. Time-lag effects of climatic change and drought on vegetation dynamics in an alpine river basin of the Tibet Plateau, China. J. Hydrol. 2021, 600, 126532. [Google Scholar] [CrossRef]
- Wu, D.; Zhao, X.; Liang, S.; Zhou, T.; Huang, K.; Tang, B.; Zhao, W. Time-lag effects of global vegetation responses to climate change. Glob. Chang. Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef]
- Fang, L.; Wang, L.; Chen, W.; Sun, J.; Cao, Q.; Wang, S.; Wang, L. Identifying the impacts of natural and human factors on ecosystem service in the Yangtze and Yellow River Basins. J. Clean. Prod. 2021, 314, 127995. [Google Scholar] [CrossRef]
- Omer, A.; Zhuguo, M.; Zheng, Z.; Saleem, F. Natural and anthropogenic influences on the recent droughts in Yellow River Basin, China. Sci. Total Environ. 2020, 704, 135428. [Google Scholar] [CrossRef]
- Sun, Z.; Zhu, X.; Pan, Y.; Zhang, J.; Liu, X. Drought evaluation using the GRACE terrestrial water storage deficit over the Yangtze River Basin, China. Sci. Total Environ. 2018, 634, 727–738. [Google Scholar] [CrossRef]
- Cui, L.; Wang, L.; Qu, S.; Deng, L.; Wang, Z. Impacts of temperature, precipitation and human activity on vegetation NDVI in Yangtze River basin, China. Earth Sci. 2019, 45, 1–24. [Google Scholar]
- Liu, H.; Liu, F.; Zheng, L. Effects of Climate Change and Human Activities on Vegetation Cover Change in the Yellow River Basin. J. Soil Water Conserv. 2021, 35, 143–151. [Google Scholar]
- Zhang, L.; Li, X.; Feng, J.; Rao, R.; He, T.; Chen, Y. Spatial-temporal Changes of NDVI in Yellow River Basin and Its Dual Response to Climate Change and Human Activities. Bull. Soil Water Conserv. 2021, 41, 1–11. [Google Scholar]
- Chen, S.; Wen, Z.; Zhang, S.; Huang, P.; Ma, M.; Zhou, X.; Liao, T.; Wu, S. Effects of long-term and large-scale ecology projects on forest dynamics in Yangtze River Basin, China. For. Ecol. Manag. 2021, 496, 119463. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, L.; Xiang, F.; Qin, W.; Jiang, W. Vegetation dynamics and the relations with climate change at multiple time scales in the Yangtze River and Yellow River Basin, China. Ecol. Indic. 2020, 110, 105892. [Google Scholar] [CrossRef]
- Li, Y.; Xie, Z.; Qin, Y.; Zheng, Z. Responses of the Yellow River basin vegetation: Climate change. Int. J. Clim. Chang. Strateg. Manag. 2019, 11, 483–498. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, Y.; Ma, X.; Ren, L.; Singh, V. Drought Analysis in the Yellow River Basin Based on a Short-Scalar Palmer Drought Severity Index. Water 2018, 10, 1526. [Google Scholar] [CrossRef] [Green Version]
- Qu, S.; Wang, L.; Lin, A.; Yu, D.; Yuan, M.; Li, C.A. Distinguishing the impacts of climate change and anthropogenic factors on vegetation dynamics in the Yangtze River Basin, China. Ecol. Indic. 2020, 108, 105724. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Wang, Z.; Shang, P. Drought evaluation over Yangtze River basin based on weighted water storage deficit. J. Hydrol. 2020, 591, 125283. