
Wavelet Analysis Reveals Phenology Mismatch between Leaf Phenology of Temperate Forest Plants and the Siberian Roe Deer Molting under Global Warming


 , and
, and
Abstract
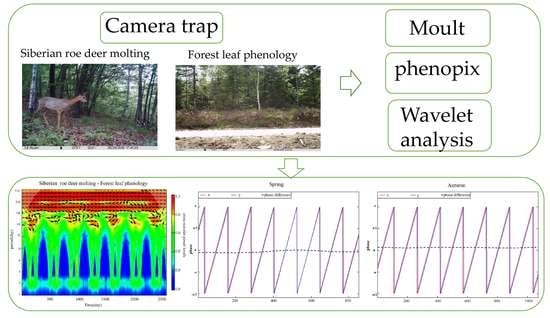
:
1. Introduction
2. Materials and Methods
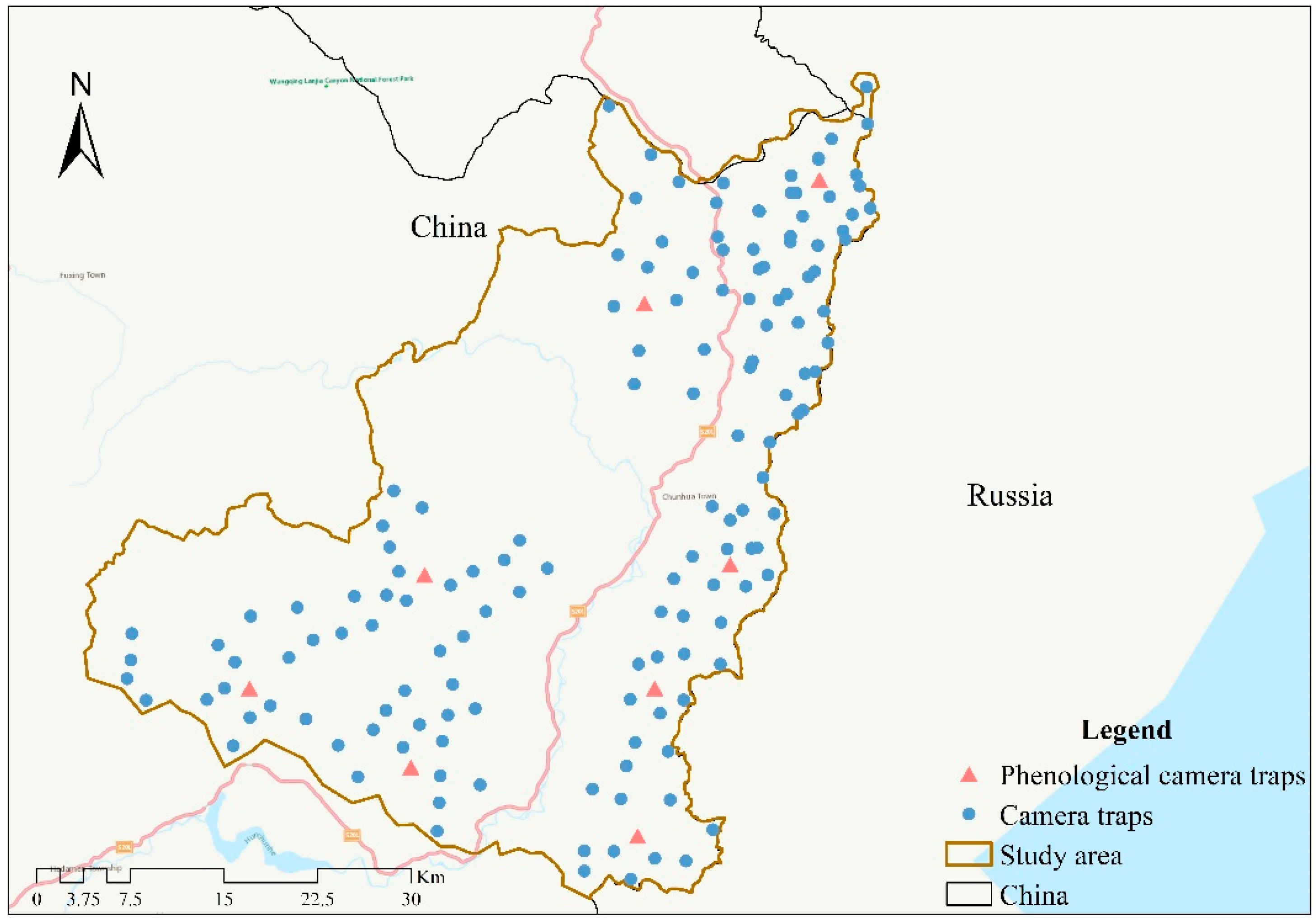
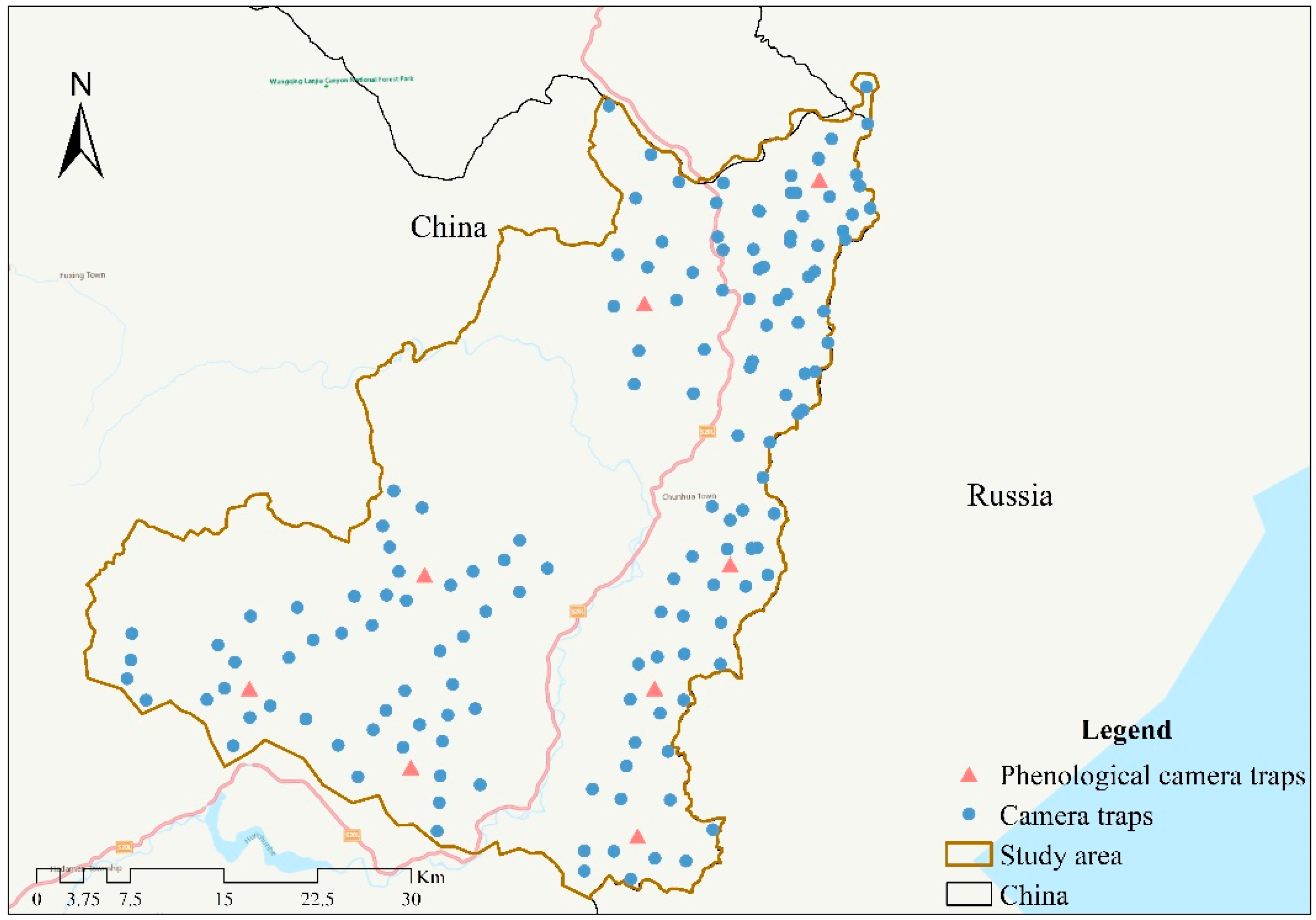
2.1. Data Collection
2.2. Forest Leaf Phenology Analysis
2.3. Measurement and Determination of Siberian Roe Deer Molting
2.4. Trend Analysis
2.5. Relationship between Meteorological Factors and Forest Leaf Phenology
2.6. Relationship between Meteorological Factors and Siberian Roe Deer Molting
2.7. Synchronization Analysis of Forest Leaf Phenology and Siberian Roe Deer Molting
3. Results
3.1. Forest Leaf Phenology Monitor
3.1.1. Key Phenophases of Forest Leaf Phenology
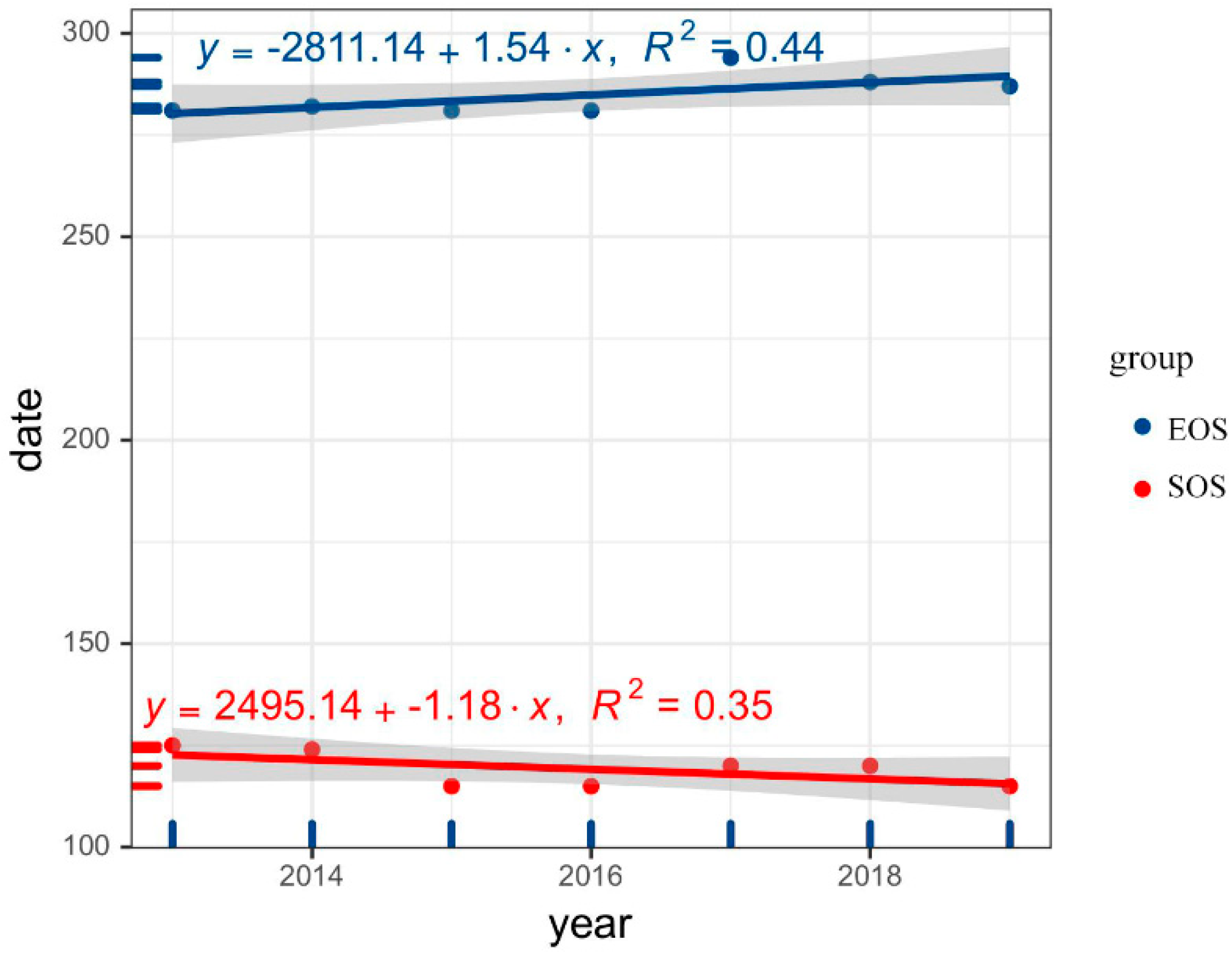
3.1.2. Interannual Variation Trend of Forest Leaf Phenology 2013–2019
3.2. Camera-Trap Monitoring of Siberian Roe Deer Molting
3.2.1. Start Date and Duration of Siberian Roe Deer Molting
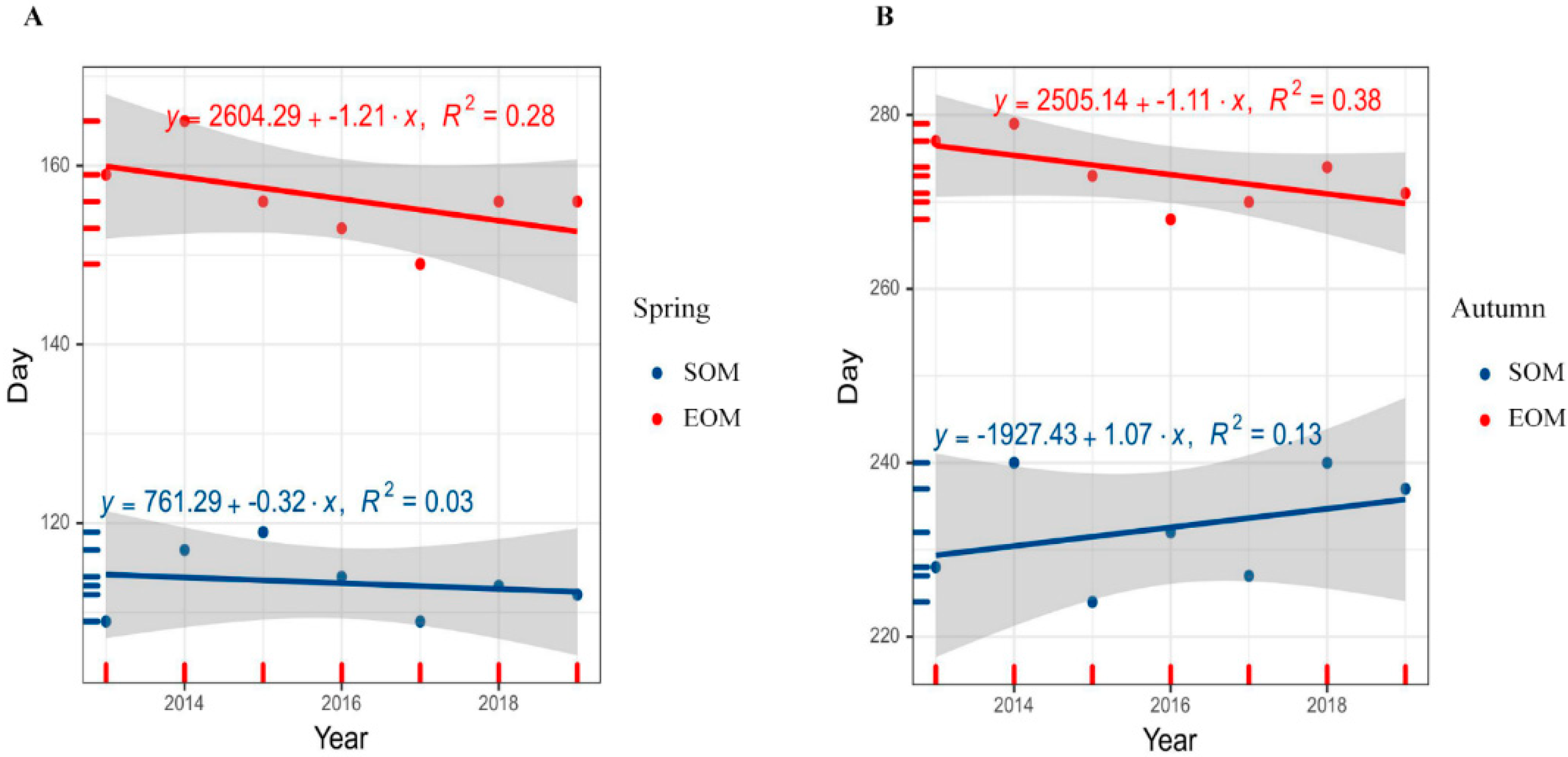
3.2.2. Interannual Variation Trend of Siberian Roe Deer Molting from 2013 to 2019
