
Estimation of Maize Yield and Flowering Time Using Multi-Temporal UAV-Based Hyperspectral Data

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Trials and Ground Truth Data Collection
2.2. Hyperspectral Image Acquisition and Processing
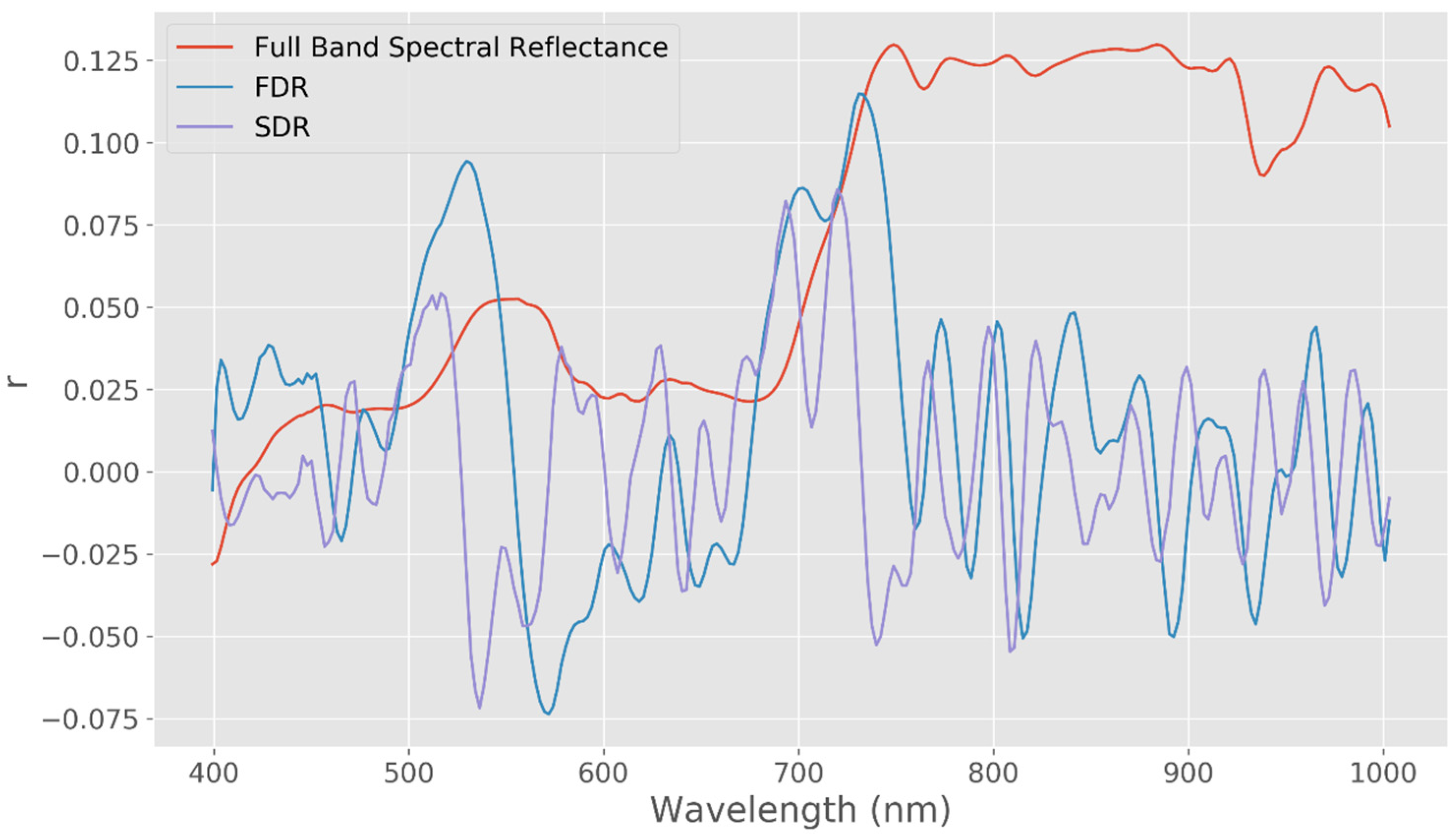
2.3. Spectral Feature Extraction
2.4. Model Development and Performance Evaluation
3. Results and Discussion
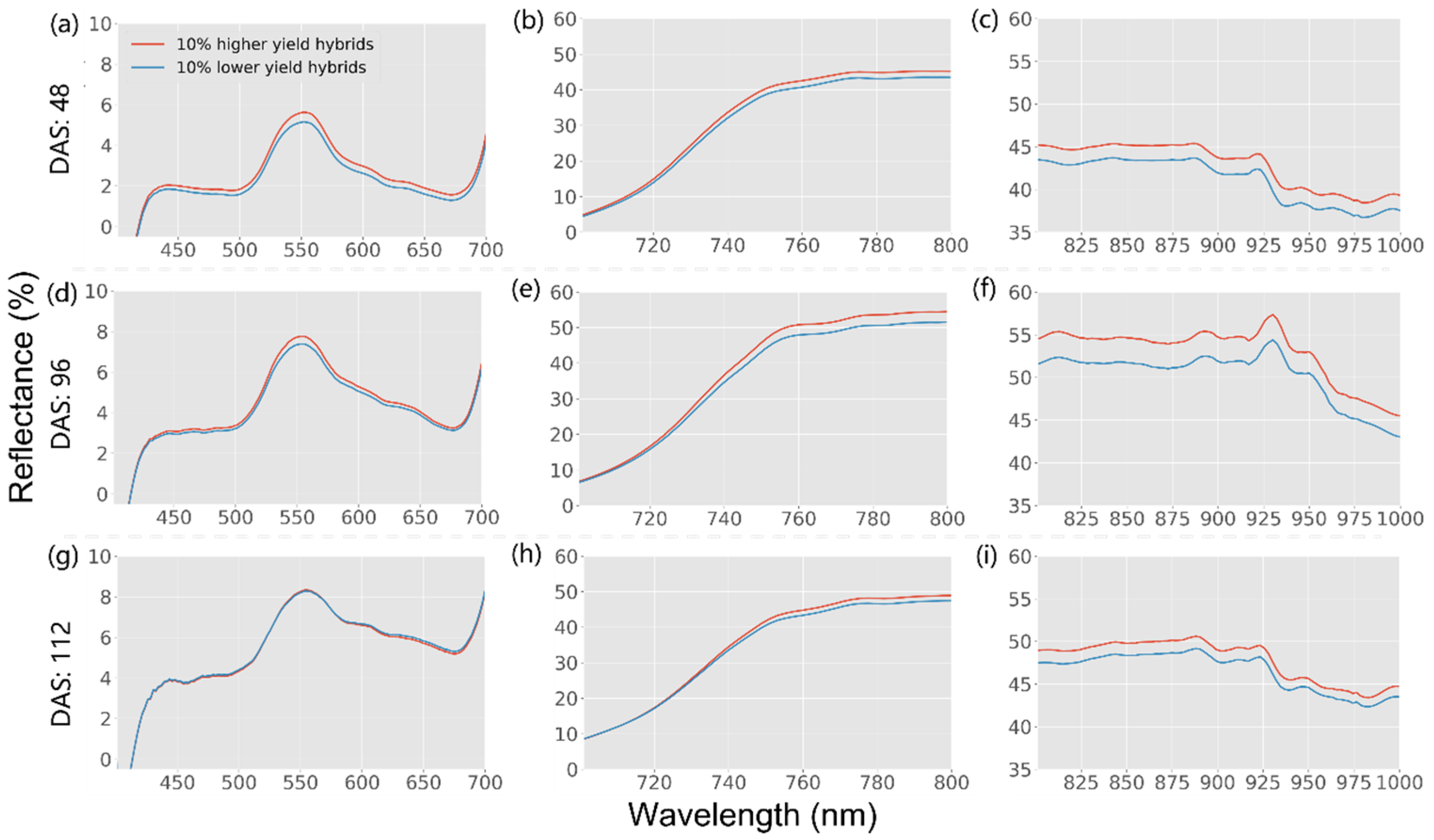
3.1. Ground Truth Field Data and Spectral Profiles
3.2. Estimation Performance by Models and Image Features
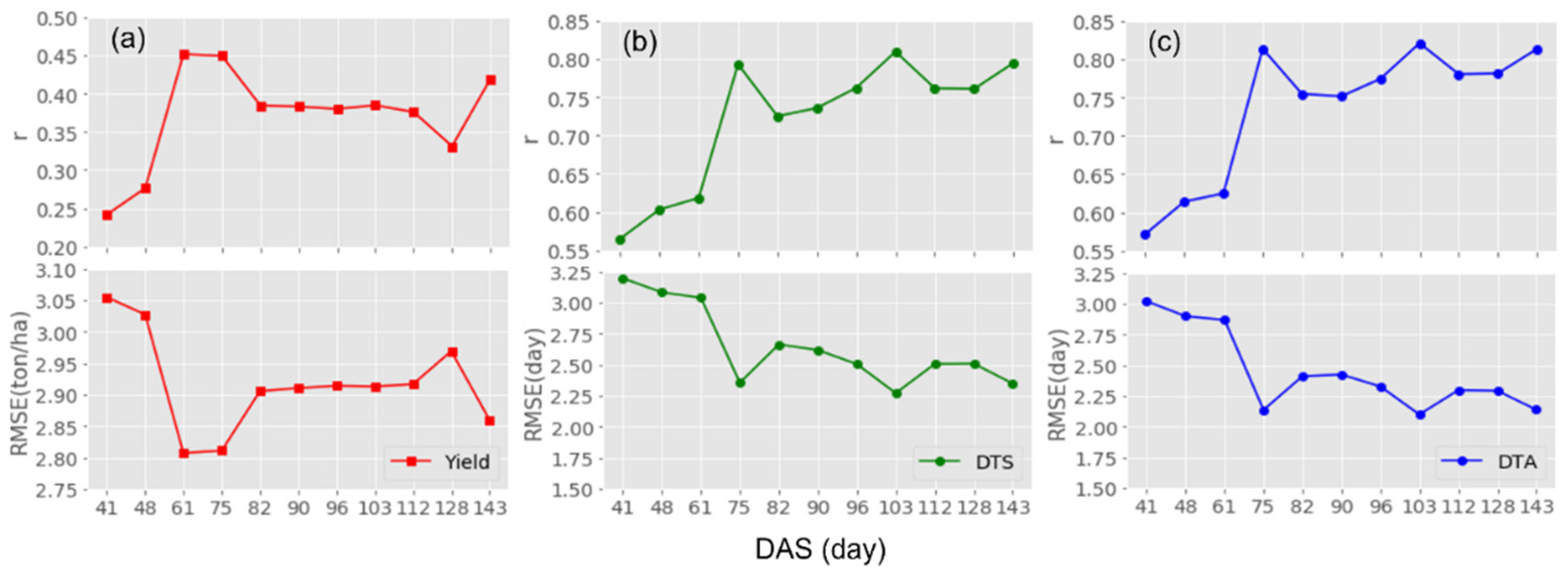
3.3. Model Performance by the UAV Survey Timing
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global Food Demand and the Sustainable Intensification of Agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosekov, A.Y.; Ivanova, S.A. Food Security: The Challenge of the Present. Geoforum 2018, 91, 73–77. [Google Scholar] [CrossRef]
- Pingali, P. Westernization of Asian Diets and the Transformation of Food Systems: Implications for Research and Policy. Food Policy 2007, 32, 281–298. [Google Scholar] [CrossRef] [Green Version]
- Breene, K. Food Security and Why It Matters. Available online: https://www.weforum.org/agenda/2016/01/food-security-and-why-it-matters/ (accessed on 10 January 2022).
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global Food Security, Biodiversity Conservation and the Future of Agricultural Intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Phalan, B.; Onial, M.; Balmford, A.; Green, R.E. Reconciling Food Production and Biodiversity Conservation: Land Sharing and Land Sparing Compared. Science 2011, 333, 1289–1291. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a Cultivated Planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Phalan, B.; Balmford, A.; Green, R.E.; Scharlemann, J.P.W. Minimising the Harm to Biodiversity of Producing More Food Globally. Food Policy 2011, 36, S62–S71. [Google Scholar] [CrossRef]
- Tanumihardjo, S.A.; McCulley, L.; Roh, R.; Lopez-Ridaura, S.; Palacios-Rojas, N.; Gunaratna, N.S. Maize Agro-Food Systems to Ensure Food and Nutrition Security in Reference to the Sustainable Development Goals. Glob. Food Secur. 2020, 25, 100327. [Google Scholar] [CrossRef]
- Maize Production Quantity. Available online: https://knoema.com/atlas/topics/Agriculture/Crops-Production-Quantity-tonnes/Maize-production?type=maps (accessed on 10 January 2022).
