
Grassland Biomass Inversion Based on a Random Forest Algorithm and Drought Risk Assessment



Abstract
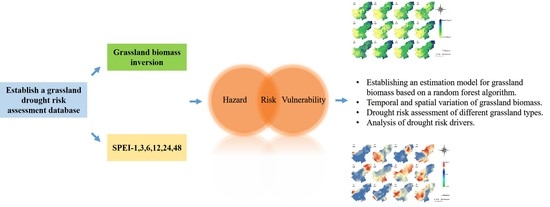
1. Introduction
2. Materials and Methods
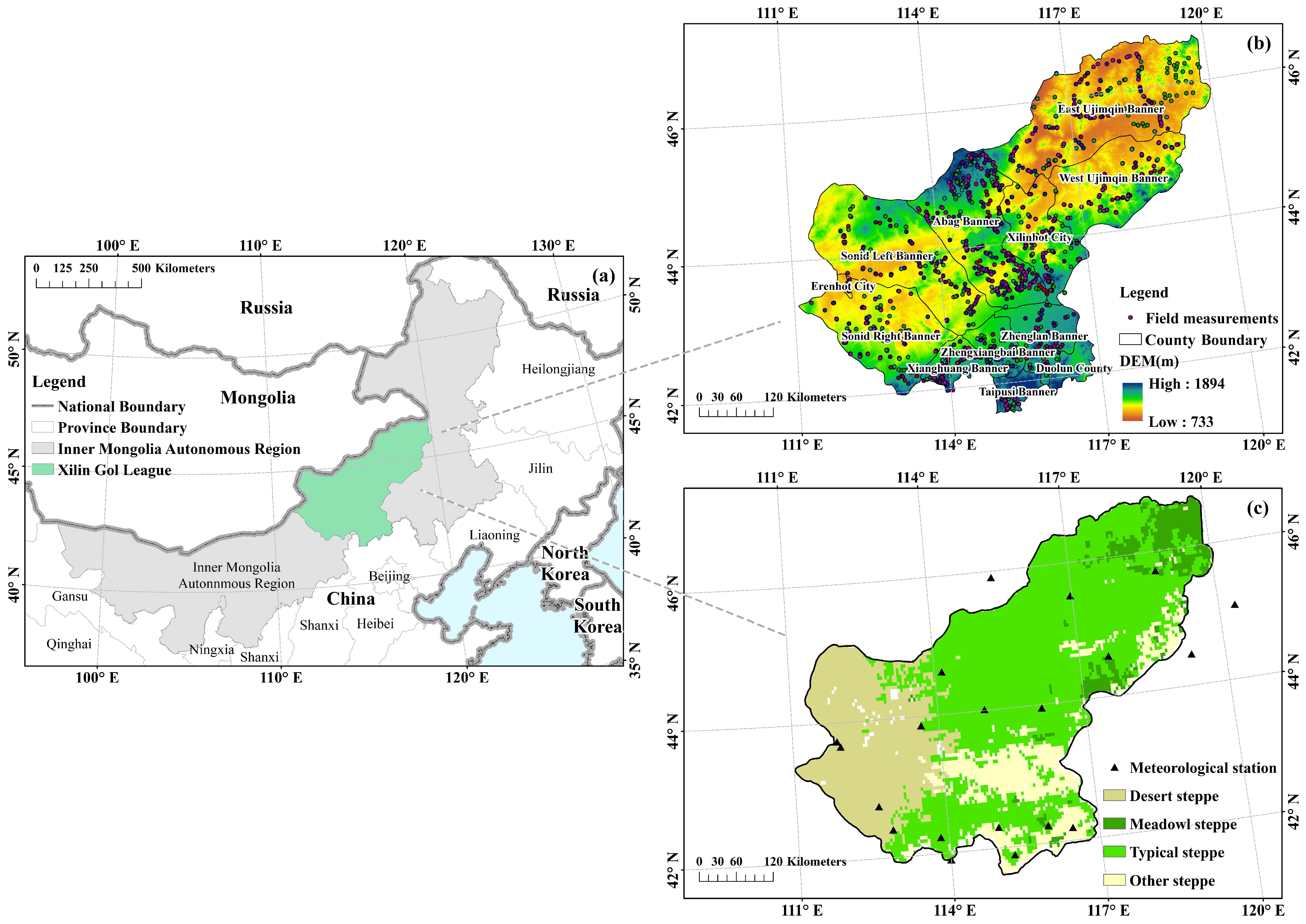
2.1. Study Area
2.2. Data Sources
2.2.1. Field Measurement Data
2.2.2. Remote Sensing Data
2.2.3. Meteorological Station Dataset
2.2.4. Socioeconomic Dataset
2.3. Methods
2.3.1. Calculation of the Vegetation Index
2.3.2. Random Forest
2.3.3. Standardized Precipitation Evapotranspiration Index
2.3.4. Spearman Correlation Analysis
2.3.5. Risk Assessment