3.3. Relationship between Meteorological Factors and Forest Leaf Phenology
3.4. Relationship between Meteorological Factors and Siberian Roe Deer Molting
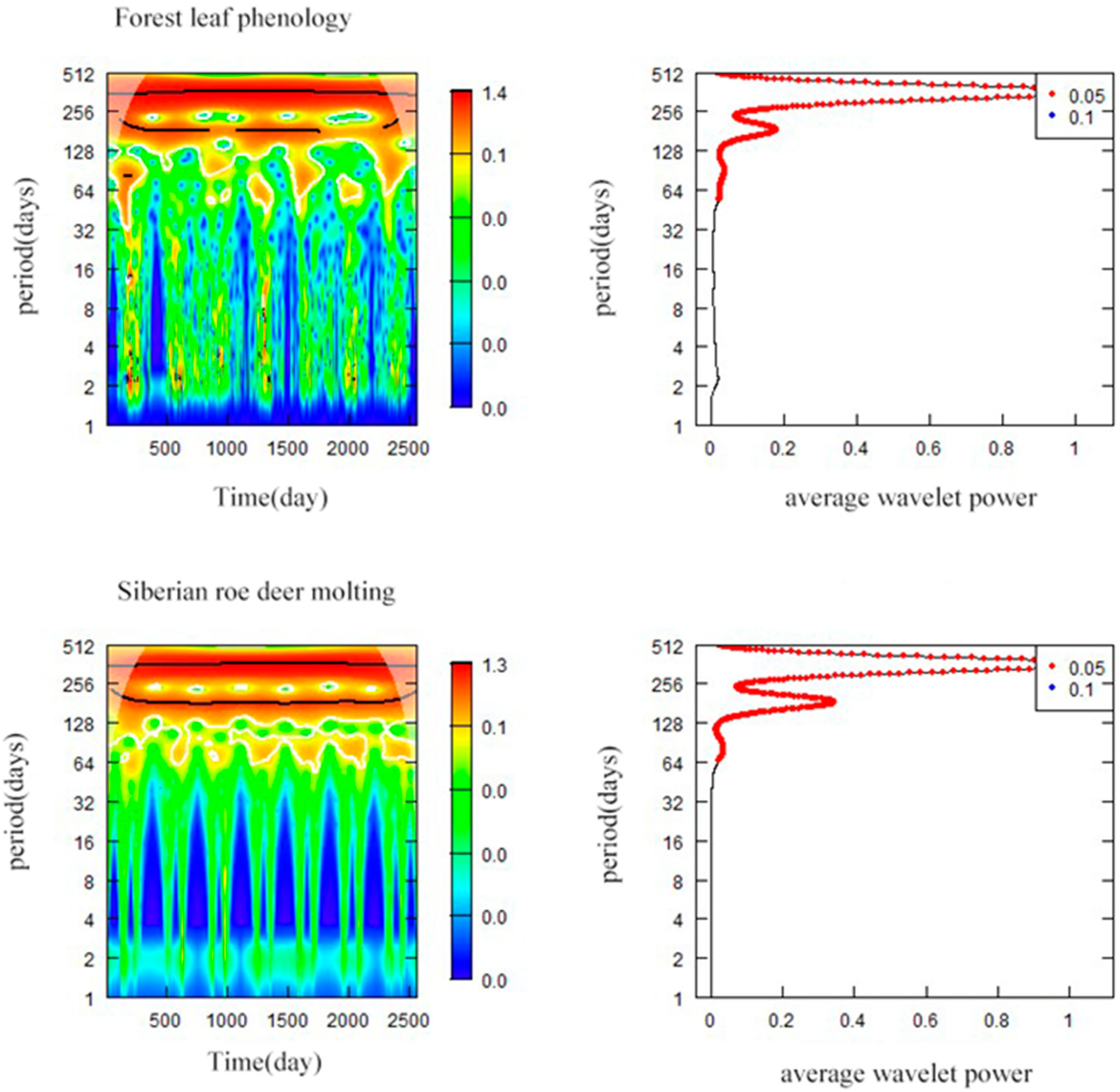
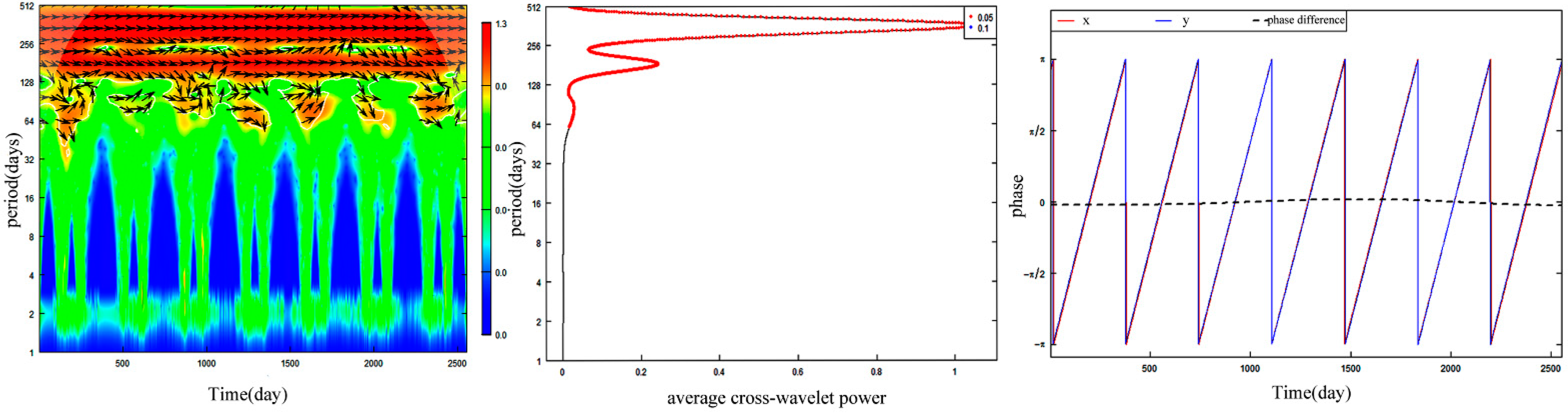
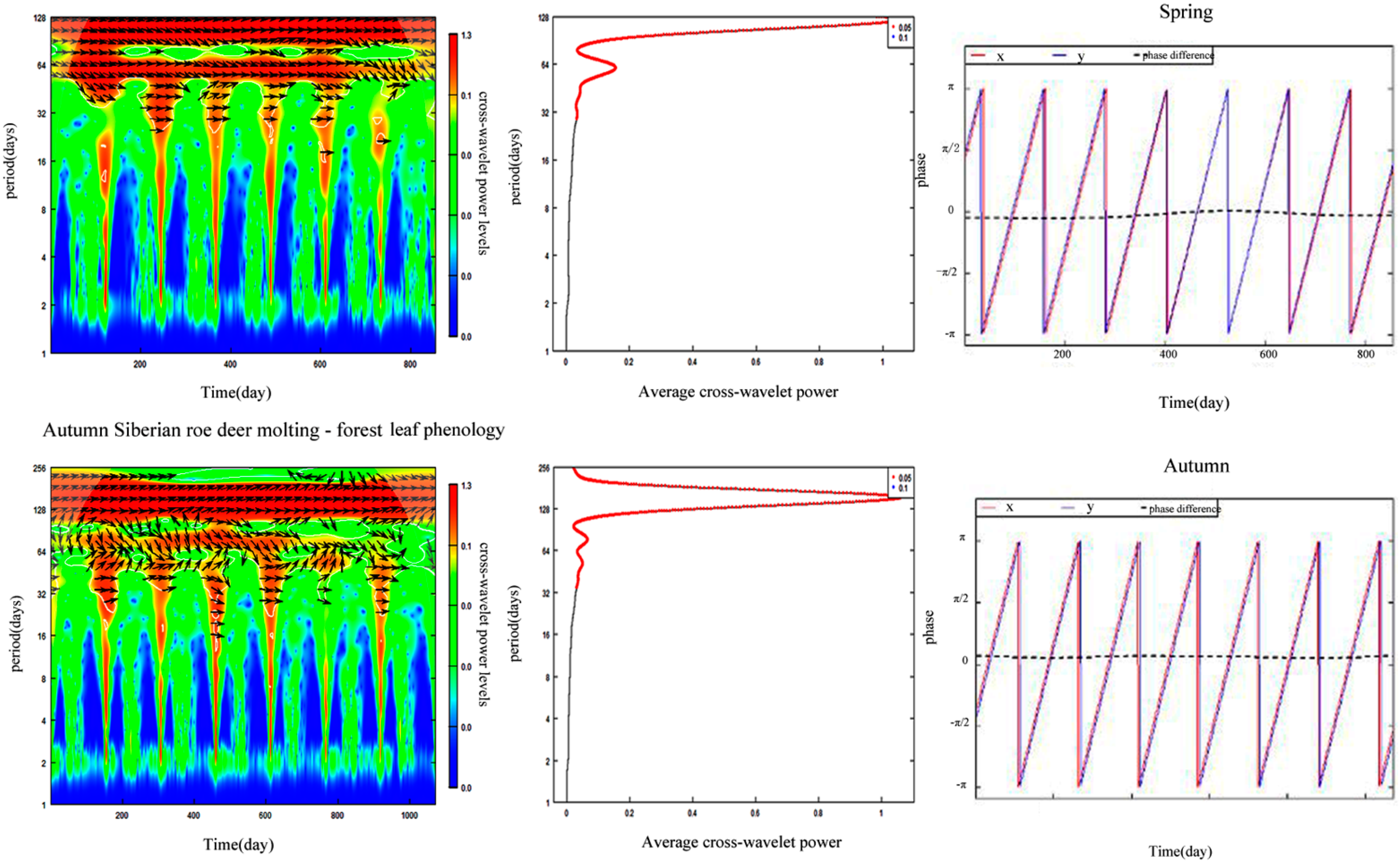
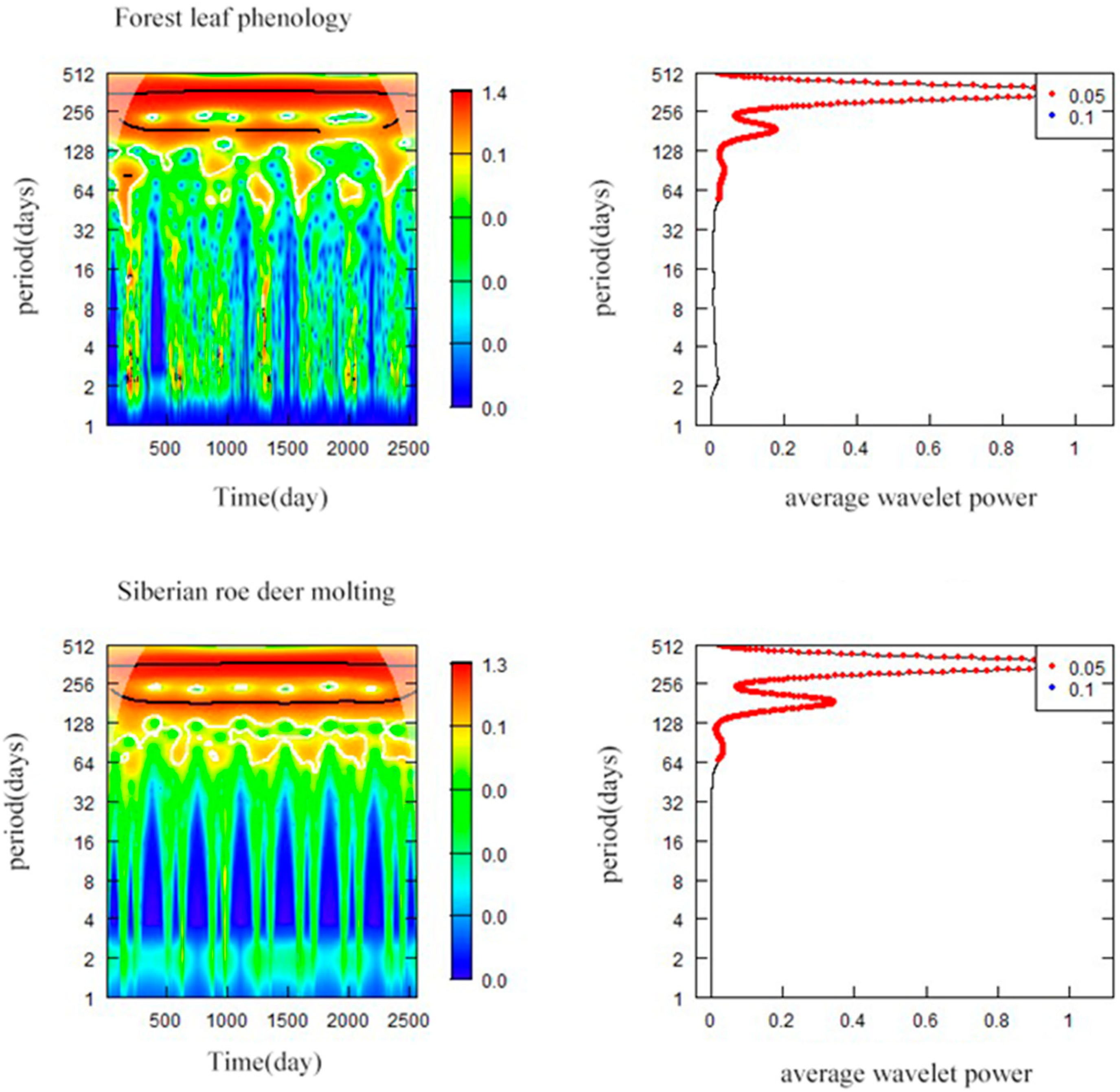
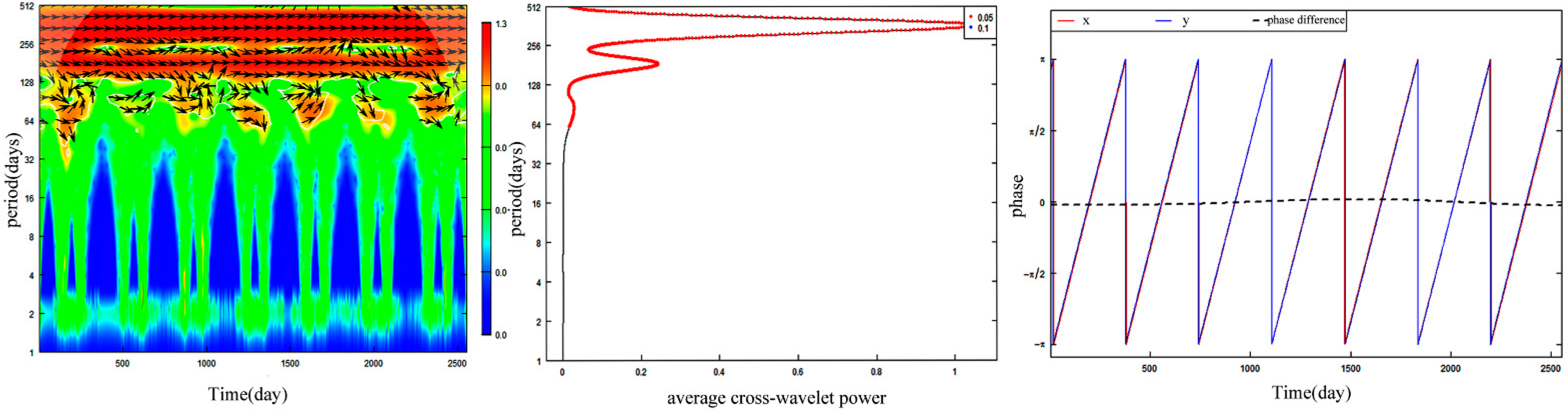
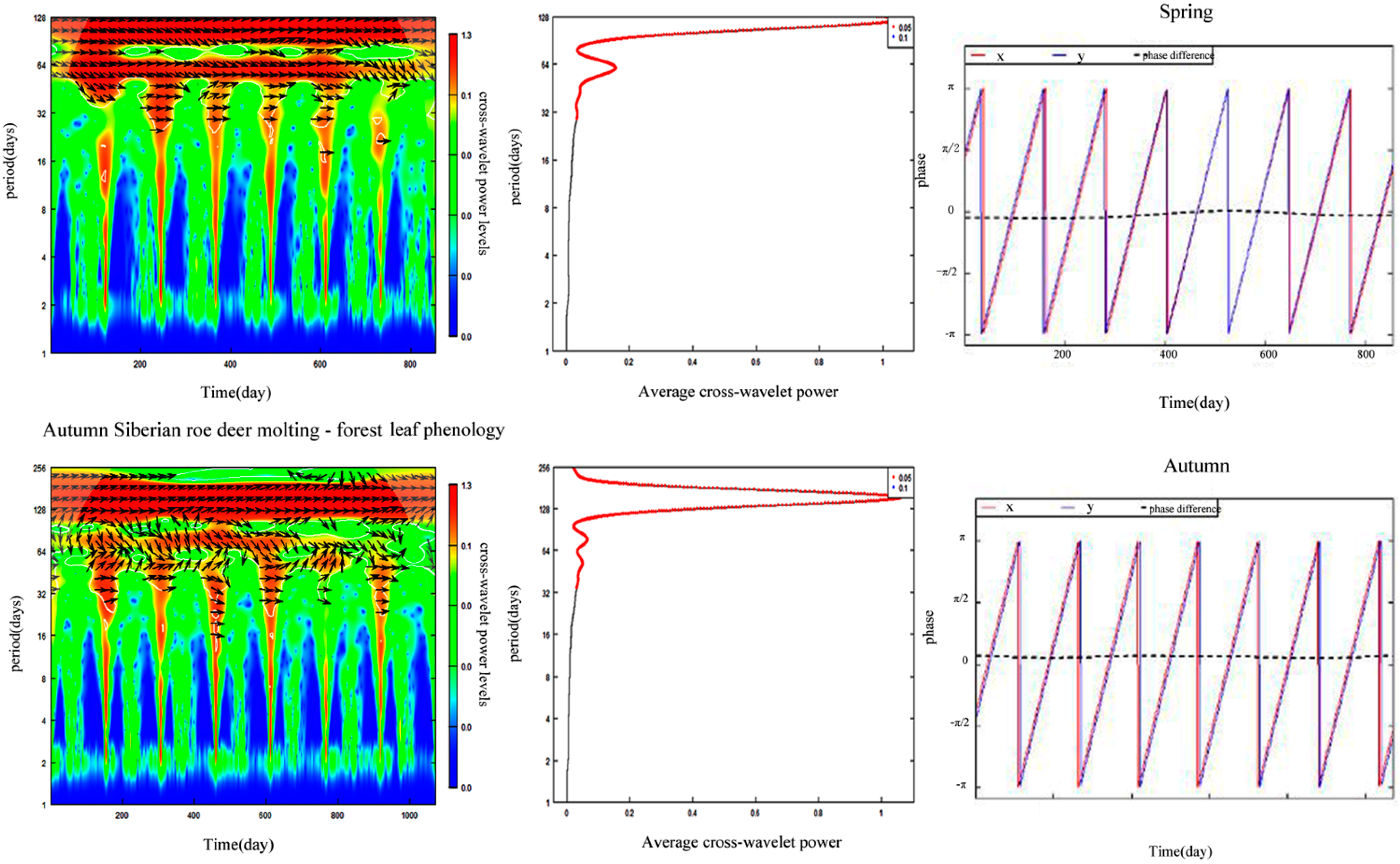
3.5. Synchronization between Forest Leaf Phenology and Siberian Roe Deer Molting
4. Discussion
4.1. Effects of Climate Change on Forest Leaf Phenology
4.2. Effects of Climate Change on Siberian Roe Deer Molting
4.3. Effects of Climate Change on Phenological Synchrony of Plants and Animals
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penuelas, J.; Rutishauser, T.; Filella, I. Phenology feedbacks on climate change. Science 2009, 324, 887–888. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.R.; Townsend, P.A.; Mladenoff, D.J. Mapping asynchrony between gypsy moth egg-hatch and forest leaf-out: Putting the phenological window hypothesis in a spatial context. For. Ecol. Manag. 2013, 287, 67–76. [Google Scholar] [CrossRef]