- Bruce, W.B.; Edmeades, G.O.; Barker, T.C. Molecular and Physiological Approaches to Maize Improvement for Drought Tolerance. J. Exp. Bot. 2002, 53, 13–25. [Google Scholar] [CrossRef]
- Rajcan, I.; Swanton, C.J. Understanding Maize-Weed Competition: Resource Competition, Light Quality and the Whole Plant. Field Crops Res. 2001, 71, 139–150. [Google Scholar] [CrossRef]
- Yang, W.; Nigon, T.; Hao, Z.; Dias Paiao, G.; Fernández, F.G.; Mulla, D.; Yang, C. Estimation of Corn Yield Based on Hyperspectral Imagery and Convolutional Neural Network. Comput. Electron. Agric. 2021, 184, 106092. [Google Scholar] [CrossRef]
- Rouf Shah, T.; Prasad, K.; Kumar, P. Maize—A Potential Source of Human Nutrition and Health: A Review. Cogent Food Agric. 2016, 2, 1166995. [Google Scholar] [CrossRef]
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops That Feed the World 6. Past Successes and Future Challenges to the Role Played by Maize in Global Food Security. Food Secur. 2011, 3, 307–327. [Google Scholar] [CrossRef] [Green Version]
- Poland, J. Breeding-Assisted Genomics. Curr. Opin. Plant Biol. 2015, 24, 119–124. [Google Scholar] [CrossRef] [PubMed]
- McMullen, M.D.; Kresovich, S.; Villeda, H.S.; Bradbury, P.; Li, H.; Sun, Q.; Flint-Garcia, S.; Thornsberry, J.; Acharya, C.; Bottoms, C.; et al. Genetic Properties of the Maize Nested Association Mapping Population. Science 2009, 325, 737–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chivasa, W.; Mutanga, O.; Burgueño, J. UAV-Based High-Throughput Phenotyping to Increase Prediction and Selection Accuracy in Maize Varieties under Artificial MSV Inoculation. Comput. Electron. Agric. 2021, 184, 106128. [Google Scholar] [CrossRef]
- Pugh, N.A.; Horne, D.W.; Murray, S.C.; Carvalho, G.; Malambo, L.; Jung, J.; Chang, A.; Maeda, M.; Popescu, S.; Chu, T.; et al. Temporal Estimates of Crop Growth in Sorghum and Maize Breeding Enabled by Unmanned Aerial Systems. Plant Phenome J. 2018, 1, 1–10. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Q.; Huang, D. A Review of Imaging Techniques for Plant Phenotyping. Sensors 2014, 14, 20078–20111. [Google Scholar] [CrossRef]
- Naik, H.S.; Zhang, J.; Lofquist, A.; Assefa, T.; Sarkar, S.; Ackerman, D.; Singh, A.; Singh, A.K.; Ganapathysubramanian, B. A Real-Time Phenotyping Framework Using Machine Learning for Plant Stress Severity Rating in Soybean. Plant Methods 2017, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Sankaran, S.; Zhou, J.; Khot, L.R.; Trapp, J.J.; Mndolwa, E.; Miklas, P.N. High-Throughput Field Phenotyping in Dry Bean Using Small Unmanned Aerial Vehicle Based Multispectral Imagery. Comput. Electron. Agric. 2018, 151, 84–92. [Google Scholar] [CrossRef]
- Singh, D.; Wang, X.; Kumar, U.; Gao, L.; Noor, M.; Imtiaz, M.; Singh, R.P.; Poland, J. High-Throughput Phenotyping Enabled Genetic Dissection of Crop Lodging in Wheat. Front. Plant Sci. 2019, 10, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Q.; Sun, N.; Bai, H.; Wang, N.; Fan, Z.; Wang, Y.; Meng, Z.; Li, B.; Cong, Y. Field-Based High-Throughput Phenotyping for Maize Plant Using 3d LIDAR Point Cloud Generated with a “Phenomobile”. Front. Plant Sci. 2019, 10, 554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehr, W.R. Principles of Cultivar Development; Macmillan: New York, NY, USA, 1987. [Google Scholar]
- Lee, E.A.; Tracy, W.F. Handbook of Maize: Genetics and Genomics; Springer: New York, NY, USA, 2009; ISBN 9780387778624. [Google Scholar]
- Zhou, J.; Zhou, J.; Ye, H.; Ali, M.L.; Chen, P.; Nguyen, H.T. Yield Estimation of Soybean Breeding Lines under Drought Stress Using Unmanned Aerial Vehicle-Based Imagery and Convolutional Neural Network. Biosyst. Eng. 2021, 204, 90–103. [Google Scholar] [CrossRef]
- Moreira, F.F.; Hearst, A.A.; Cherkauer, K.A.; Rainey, K.M. Improving the Efficiency of Soybean Breeding with High-Throughput Canopy Phenotyping. Plant Methods 2019, 15, 139. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Poland, J.A.; Mondal, S.; Crossa, J.; Juliana, P.; Singh, R.P.; Rutkoski, J.E.; Jannink, J.L.; Crespo-Herrera, L.; Velu, G.; et al. High-Throughput Phenotyping Platforms Enhance Genomic Selection for Wheat Grain Yield across Populations and Cycles in Early Stage. Theor. Appl. Genet. 2019, 132, 1705–1720. [Google Scholar] [CrossRef]
- Tanger, P.; Klassen, S.; Mojica, J.P.; Lovell, J.T.; Moyers, B.T.; Baraoidan, M.; Naredo, M.E.B.; McNally, K.L.; Poland, J.; Bush, D.R.; et al. Field-Based High Throughput Phenotyping Rapidly Identifies Genomic Regions Controlling Yield Components in Rice. Sci. Rep. 2017, 7, 42839. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Kim, H.C.; Shin, S.; Kim, K.H.; Moon, J.C.; Kim, J.Y.; Lee, B.M. Transcriptome Analysis of Flowering Time Genes under Drought Stress in Maize Leaves. Front. Plant Sci. 2017, 8, 267. [Google Scholar] [CrossRef] [Green Version]