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Miralles, D.G.; Domínguez-Castro, F.; Azorin-Molina, C.; El Kenawy, A.; McVicar, T.R.; Tomás-Burguera, M.; Beguería, S.; Maneta, M.; Peña-Gallardo, M. Global Assessment of the Standardized Evapotranspiration Deficit Index (SEDI) for Drought Analysis and Monitoring. J. Clim. 2018, 31, 5371–5393. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.; Miralles, D.; Dominguez-Castro, F.; Azorin-Molina, C.; El Kenawy, A.; McVicar, T.; Tomas-Burguera, M.; Begueria, S.; Maneta, M.; Peña-Gallardo, M. A proposed robust approach for calculating the standardized evapotranspiration deficit index (SEDI) at the global scale. In Proceedings of the First International Electronic Conference on the Hydrological Cycle, Online, 12–16 November 2017. [Google Scholar]
- Zhang, X.; Li, M.; Ma, Z.; Yang, Q.; Lv, M.; Clark, R. Assessment of an Evapotranspiration Deficit Drought Index in Relation to Impacts on Ecosystems. Adv. Atmos. Sci. 2019, 36, 1273–1287. [Google Scholar] [CrossRef]
- Zhou, X.; Yamaguchi, Y.; Arjasakusuma, S. Distinguishing the vegetation dynamics induced by anthropogenic factors using vegetation optical depth and AVHRR NDVI: A cross-border study on the Mongolian Plateau. Sci. Total Environ. 2018, 616–617, 730–743. [Google Scholar] [CrossRef]
- Huang, S.; Tang, L.; Hupy, J.P.; Wang, Y.; Shao, G. A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing. J. For. Res. 2021, 32, 1–6. [Google Scholar] [CrossRef]
- Tian, J.; Wang, L.; Li, X.; Gong, H.; Shi, C.; Zhong, R.; Liu, X. Comparison of UAV and WorldView-2 imagery for mapping leaf area index of mangrove forest. Int. J. Appl. Earth Obs. Geoinform. 2017, 61, 22–31. [Google Scholar] [CrossRef]
- Dutrieux, L.P.; Verbesselt, J.; Kooistra, L.; Herold, M. Monitoring forest cover loss using multiple data streams, a case study of a tropical dry forest in Bolivia. ISPRS J. Photogramm. 2015, 107, 112–125. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, D. Improving forest aboveground biomass estimation using seasonal Landsat NDVI time-series. ISPRS J. Photogramm. 2015, 102, 222–231. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Xu, Y.; Gao, X.; Shen, Y.; Xu, C.; Shi, Y.; Giorgi, F. A daily temperature dataset over China and its application in validating a RCM simulation. Adv. Atmos. Sci. 2009, 26, 763–772. [Google Scholar] [CrossRef]
- Wu, J.; Gao, X.J. A gridded daily observation dataset over China region and comparison with the other datasets. Chin. J. Geophys. 2013, 56, 1102–1111. [Google Scholar]
- Yang, F.; Lu, H.; Yang, K.; He, J.; Wang, W.; Wright, J.S.; Li, C.; Han, M.; Li, Y. Evaluation of multiple forcing data sets for precipitation and shortwave radiation over major land areas of China. Hydrol. Earth Syst. Sci. 2017, 21, 5805–5821. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Sun, J. Evaluation of high-resolution precipitation products over southwest China. J. Hydrometeorol. 2020, 21, 2691–2712. [Google Scholar] [CrossRef]
- Rukundo, O. Nearest Neighbor Value Interpolation. Int. J. Adv. Comput. Sci. 2012, 3, 1–6. [Google Scholar]
