
Outbreak of Moroccan Locust in Sardinia (Italy): A Remote Sensing Perspective

 ,
,  ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
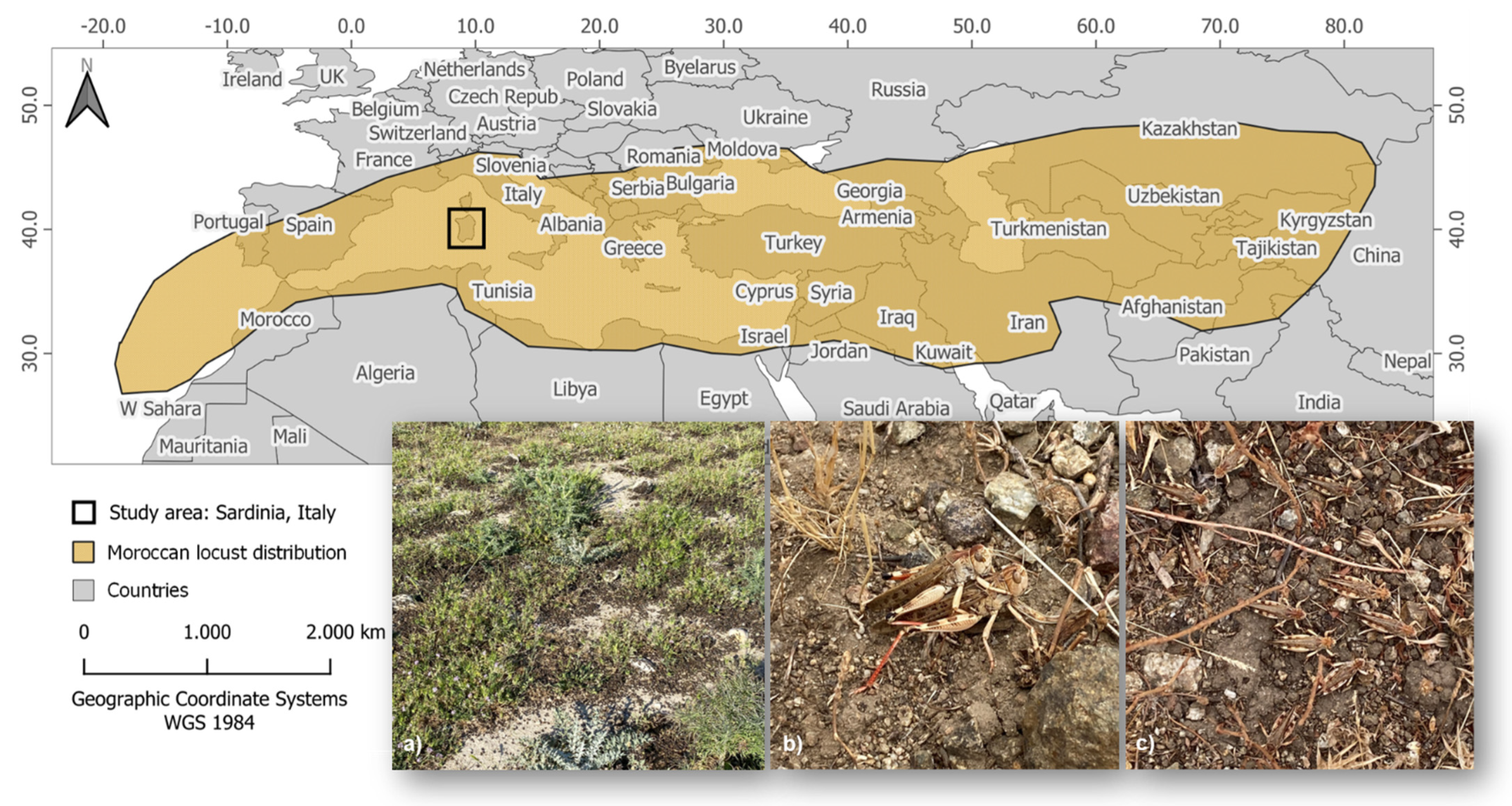
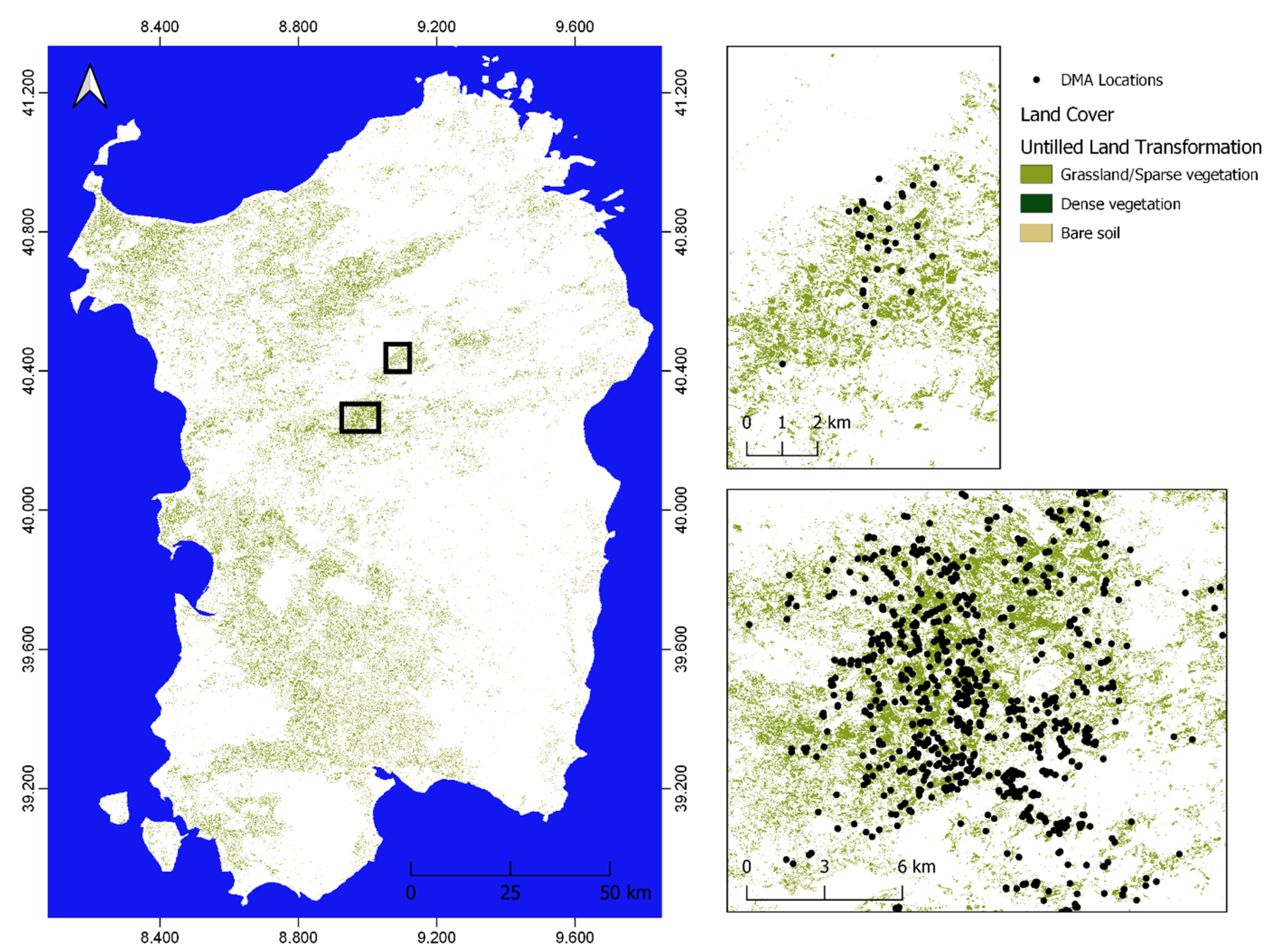
2.1. Study Area
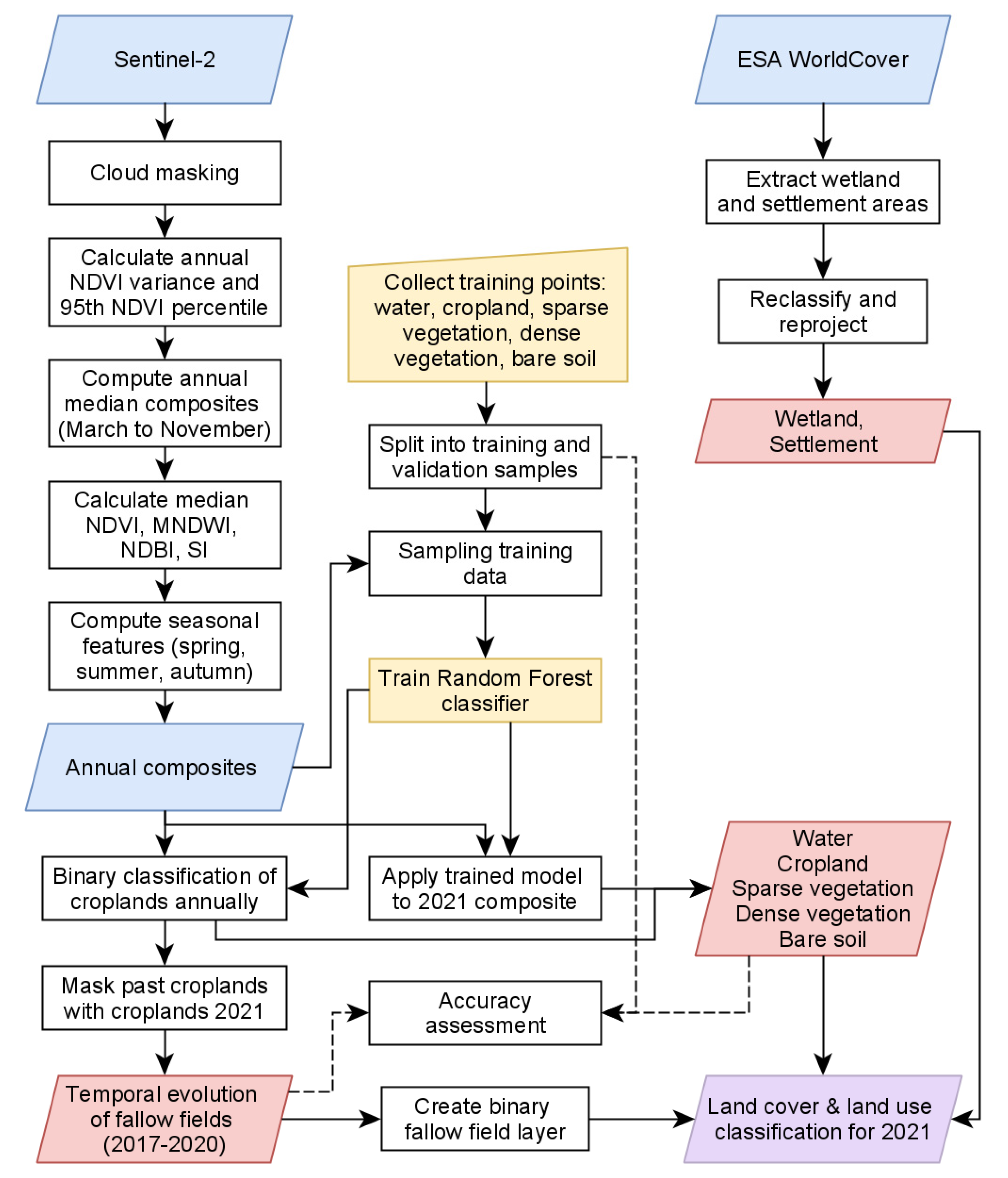
2.2. Classification of Actual State of LCLU with Focus on DMA Relevant Land Characteristics
