

Study of the Spatiotemporal Variability of Oceanographic Parameters and Their Relationship to Holothuria Species Abundance in a Marine Protected Area of the Mediterranean Using Satellite Imagery



Abstract
1. Introduction
2. Materials and Methods
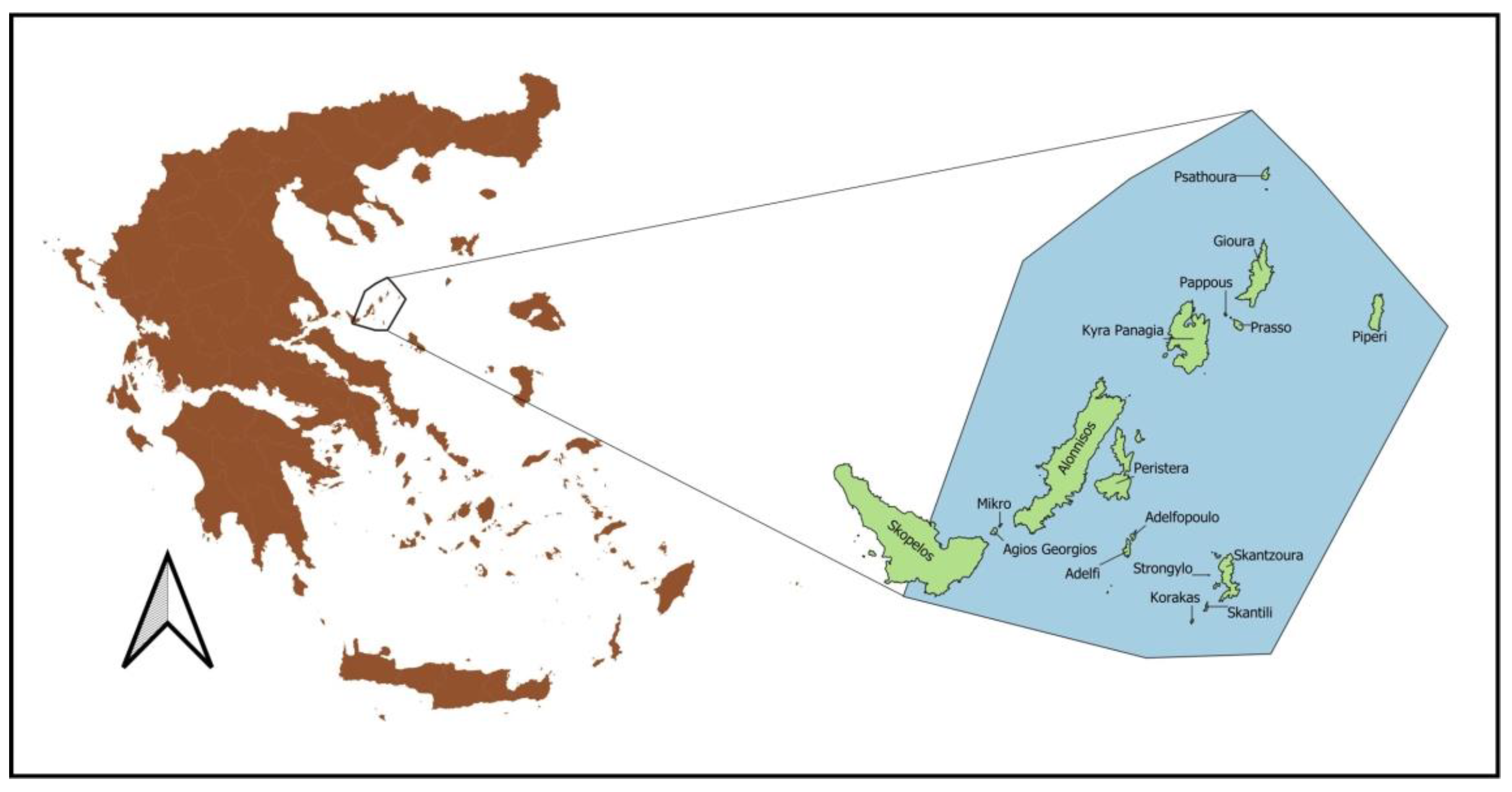
2.1. Study Area
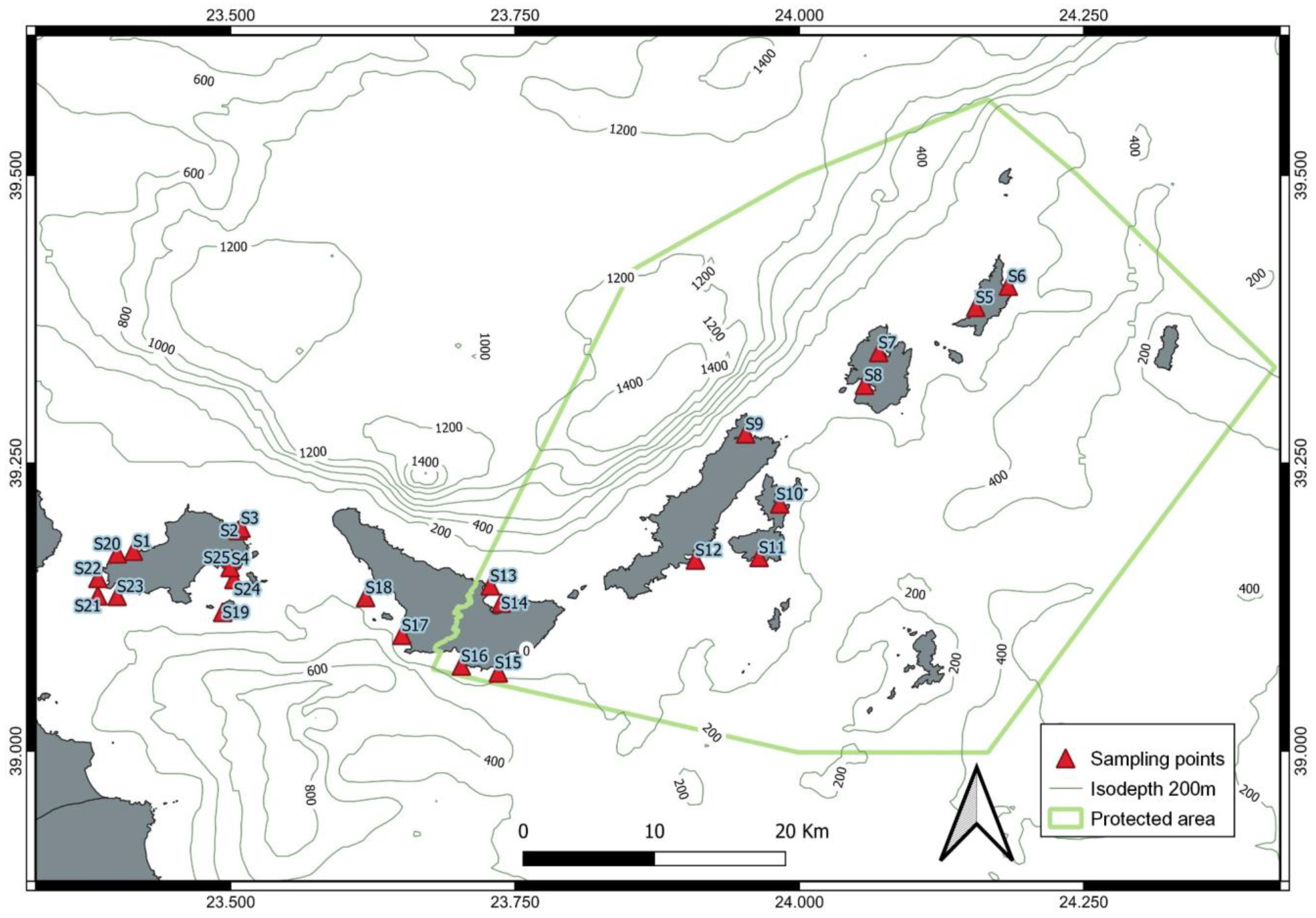
2.2. In Situ Data
2.3. Satellite Database
3. Results
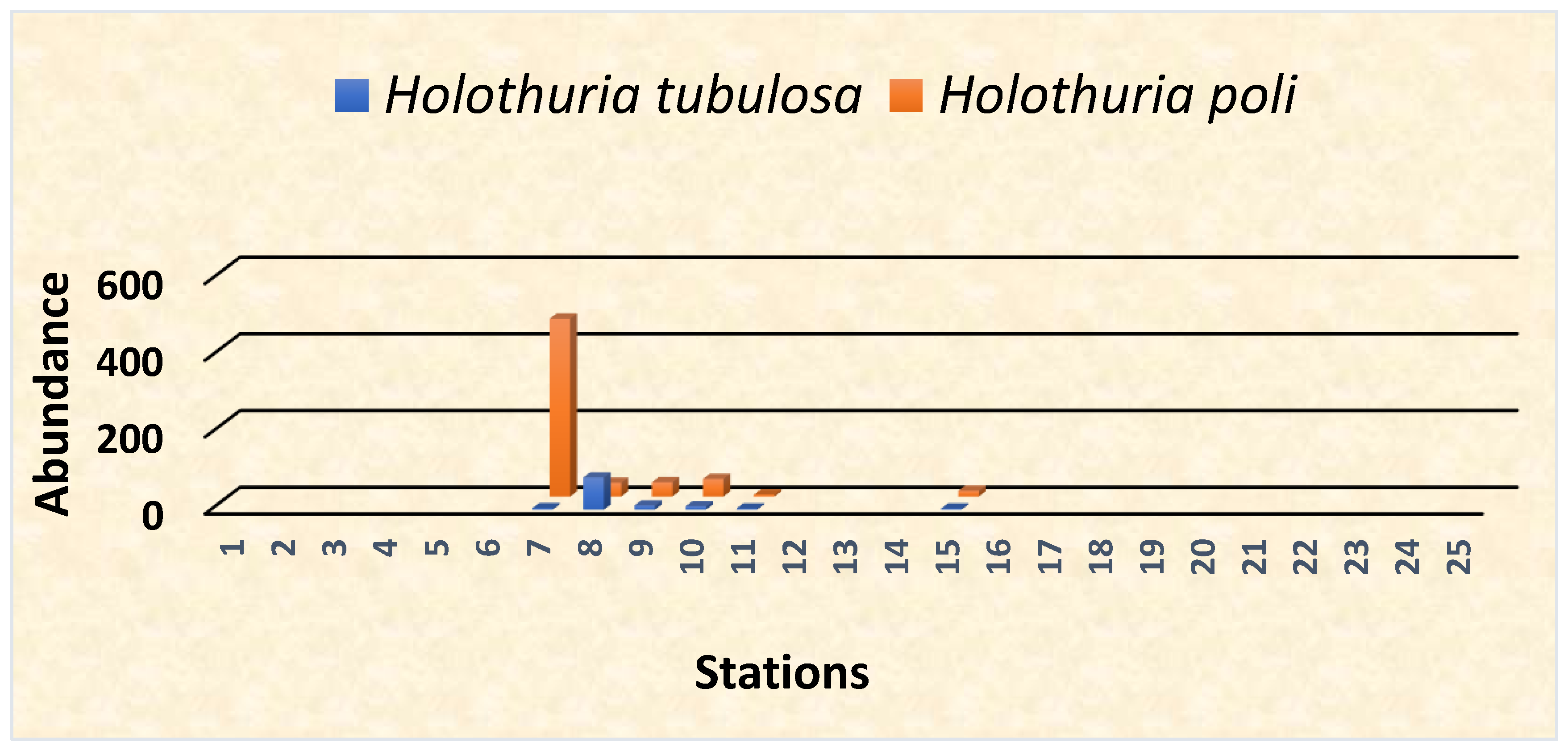
3.1. Species Abundance
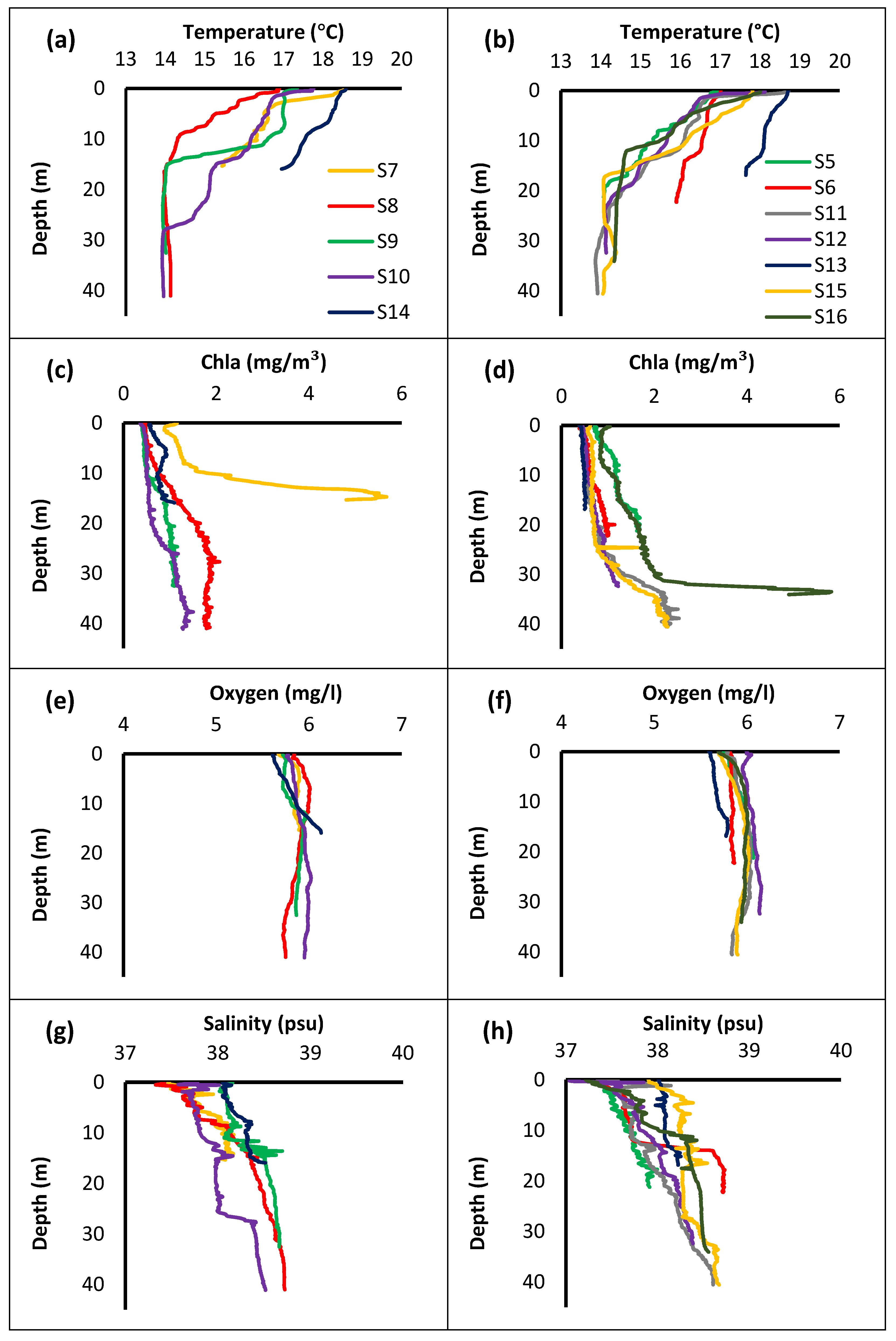
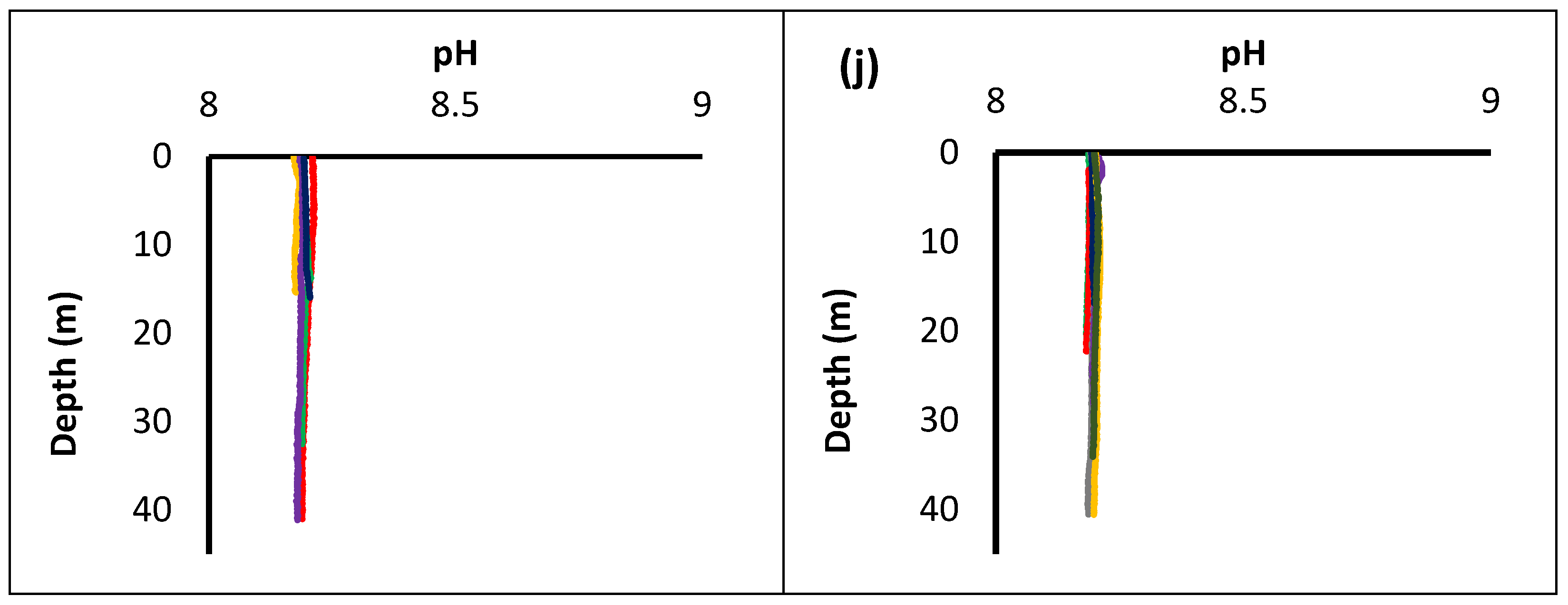
3.2. Physicochemical Parameters
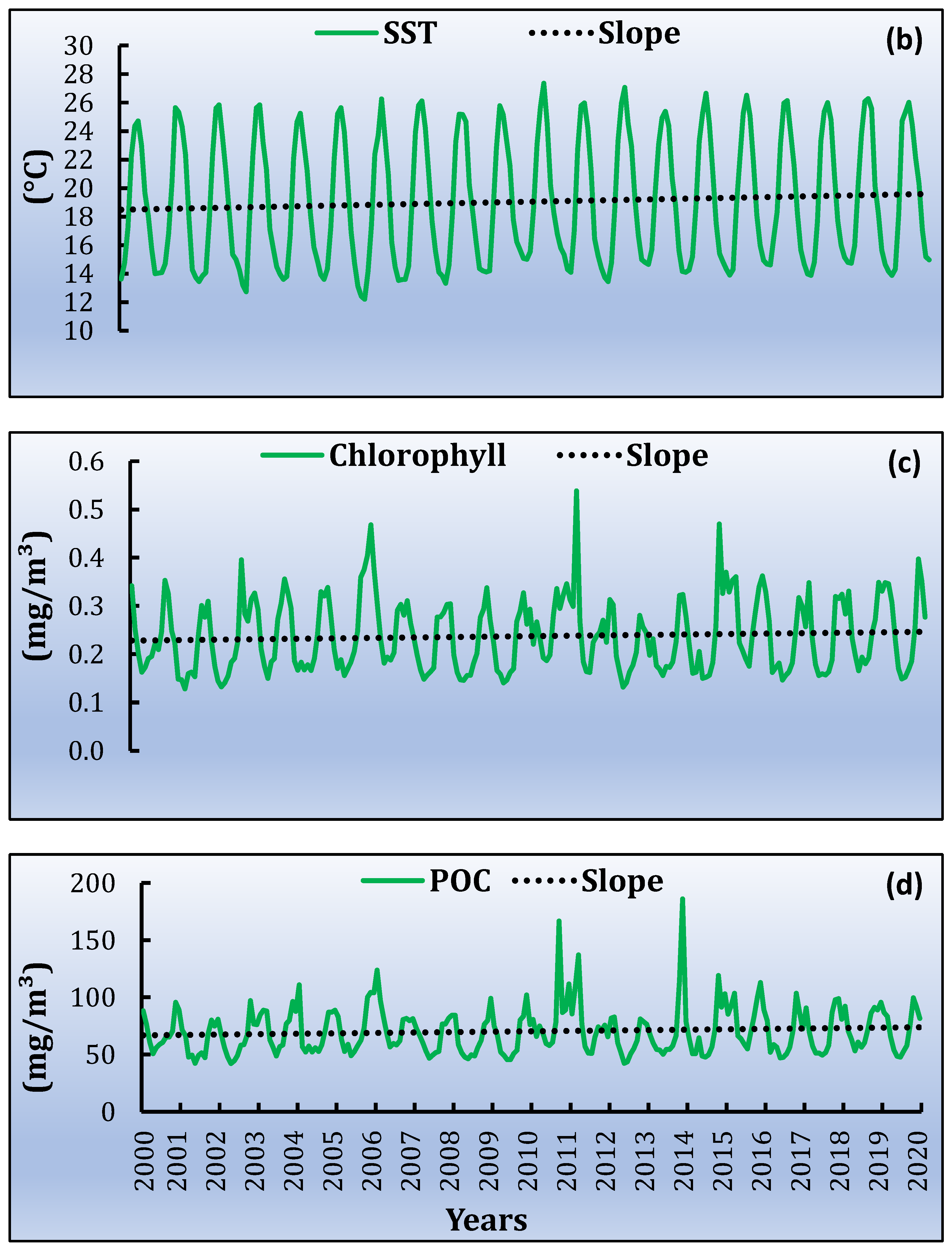
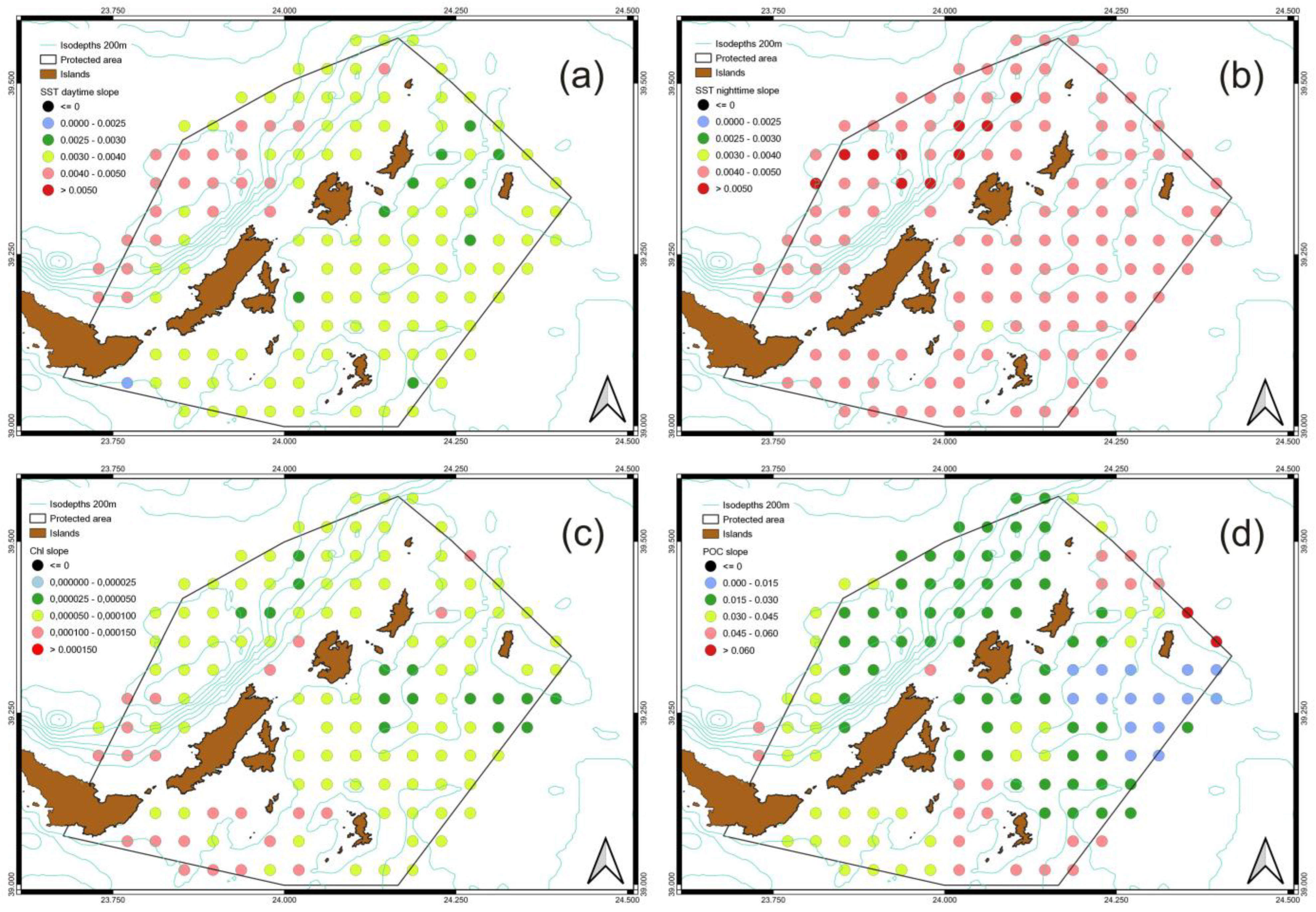
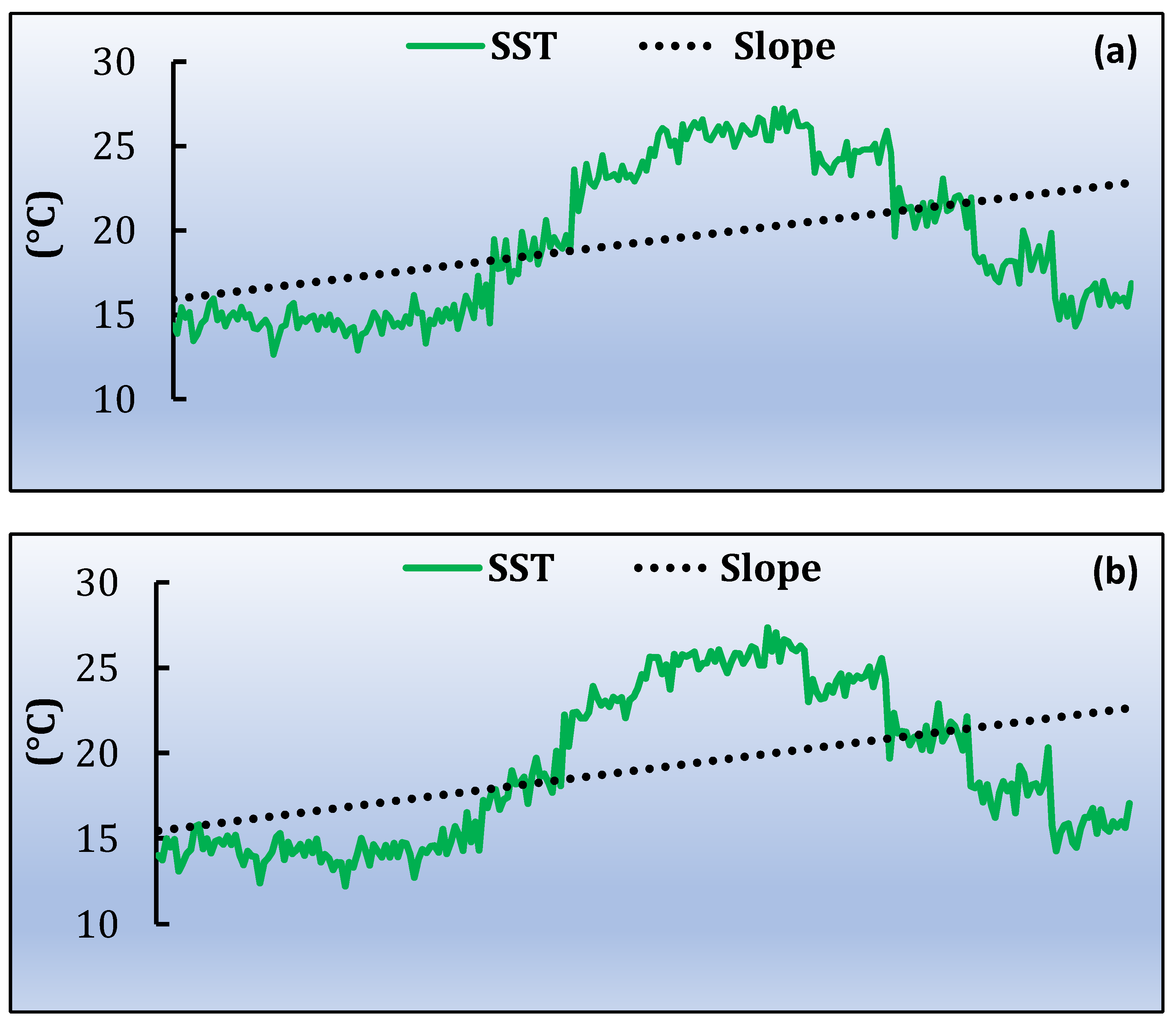
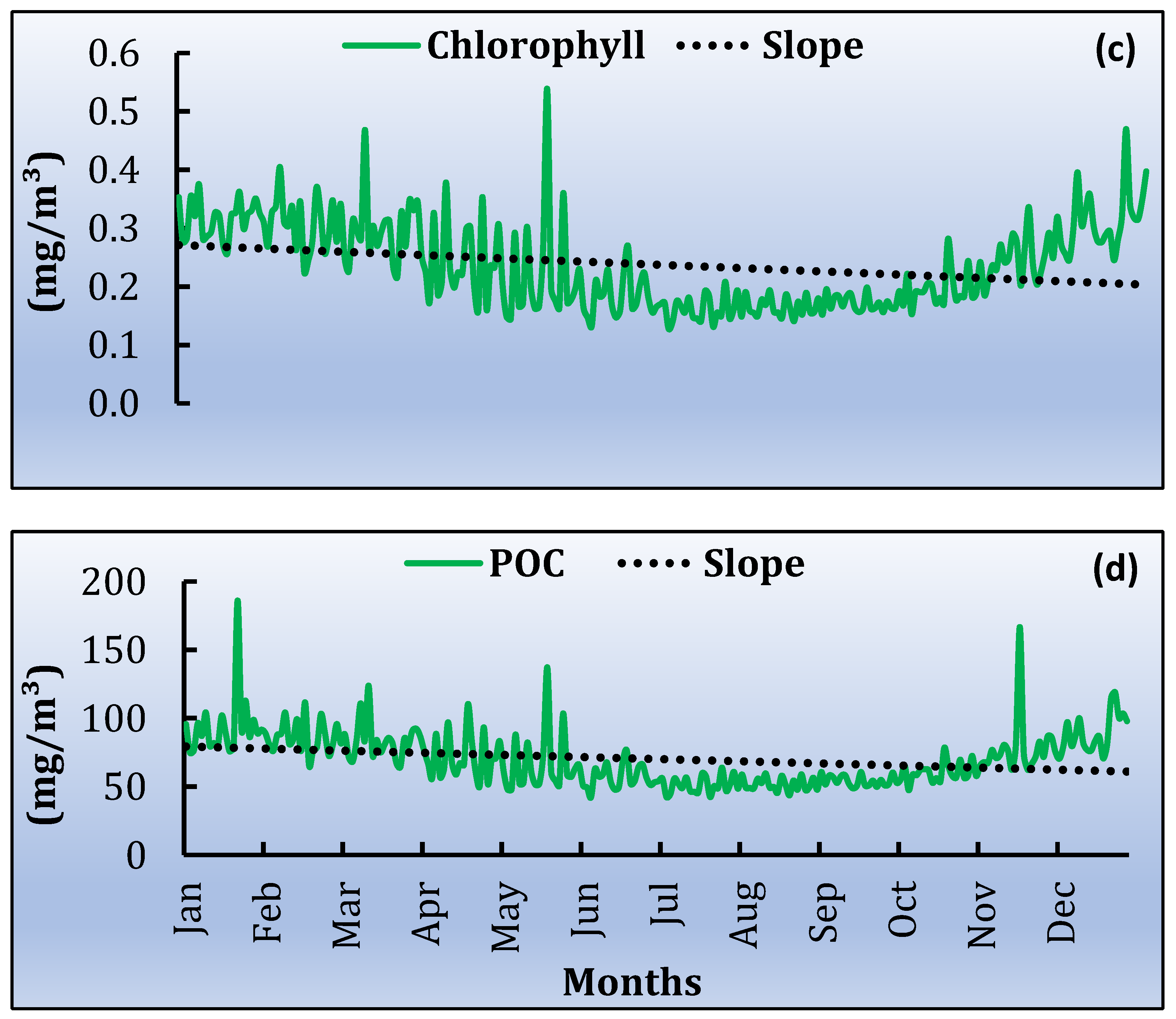
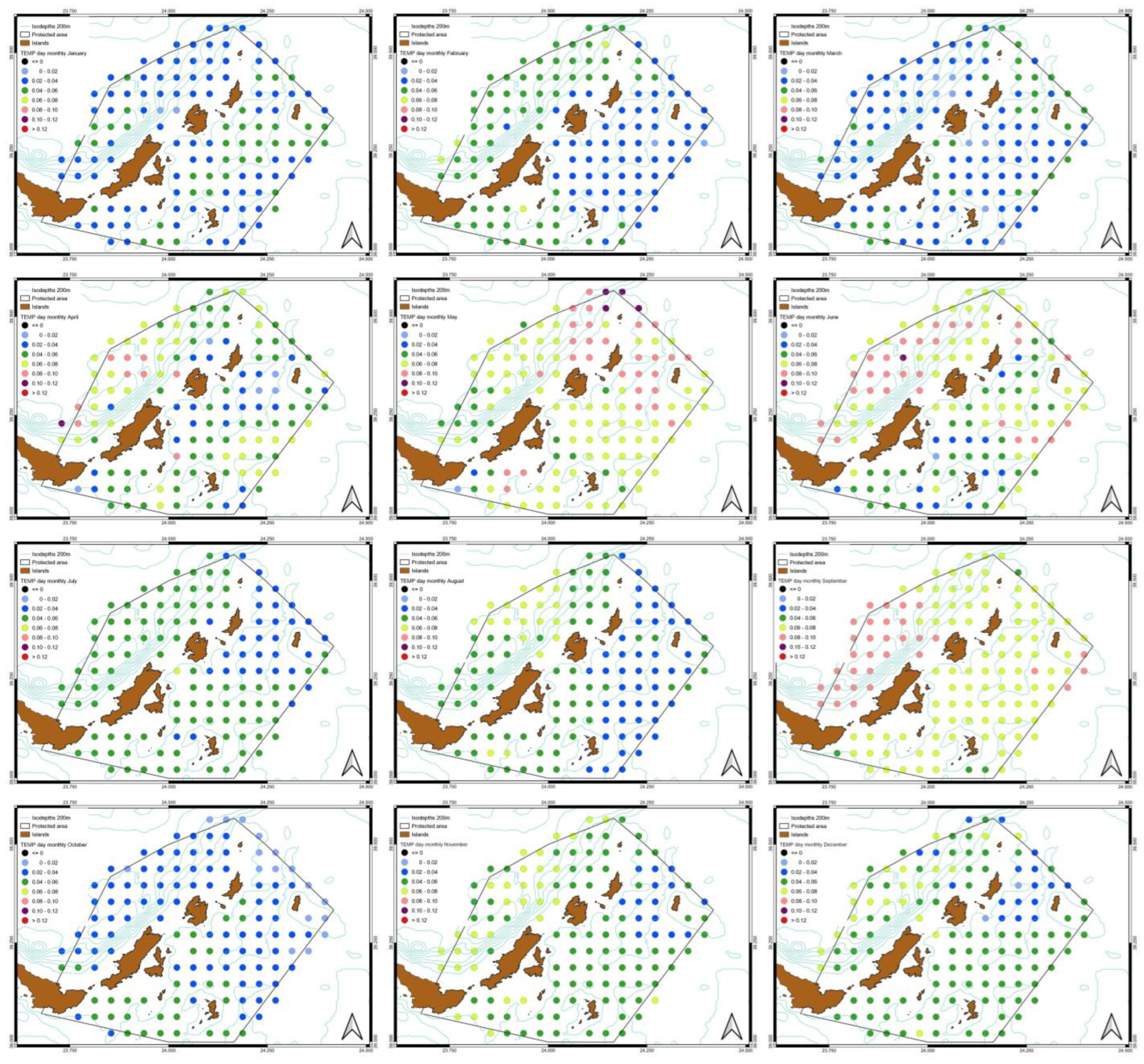
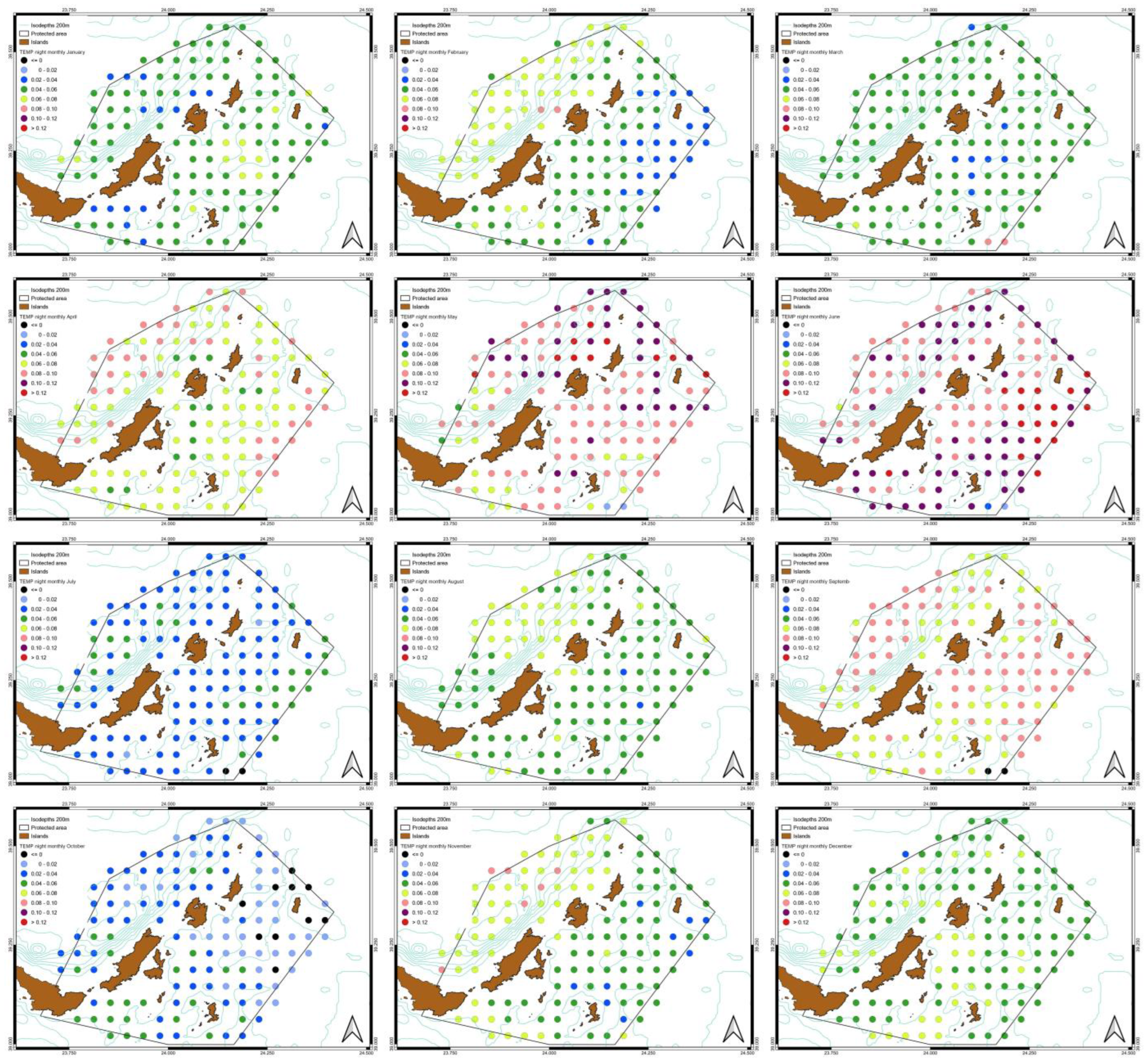
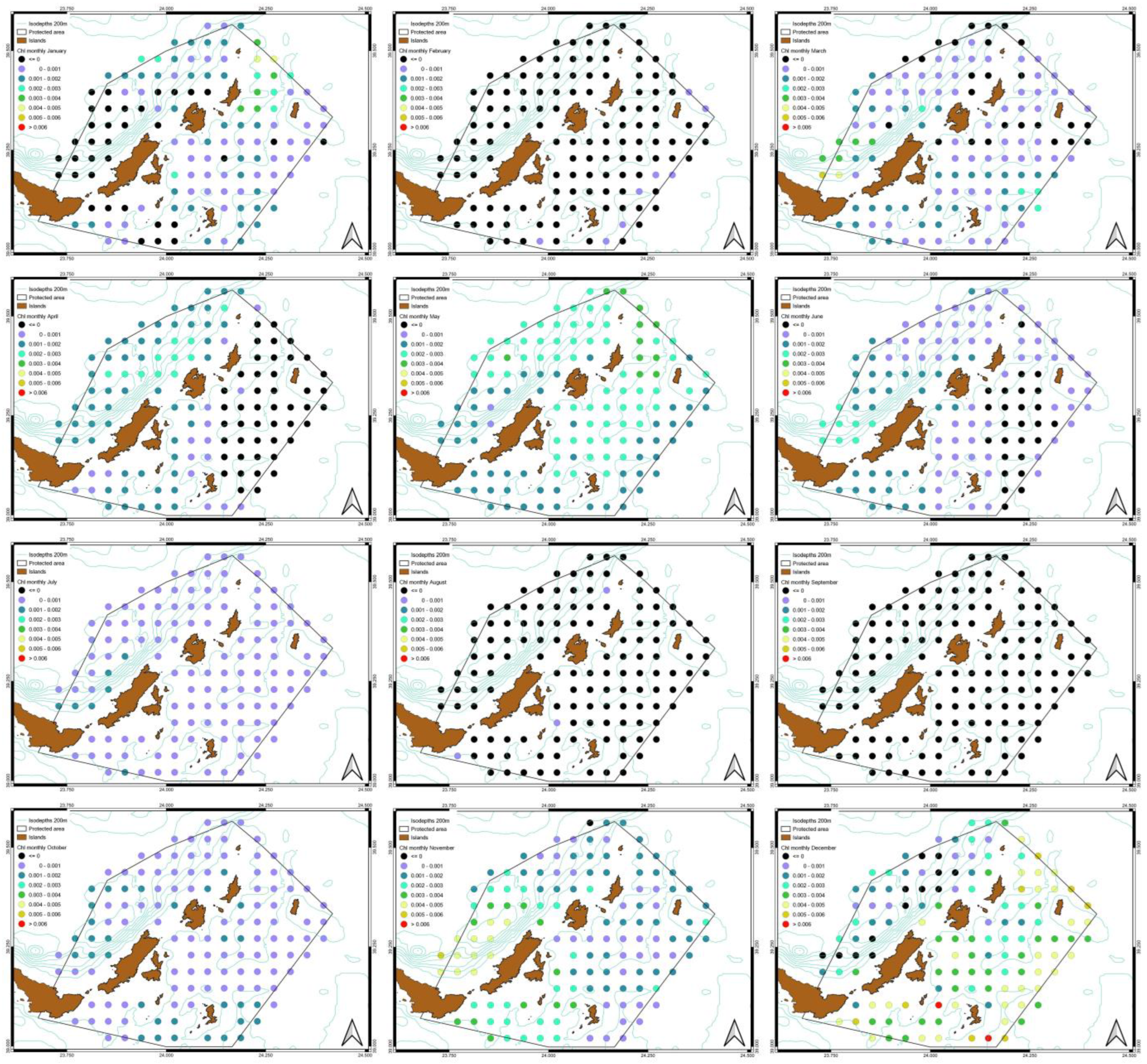
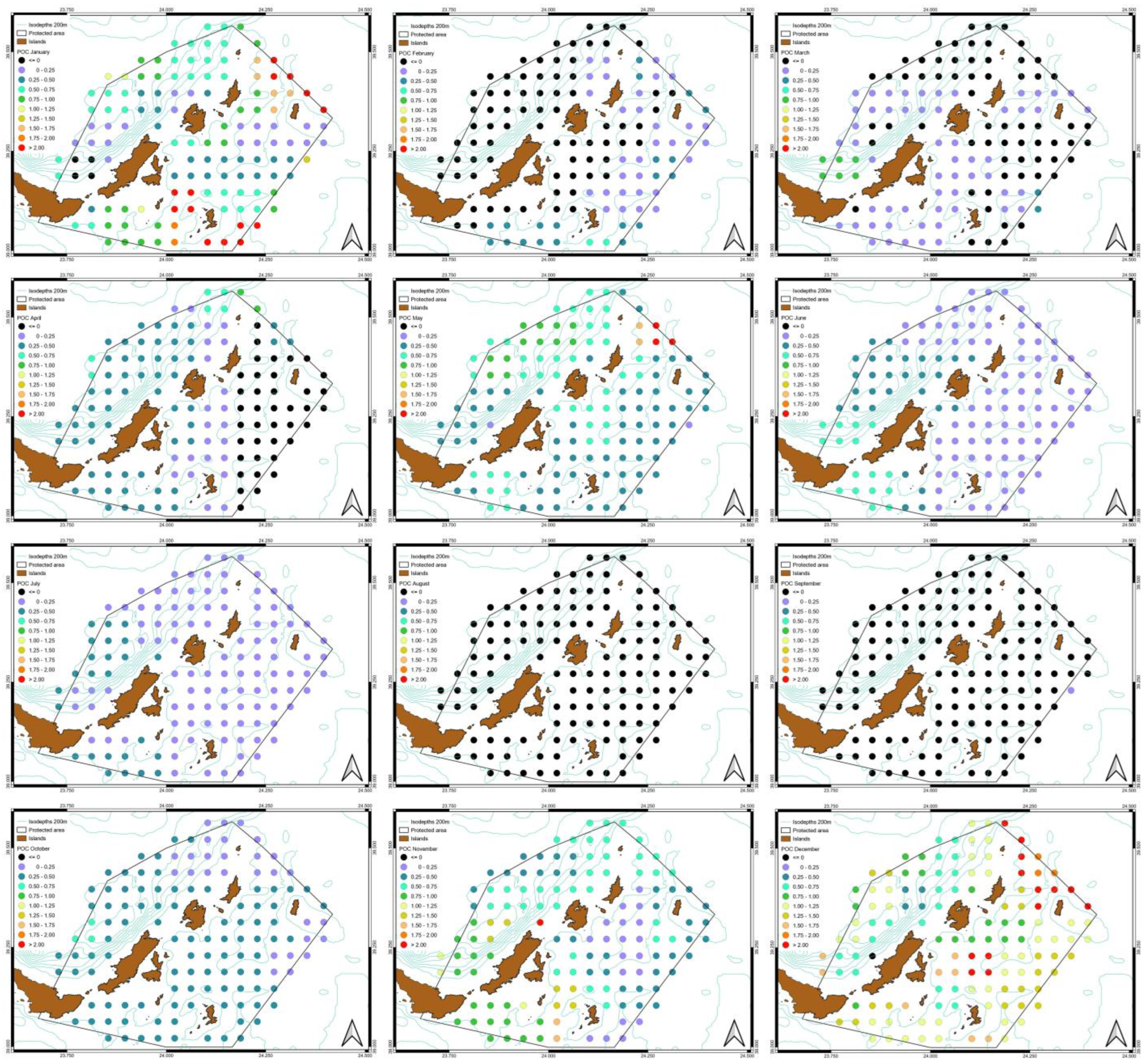
3.3. Spatiotemporal Variati Ons of SST, Chl-a and Particulate Organic Carbon [POC]
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Day, J.; Dudley, N.; Hockings, M.; Holmes, G.; Laffoley, D.; Stolton, S.; Wells, S.; Wenzel, L. Guidelines for Applying the IUCN Protected Area Management Categories to Marine Protected Areas, 2nd ed.; World Comission on Protected Areas: Gland, Switzerland, 2019; ISBN 9782831719412. [Google Scholar]
- Reuchlin-Hugenholtz, E.; McKenzie, E. Marine Protected Areas: Smart Investments in Ocean Health; Tanzer, J., Ed.; WWF: Gland, Switzerland, 2015; ISBN 9782940529216. [Google Scholar]
- Bertzk, B.; Corrigan, C.; Kemsey, J.; Kenney, S.; Ravilious, C.; Besançon, C.; Burgess, N. Protected Planet Report 2012: Tracking Progress Towards Global Targets for Protected Areas; IUCN: Gland, Switzerland, 2012; ISBN 9789280731897. [Google Scholar]
- Sala, E.; Giakoumi, S. No-Take Marine Reserves Are the Most Effective Protected Areas in the Ocean. ICES J. Mar. Sci. 2018, 75, 1166–1168. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Y.; Chen, Y.; Liu, K. Remote sensing approach for the estimation of particulate organic carbon in coastal waters based on suspended particulate concentration and particle median size. Mar. Pollut. Bull. 2020, 158, 111382. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Minnett, P.J. High latitude sea surface temperatures derived from MODIS infrared measurements. Remote Sens. Environ. 2020, 251, 112094. [Google Scholar] [CrossRef]
- Minnett, P.J.; Alvera-Azcárate, A.; Chin, T.M.; Corlett, G.K.; Gentemann, C.L.; Karagali, I.; Li, X.; Marsouin, A.; Marullo, S.; Maturi, E.; et al. Half a Century of Satellite Remote Sensing of Sea-Surface Temperature. Remote Sens. Environ. 2019, 233, 111366. [Google Scholar] [CrossRef]
- Kilpatrick, K.A.; Podestá, G.; Walsh, S.; Williams, E.; Halliwell, V.; Szczodrak, M.; Brown, O.B.; Minnett, P.J.; Evans, R. A decade of sea surface temperature from MODIS. Remote Sens. Environ. 2015, 165, 27–41. [Google Scholar] [CrossRef]
- Pahlevan, N.; Smith, B.; Binding, C.; Gurlin, D.; Li, L.; Bresciani, M.; Giardino, C. Hyperspectral retrievals of phytoplankton absorption and chlorophyll-a in inland and nearshore coastal waters. Remote Sens. Environ. 2021, 253, 112200. [Google Scholar] [CrossRef]
- Otsuka, A.Y.; Feitosa, F.A.N.; Flores-Montes, M.J.; Silva, A. Dynamics of Chlorophyll a and Oceanographic Parameters in the Coastal Zone: Barra das Jangadas-Pernambuco, Brazil. J. Coast. Res. 2016, 32, 490–499. [Google Scholar] [CrossRef]