2.3.6. Structural Equation Modeling (SEM)
3. Results
3.1. Establishing an Estimation Model for Grassland Biomass in Xilin Gol League Based on a Random Forest Algorithm
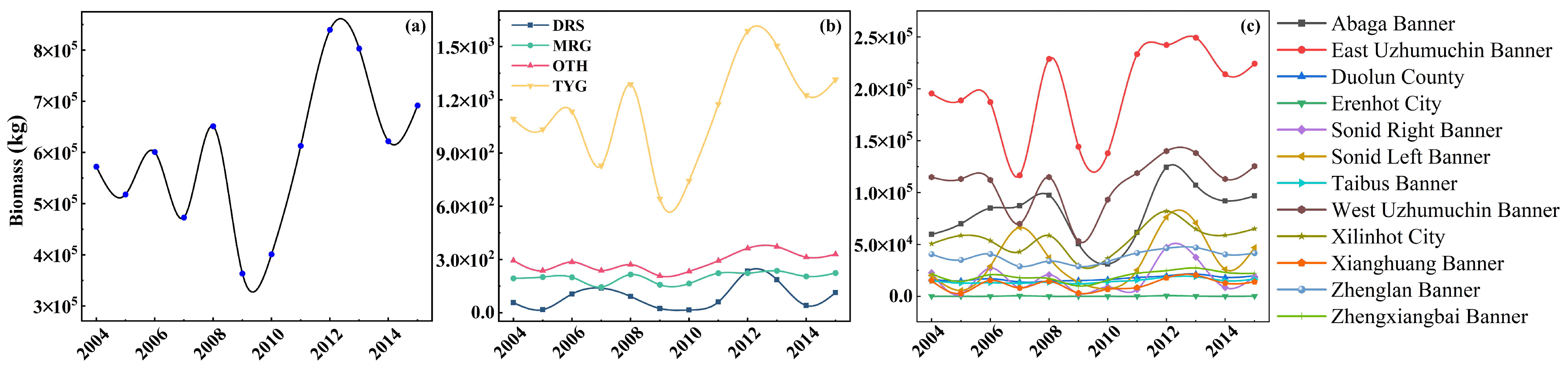
3.2. Temporal and Spatial Variation in Grassland Biomass
3.2.1. Temporal Variation in Grassland Biomass
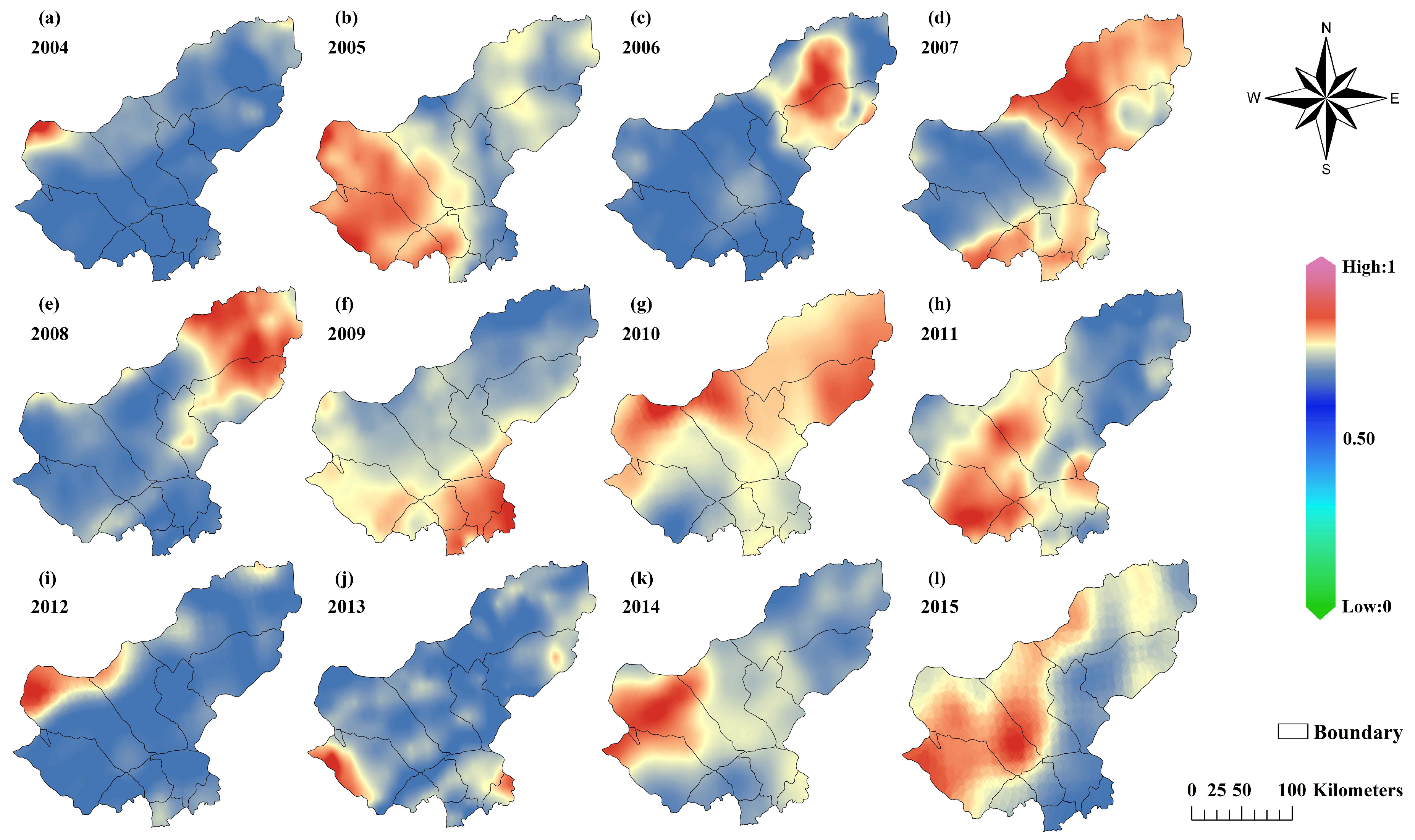
3.2.2. Spatial Variation in Grassland Biomass
3.3. Optimal SPEI Time Scale Selection for Drought Risk Assessment of Grassland Biomass
3.4. Drought Risk Assessment of Different Grassland Types
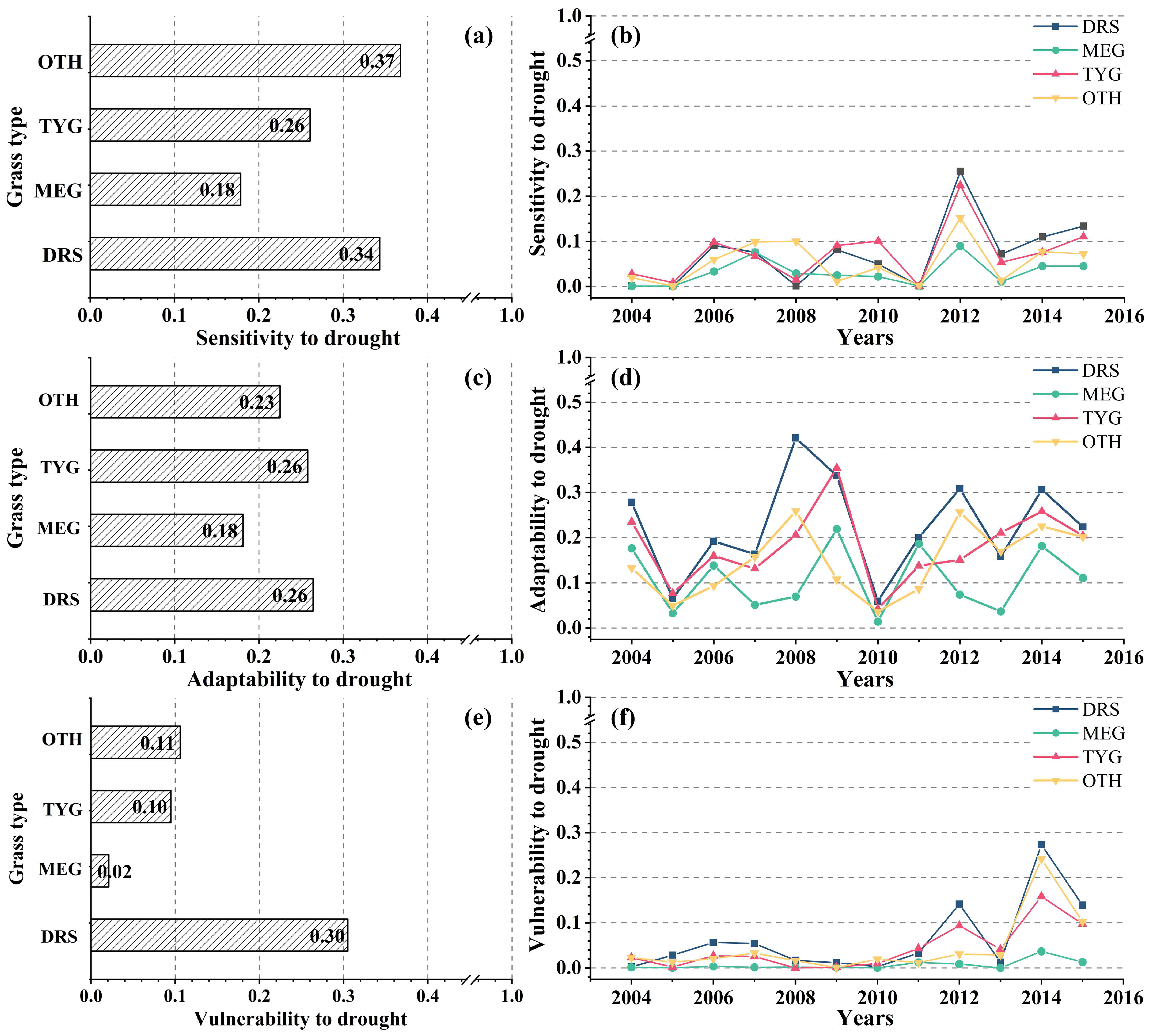
3.4.1. Drought Hazard Assessment of Different Grassland Types
3.4.2. Drought Vulnerability Assessment of the Different Grassland Types
3.4.3. Drought Risk Assessment of Different Grassland Types
3.5. Analysis of Drought Risk Drivers
4. Discussion
4.1. Model Rationalization and Evaluation Results
4.2. Analysis of Grassland Biomass and Drivers of Drought Risk
4.3. Limitations
5. Conclusions
- (1)
- Among the eight selected variables affecting grassland biomass, the NDVI, Prcp, SM and Lon were combined as input variables, and the highest model accuracy for estimating biomass was R = 0.90 and RMSE = 0.09 kg/m2. During the study period, the biomass of typical grassland was the highest, and the biomass of desert grassland was the lowest, showing a spatial distribution that gradually decreases from northeast to southwest.
- (2)
- Drought risk to grassland biomass in the Xilin Gol League shows a fluctuating trend over time. The years 2012 and 2014 had the highest drought risk during the study period. There were differences in drought risk among grassland types, among which the highest drought risk was found in desert grassland (DR = 0.30) and the lowest in meadow grassland (DR = 0.02).