2.3. Moroccan Locust Record Locations
2.4. Combination of Nymph Locations with Data from Remote Sensing
3. Results
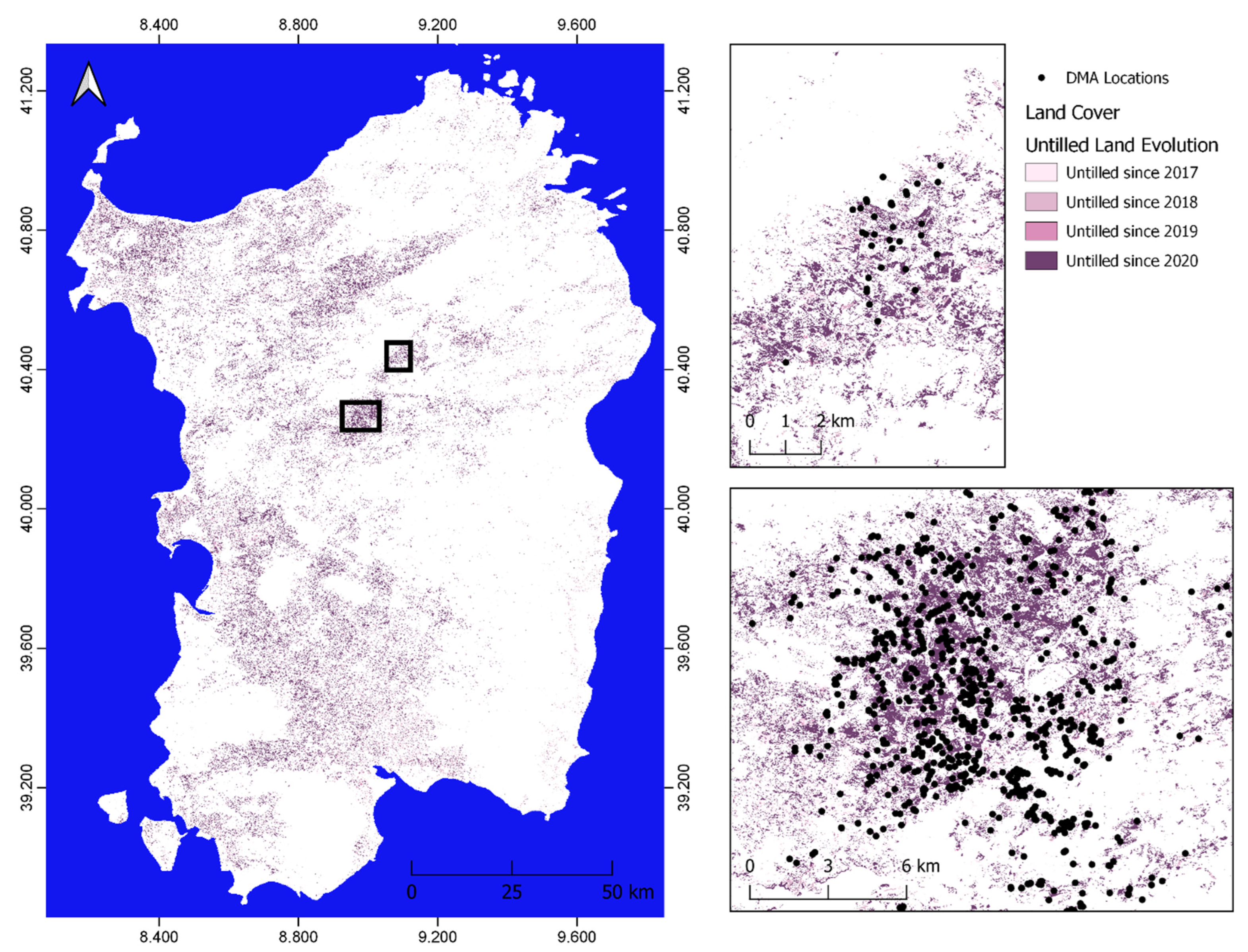
3.1. Relation of DMA Locations with Previous and Actual Land Cover
3.2. Accuracy Assessment
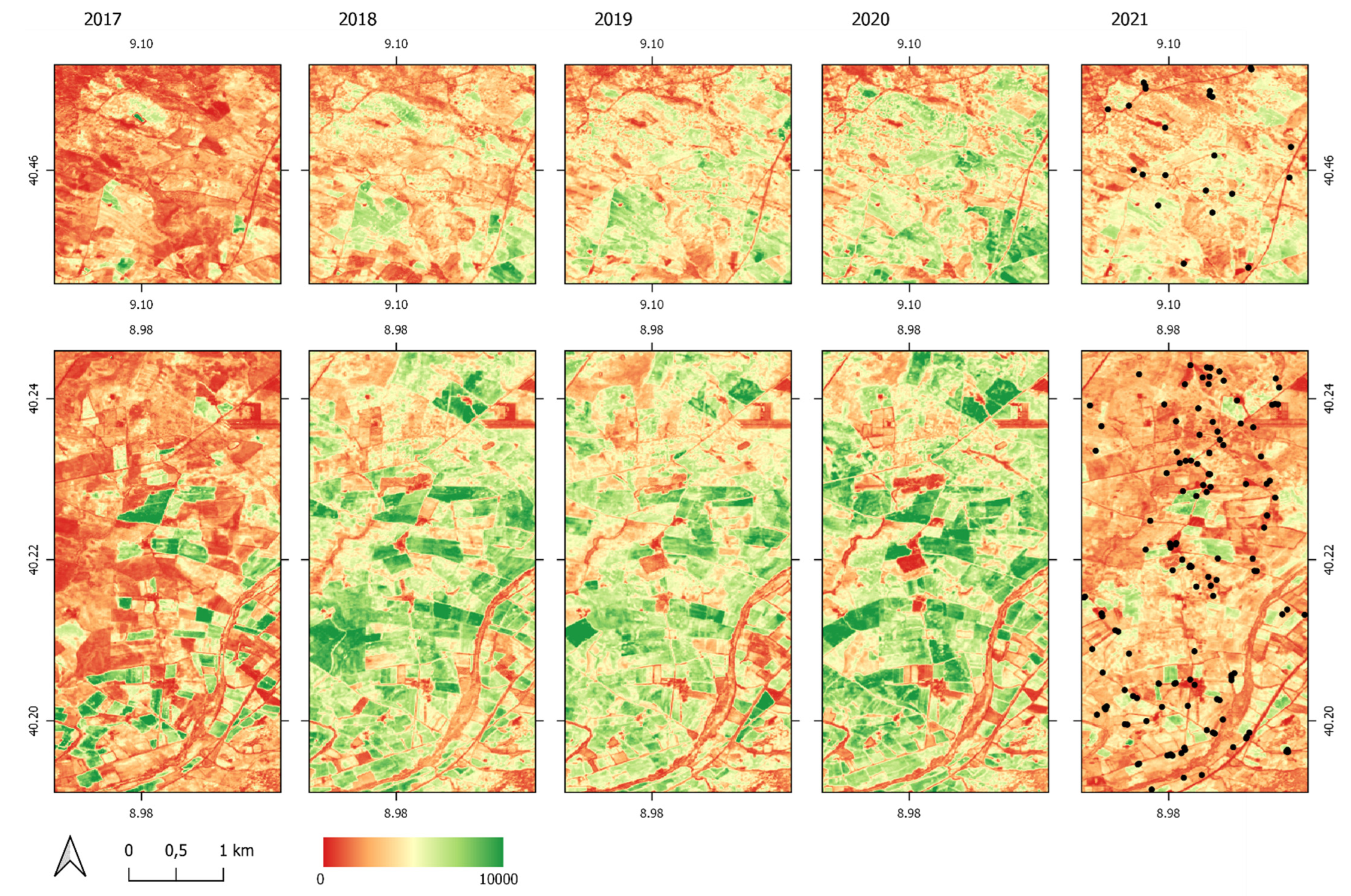
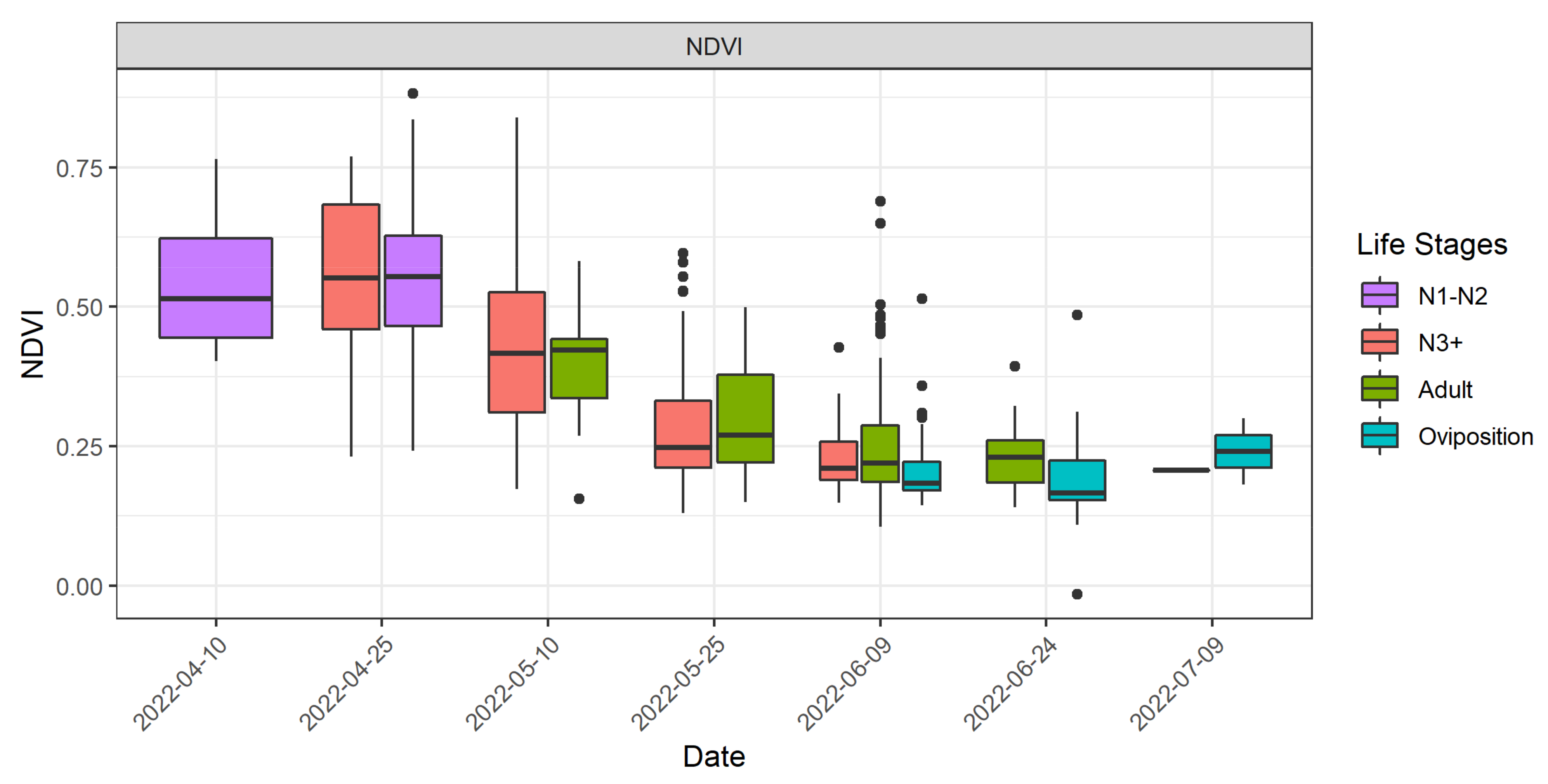
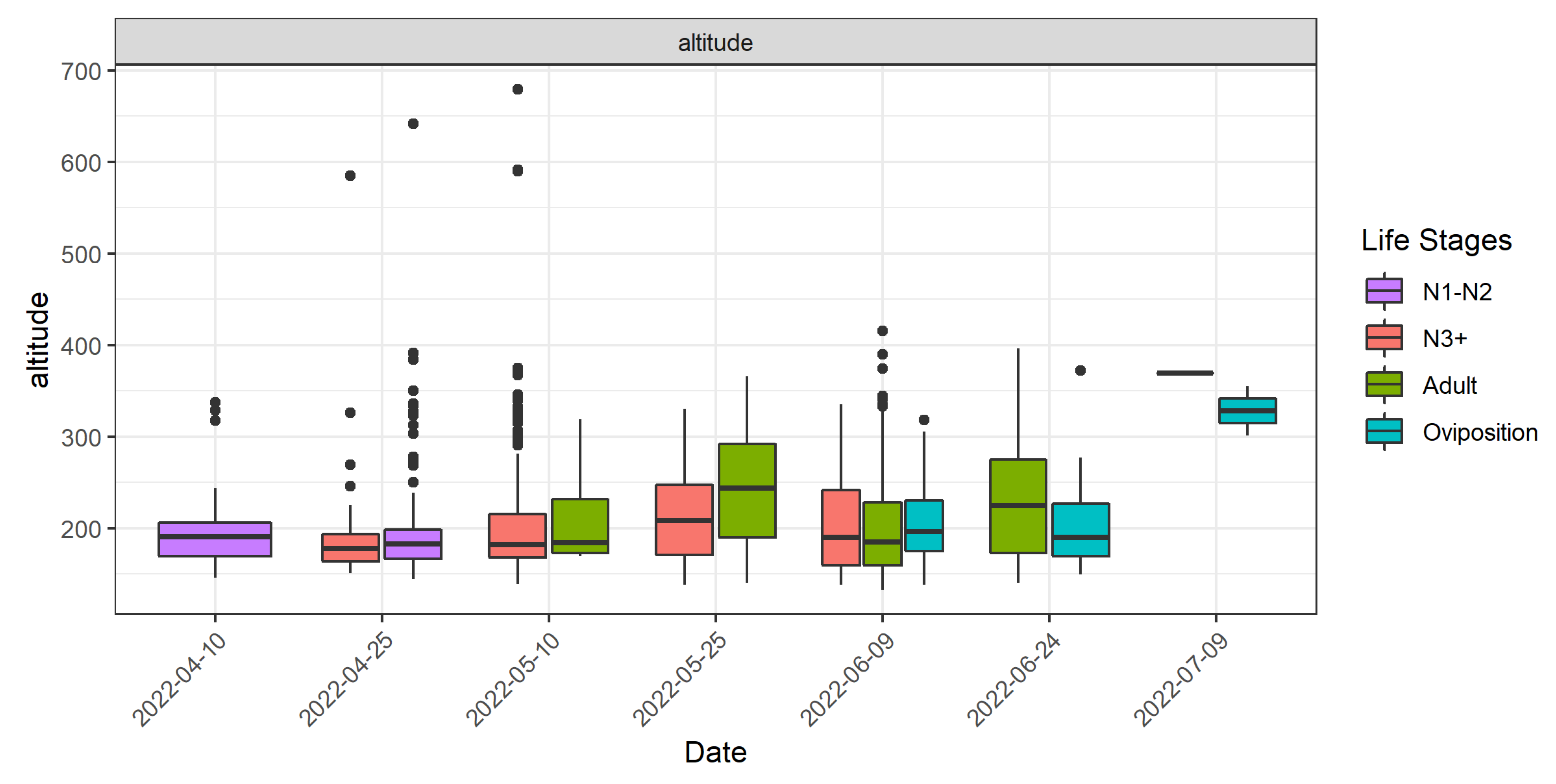
3.3. Relation of DMA Locations with Vegetation Development and Elevation
4. Discussion
5. Conclusions
- 43% were located on land that was previously used for agriculture purposes (fallow or previously tilled land);
- 23% were located on cropland within a radius of 100 m to abandoned, fallow, or untilled land, due to possible displacement after hatching as well as possible inaccuracy of land cover classification;
- The majority of locations detected on abandoned, fallow, or untilled land were occupied by active agriculture until 2020, which indicates that DMA occupied this territory immediately;
- Considering the transformation of abandoned, fallow, or untilled land, the majority of locations are found on the sparse vegetation/grassland land cover class (97%).