- Fan, H.; Wang, X.; Zhang, H.; Yu, Z. Spatial and temporal variations of particulate organic carbon in the Yellow-Bohai Sea over 2002–2016. Sci. Rep. 2018, 8, 7971. [Google Scholar] [CrossRef]
- Guan, Q.; Feng, L.; Hou, X.; Schurgers, G.; Zheng, Y.; Tang, J. Eutrophication changes in fifty large lakes on the Yangtze Plain of China derived from MERIS and OLCI observations. Remote Sens. Environ. 2020, 246, 111890. [Google Scholar] [CrossRef]
- Zhang, L.; Liao, Q.; Gao, R.; Luo, R.; Liu, C.; Zhong, J.; Wang, Z. Spatial variations in diffusive methane fluxes and the role of eutrophication in a subtropical shallow lake. Sci. Total Environ. 2021, 759, 143495. [Google Scholar] [CrossRef]
- Henryson, K.; Kätterer, T.; Tidåker, P.; Sundberg, C. Soil N2O emissions, N leaching and marine eutrophication in life cycle assessment—A comparison of modelling approaches. Sci. Total Environ. 2020, 725, 138332. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.M.; Townsend-Small, A.; Zastepa, A.; Watson, S.B.; Brandes, J.A. Methane and nitrous oxide measured throughout Lake Erie over all seasons indicate highest emissions from the eutrophic Western Basin. J. Great Lakes Res. 2020, 46, 1604–1614. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, M.; Tang, X. Eutrophication assessment of seasonal urban lakes in China Yangtze River Basin using Landsat 8-derived Forel-Ule index: A six-year (2013–2018) observation. Sci. Total Environ. 2020, 745, 135392. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, P.; Zhang, Z. Interaction between carbon dioxide emissions and eutrophication in a drinking water reservoir: A three-dimensional ecological modeling approach. Sci. Total Environ. 2019, 663, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, S.; Vethamony, P.; Sadooni, F.N.; Al-kuwari, H.A.; Al-khayat, J.A.; Seegobin, V.O.; Govil, H.; Nasir, S. Detection of Wakashio oil spill off Mauritius using Sentinel-1 and 2 data: Capability of sensors, image transformation methods and. Environ. Pollut. 2021, 274, 116618. [Google Scholar] [CrossRef]
- Balogun, A.L.; Yekeen, S.T.; Pradhan, B.; Wan Yusof, K.B. Oil spill trajectory modelling and environmental vulnerability mapping using GNOME model and GIS. Environ. Pollut. 2021, 268, 115812. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.R.; Grubesic, T.H. A spatiotemporal analysis of oil spill severity using a multi-criteria decision framework. Ocean Coast. Manag. 2021, 199, 105410. [Google Scholar] [CrossRef]
- Obida, C.B.; Blackburn, G.A.; Whyatt, J.D.; Semple, K.T. Counting the cost of the Niger Delta’s largest oil spills satellite remote sensing reveals extensive environmental damage with 1million people in the impact zone. Sci. Total Environ. 2021, 775, 145854. [Google Scholar] [CrossRef]
- Klemas, V. Remote sensing techniques for studying coastal ecosystems: An overview. J. Coast. Res. 2011, 27, 2–17. [Google Scholar] [CrossRef]
- Ferreira, M.A.; Andrade, F.; Mendes, R.N.; Paula, J. Use of satellite remote sensing for coastal conservation in the eastern african coast: Advantages and shortcomings. Eur. J. Remote Sens. 2012, 45, 293–304. [Google Scholar] [CrossRef]
- Namukose, M.; Msuya, F.E.; Ferse, S.C.A.; Slater, M.J.; Kunzmann, A. Growth performance of the sea cucumber Holothuria scabra and the seaweed Eucheuma denticulatum: Integrated mariculture and effects on sediment organic characteristics. Aquac. Environ. Interactactions 2016, 8, 179–189. [Google Scholar] [CrossRef]