- Buckler, E.S.; Holland, J.B.; Bradbury, P.J.; Acharya, C.B.; Brown, P.J.; Browne, C.; Ersoz, E.; Flint-Garcia, S.; Garcia, A.; Glaubitz, J.C.; et al. The Genetic Architecture of Maize Flowering Time. Science 2009, 325, 714–719. [Google Scholar] [CrossRef] [Green Version]
- Durand, E.; Bouchet, S.; Bertin, P.; Ressayre, A.; Jamin, P.; Charcosset, A.; Dillmann, C.; Tenaillon, M.I. Flowering Time in Maize: Linkage and Epistasis at a Major Effect Locus. Genetics 2012, 190, 1547–1562. [Google Scholar] [CrossRef] [Green Version]
- Franks, S.J. Plasticity and Evolution in Drought Avoidance and Escape in the Annual Plant Brassica rapa. New Phytol. 2011, 190, 249–257. [Google Scholar] [CrossRef]
- Andrade, F.H. Kernel Number Determination in Maize. Crop Sci. 1999, 39, 453–459. [Google Scholar] [CrossRef]
- Wu, G.; Miller, N.D.; de Leon, N.; Kaeppler, S.M.; Spalding, E.P. Predicting Zea mays Flowering Time, Yield, and Kernel Dimensions by Analyzing Aerial Images. Front. Plant Sci. 2019, 10, 1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghimi, A.; Yang, C.; Anderson, J.A. Aerial Hyperspectral Imagery and Deep Neural Networks for High-Throughput Yield Phenotyping in Wheat. Comput. Electron. Agric. 2020, 172, 105299. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhou, J.; Ye, H.; Ali, L.; Nguyen, H.T.; Chen, P. Classification of Soybean Leaf Wilting Due to Drought Stress Using UAV-Based Imagery. Comput. Electron. Agric. 2020, 175, 105576. [Google Scholar] [CrossRef]
- Gu, S.; Liao, Q.; Gao, S.; Kang, S.; Du, T.; Ding, R. Crop Water Stress Index as a Proxy of Phenotyping Maize Performance under Combined Water and Salt Stress. Remote Sens. 2021, 13, 4710. [Google Scholar] [CrossRef]
- Nikoli, A. Dynamics of Maize Vegetative Growth and Drought Adaptability Using Image-Based Phenotyping Under Controlled Conditions. Front. Plant Sci. 2021, 12, 571. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, H.; Wu, Z.; Wang, S.; Sun, H.; Senthilnath, J.; Wang, J.; Bryant, C.R.; Fu, Y. Modified Red Blue Vegetation Index for Chlorophyll Estimation and Yield Prediction of Maize from Visible Images Captured by Uav. Sensors 2020, 20, 5055. [Google Scholar] [CrossRef]
- Peng, X.; Han, W.; Ao, J.; Wang, Y. Assimilation of Lai Derived from UAV Multispectral Data into the Safy Model to Estimate Maize Yield. Remote Sens. 2021, 13, 1094. [Google Scholar] [CrossRef]
- Quemada, M.; Gabriel, J.L.; Zarco-Tejada, P. Airborne Hyperspectral Images and Ground-Level Optical Sensors as Assessment Tools for Maize Nitrogen Fertilization. Remote Sens. 2014, 6, 2940–2962. [Google Scholar] [CrossRef] [Green Version]
- Danilevicz, M.F.; Bayer, P.E.; Boussaid, F.; Bennamoun, M.; Edwards, D. Maize Yield Prediction at an Early Developmental Stage Using Multispectral Images and Genotype Data for Preliminary Hybrid Selection. Remote Sens. 2021, 13, 3976. [Google Scholar] [CrossRef]
- Herrmann, I.; Bdolach, E.; Montekyo, Y.; Rachmilevitch, S.; Townsend, P.A.; Karnieli, A. Assessment of Maize Yield and Phenology by Drone-Mounted Superspectral Camera. Precis. Agric. 2020, 21, 51–76. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, L. Technical Focus: Visible and near-Infrared Reflectance Techniques for Diagnosing Plant Physiological Status. Trends Plant Sci. 1998, 3, 151–156. [Google Scholar] [CrossRef]
- Babar, M.A.; Reynolds, M.P.; Van Ginkel, M.; Klatt, A.R.; Raun, W.R.; Stone, M.L. Spectral Reflectance to Estimate Genetic Variation for In-Season Biomass, Leaf Chlorophyll, and Canopy Temperature in Wheat. Crop Sci. 2006, 46, 1046–1057. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Cavender-Bares, J.; Townsend, P.A.; Hobbie, S.E.; Madritch, M.D.; Wang, R.; Tilman, D.; Gamon, J.A. Plant Spectral Diversity Integrates Functional and Phylogenetic Components of Biodiversity and Predicts Ecosystem Function. Nat. Ecol. Evol. 2018, 2, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Kycko, M.; Zagajewski, B.; Lavender, S.; Romanowska, E.; Zwijacz-Kozica, M. The Impact of Tourist Traffic on the Condition and Cell Structures of Alpine Swards. Remote Sens. 2018, 10, 220. [Google Scholar] [CrossRef] [Green Version]
- Yoosefzadeh-Najafabadi, M.; Earl, H.J.; Tulpan, D.; Sulik, J.; Eskandari, M. Application of Machine Learning Algorithms in Plant Breeding: Predicting Yield From Hyperspectral Reflectance in Soybean. Front. Plant Sci. 2021, 11, 624273. [Google Scholar] [CrossRef]
- Gao, S.; Niu, Z.; Huang, N.; Hou, X. Estimating the Leaf Area Index, Height and Biomass of Maize Using HJ-1 and RADARSAT-2. Int. J. Appl. Earth Obs. Geoinf. 2013, 24, 1–8. [Google Scholar] [CrossRef]
- Gnyp, M.L.; Miao, Y.; Yuan, F.; Ustin, S.L.; Yu, K.; Yao, Y.; Huang, S.; Bareth, G. Hyperspectral Canopy Sensing of Paddy Rice Aboveground Biomass at Different Growth Stages. Field Crops Res. 2014, 155, 42–55. [Google Scholar] [CrossRef]