- Sen, P.K. Estimates of the regression coefficient based on Kendall’s tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Theil, H. A rank-invariant method of linear and polynomial regression analysis. Indagat. Math. 1950, 12, 173. [Google Scholar]
- Kendall, M.G. A New Measure of Rank Correlation. Biometrika 1938, 30, 81–93. [Google Scholar] [CrossRef]
- Mann, H.B. Nonparametric Tests Against Trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Qu, S.; Wang, L.; Lin, A.; Zhu, H.; Yuan, M. What drives the vegetation restoration in Yangtze River basin, China: Climate change or anthropogenic factors? Ecol. Indic. 2018, 90, 438–450. [Google Scholar] [CrossRef]
- Chu, H.; Venevsky, S.; Wu, C.; Wang, M. NDVI-based vegetation dynamics and its response to climate changes at Amur-Heilongjiang River Basin from 1982 to 2015. Sci. Total Environ. 2019, 650, 2051–2062. [Google Scholar] [CrossRef]
- Cui, L.F.; Wang, Z.D.; Deng, L.H.; Qu, S. Vegetation Dynamics and Their Relations with Climate Change at Seasonal Scales in the Yangtze River Basin, China. Appl. Ecol. Environ. Res. 2020, 18, 3543–3556. [Google Scholar] [CrossRef]
- Ding, Y.; Li, Z.; Peng, S. Global analysis of time-lag and -accumulation effects of climate on vegetation growth. Int. J. Appl. Earth Obs. Geoinform. 2020, 92, 102179. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Begueria, S.; Trigo, R.; Lopez-Moreno, J.I.; Azorin-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Yang, Y.; Zhang, L.; Wang, Z. The relative roles of climate variations and human activities in vegetation change in North China. Phys. Chem. Earth Parts A/B/C 2015, 87, 67–78. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Z.; Yang, H.; Zhao, Y. Study of the temporal and spatial patterns of drought in the Yellow River basin based on SPEI. Sci. China Earth Sci. 2018, 61, 1098–1111. [Google Scholar] [CrossRef]
- Knapp, A.K.; Carroll, C.J.; Denton, E.M.; la Pierre, K.J.; Collins, S.L.; Smith, M.D. Differential sensitivity to regional-scale drought in six central US grasslands. Oecologia 2015, 177, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Craine, J.M.; Ocheltree, T.W.; Nippert, J.B.; Towne, E.G.; Skibbe, A.M.; Kembel, S.W.; Fargione, J.E. Global diversity of drought tolerance and grassland climate-change resilience. Nat. Clim. Chang. 2013, 3, 63–67. [Google Scholar] [CrossRef]
- Kath, J.; le Brocque, A.F.; Reardon-Smith, K.; Apan, A. Remotely sensed agricultural grassland productivity responses to land use and hydro-climatic drivers under extreme drought and rainfall. Agric. For. Meteorol. 2019, 268, 11–22. [Google Scholar] [CrossRef]
- Piao, S.; Mohammat, A.; Fang, J.; Cai, Q.; Feng, J. NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Glob. Environ. Chang. 2006, 16, 340–348. [Google Scholar] [CrossRef]
- Wang, A.; Lettenmaier, D.P.; Sheffield, J. Soil moisture drought in China, 1950–2006. J. Clim. 2011, 24, 3257–3271. [Google Scholar] [CrossRef]