- Parra-luna, M.; Martín-pozo, L.; Hidalgo, F.; Zafra-Gómez, A. Common sea urchin (Paracentrotus lividus) and sea cucumber of the genus Holothuria as bioindicators of pollution in the study of chemical contaminants in aquatic media. A revision. Ecol. Indic. 2020, 113, 106185. [Google Scholar] [CrossRef]
- Mezali, K.; Soualili, D.L. The Ability of Holothurians to Select Sediment Particles and Organic Matter. SPC Beche Mer Inf. Bull. 2013, 1, 38–43. [Google Scholar]
- Kovos, D.; Karagiannis, G. Field study on the “National Marine Park” of Alonissos, Greece. J. Tour. Res. 2018, 19, 243–253. [Google Scholar]
- Karamanlidis, A.A.; Androukaki, E.; Adamantopoulou, S.; Chatzispyrou, A.; Johnson, W.M.; Kotomatas, S.; Papadopoulos, A.; Paravas, V.; Paximadis, G.; Pires, R.; et al. Assessing Accidental Entanglement as a Threat to the Mediterranean Monk Seal Monachus Monachus. Endanger. Species Res. 2008, 5, 205–213. [Google Scholar] [CrossRef]
- Oikonomou, Z.-S.; Dikou, A. Integrating Conservation and Development at the National Marine Park of Alonissos, Northern Sporades, Greece: Perception and Practice. Environ. Manag. 2008, 42, 847–866. [Google Scholar] [CrossRef]
- Konaxis, I. Alonissos Island and the Northern Sporades Marine National Park as a Strategic Socio-Economic Node for The Culture of the Aegean Sea. Am. Res. J. Humanit. Soc. Sci. 2020, 3, 49–53. [Google Scholar]
- Trivourea, M.N.; Karamanlidis, A.A.; Tounta, E.; Dendrinos, P.; Kotomatas, S. People and the Mediterranean Monk Seal (Monachus monachus): A Study of the Socioeconomic Impacts of the National Marine Park of Alonissos, Northern Sporades, Greece. Aquat. Mamm. 2011, 37, 305–318. [Google Scholar] [CrossRef][Green Version]
- Androulidakis, Y.S.; Krestenitis, Y.N.; Psarra, S. Coastal upwelling over the North Aegean Sea: Observations and simulations. Cont. Shelf Res. 2017, 149, 32–51. [Google Scholar] [CrossRef]
- Poulos, S.E.; Drakopoulos, P.G.; Collins, M.B. Seasonal variability in sea surface oceanographic conditions in the Aegean Sea (Eastern Mediterranean): An overview. J. Mar. Syst. 1997, 13, 225–244. [Google Scholar] [CrossRef]
- Al-Wassai, F.A.; Kalyankar, N.V. Major Limitations of Satellite images. J. Glob. Res. Comput. Sci. 2013, 4, 51–59. [Google Scholar]
- Gao, J. Bathymetric mapping by means of remote sensing: Methods, accuracy and limitations. Prog. Phys. Geogr. 2009, 33, 103–116. [Google Scholar] [CrossRef]
- Lawson, K.; Larson, N.G. CTD. In Encyclopedia of Ocean Sciences; Steele, J.H., Ed.; Sea-Bird Electronics Inc.: Bellevue, DC, USA, 2001; pp. 579–588. ISBN 9780122274305. [Google Scholar]
- Fingas, M. Remote Sensing for Marine Management, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2018; ISBN 9780128050521. [Google Scholar]
- Lotfinasabasl, S.; Gunale, V.R.; Khosroshahi, M. Applying geographic information systems and remote sensing for water quality assessment of mangrove forest. Acta Ecol. Sin. 2018, 38, 135–143. [Google Scholar] [CrossRef]
- Cai, J.-Q.; Liu, X.-M.; Gao, Z.-J.; Li, L.-L.; Wang, H. Chlorophylls derivatives: Photophysical properties, assemblies, nanostructures and biomedical applications. Mater. Today 2021, 45, 77–92. [Google Scholar] [CrossRef]
- Nazeer, M.; Nichol, J.E. Development and application of a remote sensing-based Chlorophyll-a concentration prediction model for complex coastal waters of Hong Kong. J. Hydrol. 2016, 532, 80–89. [Google Scholar] [CrossRef]