- Lai, J.Y. Seasonal Hair Change of Siberian Roe Deer (Capreolus pygargus) Based on Automatic Camera Technology; Northeast Forestry University: Harbin, China, 2020. [Google Scholar]
- Hua, Y.; Zhang, W.; Xu, Y.C. Seasonal variation of pelage characteristics in Siberian weasel (Mustela sibirica) of Xiaoxing’ anling area, Heilongjiang, China. Acta Theriol. Sin. 2010, 30, 110–114. [Google Scholar]
- Zimova, M.; Mills, L.S.; Lukacs, P.M.; Mitchell, M.S. Snowshoe hares display limited phenotypic plasticity to mismatch in seasonal camouflage. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140029. [Google Scholar] [CrossRef]
- Mills, L.S.; Zimova, M.; Oyler, J.; Running, S.; Abatzoglou, J.T.; Lukacs, P.M. Camouflage mismatch in seasonal coat color due to decreased snow duration. Proc. Natl. Acad. Sci. USA 2013, 110, 7360–7365. [Google Scholar] [CrossRef]
- Mills, L.S.; Bragina, E.V.; Kumar, A.V.; Zimova, M.; Lafferty, D.J.R.; Feltner, J.; Davis, B.M.; Hackländer, K.; Alves, P.C.; Good, J.M.; et al. Winter color polymorphisms identify global hot spots for evolutionary rescue from climate change. Science 2018, 359, 1033–1036. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Forrest, J.R. Complex responses of insect phenology to climate change. Curr. Opin. Insect Sci. 2016, 17, 49–54. [Google Scholar] [CrossRef]