- (3)
- The effects of various factors on drought risk differed depending on the grassland type. Tem mainly drives TYG drought risk, while SM greatly impacts drought risk in MEG. ET has a relatively high contribution to the drought risk of DRS. Drought risks in the other grasslands are more sensitive to Prcp.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, A.; Zhang, A.; Liu, X.; Cao, S. Spatiotemporal changes of normalized difference vegetation index (NDVI) and response to climate extremes and ecological restoration in the Loess Plateau, China. Theor. Appl. Climatol. 2018, 132, 555–567. [Google Scholar]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar]
- He, B.; Liu, J.; Guo, L.; Wu, X.; Xie, X.; Zhang, Y.; Chen, C.; Zhong, Z.; Chen, Z. Recovery of Ecosystem Carbon and Energy Fluxes From the 2003 Drought in Europe and the 2012 Drought in the United States. Geophys. Res. Lett. 2018, 45, 4879–4888. [Google Scholar]
- Huang, L.; He, B.; Chen, A.; Wang, H.; Liu, J.; Lű, A.; Chen, Z. Drought dominates the interannual variability in global terrestrial net primary production by controlling semi-arid ecosystems. Sci. Rep. 2016, 6, 24639. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; He, B.; Han, L.; Liu, J.; Wang, H.; Chen, Z. A global examination of the response of ecosystem water-use efficiency to drought based on MODIS data. Sci. Total Environ. 2017, 601–602, 1097. [Google Scholar]
- He, B.; Lü, A.; Wu, J.; Lin, Z.; Ming, L. Drought hazard assessment and spatial characteristics analysis in China. J. Geogr. Sci. 2011, 21, 235–249. [Google Scholar] [CrossRef]
- Qu, Y.; Zhao, Y.; Ding, G.; Chi, W.; Gao, G. Spatiotemporal patterns of the forage-livestock balance in the Xilin Gol steppe, China: Implications for sustainably utilizing grassland-ecosystem services. J. Arid Land 2021, 13, 135–151. [Google Scholar]
- You, L.Q.; Ping, T.Y. The effects of land use change on the eco-environmental evolution of farming-pastoral region in Northern China:With an emphasis on Duolun County in Inner Mongolia. Acta Ecol. Sin. 2003, 23, 1025–1030. [Google Scholar]
- Lu, D. The potential and challenge of remote sensing-based biomass estimation. Int. J. Remote Sens. 2006, 27, 1297–1328. [Google Scholar]
- Iftikhar, A.; Fiona, C.; Edward, D.; Brian, B.; Stuart, G. Satellite remote sensing of grasslands: From observation to management—A review. J. Plant Ecol. 2016, 9, 649–671. [Google Scholar]
- Prasad, R.; Pandey, A.; Singh, K.P.; Singh, V.P.; Mishra, R.K.; Singh, D. Retrieval of spinach crop parameters by microwave remote sensing with back propagation artificial neural networks: A comparison of different transfer functions. Adv. Space Res. 2012, 50, 363–370. [Google Scholar]
- Wang, D.C.; Wang, J.H.; Jin, N.; Wang, Q.; Huang, F. ANN-based wheat biomass estimation using canopy hyperspectral vegetation indices. Trans. Chin. Soc. Agric. Eng. 2008, 24, 196–201. [Google Scholar]
- Li, Q.; Chen, L.; Xu, Y. Drought risk and water resources assessment in the Beijing-Tianjin-Hebei region, China. Sci. Total Environ. 2022, 832, 154915. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Zhang, J.; Wang, R.; Zhang, M.; Bao, Y.; Guo, E.; Wang, Y. Analysis for Spatio-Temporal Variation Characteristics of Droughts in Different Climatic Regions of the Mongolian Plateau Based on SPEI. Sustainability 2019, 11, 5767. [Google Scholar]
- Hao, Z.; Singh, V.P. Drought characterization from a multivariate perspective: A review. J. Hydrol. 2015, 527, 668–678. [Google Scholar] [CrossRef]
- Mckee, T.B.; Doesken, N.J.; Kleist, J. The Relationship of drought frequency and duration to time scales. In Proceedings of the 8th Conference on Applied Climatology, Anaheim, CA, USA, 17–22 January 1993; pp. 179–183. [Google Scholar]
- Vousdoukas, M.I.; Mentaschi, L.; Voukouvalas, E.; Bianchi, A.; Dottori, F.; Feyen, L. Climatic and socioeconomic controls of future coastal flood risk in Europe. Nat. Clim. Chang. 2018, 8, 776–780. [Google Scholar] [CrossRef]
- Du, L.; Tian, Q.; Yu, T.; Meng, Q.; Jancso, T.; Udvardy, P.; Huang, Y. A comprehensive drought monitoring method integrating MODIS and TRMM data. Int. J. Appl. Earth Obs. Geoinf. 2013, 23, 245–253. [Google Scholar]
- Dai, A. Drought under global warming: A review. Wiley Interdiscip. Rev. Clim. Chang. 2011, 2, 45–65. [Google Scholar]
- Li, K.; Tong, Z.; Liu, X.; Zhang, J.; Tong, S. Quantitative assessment and driving force analysis of vegetation drought risk to climate change:Methodology and application in Northeast China. Agric. For. Meteorol. 2019, 282–283, 107865. [Google Scholar]
- Xu, H.J.; Wang, X.P.; Zhao, C.Y.; Yang, X.M. Diverse responses of vegetation growth to meteorological drought across climate zones and land biomes in northern China from 1981 to 2014. Agric. For. Meteorol. 2018, 262, 1–13. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.I.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [PubMed]
- Wilhite, D.A. Drought as a Natural Hazard: Concepts and Definitions. 2000. Available online: http://digitalcommons.unl.edu/droughtfacpub (accessed on 26 September 2022).