- Yu, X.; Shen, J. A data-driven approach to simulate the spatiotemporal variations of chlorophyll-a in Chesapeake Bay. Ocean Model. 2021, 159, 101748. [Google Scholar] [CrossRef]
- Chawla, I.; Karthikeyan, L.; Mishra, A.K. A review of remote sensing applications for water security: Quantity, quality, and extremes. J. Hydrol. 2020, 585, 124826. [Google Scholar] [CrossRef]
- Gholizadeh, M.H.; Melesse, A.M.; Reddi, L. A Comprehensive Review on Water Quality Parameters Estimation Using Remote Sensing Techniques. Sensors 2016, 16, 1298. [Google Scholar] [CrossRef]
- Bennion, D.H.; Warner, D.M.; Esselman, P.C.; Hobson, B.; Kieft, B. A comparison of chlorophyll a values obtained from an autonomous underwater vehicle to satellite-based measures for Lake Michigan. J. Great Lakes Res. 2019, 45, 726–734. [Google Scholar] [CrossRef]
- Wei, G.F.; Tang, D.L.; Wang, S. Distribution of chlorophyll and harmful algal blooms (HABs): A review on space based studies in the coastal environments of Chinese marginal seas. Adv. Space Res. 2008, 41, 12–19. [Google Scholar] [CrossRef]
- Rodríguez-López, L.; Duran-Llacer, I.; González-Rodríguez, L.; Abarca-del-Rio, R.; Cárdenas, R.; Parra, O.; Martínez-Retureta, R.; Urrutia, R. Spectral analysis using LANDSAT images to monitor the chlorophyll-a concentration in Lake Laja in Chile. Ecol. Inform. 2020, 60, 101183. [Google Scholar] [CrossRef]
- Flores-cervantes, D.X.; Plata, D.L.; Macfarlane, J.K.; Reddy, C.M.; Gschwend, P.M. Black carbon in marine particulate organic carbon: Inputs and cycling of highly recalcitrant organic carbon in the Gulf of Maine. Mar. Chem. 2009, 113, 172–181. [Google Scholar] [CrossRef]
- Kharbush, J.J.; Close, H.G.; Van Mooy, B.A.S.; Arnosti, C.; Smittenberg, R.H.; Le Moigne, F.A.C.; Mollenhauer, G.; Scholz-Böttcher, B.; Obreht, I.; Koch, B.P.; et al. Particulate Organic Carbon Deconstructed: Molecular and Chemical Composition of Particulate Organic Carbon in the Ocean. Front. Mar. Sci. 2020, 7, 518. [Google Scholar] [CrossRef]
- McNichol, A.P.; Aluwihare, L.I. The Power of Radiocarbon in Biogeochemical Studies of the Marine Carbon Cycle: Insights from Studies of Dissolved and Particulate Organic Carbon (DOC and POC). Chem. Rev. 2007, 107, 443–466. [Google Scholar] [CrossRef]
- Son, B.Y.; Gardner, W.D.; Mishonov, A.V.; Jo, M. Multispectral remote-sensing algorithms for particulate organic carbon (POC): The Gulf of Mexico. Remote Sens. Environ. 2009, 113, 50–61. [Google Scholar] [CrossRef]
- Xu, J.; Lei, S.; Bi, S.; Li, Y.; Lyu, H.; Xu, J. Tracking spatio-temporal dynamics of POC sources in eutrophic lakes by remote sensing Tracking spatio-temporal dynamics of POC sources in eutrophic lakes by remote sensing. Water Res. 2019, 168, 115162. [Google Scholar] [CrossRef]
- Yakushev, E.V.; Wallhead, P.; Renaud, P.E.; Ilinskaya, A.; Protsenko, E.; Yakubov, S.; Pakhomova, S.; Sweetman, A.K.; Dunlop, K.; Berezina, A.; et al. Understanding the Biogeochemical Impacts of Fish Farms Using a Benthic-Pelagic Model. Water 2020, 12, 2384. [Google Scholar] [CrossRef]
- Noroi, G.Á.; Glud, R.N.; Gaard, E.; Simonsen, K. Environmental Impacts of Coastal Fish Farming: Carbon and Nitrogen Budgets for Trout Farming in Kaldbaksfjørour (Faroe Islands). Mar. Ecol. Prog. Ser. 2011, 431, 223–241. [Google Scholar]