- Anderegg, W.R.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Peng, S.; Lin, X.; Chang, J. Declining snow cover may affect spring phenological trend on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2013, 110, E2854–E2855. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wang, D.; Wang, G.; Qiong, M.; Zhang, G.; Ji, D.M. Vegetation cover change over the Southwest China and its relation to climatic factors. Resour. Environ. Yangtze Basin 2015, 24, 956–964. [Google Scholar]
- Wang, J.; Wang, K.; Zhang, M.; Zhang, C. Impacts of climate change and human activities on vegetation cover in hilly southern China. Ecol. Eng. 2015, 81, 451–461. [Google Scholar] [CrossRef]
- Busetto, L.; Colombo, R.; Migliavacca, M.; Cremonese, E.; Meroni, M.; Galvagno, M.; Rossini, M.; Siniscalco, C.; Morra Di Cella, U.; Pari, E. Remote sensing of larch phenological cycle and analysis of relationships with climate in the Alpine region. Glob. Chang. Biol. 2010, 16, 2504–2517. [Google Scholar] [CrossRef]
- McKee, T.B.; Doesken, N.J.; Kleist, J. The relationship of drought frequency and duration to time scales. In Proceedings of the 8th Conference on Applied Climatology, Boston, MA, USA, 17–22 January 1993. [Google Scholar]
- Wells, N.; Goddard, S.; Hayes, M.J. A self-calibrating Palmer drought severity index. J. Clim. 2004, 17, 2335–2351. [Google Scholar] [CrossRef]
- Dai, A. Characteristics and trends in various forms of the Palmer Drought Severity Index during 1900–2008. J. Geophys. Res.-Atmos. 2011, 116, D12115. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Zhang, H.; Chen, X.; Hagan, D.F.; Wang, G.; Gao, Z.; Shi, T. Robust drying and wetting trends found in regions over China based on Köppen climate classifications. J. Geophys. Res.-Atmos. 2017, 122, 4228–4237. [Google Scholar] [CrossRef]
- McEvoy, D.J.; Huntington, J.L.; Hobbins, M.T.; Wood, A.; Morton, C.; Anderson, M.; Hain, C. The evaporative demand drought index. Part II: CONUS-wide assessment against common drought indicators. J. Hydrometeorol. 2016, 17, 1763–1779. [Google Scholar] [CrossRef]
- Kim, D.; Rhee, J. A drought index based on actual evapotranspiration from the Bouchet hypothesis. Geophys. Res. Lett. 2016, 43, 10,277–10,285. [Google Scholar] [CrossRef]
- Alsafadi, K.; Al-Ansari, N.; Mokhtar, A.; Mohammed, S.; Elbeltagi, A.; Sh Sammen, S.; Bi, S. An evapotranspiration deficit-based drought index to detect variability of terrestrial carbon productivity in the Middle East. Environ. Res. Lett. 2022, 17, D12115. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, S.; Regnier, P.; Yuan, W. New insights on plant phenological response to temperature revealed from long-term widespread observations in China. Glob. Chang. Biol. 2018, 24, 2066–2078. [Google Scholar] [CrossRef]
- Ivits, E.; Horion, S.; Erhard, M.; Fensholt, R. Assessing European ecosystem stability to drought in the vegetation growing season. Glob. Ecol. Biogeogr. 2016, 25, 1131–1143. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, X.; Xin, Q.; Wu, J.; Xu, X.; Pei, F.; Li, X.; Du, G.; Cai, Y.; Lin, K.; et al. Cumulative Effects of Climatic Factors on Terrestrial Vegetation Growth. J. Geophys. Res.-Biogeo. 2019, 124, 789–806. [Google Scholar] [CrossRef]