- Charmantier, A.; McCleery, R.H.; Cole, L.R.; Perrins, C.; Kruuk, L.E.B.; Sheldon, B.C. Adaptive Phenotypic Plasticity in Response to Climate Change in a Wild Bird Population. Science 2008, 320, 800–803. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, I.R.; Bryant, D.M. Avian phenology: Climate change and constraints on breeding. Nature 2000, 406, 366–367. [Google Scholar] [CrossRef]
- Roxanne, S.B.; Jennifer, M.B.; Greg, A.B. Convergence of biannual moulting strategies across birds and mammals. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180318. [Google Scholar]
- Mundim, F.M.; Bruna, E.M. Is There a Temperate Bias in Our Understanding of How Climate Change Will Alter Plant-Herbivore Interactions? A Meta-analysis of Experimental Studies. Am. Nat. 2016, 188, S74–S89. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.D.; Jenkins, J.P.; Braswell, B.H.; Hollinger, D.Y.; Ollinger, S.V.; Smith, M.-L. Use of digital webcam images to track spring green-up in a deciduous broadleaf forest. Oecologia 2007, 152, 323–334. [Google Scholar] [CrossRef]
- Burns, P.A.; Parrott, M.L.; Rowe, K.C.; Phillips, B.L. Identification of threatened rodent species using infrared and white-flash camera traps. Aust. Mammal. 2018, 40, 188–197. [Google Scholar] [CrossRef]
- Nowak, K.; Berger, J.; Panikowski, A.; Reid, D.G.; Jacob, A.L.; Newman, G.; Young, N.E.; Beckmann, J.P.; Richards, S.A. Using community photography to investigate phenology: A case study of coat molt in the mountain goat (Oreamnos americanus) with missing data. Ecol. Evol. 2020, 10, 13488–13499. [Google Scholar] [CrossRef]
- Filippa, G.; Cremonese, E.; Migliavacca, M.; Galvagno, M.; Forkel, M.; Wingate, L.; Tomelleri, E.; di Cella, U.M.; Richardson, A.D. Phenopix: A R package for image-based vegetation phenology. Agricutural Meteorol. 2016, 220, 141–150. [Google Scholar] [CrossRef]
- Erni, B.; Bonnevie, B.T.; Underhill, L. Moult: An R package to analyses moult in birds. J. Stat. Softw. 2012, 52, 1–23. [Google Scholar]
- Underhill, L.G.; Zucchini, W.; Summers, R.W. A model for avian primary moult-data types based on migration strategies and an example using the Redshank Tringatotanus. IBIS 1990, 130, 358–372. [Google Scholar] [CrossRef]