- Fu, Y.; Yang, G.; Wang, J.; Song, X.; Feng, H. Winter Wheat Biomass Estimation Based on Spectral Indices, Band Depth Analysis and Partial Least Squares Regression Using Hyperspectral Measurements. Comput. Electron. Agric. 2014, 100, 51–59. [Google Scholar] [CrossRef]
- Kross, A.; McNairn, H.; Lapen, D.; Sunohara, M.; Champagne, C. Assessment of RapidEye Vegetation Indices for Estimation of Leaf Area Index and Biomass in Corn and Soybean Crops. Int. J. Appl. Earth Obs. Geoinf. 2015, 34, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Zhang, Z.; Ma, Y.; Du, Q.; Williams, P.; Drewry, J.; Luck, B. Alfalfa Yield Prediction Using UAV-Based Hyperspectral Imagery and Ensemble Learning. Remote Sens. 2020, 12, 2028. [Google Scholar] [CrossRef]
- Thorp, K.R.; Wang, G.; Bronson, K.F.; Badaruddin, M.; Mon, J. Hyperspectral Data Mining to Identify Relevant Canopy Spectral Features for Estimating Durum Wheat Growth, Nitrogen Status, and Grain Yield. Comput. Electron. Agric. 2017, 136, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tsai, F.; Philpot, W. Derivative Analysis of Hyperspectral Data. Remote Sens. Environ. 1998, 66, 41–51. [Google Scholar] [CrossRef]
- Tsai, F.; Philpot, W.D. A Derivative-Aided Hyperspectral Image Analysis System for Land-Cover Classification. IEEE Trans. Geosci. Remote Sens. 2002, 40, 416–425. [Google Scholar] [CrossRef]
- Bao, J.; Chi, M.; Benediktsson, J.A. Spectral Derivative Features for Classification of Hyperspectral Remote Sensing Images: Experimental Evaluation. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2013, 6, 594–601. [Google Scholar] [CrossRef]
- Kalluri, H.R.; Prasad, S.; Bruce, L.M. Decision-Level Fusion of Spectral Reflectance and Derivative Information for Robust Hyperspectral Land Cover Classification. IEEE Trans. Geosci. Remote Sens. 2010, 48, 4047–4058. [Google Scholar] [CrossRef]
- Smith, K.L.; Steven, M.D.; Colls, J.J. Use of Hyperspectral Derivative Ratios in the Red-Edge Region to Identify Plant Stress Responses to Gas Leaks. Remote Sens. Environ. 2004, 92, 207–217. [Google Scholar] [CrossRef]
- Thorp, K.R.; Tian, L.; Yao, H.; Tang, L. Narrow-Band and Derivative-Based Vegetation Indices for Hyperspectral Data. Trans. ASAE 2004, 47, 291–299. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J.E. Smoothing and Differentiation. Anal. Chem 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Hoerl, A.E.; Kennard, R.W. Ridge Regression: Biased Estimation for Nonorthogonal Problems. Technometrics 1970, 12, 55–67. [Google Scholar] [CrossRef]
- Vogt, W. Ridge Regression. In Dictionary of Statistics and Methodology; Sage: New York, NY, USA, 2015; pp. 1–20. [Google Scholar] [CrossRef]
- Vapnik, V. The Nature of Statistical Learning Theory; Springer: New York, NY, USA, 2013. [Google Scholar]
- Auria, L.; Moro, R.A. Support Vector Machines (SVM) as a Technique for Solvency Analysis. SSRN Electron. J. 2011. [Google Scholar] [CrossRef] [Green Version]
- Drucker, H.; Surges, C.J.C.; Kaufman, L.; Smola, A.; Vapnik, V. Support Vector Regression Machines. Adv. Neural Inf. Process. Syst. 1997, 1, 155–161. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhou, X.; Zhu, X.; Dong, Z.; Guo, W. Estimation of Biomass in Wheat Using Random Forest Regression Algorithm and Remote Sensing Data. Crop J. 2016, 4, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Z.; Feng, L.; Du, Q. Combining Multi-Source Data and Machine Learning Approaches to Predict Winter Wheat Yield in The. Remote Sens. 2020, 12, 1232. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Li, Y.; Chen, Y.; Du, K.; Zheng, F.; Zhang, L.; Sun, Z. Estimating above Ground Biomass of Winter Wheat at Early Growth Stages Using Digital Images and Deep Convolutional Neural Network. Eur. J. Agron. 2019, 103, 117–129. [Google Scholar] [CrossRef]
- Feng, L.; Wang, Y.; Zhang, Z.; Du, Q. Geographically and Temporally Weighted Neural Network for Winter Wheat Yield Prediction. Remote Sens. Environ. 2021, 262, 112514. [Google Scholar] [CrossRef]
- Feng, L.; Zhang, Z.; Ma, Y.; Sun, Y.; Du, Q.; Williams, P.; Drewry, J.; Luck, B. Multitask Learning of Alfalfa Nutritive Value From UAV-Based Hyperspectral Images. IEEE Geosci. Remote Sens. Lett. 2021, 19, 1–5. [Google Scholar] [CrossRef]
- Masjedi, A.; Crawford, M.M.; Carpenter, N.R.; Tuinstra, M.R. Multi-Temporal Predictive Modelling of Sorghum Biomass Using UAV-Based Hyperspectral and LiDAR Data. Remote Sens. 2020, 12, 3587. [Google Scholar] [CrossRef]
- Zhou, J.; Ye, H.; Nguyen, H.; Chen, P.; Zhou, J. Yield Estimation of Soybean Breeding Lines Using UAV Multispectral Imagery and Convolutional Neuron Network Jing. In Proceedings of the 2020 ASABE Annual International Virtual Meeting, Virtual, 13–15 July 2020; pp. 2–14. [Google Scholar]