- Young nymphs were detected in April within the peak of the vegetative period;
- Older nymphs and adults were found in areas with significantly decreased vegetation greenness;
- In terms of altitude, the majority (79%) of DMA locations were found between 137 and 250 m a.s.l.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reuters. Sardinian Farmers Suffer Worst Locust Invasion in over 30 Years 2022. Available online: https://www.reuters.com/world/europe/sardinian-farmers-suffer-worst-locust-invasion-over-30-years-2022-07-13/ (accessed on 25 July 2022).
- Reuters. Sardinia Hit by Worst Locust Invasion for 70 Years 2019. Available online: https://www.reuters.com/article/us-italy-locusts-idUSKCN1TC1BY (accessed on 14 August 2020).
- Latchininsky, A.V. Moroccan Locust Dociostaurus maroccanus (Thunberg, 1815): A Faunistic Rarity or an Important Economic Pest? J. Insect Conserv. 1998, 2, 167–178. [Google Scholar] [CrossRef]
- Pantaleoni, R.A.; Molinu, A.; Cesaroni, C. Some Aspects of Locust Control in Sardinia in the First Half of the Twentieth Century. In Arsenic Locusts—The Control of Locusts in Sardinia in the First Half of Twentieth Century; Molinu, A., Cesaroni, C., Pantaleoni, R.A., Eds.; Composita: Sassari, Italy, 2004; pp. 17–50. [Google Scholar]
- Malakhov, D.V.; Zlatanov, B.V. An Ecological Niche Model for Dociostaurus maroccanus, Thunberg, 1815 (Orthoptera, Acrididae): The Nesting Environment and Survival of Egg-Pods. Biosis Biol. Syst. 2020, 1, 8–24. [Google Scholar] [CrossRef][Green Version]
- FAO Locust Watch—Locusts in Caucasus and Central Asia. Food and Agriculture Organization of the United Nations (FAO). 2021. Available online: https://www.fao.org/locusts-cca/bioecology/moroccan-locust-dma/en/ (accessed on 29 September 2022).
- Aragón, P.; Coca-Abia, M.M.; Llorente, V.; Lobo, J.M. Estimation of Climatic Favourable Areas for Locust Outbreaks in Spain: Integrating Species’ Presence Records and Spatial Information on Outbreaks. J. Appl. Entomol. 2013, 137, 610–623. [Google Scholar] [CrossRef]
- Kambulin, V.E. Locust—Methods of Assessing Harm, Forecasting the Number and Technologies for Identifying Populated Areas. Kazakh Research Institute of Plant Protection and Quarantine: Almaty, Kazakhstan, 2018; ISBN 978-601-7416-92-8. [Google Scholar]
- Showler, A.T.; Lecoq, M. Incidence and Ramifications of Armed Conflict in Countries with Major Desert Locust Breeding Areas. Agronomy 2021, 11, 114. [Google Scholar] [CrossRef]
- Ortu, S.; Prota, R. Possibilità di Lotta Biologica Contro le Cavallette: Il Caso del Dociostaurus maroccanus Thunb. (Osservazioni Preliminari). Proc. S.I.T.E. 1989, 8, 89–97. [Google Scholar]
- Latchininsky, A.V. Locusts and Remote Sensing: A Review. J. Appl. Remote Sens 2013, 7, 075099. [Google Scholar] [CrossRef]
- Cressman, K. Role of Remote Sensing in Desert Locust Early Warning. J. Appl. Remote Sens 2013, 7, 075098. [Google Scholar] [CrossRef]
- Zhang, L.; Lecoq, M.; Latchininsky, A.; Hunter, D. Locust and Grasshopper Management. Annu. Rev. Entomol. 2019, 64, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Klein, I.; Oppelt, N.; Kuenzer, C. Application of Remote Sensing Data for Locust Research and Management—A Review. Insects 2021, 12, 233. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.M. Advances in the Control of Locusts (Orthoptera: Acrididae) in Eastern Australia: From Crop Protection to Preventive Control. Aust J. Entomol. 2004, 43, 293–303. [Google Scholar] [CrossRef]
- Magor, J.I.; Lecoq, M.; Hunter, D.M. Preventive Control and Desert Locust Plagues. Crop Prot. 2008, 27, 1527–1533. [Google Scholar] [CrossRef]
- Pedgley, D.E. ERTS Surveys a 500 Km2 Locust Breeding Site in Saudi Arabia. In Third Earth Resources Technology Satellite—Symposium; Frieden, S.C., Mercanti, E.P., Becker, M.A., Eds.; NASA: Washington, DC, USA; Volume 1, pp. 233–246.