- Guangjia, J.; Ronghua, M.; Loiselle, S.A.; Duan, H.; Su, W.; Huang, C.; Jie Yang, W.Y. Remote sensing of particulate organic carbon dynamics in a eutrophic lake (Taihu Lake, China). Sci. Total Environ. 2015, 532, 245–254. [Google Scholar] [CrossRef]
- Djurhuus, A.; Read, J.F.; Rogers, A.D. The spatial distribution of particulate organic carbon and microorganisms on seamounts of the South West Indian Ridge. Deep Sea Res. Part II 2017, 136, 73–84. [Google Scholar] [CrossRef]
- Legendre, L.; Michaud, J. Chlorophyll a to estimate the particulate organic carbon available as food to large zooplankton in the euphotic zone of oceans. J. Plankton Res. 1999, 21, 2067–2083. [Google Scholar] [CrossRef]
- Abou Samra, R.M.; El-Gammal, M.; Eissa, R. Oceanographic Factors of Oil Pollution Dispersion Offshore the Nile Delta (Egypt) Using GIS. Environ. Sci. Pollut. Res. 2021, 28, 25830–25843. [Google Scholar] [CrossRef] [PubMed]
- Siepak, J. Total Organic Carbon (TOC) as a Sum Parameter of Water Pollution in Selected Polish Rivers (Vistula, Odra, and Warta). Acta Hydrochim. Hydrobiol. 1999, 27, 282–285. [Google Scholar] [CrossRef]
- Chaithanya, M.S.; Das, B.; Vidya, R. Assessment of metals pollution and subsequent ecological risk in water, sediments and vegetation from a shallow lake: A case study from Ranipet industrial town, Tamil Nadu, India. Int. J. Environ. Anal. Chem. 2021, 1–18. [Google Scholar] [CrossRef]
- Minnett, P.J.; Brown, O.B.; Evans, R.H.; Key, E.L.; Kearns, E.J.; Kilpatrick, K.; Kumar, A.; Maillet, K.A.; Szczodrak, G. Sea-Surface Temperature Measurements from the Moderate-Resolution Imaging Spectroradiometer (MODIS) on Aqua and Terra. Int. Geosci. Remote Sens. Symp. 2004, 7, 4576–4579. [Google Scholar] [CrossRef]
- Gmelin World Register of Marine Species (WoRMS). Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=125182 (accessed on 9 August 2022).
- Rakaj, A.; Fianchini, A.; Boncagni, P.; Scardi, M.; Cataudella, S. Artificial Reproduction of Holothuria Polii: A New Candidate for Aquaculture. Aquaculture 2019, 498, 444–453. [Google Scholar] [CrossRef]
- Tolon, M.T.; Engin, S. Gonadal Development of the Holothurian Holothuria Polii (Delle Chiaje, 1823) in Spawning Period at the Aegean Sea (Mediterranean Sea). Ege J. Fish. Aquat. Sci. 2019, 36, 379–385. [Google Scholar] [CrossRef]
- Toscano, A.; Cirino, P. First Evidence of Artificial Fission in Two Mediterranean Species of Holothurians: Holothuria Tubulosa and Holothuria Polii. Turk. J. Fish. Aquat. Sci. 2018, 18, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Slimane-tamacha, F.; Soualili, D.L.; Mezali, K. Reproductive biology of Holothuria (Roweothuria) poli (Holothuroidea: Echinodermata) from Oran Bay, Algeria. SPC Beche Mer Inf. Bull. 2019, 39, 47–53. [Google Scholar]
- Gonzalez-Wanguemert, M.; Valente, S.; Aydin, M. Effects of fishery protection on biometry and genetic structure of two target sea cucumber species from the Mediterranean Sea. Hydrobiologia 2015, 743, 65–74. [Google Scholar] [CrossRef]
- Simunovic, A.; Grubelic, I. A contribution to the knowledge of the species Holothuria tubulosa GMELIN, 1788 (Holothuria, Echinodermata) in the coastal area of the central eastern Adriatic. Acta Adriat. 1998, 39, 13–23. [Google Scholar]
- Neofitou, N.; Lolas, A.; Ballios, I.; Skordas, K.; Tziantziou, L.; Vafidis, D. Contribution of Sea Cucumber Holothuria Tubulosa on Organic Load Reduction from Fish Farming Operation. Aquaculture 2019, 501, 97–103. [Google Scholar] [CrossRef]