- Gutierrez-Rodriguez, M.; Escalante-Estrada, J.A.; Rodriguez-Gonzalez, M.T. Canopy Reflectance, Stomatal Conductance, and Yield of Phaseolus vulgaris L. and Phaseolus coccinues L. Under Saline Field Conditions. Int. J. Agric. Biol. 2005, 7, 491–494. [Google Scholar]
- Candiago, S.; Remondino, F.; De Giglio, M.; Dubbini, M.; Gattelli, M. Evaluating Multispectral Images and Vegetation Indices for Precision Farming Applications from UAV Images. Remote Sens. 2015, 7, 4026–4047. [Google Scholar] [CrossRef] [Green Version]
- Condorelli, G.E.; Maccaferri, M.; Newcomb, M.; Andrade-Sanchez, P.; White, J.W.; French, A.N.; Sciara, G.; Ward, R.; Tuberosa, R. Comparative Aerial and Ground Based High Throughput Phenotyping for the Genetic Dissection of NDVI as a Proxy for Drought Adaptive Traits in Durum Wheat. Front. Plant Sci. 2018, 9, 893. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Chen, J.; Zhu, X.; Tang, Y. Yellow Flowers Can Decrease NDVI and EVI Values: Evidence from a Field Experiment in an Alpine Meadow. Can. J. Remote Sens. 2009, 35, 99–106. [Google Scholar] [CrossRef]
- Shen, M.; Chen, J.; Zhu, X.; Tang, Y.; Chen, X. Do Flowers Affect Biomass Estimate Accuracy from NDVI and EVI? Int. J. Remote Sens. 2010, 31, 2139–2149. [Google Scholar] [CrossRef]
- Bernardo, R. Parental Selection, Number of Breeding Populations, and Size of Each Population in Inbred Development. Theor. Appl. Genet. 2003, 107, 1252–1256. [Google Scholar] [CrossRef]
- Rogers, A.R.; Dunne, J.C.; Romay, C.; Bohn, M.; Buckler, E.S.; Ciampitti, I.A.; Edwards, J.; Ertl, D.; Flint-Garcia, S.; Gore, M.A.; et al. The Importance of Dominance and Genotype-by-Environment Interactions on Grain Yield Variation in a Large-Scale Public Cooperative Maize Experiment. G3 Genes Genomes Genet. 2021, 11, jkaa050. [Google Scholar] [CrossRef]
- Kumar, A.; Rajalakshmi, P.; Guo, W.; Naik, B.B.; Marathi, B.; Desai, U.B. Detection and Counting of Tassels for Maize Crop Monitoring Using Multispectral Images. In Proceedings of the 2020 IEEE International Conference on Computing, Power and Communication Technologies (GUCON), Greater Noida, India, 2–4 October 2020; pp. 789–793. [Google Scholar] [CrossRef]
- Lambert, R.J.; Johnson, R.R. Leaf Angle, Tassel Morphology, and the Performance of Maize Hybrids 1. Crop Sci. 1978, 18, 499–502. [Google Scholar] [CrossRef]
- Hunter, R.B.; Daynard, T.B.; Hume, D.J.; Tanner, J.W.; Curtis, J.D.; Kannenberg, L.W. Effect of Tassel Removal on Grain Yield of Corn (Zea mays L.) 1. Crop Sci. 1969, 9, 405–406. [Google Scholar] [CrossRef]
- Gage, J.L.; Miller, N.D.; Spalding, E.P.; Kaeppler, S.M.; de Leon, N. TIPS: A System for Automated Image-Based Phenotyping of Maize Tassels. Plant Methods 2017, 13, 21. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Feng, L.; Zhang, Z.; Ma, Y.; Crosby, T.; Naber, M.; Wang, Y. Prediction of End-of-Season Tuber Yield and Tuber Set in Potatoes Using in-Season Uav-Based Hyperspectral Imagery and Machine Learning. Sensors 2020, 20, 5293. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Smith, R.B.; De Pauw, E. Hyperspectral Vegetation Indices and Their Relationships with Agricultural Crop Characteristics. Remote Sens. Environ. 2000, 71, 158–182. [Google Scholar] [CrossRef]
- Myneni, R.B.; Williams, D.L. On the Relationship between FAPAR and NDVI. Remote Sens. Environ. 1994, 49, 200–211. [Google Scholar] [CrossRef]
- Prabhakara, K.; Hively, W.D.; Mccarty, G.W. Evaluating the Relationship between Biomass, Percent Groundcover and Remote Sensing Indices across Six Winter Cover Crop Fields in Maryland, United States. Int. J. Appl. Earth Obs. Geoinf. 2015, 39, 88–102. [Google Scholar] [CrossRef] [Green Version]
- Viña, A.; Leavitt, B.; Rundquist, D.C.; Keydan, G.; Leavitt, B.; Schepers, J. Monitoring Maize (Zea mays L.) Phenology with Remote Sensing. Agron. J. 2004, 96, 1139–1147. [Google Scholar] [CrossRef]
- Tesfaye, A.A. Evaluation of the Saturation Property of Vegetation Indices Derived from Sentinel-2 in Mixed Crop-Forest Ecosystem. Spat. Inf. Res. 2021, 29, 109–121. [Google Scholar] [CrossRef]
- Gitelson, A.A. Wide Dynamic Range Vegetation Index for Remote Quantification of Biophysical Characteristics of Vegetation. J. Plant Physiol. 2004, 161, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Abendroth, L.J.; Elmore, R.W.; Boyer, M.J.; Marlay, S.K. Corn Growth and Development; Iowa State University: Ames, IA, USA, 2011. [Google Scholar]
- Nielsen, R.L. Ear Size Determination in Corn. Available online: https://www.agry.purdue.edu/ext/corn/news/timeless/EarSize.html (accessed on 10 January 2022).
- Hanway, J.J. How a Corn Plant Develops; Special Report; Iowa Agricultural and Home Economics Experiment State Publications: Ames, IA, USA, 1966; Volume 48, pp. 1–18. [Google Scholar]
- Nielsen, R.L. Grain Fill Stages in Corn. Available online: https://www.agry.purdue.edu/ext/corn/news/timeless/grainfill.html (accessed on 10 January 2022).