- Wisner, B.; Blaikie, P.; Cannon, T.; Davis, I. At Risk: Natural Hazards, People’s Vulnerability and Disasters; Routledge: London, UK, 2014. [Google Scholar]
- Zhang, Q.; Zhang, J.; Wang, C. Risk assessment of drought disaster in typical area of corn cultivation in China. Theor. Appl. Climatol. 2017, 128, 533–540. [Google Scholar]
- Pachauri, R.; Meyer, L. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. 2014. Available online: https://epic.awi.de/id/eprint/37530/ (accessed on 26 September 2022).
- Yang, X.; Li, Y.P.; Huang, G. A maximum entropy copula-based frequency analysis method for assessing bivariate drought risk: A case study of the Kaidu River Basin. J. Water Clim. Chang. 2022, 13, 175–189. [Google Scholar]
- Zhang, L.; Chu, Q.Q.; Jiang, Y.L.; Fu, C.; Lei, Y.D. Impacts of climate change on drought risk of winter wheat in the North China Plain. J. Integr. Agric. 2021, 20, 2601–2612. [Google Scholar]
- Tong, C.; Wu, J.; Yong, S.-p.; Yang, J.; Yong, W. A landscape-scale assessment of steppe degradation in the Xilin River Basin, Inner Mongolia, China. J. Arid Environ. 2004, 59, 133–149. [Google Scholar] [CrossRef]
- Wang, Z.; Yu, Q.; Guo, L. Quantifying the impact of the grain-for-green program on ecosystem health in the typical agro-pastoral ecotone: A case study in the Xilin Gol league, Inner Mongolia. Int. J. Environ. Res. Public Health 2020, 17, 5631. [Google Scholar] [CrossRef]
- Javed, T.; Li, Y.; Feng, K.; Ayantobo, O.O.; Ahmad, S.; Chen, X.; Rashid, S.; Suon, S. Monitoring responses of vegetation phenology and productivity to extreme climatic conditions using remote sensing across different sub-regions of China. Environ. Sci. Pollut. Res. 2021, 28, 3644–3659. [Google Scholar]
- Jiang, R.; Wang, P.; Xu, Y.; Zhou, Z.; Luo, X.; Lan, Y.; Zhao, G.; Sanchez-Azofeifa, A.; Laakso, K. Assessing the operation parameters of a low-altitude UAV for the collection of NDVI values over a paddy rice field. Remote Sens. 2020, 12, 1850. [Google Scholar] [CrossRef]
- Matsushita, B.; Yang, W.; Chen, J.; Onda, Y.; Qiu, G. Sensitivity of the enhanced vegetation index (EVI) and normalized difference vegetation index (NDVI) to topographic effects: A case study in high-density cypress forest. Sensors 2007, 7, 2636–2651. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, T.; Li, Y.; Li, Y.; Bu, C.; Zhang, Q. Comparative analysis of fractional vegetation cover estimation based on multi-sensor data in a semi-arid sandy area. Chin. Geogr. Sci. 2019, 29, 166–180. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Zhou, X.; Zhu, X.; Dong, Z.; Guo, W. Estimation of biomass in wheat using random forest regression algorithm and remote sensing data. Crop J. 2016, 4, 212–219. [Google Scholar]
- Chan, J.C.-W.; Paelinckx, D. Evaluation of Random Forest and Adaboost tree-based ensemble classification and spectral band selection for ecotope mapping using airborne hyperspectral imagery. Remote Sens. Environ. 2008, 112, 2999–3011. [Google Scholar]
- Gong, H.; Sun, Y.; Shu, X.; Huang, B. Use of random forests regression for predicting IRI of asphalt pavements. Constr. Build. Mater. 2018, 189, 890–897. [Google Scholar] [CrossRef]
- Potopová, V.; Stepanek, P.; Mozny, M.; Soukup, J. Performance of the standardised precipitation evapotranspiration index at various lags for agricultural drought risk assessment in the Czech Republic. Agric. For. Meteorol. 2015, 202, 26–38. [Google Scholar]