- Antoniadou, C.; Vafidis, D. Population structure of the traditionally exploited holothurian Holothuria tubulosa in the south Aegean Sea. Cah. Biol. Mar. 2011, 52, 171–175. [Google Scholar]
- Kazanidis, G.; Lolas, A.; Vafidis, D. Reproductive cycle of the traditionally exploited sea cucumber Holothuria tubulosa (Holothuroidea: Aspidochirotida) in Pagasitikos Gulf, western Aegean Sea, Greece. Turk. J. Zool. 2014, 38, 306–315. [Google Scholar] [CrossRef]
- Kazanidis, G.; Antoniadou, C.; Lolas, A.P.; Neofitou, N.; Vafidis, D.; Chintiroglou, C.; Neofitou, C. Population Dynamics and Reproduction of Holothuria Tubulosa (Holothuroidea: Echinodermata) in the Aegean Sea. J. Mar. Biol. Assoc. UK 2010, 90, 895–901. [Google Scholar] [CrossRef]
- Despalatovic, M.; Grubelic, I.; Simunovic, A.; Antolic, B.; Zuljevic, A. Reproductive biology of the holothurian Holothuria tubulosa (Echinodermata) in the Adriatic Sea. Mar. Biol. Assoc. UK 2004, 84, 409–414. [Google Scholar] [CrossRef]
- Yuval, B.; Sudai, L.; Ziv, Y. Abundance and Diversity of Holothuroids in Shallow Habitats of the Northern Red Sea. J. Mar. Biol. 2014, 2014, 631309. [Google Scholar] [CrossRef]
- Lampe-Ramdoo, K.; Moothien Pillay, R.; Conand, C. An assessment of holothurian diversity, abundance and distribution in the shallow lagoons of Mauritius. SPC Beche Mer Inf. Bull. 2014, 34, 17–24. [Google Scholar]
- Arsad, N.A.; Othman, R.; Raehanah, S.; Shaleh, M.; Abdullah, F.C.; Matsumoto, M.M. Effects of physicochemical parameters on the reproductive pattern of sea cucumber Holothuria scabra in Sabah. Songklanakarin J. Sci. Technol. 2020, 42, 109–116. [Google Scholar]
- Dissanayake, D.C.T.; Stefansson, G. Abundance and distribution of commercial sea cucumber species in the coastal waters of Sri Lanka. Aquat. Living Resour. 2011, 313, 303–313. [Google Scholar] [CrossRef]
- Collard, M.; Eeckhaut, I.; Dehairs, F.; Dubois, P. Acid–Base Physiology Response to Ocean Acidification of Two Ecologically and Economically Important Holothuroids from Contrasting Habitats, Holothuria Scabra and Holothuria Parva. Environ. Sci. Pollut. Res. 2014, 21, 13602–13614. [Google Scholar] [CrossRef]
- Herna, C.; Clemente, S.; Tuya, F. Is there a link between the type of habitat and the patterns of abundance of holothurians in shallow rocky reefs? Hydrobiologia 2006, 571, 191–199. [Google Scholar] [CrossRef]
- Aydin, M. Density and Biomass of Commercial Sea Cucumber Species Relative to Depth in the Northern Aegean Sea. Thalass. Int. J. Mar. Sci. 2019, 35, 541–550. [Google Scholar] [CrossRef]
- Nazari-Sharabian, M.; Ahmad, S.; Moses, K. Climate Change and Eutrophication: A Short Review. Eng. Technol. Appl. Sci. Res. 2018, 8, 3668–3672. [Google Scholar] [CrossRef]
- Nurdin, S.; Mustapha, M.A.; Lihan, T. The Relationship between Sea Surface Temperature and Chlorophyll-a Concentration in Fisheries Aggregation Area in the Archipelagic Waters of Spermonde Using Satellite Images. AIP Conf. Proc. 2013, 1571, 466–472. [Google Scholar] [CrossRef]
- Bulteel, P.; Jangoux, M.; Coulon, P. Biometry, Bathymetric Distribution, and Reproductive Cycle of the Holothuroid from Mediterranean Seagrass Beds. Mar. Ecol. 1992, 13, 53–62. [Google Scholar] [CrossRef]
- Vafeiadou, A.M.; Antoniadou, C.; Vafidis, D.; Fryganiotis, K.; Chintiroglou, C.; Density, K.; Sea, A. Density and Biometry of the Exploited Holothurian Holothuria Tubulosa at the Dodecanese, South Aegean Sea. Rapp. Comm. Int. Mer Medit. 2010, 39, 661. [Google Scholar]