- Hielkema, J.U. Application of Landsat Data in Desert Locust Survey and Control; Report of the Desert Locust satellite Applications Projects, Stage II; FAO: Rome, Italy, 1977. [Google Scholar]
- Hielkema, J.U.; Snijders, F.L. Operational Use of Environmental Satellite Remote Sensing and Satellite Communications Technology for Global Food Security and Locust Control by FAO: The ARTEMIS and DIANA Systems. Acta Astronaut. 1994, 32, 603–616. [Google Scholar] [CrossRef]
- Ceccato, P.; Bell, M.; Blumenthal, M.; Connor, S.; Dinku, T.; Grover-Kopec, E.; Ropelewski, C.; Thomson, M. Use of Remote Sensing for Monitoring Climate Variability for Integrated Early Warning Systems: Applications for Human Diseases and Desert Locust Management. In Proceedings of the 2006 IEEE International Symposium on Geoscience and Remote Sensing, Denver, CO, USA, 31 July–4 August 2006; IEEE: Denver, CO, USA, 2006; pp. 270–274. [Google Scholar]
- Pekel, J.-F.; Ceccato, P.; Vancutsem, C.; Cressman, K.; Vanbogaert, E.; Defourny, P. Development and Application of Multi-Temporal Colorimetric Transformation to Monitor Vegetation in the Desert Locust Habitat. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2011, 4, 318–326. [Google Scholar] [CrossRef]
- Bryceson, K.P.; Hunter, D.M.; Hamilton, G.L. Use of Remotely Sensed Data in the Australian Plague Locust Commission. In Proceedings of the Pest Control & Sustainable Agriculture; 5th Australian Applied Entomological Research Conf: Canberra, Australia, 1993; pp. 435–439. [Google Scholar]
- Piou, C.; Lebourgeois, V.; Benahi, A.S.; Bonnal, V.; el Hacen Jaavar, M.; Lecoq, M.; Vassal, J.-M. Coupling Historical Prospection Data and a Remotely-Sensed Vegetation Index for the Preventative Control of Desert Locusts. Basic Appl. Ecol. 2013, 14, 593–604. [Google Scholar] [CrossRef]
- Piou, C.; Gay, P.; Benahi, A.S.; Babah Ebbe, M.A.O.; Chihrane, J.; Ghaout, S.; Cisse, S.; Diakite, F.; Lazar, M.; Cressman, K.; et al. Soil Moisture from Remote Sensing to Forecast Desert Locust Presence. J. Appl. Ecol. 2019, 56, 966–975. [Google Scholar] [CrossRef]
- Crooks, W.T.S.; Cheke, R.A. Soil Moisture Assessments for Brown Locust Locustana pardalina Breeding Potential Using Synthetic Aperture Radar. J. Appl. Remote Sens 2014, 8, 084898. [Google Scholar] [CrossRef]
- Escorihuela, M.J.; Merlin, O.; Stefan, V.; Moyano, G.; Eweys, O.A.; Zribi, M.; Kamara, S.; Benahi, A.S.; Ebbe, M.A.B.; Chihrane, J.; et al. SMOS Based High Resolution Soil Moisture Estimates for Desert Locust Preventive Management. Remote Sens. Appl. Soc. Environ. 2018, 11, 140–150. [Google Scholar] [CrossRef]
- Gómez, D.; Salvador, P.; Sanz, J.; Casanova, C.; Taratiel, D.; Casanova, J.L. Machine Learning Approach to Locate Desert Locust Breeding Areas Based on ESA CCI Soil Moisture. J. Appl. Remote Sens. 2018, 12, 036011. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Rivas-Sáenz, S.; Penas, A.S. Worldwide Bioclimatic Classification System. Glob. Geobot. 2011, 1, 634. [Google Scholar]
- Canu, S.; Rosati, L.; Fiori, M.; Motroni, A.; Filigheddu, R.; Farris, E. Bioclimate Map of Sardinia (Italy). J. Maps 2015, 11, 711–718. [Google Scholar] [CrossRef]
- Secci, D.; Patriche, C.V.; Ursu, A.; Sfica, L. Spatial Interpolation of Mean Annual Precipitations in Sardinia. A Comparative Analysis of Several Methods. Geogr. Tech. 2010, 9, 67–75. [Google Scholar]
- Bartholomé, E.; Belward, A.S. GLC2000: A New Approach to Global Land Cover Mapping from Earth Observation Data. Int. J. Remote Sens. 2005, 26, 1959–1977. [Google Scholar] [CrossRef]
- Friedl, M.A.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. MODIS Collection 5 Global Land Cover: Algorithm Refinements and Characterization of New Datasets. Remote Sens. Environ. 2010, 114, 168–182. [Google Scholar] [CrossRef]
- Winkler, K.; Fuchs, R.; Rounsevell, M.; Herold, M. Global Land Use Changes Are Four Times Greater than Previously Estimated. Nat Commun 2021, 12, 2501. [Google Scholar] [CrossRef] [PubMed]
- Pekel, J.-F.; Cottam, A.; Gorelick, N.; Belward, A.S. High-Resolution Mapping of Global Surface Water and Its Long-Term Changes. Nature 2016, 540, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Zanaga, D.; Van De Kerchove, R.; De Keersmaecker, W.; Souverijns, N.; Brockmann, C.; Quast, R.; Wevers, J.; Grosu, A.; Paccini, A.; Vergnaud, S.; et al. ESA WorldCover 10 m 2020 V100 2021. Available online: https://pure.iiasa.ac.at/id/eprint/18398/ (accessed on 25 July 2022).
- Gessner, U.; Machwitz, M.; Esch, T.; Tillack, A.; Naeimi, V.; Kuenzer, C.; Dech, S. Multi-Sensor Mapping of West African Land Cover Using MODIS, ASAR and TanDEM-X/TerraSAR-X Data. Remote Sens. Environ. 2015, 164, 282–297. [Google Scholar] [CrossRef]
- Klein, I.; Gessner, U.; Kuenzer, C. Regional Land Cover Mapping and Change Detection in Central Asia Using MODIS Time-Series. Appl. Geogr. 2012, 35, 219–234. [Google Scholar] [CrossRef]
- Leinenkugel, P.; Kuenzer, C.; Oppelt, N.; Dech, S. Characterisation of Land Surface Phenology and Land Cover Based on Moderate Resolution Satellite Data in Cloud Prone Areas—A Novel Product for the Mekong Basin. Remote Sens. Environ. 2013, 136, 180–198. [Google Scholar] [CrossRef]
- Pickens, A.H.; Hansen, M.C.; Hancher, M.; Stehman, S.V.; Tyukavina, A.; Potapov, P.; Marroquin, B.; Sherani, Z. Mapping and Sampling to Characterize Global Inland Water Dynamics from 1999 to 2018 with Full Landsat Time-Series. Remote Sens. Environ. 2020, 243, 111792. [Google Scholar] [CrossRef]
- Le Gall, M.; Overson, R.; Cease, A. A Global Review on Locusts (Orthoptera: Acrididae) and Their Interactions with Livestock Grazing Practices. Front. Ecol. Evol. 2019, 7, 263. [Google Scholar] [CrossRef]