- Dai, J.; Wang, H.; Ge, Q. The spatial pattern of leaf phenology and its response to climate change in China. Int. J. Biometeorol. 2014, 58, 521–528. [Google Scholar] [CrossRef]
- Cazelles, B.; Chavez, M.; Berteaux, D.; Ménard, F.; Vik, J.O.; Jenouvrier, S.; Stenseth, N.C. Wavelet analysis of ecological time series. Oecologia 2008, 156, 287–304. [Google Scholar] [CrossRef]
- Spies, B.T.A. Characterizing canopy gap structure in forests using wavelet analysis. J. Ecol. 1992, 80, 205–215. [Google Scholar] [CrossRef]
- Grenfell, B.T.; Rnstad, O.N.; Kappey, J. Travelling waves and spatial hierarchies in measles epidemics. Nature 2001, 414, 716–723. [Google Scholar] [CrossRef]
- Qi, J.Z.; Holyoak, M.; Dobbins, M.T.; Huang, C.; Li, Q.; She, W.; Ning, Y.; Sun, Q.; Jiang, G.; Wang, X. Wavelet methods reveal big cat activity patterns and synchrony of activity with prey. Integr. Zool. 2021, 17, 1–15. [Google Scholar] [CrossRef]
- Roschand, A.; Schmidbauer, H. WaveletComp: A Guide Tour through the R-Package. Online Version Paper, 2014. Available online: https://cran.r-project.org/package=WaveletComp (accessed on 9 May 2020).