- Lavitra, T.; Fohy, N.; Gestin, P.; Rasolofonirina, R.; Eeckhaut, I. Effect of water temperature on the survival and growth of endobenthic Holothuria scabra (Echinodermata: Holothuroidea) juveniles reared in outdoor ponds. SPC Beche Mer Inf. Bull. 2010, 30, 25–28. [Google Scholar]
- Yang, H.; Yuan, X.; Zhou, Y.; Mao, Y.; Zhang, T.; Liu, Y. Effects of body size and water temperature on food consumption and growth in the sea cucumber Apostichopus japonicus (Selenka) with special reference to aestivation. Aquac. Res. 2005, 36, 1085–1092. [Google Scholar] [CrossRef]
- Marquet, N.; Conand, C.; Power, D.M.; Canário, A.V.M.; González-wangüemert, M. Sea cucumbers, Holothuria arguinensis and H. mammata, from the southern Iberian Peninsula: Variation in reproductive activity between populations from different habitats. Fish. Res. 2017, 191, 120–130. [Google Scholar] [CrossRef]
- Zamora, L.N.; Jeffs, A.G. Feeding, metabolism and growth in response to temperature in juveniles of the Australasian sea cucumber, Australostichopus mollis. Aquaculture 2012, 358–359, 92–97. [Google Scholar] [CrossRef]
- Lampe, K. Holothurian density, distribution and diversity comparing sites with different degrees of exploitation in the shallow lagoons of Mauritius. SPC Beche Mer Inf. Bull. 2013, 33, 23–29. [Google Scholar]
- Seeruttun, R.; Appadoo, C.; Laxminarayana, A.; Codabaccus, B. A Study on the Factors Influencing the Growth and Survival of Juvenile Sea Cucumber, Holothuria atra, under Laboratory Conditions. Univ. Maurit. Res. J. 2008, 14, 1–15. [Google Scholar]
- Tolon, T. Effect of salinity on growth and survival of the juvenile sea cucumbers Holothuria tubulosa (Gmelin, 1788) and Holothuria poli (Delle Chiaje, 1923). Fresenius Environ. Bull. 2017, 26, 3930–3935. [Google Scholar]
- Darmaraki, S.; Somot, S.; Sevault, F.; Nabat, P.; Cabos Narvaez, W.D.; Cavicchia, L.; Djurdjevic, V.; Li, L.; Sannino, G.; Sein, D.V. Future evolution of Marine Heatwaves in the Mediterranean Sea. Clim. Dyn. 2019, 53, 1371–1392. [Google Scholar] [CrossRef]
- Xiong, X.; Salomonson, V.V.; Barnes, W.L.; Guenther, B.; Xie, X.; Sun, J. An overview of terra MODIS reflective solar bands on-orbit calibration. In Proceedings of the 2006 IEEE International Symposium on Geoscience and Remote Sensing, Denver, CO, USA, 31 July–4 August 2006; pp. 1103–1106. [Google Scholar] [CrossRef]
- Pisano, A.; Marullo, S.; Artale, V.; Falcini, F.; Yang, C.; Leonelli, F.E.; Santoleri, R.; Nardelli, B.B. New evidence of Mediterranean climate change and variability from Sea Surface Temperature observations. Remote Sens. 2020, 12, 132. [Google Scholar] [CrossRef]
- Pastor, F.; Valiente, J.A.; Khodayar, S. A warming Mediterranean: 38 years of increasing sea surface temperature. Remote Sens. 2020, 12, 2687. [Google Scholar] [CrossRef]
- Pastor, F.; Valiente, J.A.; Palau, J.L. Sea Surface Temperature in the Mediterranean: Trends and Spatial Patterns (1982–2016). Pure Appl. Geophys. 2018, 175, 4017–4029. [Google Scholar] [CrossRef]
- Tyrlis, E.; Lelieveld, J. Climatology and Dynamics of the Summer Etesian Winds over the Eastern Mediterranean. J. Atmos. Sci. 2013, 70, 3374–3396. [Google Scholar] [CrossRef]
- Poupkou, A.; Zanis, P.; Nastos, P.; Papanastasiou, D.; Melas, D.; Tourpali, K.; Zerefos, C. Present climate trend analysis of the Etesian winds in the Aegean Sea. Theor. Appl. Climatol. 2011, 106, 459–472. [Google Scholar] [CrossRef]