- Sivanpillai, R.; Latchininsky, A.V.; Peveling, R.; Pankov, V.I.; Diagnosis, P. Utility of the IRS-AWiFS Data to Map the Potential Italian Locust (Calliptamus italicus) Habitats in Northern Kazakhstan; ASPRS: Baltimore, MD, USA, 2009. [Google Scholar]
- Bryceson, K.P. The Use of Landsat MSS Data to Determine the Locust Eggbeds of Locust Eggbeds in the Riverina Region of New South Wales, Australia. Int. J. Remote Sens. 1989, 10, 1749–1762. [Google Scholar] [CrossRef]
- de Miranda, E.E.; Duranton, J.-F.; Lecoq, M. Static and Dynamic Cartographies of the Biotopes of the Grasshopper Rhammatocerus Schistocercoides (Rehn, 1906) in the State of Mato Grosso, Brazil; Anais do International Symposium on Resource and Environmental Monitoring – ECO RIO´94: Rio de Janeiro, Brazil, 1994; pp. 67–72. [Google Scholar]
- Latchininsky, A.V.; Sivanpillai, R.; Driese, K.L.; Wilps, H. Can Early Season Landsat Images Improve Locust Habitat Monitoring in the Amudarya River Delta of Uzbekistan. J. Orthoptera Res. 2007, 16, 167–173. [Google Scholar] [CrossRef]
- Lazar, M.; Aliou, D.; Jeng-Tze, Y.; Doumandji-Mitiche, B.; Lecoq, M. Location and Characterization of Breeding Sites of Solitary Desert Locust Using Satellite Images Landsat 7 ETM+ and Terra MODIS. Adv. Entomol. 2015, 3, 6–15. [Google Scholar] [CrossRef][Green Version]
- Löw, F.; Waldner, F.; Latchininsky, A.; Biradar, C.; Bolkart, M.; Colditz, R.R. Timely Monitoring of Asian Migratory Locust Habitats in the Amudarya Delta, Uzbekistan Using Time Series of Satellite Remote Sensing Vegetation Index. J. Environ. Manag. 2016, 183, 562–575. [Google Scholar] [CrossRef]
- McCulloch, L.; Hunter, D.M. Identification and Monitoring of Australian Plague Locust Habitats from Landsat. Remote Sens. Environ. 1983, 13, 95–102. [Google Scholar] [CrossRef]
- Shi, Y.; Huang, W.; Dong, Y.; Peng, D.; Zheng, Q.; Yang, P. The Influence of Landscape’s Dynamics on the Oriental Migratory Locust Habitat Change Based on the Time-Series Satellite Data. J. Environ. Manag. 2018, 218, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Huang, W.; Chen, J.; Dong, Y.; Ren, B.; Geng, Y. Land Use/Cover Changes in the Oriental Migratory Locust Area of China: Implications for Ecological Control and Monitoring of Locust Area. Agric. Ecosyst. Environ. 2020, 303, 107110. [Google Scholar] [CrossRef]
- Latchininsky, A.; Sword, G.; Sergeev, M.; Cigliano, M.M.; Lecoq, M. Locusts and Grasshoppers: Behavior, Ecology, and Biogeography. Psyche A J. Entomol. 2011, 2011, 578327. [Google Scholar] [CrossRef]
- Monard, A.; Chiris, M.; Latchininsky, A.V. Analytical Report on Locust Situations and Management in Caucasus and Central Asia (Cca); FAO: Rome, Italy, 2009. [Google Scholar]
- Sergeev, M.G. Ups and Downs of the Italian Locust (Calliptamus italicus L.) Populations in the Siberian Steppes: On the Horns of Dilemmas. Agronomy 2021, 11, 746. [Google Scholar] [CrossRef]
- Sergeev, M.G.; Childebaev, M.K.; Vankova, I.A.; Gapparov, F.A.; Kambulin, V.E.; Kokanova, E.O.; Latchininsky, A.V.; Pshenitsyna, L.B.; Temreshev, I.I.; Chernyakhovsky, M.E.; et al. Italian Locust Calliptamus italicus (Linnaeus, 1758). Morphology, Distribution, Ecology, Population Management; FAO: Rome, Italy, 2022; ISBN 978-92-5-135438-4. [Google Scholar]
- Estel, S.; Kuemmerle, T.; Alcántara, C.; Levers, C.; Prishchepov, A.; Hostert, P. Mapping Farmland Abandonment and Recultivation across Europe Using MODIS NDVI Time Series. Remote Sens. Environ. 2015, 163, 312–325. [Google Scholar] [CrossRef]
- Orynbaikyzy, A.; Gessner, U.; Mack, B.; Conrad, C. Crop Type Classification Using Fusion of Sentinel-1 and Sentinel-2 Data: Assessing the Impact of Feature Selection, Optical Data Availability, and Parcel Sizes on the Accuracies. Remote Sens. 2020, 12, 2779. [Google Scholar] [CrossRef]
- Prishchepov, A.V.; Müller, D.; Dubinin, M.; Baumann, M.; Radeloff, V.C. Determinants of Agricultural Land Abandonment in Post-Soviet European Russia. Land Use Policy 2013, 30, 873–884. [Google Scholar] [CrossRef]
- Verbesselt, J.; Hyndman, R.; Newnham, G.; Culvenor, D. Detecting Trend and Seasonal Changes in Satellite Image Time Series. Remote Sens. Environ. 2010, 114, 106–115. [Google Scholar] [CrossRef]
- Zeng, L.; Wardlow, B.D.; Xiang, D.; Hu, S.; Li, D. A Review of Vegetation Phenological Metrics Extraction Using Time-Series, Multispectral Satellite Data. Remote Sens. Environ. 2020, 237, 111511. [Google Scholar] [CrossRef]
- Benjamin, K.; Domon, G.; Bouchard, A. Vegetation Composition and Succession of Abandoned Farmland: Effects of Ecological, Historical and Spatial Factors. Landsc. Ecol 2005, 20, 627–647. [Google Scholar] [CrossRef]
- ARPAS. Analisi Agrometeorologica e Climatologica Della Sardegna Analisi Delle Condizioni Meteorologiche e Conseguenze Sul Territorio Regionale Nel Periodo Ottobre 2020–Settembre 2021; ARPAS (Agenzia Regionale per la Protezione dell’Ambiente della Sardegna): Cagliari, Italy, 2021. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Dirscherl, M.; Dietz, A.J.; Kneisel, C.; Kuenzer, C. Automated Mapping of Antarctic Supraglacial Lakes Using a Machine Learning Approach. Remote Sens. 2020, 12, 1203. [Google Scholar] [CrossRef]
- Dubertret, F.; Le Tourneau, F.-M.; Villarreal, M.L.; Norman, L.M. Monitoring Annual Land Use/Land Cover Change in the Tucson Metropolitan Area with Google Earth Engine (1986–2020). Remote Sens. 2022, 14, 2127. [Google Scholar] [CrossRef]
- Phan, T.N.; Kuch, V.; Lehnert, L.W. Land Cover Classification Using Google Earth Engine and Random Forest Classifier—The Role of Image Composition. Remote Sens. 2020, 12, 2411. [Google Scholar] [CrossRef]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Copernicus DEM GLO-30 © DLR, e.V. (2014–2018) and © Airbus Defence and Space GmbH 2022 Provided under COPERNICUS by the European Union and ESA; All Rights Reserved 2022. Available online: https://spacedata.copernicus.eu/web/cscda (accessed on 25 July 2022).