- Begueria-Portugues, S.; Vicente-Serrano, S.; Angulo-Martínez, M.; López-Moreno, J.; EIKenawy, A. The Standardized Precipitation-Evapotranspiration Index (SPEI): A multiscalar drought index. In Proceedings of the 10th EMS Annual Meeting, Zürich, Switzerland, 13–17 September 2010; p. EMS2010-2562. [Google Scholar]
- Zhou, Q.; Luo, Y.; Zhou, X.; Cai, M.; Zhao, C. Response of vegetation to water balance conditions at different time scales across the karst area of southwestern China—A remote sensing approach. Sci. Total Environ. 2018, 645, 460–470. [Google Scholar]
- Zhao, A.; Zhang, A.; Cao, S.; Liu, X.; Liu, J.; Cheng, D. Responses of vegetation productivity to multi-scale drought in Loess Plateau, China. Catena 2018, 163, 165–171. [Google Scholar]
- Gao, J.; Jiao, K.; Wu, S. Quantitative assessment of ecosystem vulnerability to climate change: Methodology and application in China. Environ. Res. Lett. 2018, 13, 094016. [Google Scholar] [CrossRef]
- Jin, J.; Wang, Q. Assessing ecological vulnerability in western China based on Time-Integrated NDVI data. J. Arid Land 2016, 8, 533–545. [Google Scholar] [CrossRef]
- Keenan, R.J. Climate change impacts and adaptation in forest management: A review. Ann. For. Sci. 2015, 72, 145–167. [Google Scholar] [CrossRef]
- Kozak, M.; Kang, M. S. Note on modern path analysis in application to crop science. Commun. Biometry Crop Sci. 2006, 1, 32–34. [Google Scholar]
- Gleason, C.J.; Im, J. Forest biomass estimation from airborne LiDAR data using machine learning approaches. Remote Sens. Environ. 2012, 125, 80–91. [Google Scholar]
- Montes, J.M.; Technow, F.; Dhillon, B.S.; Mauch, F.; Melchinger, A.E. High-throughput non-destructive biomass determination during early plant development in maize under field conditions. Field Crops Res. 2011, 121, 268–273. [Google Scholar]
- Jin, X.L.; Diao, W.Y.; Xiao, C.H.; Wang, F.Y.; Chen, B.; Wang, K.R.; Li, S.K.; Ive, D.S. Estimation of Wheat Agronomic Parameters using New Spectral Indices. PLoS ONE 2013, 8, e72736. [Google Scholar] [CrossRef]
- Liu, M.; Liu, G.; Gong, L.; Wang, D.; Sun, J. Relationships of biomass with environmental factors in the grassland area of Hulunbuir, China. PLoS ONE 2014, 9, e102344. [Google Scholar]
- Feng, Q.; Liu, B.; Yang, S.; Liang, T.; Huang, X. Multi-factor modeling of above-ground biomass in alpine grassland: A case study in the Three-River Headwaters Region, China. Remote Sens. Environ. Interdiscip. J. 2016, 186, 164–172. [Google Scholar]
- Meng, B.; Ge, J.; Liang, T.; Yang, S.; Gao, J.; Feng, Q.; Cui, X.; Huang, X.; Xie, H. Evaluation of remote sensing inversion error for the above-ground biomass of alpine meadow grassland based on multi-source satellite data. Remote Sens. 2017, 9, 372. [Google Scholar]
- Cabrera-Bosquet, L.; Molero, G.; Stellacci, A.; Bort, J.; Nogués, S.; Araus, J. NDVI as a potential tool for predicting biomass, plant nitrogen content and growth in wheat genotypes subjected to different water and nitrogen conditions. Cereal Res. Commun. 2011, 39, 147–159. [Google Scholar]
- Xu, B.; Yang, X.; Tao, W.; Qin, Z.; Liu, H.; Miao, J. Remote sensing monitoring upon the grass production in China. Acta Ecol. Sin. 2007, 2, 405–413. [Google Scholar] [CrossRef]
- Chi, D.; Wang, H.; Li, X.; Liu, H.; Li, X. Assessing the effects of grazing on variations of vegetation NPP in the Xilingol Grassland, China, using a grazing pressure index. Ecol. Indic. 2018, 88, 372–383. [Google Scholar]