- Anagnostopoulou, C.; Zanis, P.; Katragkou, E.; Tegoulias, I.; Tolika, K. Recent past and future patterns of the Etesian winds based on regional scale climate model simulations. Clim. Dyn. 2014, 42, 1819–1836. [Google Scholar] [CrossRef]
- Coulon, P.; Jangoux, M. Feeding rate and sediment reworking by the holothuroid Holothuria tubulosa (Echinodermata) in a Mediterranean seagrass bed off Ischia Island, Italy. Mar. Ecol. Prog. Ser. 1993, 92, 201–204. [Google Scholar] [CrossRef]
- Ru, X.; Zhang, L.; Liu, S.; Yang, H. Plasticity of Respiratory Function Accommodates High Oxygen Demand in Breeding Sea Cucumbers. Front. Physiol. 2020, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Emiroğlu, D.; Günay, D. The effect of sea cucumber Holothuria tubulosa G. 1788 on nutrient and organic matter contents of bottom sediment of oligotrophy and hypereutrophic shores. Fresenius Environ. Bull. 2007, 16, 290–294. [Google Scholar]
- Günay, D.; Emiroğlu, D.; Tolon, T.; Özden, O.; Saygi, H. Growth and Survival Rate of Juvenile Sea Cucumbers (Holothuria Tubulosa, Gmelin, 1788) at Various Temperatures. Turk. J. Fish. Aquat. Sci. 2015, 15, 533–541. [Google Scholar] [CrossRef]
- Gao, Q.F.; Wang, Y.; Dong, S.; Sun, Z.; Wang, F. Absorption of different food sources by sea cucumber Apostichopus japonicus (Selenka) (Echinodermata: Holothuroidea): Evidence from carbon stable isotope. Aquaculture 2011, 319, 272–276. [Google Scholar] [CrossRef]
- Ren, Y.; Dong, S.; Wang, F.; Gao, Q.; Tian, X.; Liu, F. Sedimentation and sediment characteristics in sea cucumber Apostichopus japonicus (Selenka) culture ponds. Aquac. Res. 2010, 42, 14–21. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SST [Daytime] | SST [Night-Time] | Chl-a | POC | |||||
|---|---|---|---|---|---|---|---|---|
| Per Month | °C | Per Month | °C | Per Month | Mg/m3 | Per Month | Mg/m3 | |
| min | 0.002 | 0.47 | 0.0040 | 0.96 | 0.000027 | 0.0064 | 0.0035 | 0.8 |
| max | 0.005 | 1.09 | 0.0053 | 1.28 | 0.000138 | 0.0331 | 0.0722 | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christou, P.; Domenikiotis, C.; Neofitou, N.; Vafidis, D. Study of the Spatiotemporal Variability of Oceanographic Parameters and Their Relationship to Holothuria Species Abundance in a Marine Protected Area of the Mediterranean Using Satellite Imagery. Remote Sens. 2022, 14, 5946. https://doi.org/10.3390/rs14235946
Christou P, Domenikiotis C, Neofitou N, Vafidis D. Study of the Spatiotemporal Variability of Oceanographic Parameters and Their Relationship to Holothuria Species Abundance in a Marine Protected Area of the Mediterranean Using Satellite Imagery. Remote Sensing. 2022; 14(23):5946. https://doi.org/10.3390/rs14235946
Chicago/Turabian StyleChristou, Panteleimon, Christos Domenikiotis, Nikos Neofitou, and Dimitris Vafidis. 2022. "Study of the Spatiotemporal Variability of Oceanographic Parameters and Their Relationship to Holothuria Species Abundance in a Marine Protected Area of the Mediterranean Using Satellite Imagery" Remote Sensing 14, no. 23: 5946. https://doi.org/10.3390/rs14235946
APA StyleChristou, P., Domenikiotis, C., Neofitou, N., & Vafidis, D. (2022). Study of the Spatiotemporal Variability of Oceanographic Parameters and Their Relationship to Holothuria Species Abundance in a Marine Protected Area of the Mediterranean Using Satellite Imagery. Remote Sensing, 14(23), 5946. https://doi.org/10.3390/rs14235946