- Li, M. The Responses of Forest Phenology on Climate Change in Changbai Mountains. Ph.D. Thesis, Northeast Normal University, Changchun, China, 2011. [Google Scholar]
- Zhou, Y.K. Comparative study of vegetation phenology extraction methods based on digital images. Prog. Geogr. 2018, 37, 23–36. [Google Scholar]
- Bothmann, L.; Menzel, A.; Menze, B.H.; Schunk, C.; Kauermann, G. Automated processing of webcam images for phenological classification. PLoS ONE 2017, 12, e0171918. [Google Scholar] [CrossRef]
- Carter, S.K.; Rudolf, H.W. Shifts in phenological mean and synchrony interact to shape competitive outcomes. Ecology 2019, 100, e02826. [Google Scholar] [CrossRef]
- De Sassi, C.; Tylianakis, J.M. Climate change disproportionately increases herbivore over plant or parasitoid biomass. PLoS ONE 2012, 7, e40557. [Google Scholar] [CrossRef]
- Bahner, K.W.; Zweig, K.A.; Leal, I.R.; Wirth, R. Robustness of plant-insect herbivore interaction networks to climate change in a fragmented temperate forest landscape. Bull. Entomol. Res. 2017, 107, 563–572. [Google Scholar] [CrossRef]
- De Frenne, P.; Van Langenhove, L.; Van Driessche, A.; Bertrand, C.; Verheyen, K.; Vangansbeke, P. Using archived television video footage to quantify phenology responses to climate change. Methods Ecol. Evol. 2018, 9, 1874–1882. [Google Scholar] [CrossRef]
- Filippa, G.; Cremonese, E.; Galvagno, M.; Migliavacca, M.; Morra di Cella, U.; Petey, M.; Siniscalco, C. Five years of phenological monitoring in a mountain grassland: Inter-annual patterns and evaluation of the sampling protocol. Int. J. Biometeorol. 2015, 59, 1927–1937. [Google Scholar] [CrossRef]
- Reyer, C.P.; Leuzinger, S.; Rammig, A.; Wolf, A.; Bartholomeus, R.; Bonfante, A.; De Lorenzi, F.; Dury, M.; Gloning, P.; Jaoudé, R.A.; et al. A plant’s perspective of extremes: Terrestrial plant responses to changing climatic variability. Glob. Chang. Biol. 2012, 19, 75–89. [Google Scholar] [CrossRef]
- Chen, X.Q.; Li, J. Relationships between Leymus chinensis phenology and meteorological factors in Inner Mongolia grasslands. Acta Ecol. Sin. 2009, 29, 5280–5290. [Google Scholar]
- Deng, L.J. Response of Plant Phenology to Climate Change in North China and Surveillance Camera-Based Monitoring of Plant Flowering Phenology. Master’s Thesis, China University of Geosciences, Beijing, China, 2017. [Google Scholar]
- Bai, J.; Ge, Q.S.; Dai, J.H. The response of first flowering dates to abrupt climate change in Beijing. Adv. Atmos. Sci. 2011, 28, 564–572. [Google Scholar] [CrossRef]
- Fu, Y.H.; Zhao, H.; Piao, S.; Peaucelle, M.; Peng, S.; Zhou, G.; Ciais, P.; Huang, M.; Menzel, A.; Peñuelas, J.; et al. Declining global warming effects on the phenology of spring leaf unfolding. Nature 2015, 526, 104–107. [Google Scholar] [CrossRef]
- Gu, L.Y.; Hua, Y.; Mo, J.X.; Wang, H.; Zhang, W. Study on the molt sequence of captive sable in spring in Dalian China. Chin. J. Wildl. 2016, 37, 258–265. [Google Scholar]
- Yang, G.Z. The regularity of seasonal molting of Mustelasibirica. J. Econ. Anim. 1980, 2, 6–8. [Google Scholar]
- Xu, Z.Q.; Xu, L.; Li, B.; Xu, H. Coat changes and hair morphology of musk deer (Moschus berezovskii) fawn. Acta Theriol. Sin. 2006, 26, 101–104. [Google Scholar]
- Hua, Y.; Lai, J.Y.; Liu, D.; Xing, X.; Jiang, G. Measurement method of molt sequence of Siberian roe deer (Capreolus Pygargus) in northeast China based on camera-trapping. Chin. J. Wildl. 2019, 40, 840–847. [Google Scholar]
- Zhang, D. Study on the Environmental Factors Driving the Seasonal Molting Characteristics of Siberian Roe Deer (Capreolus Pygargus). Master’s Thesis, Northeast Forestry University, Harbin, China, 2020. [Google Scholar]
- Berteaux, D.; Gauthier, G.; Domine, F.; Ims, R.A.; Lamoureux, S.F.; Lévesque, E.; Yoccoz, N. Effects of changing permafrost and snow conditions on tundra wildlife: Critical places and times. Arct. Sci. 2016, 3, 65–90. [Google Scholar] [CrossRef]
- Al-Khateeb, A.; Johnson, E. Seasonal changes of pelage in the vole (Microtus agrestis): The effect of daylength. Gen. Comp. Endocrinol. 1971, 16, 229–235. [Google Scholar] [CrossRef]
- Bissonnette, T.H.; Wilson, E. Shortening daylight periods between May 15 and September 12 and the pelt cycle of the mink. Science 1939, 89, 418–419. [Google Scholar] [CrossRef]
- Song, J.H.; Tong, Y.R.; Xiao, Y.J. Effects of light on reproduction and molting of Matres zibellina. Chin. J. Ecol. 1988, 6, 27–29. [Google Scholar]
- Martinet, L.; Allain, D.; Meunier, M. Regulation in pregnant mink (Mustela vison) of plasma progesterone and prolactin and regulation of the onset of the spring moult by daylight ratio and melatonin injections. Can. J. Zool. 1983, 61, 1959–1963. [Google Scholar] [CrossRef]
- Cormont, A.; Wamelink, G.W.; Jochem, R.; WallisDeVries, M.F.; Wegman, R.M. Host plant-mediated effects of climate change on the occurrence of the Alcon blue butterfly (Phengaris alcon). Ecol. Model. 2013, 250, 329–337. [Google Scholar] [CrossRef]
- Van Asch, M.; Van Tienderen, P.H.; Holleman, L.J.M.; Visser, M.E. Predicting adaptation of phenology in response to climate change, an insect herbivore example. Glob. Chang. Biol. 2007, 13, 1596–1604. [Google Scholar] [CrossRef]
- Anderson, J.T.; Inouye, D.W.; Mckinney, A.M.; Colautti, R.I.; Mitchell-Olds, T. Phenotypic plasticity and adaptive evolution contribute to advancing flowering phenology in response to climate change. Proc. R. Soc. B-Biol. Sci. 2012, 279, 3843–3852. [Google Scholar] [CrossRef]
- Lincoln, G.A.; Ebling, F.J.P. Effect of constant-release implants of melatonin on seasonal cycles in reproduction, prolactin secretion and molting in rams. J. Reprod. Fertil. 1985, 73, 241–253. [Google Scholar] [CrossRef]