- Nleya, T.; Chungu, C.; Kleinjan, J. Corn Growth and Development. In Best Management Practices; South Dakota University: Brookings, SD, USA, 2016. [Google Scholar]
- Ransom, J.; Endres, G. Corn: Growth and Management Quick Guide: Revised; North Dakota State University: Fargo, ND, USA, 2020; Volume 1173, pp. 1–8. [Google Scholar]
- Teal, R.; Tubana, B.; Arnall, B.; Walsh, O. In-Season Prediction of Corn Grain Yield Potential Using Normalized Difference Vegetation Index. Agron. J. 2006, 98, 1488–1494. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Michael, O.; Hendley, P.; Drost, D. Corn and Soybean Yield Indicators Using Remotely Sensed Vegetation Index. In Proceedings of the Fourth International Conference on Precision Agriculture, St. Paul, MN, USA, 19–22 July 1998; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 1999; pp. 1475–1481. [Google Scholar]
- Spitkó, T.; Nagy, Z.; Zsubori, Z.T.; Szőke, C.; Berzy, T.; Pintér, J.; Marton, C.L. Connection between Normalized Difference Vegetation Index and Yield in Maize. Plant Soil Environ. 2016, 62, 293–298. [Google Scholar] [CrossRef]
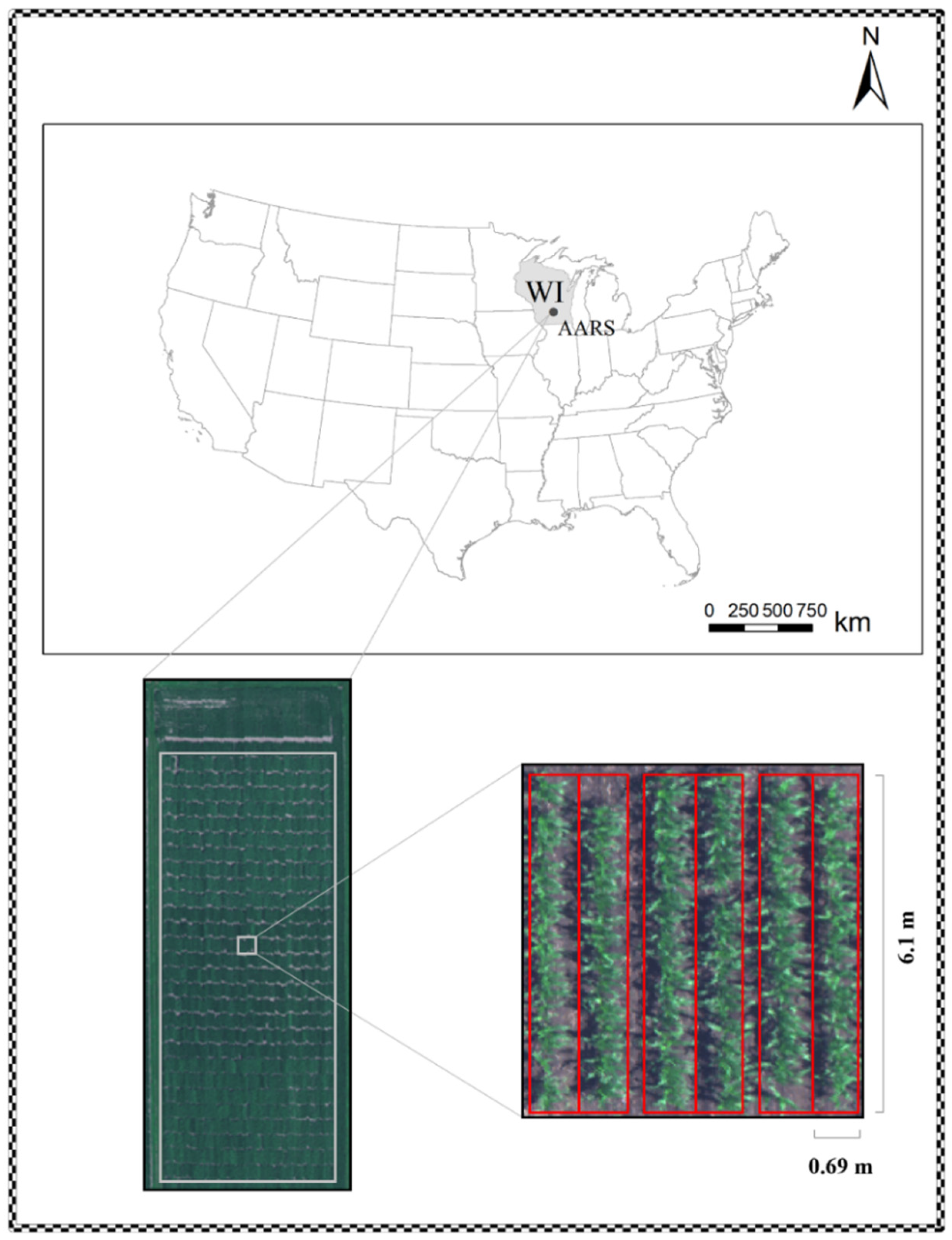




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flight Dates | DAS | Growth Stages |
|---|---|---|
| 23 June | 41 | V6–V8 (6–8 Leaf Collars) |
| 30 June | 48 | V9–V11 (9–11 Leaf Collars) |
| 13 July | 61 | VT (Tasseling) |
| 27 July | 75 | R1 (Silking) |
| 3 August | 82 | R2 (Blister) |
| 11 August | 90 | R3 (Milk) |
| 17 August | 96 | R4 (Dough) |
| 24 August | 103 | R4 (Dough) |
| 2 September | 112 | R5 (Dent) |
| 18 September | 128 | R5 (Dent) |
| 3 October | 143 | R6 (Black Layer) |
| Growth Stages | Region | t | p-Value |
|---|---|---|---|
| Early Stages, 48 DAS | 400–700 nm | −1.569091 | 0.117789 |
| 700–800 nm | −0.486646 | 0.627719 | |
| 800–100 nm | −4.490146 | 0.000013 | |
| Middle Stages, 96 DAS | 400–700 nm | −0.755478 | 0.450616 |
| 700–800 nm | −0.594924 | 0.553421 | |
| 800–100 nm | −5.709019 | 0.000000 | |
| Late Stages, 112 DAS | 400–700 nm | 0.191138 | 0.848560 |
| 700–800 nm | −0.323303 | 0.747233 | |
| 800–100 nm | −3.763716 | 0.000226 |
| Phenotypic Data | Model | Features | Metrics | ||
|---|---|---|---|---|---|
| r | MAE | RMSE | |||
| Yield (ton/ha) | Ridge | Full Band Spectral Reflectance | 0.54 | 2.10 | 2.68 |
| (0.01) | (0.02) | (0.03) | |||
| First Derivatives of Full Bands | 0.41 | 2.74 | 3.51 | ||
| (0.01) | (0.03) | (0.04) | |||
| Second Derivatives of Full Bands | 0.37 | 2.97 | 3.78 | ||
| (0.02) | (0.05) | (0.06) | |||
| 81 Vegetation Indices | 0.15 | 2.55 | 12.26 | ||
| (0.09) | (0.42) | (14.88) | |||
| SVR | Full Band Spectral Reflectance | 0.53 | 2.10 | 2.69 | |
| (0.01) | (0.02) | (0.02) | |||
| First Derivatives of Full Bands | 0.49 | 2.30 | 2.95 | ||
| (0.01) | (0.03) | (0.04) | |||
| Second Derivatives of Full Bands | 0.45 | 2.41 | 3.08 | ||
| (0.01) | (0.03) | (0.05) | |||
| 81 Vegetation Indices | 0.47 | 2.13 | 2.91 | ||
| (0.01) | (0.02) | (0.05) | |||
| RF | Full Band Spectral Reflectance | 0.44 | 2.33 | 2.83 | |
| (0.01) | (0.01) | (0.01) | |||
| First Derivatives of Full Bands | 0.48 | 2.29 | 2.78 | ||
| (0.01) | (0.01) | (0.01) | |||
| Second Derivatives of Full Bands | 0.46 | 2.34 | 2.82 | ||
| (0.01) | (0.01) | (0.01) | |||
| 81 Vegetation Indices | 0.46 | 2.30 | 2.80 | ||