- Long, H.; Li, X.; Bao, Y.; Huang, L.; Li, Z. Time lag analysis between vegetation and climate change in Inner Mongolia. In Proceedings of the 2010 IEEE International Geoscience and Remote Sensing Symposium, Honolulu, HI, USA, 25–30 July 2010. [Google Scholar]
- Peng, S.; Li, Z. Potential land use adjustment for future climate change adaptation in revegetated regions. Sci. Total Environ. 2018, 639, 476–484. [Google Scholar] [CrossRef]
- Zheng, K.; Wei, J.Z.; Pei, J.Y.; Cheng, H.; Zhang, X.L.; Huang, F.Q.; Li, F.M.; Ye, J.S. Impacts of climate change and human activities on grassland vegetation variation in the Chinese Loess Plateau. Sci. Total Environ. 2019, 660, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Cao, J.; Wang, L.; Zhang, W.; Wu, X. Urbanization effects on vegetation cover in major African cities during 2001–2017. Int. J. Appl. Earth Obs. Geoinform. 2019, 75, 44–53. [Google Scholar] [CrossRef]
- Tian, H.; Cao, C.; Chen, W.; Bao, S.; Yang, B.; Myneni, R.B. Response of vegetation activity dynamic to climatic change and ecological restoration programs in Inner Mongolia from 2000 to 2012. Ecol. Eng. 2015, 82, 276–289. [Google Scholar] [CrossRef]
- Yi, L.; Ren, Z.; Zhang, C.; Liu, W. Vegetation Cover, Climate and Human Activities on the Loess Plateau. Resour. Sci. 2014, 36, 166–174. [Google Scholar]
- Zhang, J.; Du, J.; Sheng, Z.; Zhang, Y.; Wu, J.; Liu, B. Spatio-temporal Changes of Vegetation Cover and Their Influencing Factors in the Yellow River Basin from 1982 to 2015. Ecol. Environ. Sci. 2021, 30, 929–937. [Google Scholar]
- Li, S.; Sun, Z.; Tan, M.; Li, X. Effects of rural-urban migration on vegetation greenness in fragile areas: A case study of Inner Mongolia in China. J. Geogr. Sci. 2016, 26, 313–324. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SEDI | Classification |
|---|---|
| Less than −2.0 | Extreme drought |
| −1.99 to −1.5 | Severe drought |
| −1.49 to −1.0 | Moderate drought |
| −0.99 to −0.5 | Mild drought |
| −0.5 to 0.5 | Normal |
| 0.5 to 0.99 | Mildly wet |
| 1.0 to 1.49 | Moderately wet |
| 1.5 to 1.99 | Severely wet |
| Larger than 2.0 | Extremely wet |
| YLRB | YTRB | |||
|---|---|---|---|---|
| Significant Increase (p < 0.05) | Significant Decrease (p < 0.05) | Significant Increase (p < 0.05) | Significant Decrease (p < 0.05) | |
| SEDI | 13.95% | 12.63% | 45.17% | 9.44% |
| NDVI | 69.91% | 1.17% | 54.22% | 4.54% |
| Determination Coefficients | Standard Deviations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Forests | Grasslands | Croplands | Others | Forests | Grasslands | Croplands | Others | |||
| YLRB | TEM | R2_no | 0.443 | 0.590 | 0.446 | 0.375 | 0.071 | 0.140 | 0.134 | 0.233 |
| R2_acc | 0.499 | 0.694 | 0.510 | 0.462 | 0.071 | 0.132 | 0.147 | 0.252 | ||
| R2_lag | 0.447 | 0.599 | 0.448 | 0.401 | 0.066 | 0.136 | 0.132 | 0.224 | ||
| R2_lagacc | 0.505 | 0.694 | 0.524 | 0.472 | 0.065 | 0.130 | 0.123 | 0.241 | ||
| PRE | R2_no | 0.782 | 0.670 | 0.691 | 0.478 | 0.100 | 0.111 | 0.130 | 0.256 | |
| R2_acc | 0.800 | 0.727 | 0.714 | 0.519 | 0.089 | 0.101 | 0.136 | 0.260 | ||
| R2_lag | 0.786 | 0.693 | 0.695 | 0.497 | 0.091 | 0.095 | 0.129 | 0.247 | ||