- Mo, G.; Gili, C.; Ferrando, P. Do photoperiod and temperature influence the molt cycle of Phoca vitulina. Mar. Mammal Sci. 2006, 16, 570–577. [Google Scholar] [CrossRef]
- Murray, D.M. A field study of coat shedding in cattle under conditions of equal day-length but different temperatures. J. Agric. Sci. 1965, 65, 295–300. [Google Scholar] [CrossRef]
- Zimova, M.; Hacklander, K.; Good, J.M.; Melo-Ferreira, J.; Alves, P.C.; Mills, S.L. Function and underlying mechanisms of seasonal colourmolting in mammals and birds: What keeps them changing in a warming world? Biol. Rev. 2018, 93, 1478–1498. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Year | Start of Growing Season | End of Growing Season | Length of Growing Season | Peak of Season Position |
|---|---|---|---|---|---|
| 2013 | 125 | 281 | 156 | 158 | |
| 2014 | 124 | 282 | 158 | 156 | |
| 2015 | 115 | 280 | 165 | 150 | |
| Klosterman | 2016 | 116 | 281 | 166 | 151 |
| 2017 | 119 | 294 | 174 | 155 | |
| 2018 | 120 | 288 | 168 | 151 | |
| 2019 | 114 | 287 | 172 | 148 |
| Season | Year | Duration ± SE (Day) | Start Date ± SE (Day) | SD of Start Date ± SE (Day) |
|---|---|---|---|---|
| Spring | 2013 | 50.12 ± 4.25 | 108.89 ± 2.56 | 5.11 ± 1.85 |
| 2014 | 52.26 ± 1.51 | 116.82 ± 0.65 | 1.78 ± 0.58 | |
| 2015 | 39.60 ± 2.16 | 119.76 ± 1.64 | 3.38 ± 1.11 | |
| 2016 | 39.03 ± 1.87 | 114.02 ± 1.06 | 3.97 ± 1.41 | |
| 2017 | 39.55 ± 1.21 | 109.73 ± 0.69 | 4.11 ± 1.06 | |
| 2018 | 43.28 ± 1.91 | 112.93 ± 0.74 | 3.73 ± 1.13 | |
| 2019 | 44.02 ± 1.69 | 111.74 ± 0.97 | 3.51 ± 1.14 | |
| Autumn | 2013 | 49.25 ± 2.72 | 228.72 ± 1.46 | 4.23 ± 1.70 |
| 2014 | 40.20 ± 1.38 | 234.60 ± 0.93 | 2.32 ± 1.06 | |
| 2015 | 48.99 ± 1.03 | 224.01 ± 0.87 | 3.17 ± 0.95 | |
| 2016 | 36.33 ± 1.25 | 232.18 ± 0.77 | 2.21 ± 0.75 | |
| 2017 | 43.59 ± 1.93 | 227.37 ± 0.96 | 3.28 ± 1.17 | |
| 2018 | 33.70 ± 2.43 | 240.61 ± 1.30 | 3.89 ± 1.40 | |
| 2019 | 33.11 ± 1.69 | 237.96 ± 0.79 | 3.03 ± 1.09 |
| Forest Leaf Phenology | Temperature (R2/p Value) | Sunshine Duration (R2/p Value) | Precipitation (R2/p Value) | Multiple Regression (R2/p Value) |
|---|---|---|---|---|
| Start of growing season | 0.889/<0.05 | 0.083/>0.05 | 0.197/>0.05 | 0.976/<0.05 |
| End of growing season | 0.792/<0.05 | 0.130/>0.05 | 0.807/<0.05 | 0.882/<0.05 |
| Season | Meteorological Factors | Start Date of Molting (r/p Value) | End Date of Molting (r/p Value) |
|---|---|---|---|
| Spring | Mean temperature from winter solstice to start date of molting/end date of molting | 0.206/>0.05 | −0.662/>0.05 |
| Mean sunshine duration from winter solstice to start date of molting/end date of molting | −0.739/<0.05 | 0.729/<0.05 | |
| Sunshine duration of start date of molting/end date of molting | −0.409/>0.05 | 0.714/>0.05 | |
| Mean temperature of start date of molting/end date of molting | −0.377/>0.05 | −0.559/>0.05 | |
| Mean temperature in March | 0.243/>0.05 | 0.139/>0.05 | |
| Mean temperature in April | −0.203/>0.05 | −0.496/>0.05 | |
| Mean temperature in May | 0.359/>0.05 | −0.201/>0.05 | |
| Sunshine duration in March | −0.291/>0.05 | −0.474/>0.05 | |
| Sunshine duration in April | −0.257/<0.05 | 0.167/>0.05 | |
| Sunshine duration in May | −0.460/<0.05 | 0.578/<0.05 | |
| Autumn | Mean temperature from winter solstice to start date of molting/end date of molting | 0.369/>0.05 | 0.615/>0.05 |
| Mean sunshine duration from summer solstice to start date of molting/end date of molting | −0.794/<0.05 | 0.862/<0.05 | |
| Sunshine duration of start date of molting/end date of molting | 0.665/>0.05 | 0.637/>0.05 | |
| Mean temperature of start date of molting/end date of molting | 0.525/>0.05 | −0.473/>0.05 | |
| Mean temperature in July | 0.105/>0.05 | 0.664/>0.05 | |
| Mean temperature in August | −0.451/<0.05 | −0.526/>0.05 | |
| Mean temperature in September | −0.378/>0.05 | −0.827/>0.05 | |
| Mean temperature in October | −0.493/>0.05 | 0.712/>0.05 | |
| Sunshine duration in July | −0.446/>0.05 | −0.750/>0.05 | |
| Sunshine duration in August | −0.723/<0.05 | −0.342/>0.05 | |
| Sunshine duration in September | 0.721/>0.05 | −0.750/>0.05 | |
| Sunshine duration in October | 0.475/>0.05 | 0.907/<0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, H.; Hua, Y.; Liang, X.; Long, Z.; Qi, J.; Wen, D.; Roberts, N.J.; Su, H.; Jiang, G. Wavelet Analysis Reveals Phenology Mismatch between Leaf Phenology of Temperate Forest Plants and the Siberian Roe Deer Molting under Global Warming. Remote Sens. 2022, 14, 3901. https://doi.org/10.3390/rs14163901
Cao H, Hua Y, Liang X, Long Z, Qi J, Wen D, Roberts NJ, Su H, Jiang G. Wavelet Analysis Reveals Phenology Mismatch between Leaf Phenology of Temperate Forest Plants and the Siberian Roe Deer Molting under Global Warming. Remote Sensing. 2022; 14(16):3901. https://doi.org/10.3390/rs14163901
Chicago/Turabian StyleCao, Heqin, Yan Hua, Xin Liang, Zexu Long, Jinzhe Qi, Dusu Wen, Nathan James Roberts, Haijun Su, and Guangshun Jiang. 2022. "Wavelet Analysis Reveals Phenology Mismatch between Leaf Phenology of Temperate Forest Plants and the Siberian Roe Deer Molting under Global Warming" Remote Sensing 14, no. 16: 3901. https://doi.org/10.3390/rs14163901
APA StyleCao, H., Hua, Y., Liang, X., Long, Z., Qi, J., Wen, D., Roberts, N. J., Su, H., & Jiang, G. (2022). Wavelet Analysis Reveals Phenology Mismatch between Leaf Phenology of Temperate Forest Plants and the Siberian Roe Deer Molting under Global Warming. Remote Sensing, 14(16), 3901. https://doi.org/10.3390/rs14163901