| (0.01) | (0.01) | (0.01) | |||
| DTS (day) | Ridge | Full Band Spectral Reflectance | 0.91 | 1.27 | 1.63 |
| (<0.01) | (0.01) | (0.01) | |||
| First Derivatives of Full Bands | 0.86 | 1.60 | 2.05 | ||
| (<0.01) | (0.03) | (0.03) | |||
| Second Derivatives of Full Bands | 0.82 | 1.84 | 2.34 | ||
| (0.01) | (0.02) | (0.03) | |||
| 81 Vegetation Indices | 0.24 | 1.90 | 17.40 | ||
| (0.10) | (0.34) | (12.24) | |||
| SVR | Full Band Spectral Reflectance | 0.89 | 1.35 | 1.76 | |
| (<0.01) | (0.02) | (0.03) | |||
| First Derivatives of Full Bands | 0.89 | 1.42 | 1.81 | ||
| (<0.01) | (0.02) | (0.03) | |||
| Second Derivatives of Full Bands | 0.86 | 1.57 | 2.02 | ||
| (<0.01) | (0.02) | (0.03) | |||
| 81 Vegetation Indices | 0.74 | 1.42 | 2.99 | ||
| (0.05) | (0.04) | (0.48) | |||
| RF | Full Band Spectral Reflectance | 0.84 | 1.59 | 2.12 | |
| (0.01) | (0.01) | (0.03) | |||
| First Derivatives of Full Bands | 0.85 | 1.54 | 2.02 | ||
| (<0.01) | (0.01) | (0.02) | |||
| Second Derivatives of Full Bands | 0.83 | 1.63 | 2.20 | ||
| (0.01) | (0.02) | (0.03) | |||
| 81 Vegetation Indices | 0.86 | 1.50 | 1.96 | ||
| (<0.01) | (0.01) | (0.01) | |||
| DTA (day) | Ridge | Full Band Spectral Reflectance | 0.92 | 1.16 | 1.48 |
| (<0.01) | (0.01) | (0.01) | |||
| First Derivatives of Full Bands | 0.87 | 1.49 | 1.89 | ||
| (<0.01) | (0.03) | (0.04) | |||
| Second Derivatives of Full Bands | 0.84 | 1.63 | 2.07 | ||
| (<0.01) | (0.03) | (0.02) | |||
| 81 Vegetation Indices | 0.43 | 1.54 | 10.18 | ||
| (0.22) | (0.27) | (9.29) | |||
| SVR | Full Band Spectral Reflectance | 0.90 | 1.24 | 1.61 | |
| (<0.01) | (0.02) | (0.03) | |||
| First Derivatives of Full Bands | 0.89 | 1.32 | 1.69 | ||
| (<0.01) | (0.01) | (0.02) | |||
| Second Derivatives of Full Bands | 0.87 | 1.42 | 1.81 | ||
| (<0.01) | (0.02) | (0.03) | |||
| 81 Vegetation Indices | 0.79 | 1.27 | 2.45 | ||
| (0.04) | (0.03) | (0.29) | |||
| RF | Full Band Spectral Reflectance | 0.85 | 1.48 | 1.97 | |
| (0.01) | (0.01) | (0.03) | |||
| First Derivatives of Full Bands | 0.87 | 1.39 | 1.84 | ||
| (<0.01) | (0.01) | (0.02) | |||
| Second Derivatives of Full Bands | 0.84 | 1.52 | 2.04 | ||
| (<0.01) | (0.01) | (0.02) | |||
| 81 Vegetation Indices | 0.88 | 1.35 | 1.78 | ||
| (<0.01) | (0.01) | (0.02) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Zhou, J.; Wang, B.; de Leon, N.; Kaeppler, S.M.; Lima, D.C.; Zhang, Z. Estimation of Maize Yield and Flowering Time Using Multi-Temporal UAV-Based Hyperspectral Data. Remote Sens. 2022, 14, 3052. https://doi.org/10.3390/rs14133052
Fan J, Zhou J, Wang B, de Leon N, Kaeppler SM, Lima DC, Zhang Z. Estimation of Maize Yield and Flowering Time Using Multi-Temporal UAV-Based Hyperspectral Data. Remote Sensing. 2022; 14(13):3052. https://doi.org/10.3390/rs14133052
Chicago/Turabian StyleFan, Jiahao, Jing Zhou, Biwen Wang, Natalia de Leon, Shawn M. Kaeppler, Dayane C. Lima, and Zhou Zhang. 2022. "Estimation of Maize Yield and Flowering Time Using Multi-Temporal UAV-Based Hyperspectral Data" Remote Sensing 14, no. 13: 3052. https://doi.org/10.3390/rs14133052
APA StyleFan, J., Zhou, J., Wang, B., de Leon, N., Kaeppler, S. M., Lima, D. C., & Zhang, Z. (2022). Estimation of Maize Yield and Flowering Time Using Multi-Temporal UAV-Based Hyperspectral Data. Remote Sensing, 14(13), 3052. https://doi.org/10.3390/rs14133052