| R2_lagacc | 0.800 | 0.727 | 0.715 | 0.522 | 0.089 | 0.101 | 0.133 | 0.256 | ||
| YTRB | TEM | R2_no | 0.251 | 0.352 | 0.264 | 0.227 | 0.198 | 0.261 | 0.176 | 0.229 |
| R2_acc | 0.363 | 0.494 | 0.421 | 0.417 | 0.198 | 0.241 | 0.126 | 0.192 | ||
| R2_lag | 0.327 | 0.435 | 0.354 | 0.339 | 0.161 | 0.210 | 0.116 | 0.178 | ||
| R2_lagacc | 0.395 | 0.521 | 0.441 | 0.433 | 0.165 | 0.209 | 0.109 | 0.182 | ||
| PRE | R2_no | 0.472 | 0.562 | 0.604 | 0.582 | 0.276 | 0.235 | 0.156 | 0.172 | |
| R2_acc | 0.531 | 0.620 | 0.630 | 0.601 | 0.245 | 0.206 | 0.135 | 0.167 | ||
| R2_lag | 0.526 | 0.607 | 0.624 | 0.590 | 0.225 | 0.181 | 0.127 | 0.165 | ||
| R2_lagacc | 0.547 | 0.633 | 0.634 | 0.601 | 0.222 | 0.180 | 0.127 | 0.167 | ||
| Months | Standard Deviations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Forests | Grasslands | Croplands | Others | Forests | Grasslands | Croplands | Others | |||
| YLRB | TEM | Acc | 0.647 | 1.080 | 0.645 | 1.103 | 0.512 | 0.395 | 0.518 | 0.788 |
| Lag | 0.103 | 0.516 | 0.112 | 0.552 | 0.306 | 0.502 | 0.316 | 0.502 | ||
| PRE | Acc | 1.000 | 1.013 | 0.931 | 1.103 | 0.173 | 0.211 | 0.310 | 0.484 | |
| Lag | 0.059 | 0.229 | 0.073 | 0.466 | 0.237 | 0.421 | 0.339 | 0.681 | ||
| YZRB | TEM | Acc | 1.282 | 1.112 | 0.474 | 0.524 | 1.068 | 0.924 | 0.748 | 0.681 |
| Lag | 0.758 | 0.585 | 0.209 | 0.181 | 0.996 | 0.873 | 0.533 | 0.455 | ||
| PRE | Acc | 1.774 | 1.746 | 1.782 | 2.010 | 1.072 | 0.966 | 0.943 | 0.838 | |
| Lag | 1.123 | 1.009 | 0.893 | 1.086 | 1.117 | 1.006 | 0.897 | 0.761 | ||
| YLRB | YTRB | |||||||
|---|---|---|---|---|---|---|---|---|
| Forests | Grasslands | Croplands | Others | Forests | Grasslands | Croplands | Others | |
| R2_no | 0.711 | 0.788 | 0.723 | 0.710 | 0.614 | 0.505 | 0.597 | 0.619 |
| R2_acc | 0.763 | 0.806 | 0.786 | 0.732 | 0.657 | 0.544 | 0.648 | 0.640 |
| R2_lag | 0.734 | 0.793 | 0.749 | 0.716 | 0.639 | 0.535 | 0.631 | 0.633 |
| R2_lagacc | 0.764 | 0.807 | 0.786 | 0.735 | 0.662 | 0.552 | 0.654 | 0.643 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, W.; Niu, Z.; Wang, L.; Yao, R.; Gui, X.; Xiang, F.; Ji, Y. Impacts of Drought and Climatic Factors on Vegetation Dynamics in the Yellow River Basin and Yangtze River Basin, China. Remote Sens. 2022, 14, 930. https://doi.org/10.3390/rs14040930
Jiang W, Niu Z, Wang L, Yao R, Gui X, Xiang F, Ji Y. Impacts of Drought and Climatic Factors on Vegetation Dynamics in the Yellow River Basin and Yangtze River Basin, China. Remote Sensing. 2022; 14(4):930. https://doi.org/10.3390/rs14040930
Chicago/Turabian StyleJiang, Weixia, Zigeng Niu, Lunche Wang, Rui Yao, Xuan Gui, Feifei Xiang, and Yuxi Ji. 2022. "Impacts of Drought and Climatic Factors on Vegetation Dynamics in the Yellow River Basin and Yangtze River Basin, China" Remote Sensing 14, no. 4: 930. https://doi.org/10.3390/rs14040930
APA StyleJiang, W., Niu, Z., Wang, L., Yao, R., Gui, X., Xiang, F., & Ji, Y. (2022). Impacts of Drought and Climatic Factors on Vegetation Dynamics in the Yellow River Basin and Yangtze River Basin, China. Remote Sensing, 14(4), 930. https://doi.org/10.3390/rs14040930