- Yang, S.; Feng, Q.; Liang, T.; Liu, B.; Zhang, W. Modeling grassland above-ground biomass based on artificial neural network and remote sensing in the Three-River Headwaters Region. Remote Sens. Environ. Interdiscip. J. 2018, 204, 448–455. [Google Scholar]
- Guo, E.; Wang, Y.; Wang, C.; Sun, Z.; Li, H. NDVI Indicates Long-Term Dynamics of Vegetation and Its Driving Forces from Climatic and Anthropogenic Factors in Mongolian Plateau. Remote Sens. 2021, 13, 688. [Google Scholar]
- Li, X.; Philp, J.; Cremades, R.; Roberts, A.; He, L.; Li, L.; Yu, Q. Agricultural vulnerability over the Chinese Loess Plateau in response to climate change: Exposure, sensitivity, and adaptive capacity. Ambio 2016, 45, 350–360. [Google Scholar] [PubMed]
- Mu, S.; Zhou, S.; Chen, Y.; Li, J.; Ju, W.; Odeh, I. Assessing the impact of restoration-induced land conversion and management alternatives on net primary productivity in Inner Mongolian grassland, China. Glob. Planet. Chang. 2013, 108, 29–41. [Google Scholar] [CrossRef]
- Ersi, C.; Bayaer, T.; Bao, Y.; Bao, Y.; Yong, M.; Zhang, X. Temporal and Spatial Changes in Evapotranspiration and Its Potential Driving Factors in Mongolia over the Past 20 Years. Remote Sens. 2022, 14, 1856. [Google Scholar]
- Diffenbaugh, N.S.; Swain, D.L.; Touma, D. Anthropogenic warming has increased drought risk in California. Proc. Natl. Acad. Sci. USA 2015, 112, 3931–3936. [Google Scholar]
- Zhu, X.-J.; Yu, G.-R.; Hu, Z.-M.; Wang, Q.-F.; He, H.-L.; Yan, J.-H.; Wang, H.-M.; Zhang, J.-H. Spatiotemporal variations of T/ET (the ratio of transpiration to evapotranspiration) in three forests of Eastern China. Ecol. Indic. 2015, 52, 411–421. [Google Scholar] [CrossRef]
- Chen, J.; Li, Z.; Jia, S. Temporal and spatial changes of climate aridity in Xilinguole steppe region. J. Inn. Mong. Univ. (Nat. Sci. Ed.) 2011, 42, 304–310. [Google Scholar]
- Gouveia, C.M.; Trigo, R.M.; Begueria, S.; Vicente-Serrano, S.M. Drought impacts on vegetation activity in the Mediterranean region: An assessment using remote sensing data and multi-scale drought indicators. Glob. Planet. Chang. 2016, 151, 15–27. [Google Scholar] [CrossRef]
- Payero, J.O.; Tarkalson, D.D.; Irmak, S.; Davison, D.; Petersen, J.L. Effect of timing of a deficit-irrigation allocation on corn evapotranspiration, yield, water use efficiency and dry mass. Agric. Water Manag. 2009, 96, 1387–1397. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Abbreviation | Formula | Reference |
| Normalized Difference Vegetation Index | NDVI | [32] | |
| Enhanced Vegetation Index | EVI | [33] | |
| Fractional Vegetation Cover | FVC | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bu, L.; Lai, Q.; Qing, S.; Bao, Y.; Liu, X.; Na, Q.; Li, Y. Grassland Biomass Inversion Based on a Random Forest Algorithm and Drought Risk Assessment. Remote Sens. 2022, 14, 5745. https://doi.org/10.3390/rs14225745
Bu L, Lai Q, Qing S, Bao Y, Liu X, Na Q, Li Y. Grassland Biomass Inversion Based on a Random Forest Algorithm and Drought Risk Assessment. Remote Sensing. 2022; 14(22):5745. https://doi.org/10.3390/rs14225745
Chicago/Turabian StyleBu, Lingxin, Quan Lai, Song Qing, Yuhai Bao, Xinyi Liu, Qin Na, and Yuan Li. 2022. "Grassland Biomass Inversion Based on a Random Forest Algorithm and Drought Risk Assessment" Remote Sensing 14, no. 22: 5745. https://doi.org/10.3390/rs14225745
APA StyleBu, L., Lai, Q., Qing, S., Bao, Y., Liu, X., Na, Q., & Li, Y. (2022). Grassland Biomass Inversion Based on a Random Forest Algorithm and Drought Risk Assessment. Remote Sensing, 14(22), 5745. https://doi.org/10.3390/rs14225745