- Molinu, A.; Cesaroni, C.; Pantaleoni, R.A. Arsenic Locusts—The Control of Locusts in Sardinia in the First Half of Twentieth Century; Composita: Sassari, Italy, 2004. [Google Scholar]
- Symmons, P.M.; Cressman, K. Desert Locust Guidelines—1. Biology and Behaviour, 2nd ed.; FAO: Rome, Italy, 2001. [Google Scholar]
- Cisse, S.; Ghaout, S.; Mazih, A.; Babah Ebbe, M.A.O.; Benahi, A.S.; Piou, C. Effect of Vegetation on Density Thresholds of Adult Desert Locust Gregarization from Survey Data in Mauritania. Entomol. Exp. Et Appl. 2013, 149, 159–165. [Google Scholar] [CrossRef]
- Despland, E. Fractal Index Captures the Role of Vegetation Clumping in Locust Swarming. Funct Ecol. 2003, 17, 315–322. [Google Scholar] [CrossRef]
- Deveson, E.D. Satellite Normalized Difference Vegetation Index Data Used in Managing Australian Plague Locusts. J. Appl. Remote Sens 2013, 7, 075096. [Google Scholar] [CrossRef]
- Renier, C.; Waldner, F.; Jacques, D.; Babah Ebbe, M.; Cressman, K.; Defourny, P. A Dynamic Vegetation Senescence Indicator for Near-Real-Time Desert Locust Habitat Monitoring with MODIS. Remote Sens. 2015, 7, 7545–7570. [Google Scholar] [CrossRef]
- Waldner, F.; Ebbe, M.; Cressman, K.; Defourny, P. Operational Monitoring of the Desert Locust Habitat with Earth Observation: An Assessment. ISPRS Int. J. Geo-Inf. 2015, 4, 2379–2400. [Google Scholar] [CrossRef]
- Sivanpillai, R.; Latchininsky, A.V. Mapping Locust Habitats in the Amudarya River Delta, Uzbekistan with Multi-Temporal MODIS Imagery. Environ. Manag. 2007, 39, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Zhao, L.; Dong, Y.; Huang, W.; Shi, Y.; Ren, Y.; Ren, B. Migratory Locust Habitat Analysis With PB-AHP Model Using Time-Series Satellite Images. IEEE Access 2020, 8, 166813–166823. [Google Scholar] [CrossRef]
- Lecoq, M.; Cease, A. What Have We Learned after Millennia of Locust Invasions? Agronomy 2022, 12, 472. [Google Scholar] [CrossRef]
- Kraemer, R.; Prishchepov, A.V.; Müller, D.; Kuemmerle, T.; Radeloff, V.C.; Dara, A.; Terekhov, A.; Frühauf, M. Long-Term Agricultural Land-Cover Change and Potential for Cropland Expansion in the Former Virgin Lands Area of Kazakhstan. Environ. Res. Lett. 2015, 10, 054012. [Google Scholar] [CrossRef]
- Prishchepov, A.V.; Radeloff, V.C.; Baumann, M.; Kuemmerle, T.; Müller, D. Effects of Institutional Changes on Land Use: Agricultural Land Abandonment during the Transition from State-Command to Market-Driven Economies in Post-Soviet Eastern Europe. Environ. Res. Lett. 2012, 7, 024021. [Google Scholar] [CrossRef]
- Legendre, P. Spatial Autocorrelation: Trouble or New Paradigm? Ecology 1993, 74, 1659–1673. [Google Scholar] [CrossRef]
- Adams, E.C.; Parache, H.B.; Cherrington, E.; Ellenburg, W.L.; Mishra, V.; Lucey, R.; Nakalembe, C. Limitations of Remote Sensing in Assessing Vegetation Damage Due to the 2019–2021 Desert Locust Upsurge. Front. Clim. 2021, 3, 714273. [Google Scholar] [CrossRef]
- Weiss, J.E.R. Do Locusts Seek Greener Pastures? An Evaluation of MODIS Vegetation Indices to Predict Presence, Abundance and Impact of the Australian Plague Locust in South–Eastern Australia. Ph.D. Thesis, University of Melbourne, Melbourne, Australia, 2016. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LCLU 2021 | Untilled Since | Untilled LC 2021 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DMA Stage | C | S | B | O | U | 2017 | 2018 | 2019 | 2020 | S | B |
| N1-N2 (113) | 23 | 25 | 0 | 1 | 64 (+) | 1 | 2 | 0 | 61 | 62 | 2 |
| N3+ (435) | 103 | 119 | 21 | 9 | 183 | 5 | 20 | 1 | 157 | 178 | 5 |
| Feeding/moving adults (181) | 36 | 77 (+) | 3 | 7 | 58 (2212) | 1 | 7 | 3 | 47 | 56 | 2 |
| Oviposition (85) | 25 | 15 | 2 | 1 | 42 | 0 | 0 | 0 | 42 | 42 | 0 |
| Total (814) | 187 | 236 | 26 | 18 | 347 | 7 | 29 | 4 | 307 | 338 | 9 |
| Classes | Cropland | Non-Cropland | Both | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Accuracy Measure | EO (%) | UA (%) | EC (%) | PA (%) | EO (%) | UA (%) | EC (%) | PA (%) | OA (%) | K |
| 2017 | 14.29 | 85.71 | 7.69 | 92.31 | 1.47 | 98.53 | 2.90 | 97.10 | 96.34 | 0.867 |
| 2018 | 34.55 | 65.45 | 0.00 | 100.00 | 0.00 | 100.00 | 8.92 | 91.08 | 92.37 | 0.747 |
| 2021 | 15.09 | 84.91 | 0.00 | 100.00 | 0.00 | 100.00 | 3.98 | 96.02 | 96.75 | 0.898 |
| Average | 21.31 | 78.69 | 2.56 | 97.44 | 0.49 | 99.51 | 5.23 | 94.73 | 95.15 | 0.837 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klein, I.; Cocco, A.; Uereyen, S.; Mannu, R.; Floris, I.; Oppelt, N.; Kuenzer, C. Outbreak of Moroccan Locust in Sardinia (Italy): A Remote Sensing Perspective. Remote Sens. 2022, 14, 6050. https://doi.org/10.3390/rs14236050
Klein I, Cocco A, Uereyen S, Mannu R, Floris I, Oppelt N, Kuenzer C. Outbreak of Moroccan Locust in Sardinia (Italy): A Remote Sensing Perspective. Remote Sensing. 2022; 14(23):6050. https://doi.org/10.3390/rs14236050
Chicago/Turabian StyleKlein, Igor, Arturo Cocco, Soner Uereyen, Roberto Mannu, Ignazio Floris, Natascha Oppelt, and Claudia Kuenzer. 2022. "Outbreak of Moroccan Locust in Sardinia (Italy): A Remote Sensing Perspective" Remote Sensing 14, no. 23: 6050. https://doi.org/10.3390/rs14236050
APA StyleKlein, I., Cocco, A., Uereyen, S., Mannu, R., Floris, I., Oppelt, N., & Kuenzer, C. (2022). Outbreak of Moroccan Locust in Sardinia (Italy): A Remote Sensing Perspective. Remote Sensing, 14(23), 6050. https://doi.org/10.3390/rs14236050