
Dormant Season Vegetation Phenology and Eddy Fluxes in Native Tallgrass Prairies of the U.S. Southern Plains

 ,
,  ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
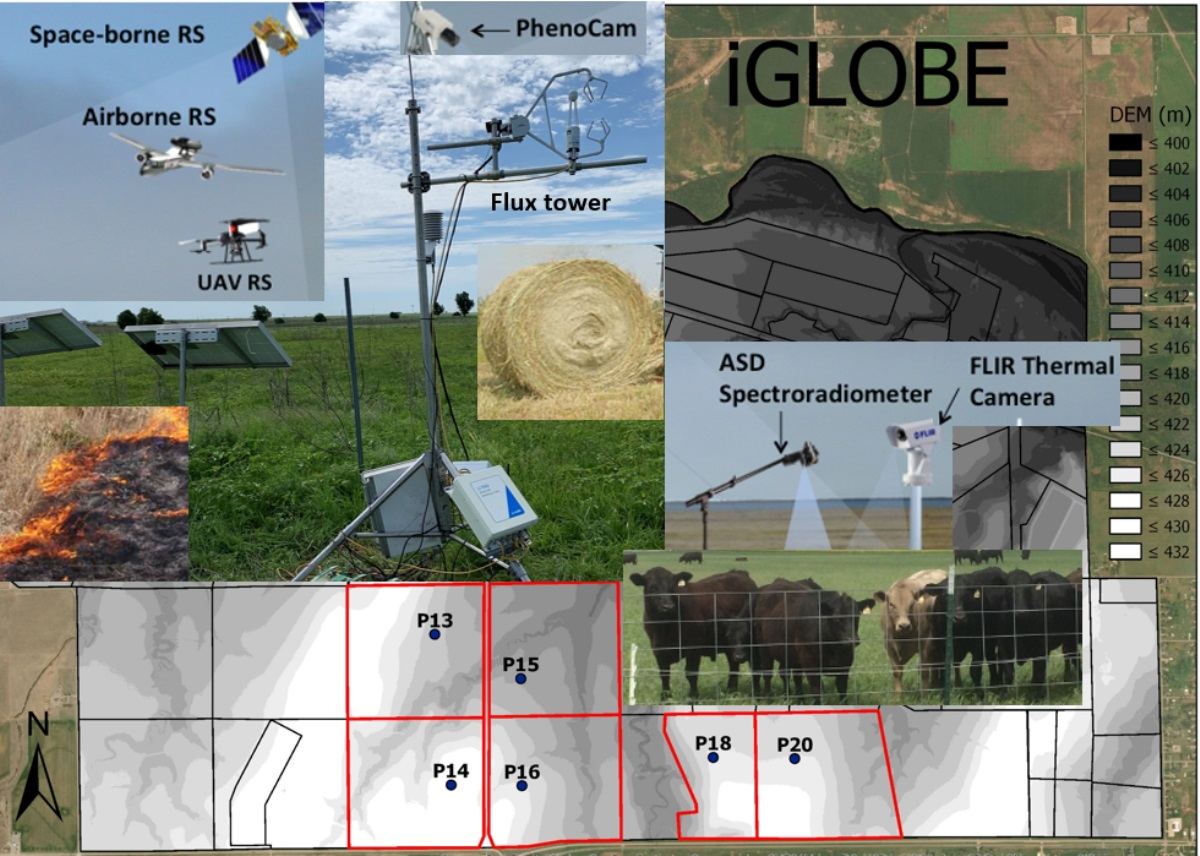
2.1. Study Sites
2.2. Eddy Covariance Measurements
2.3. Satellite Remote Sensing Data
3. Results and Discussion
3.1. Weather Conditions for the Study Period
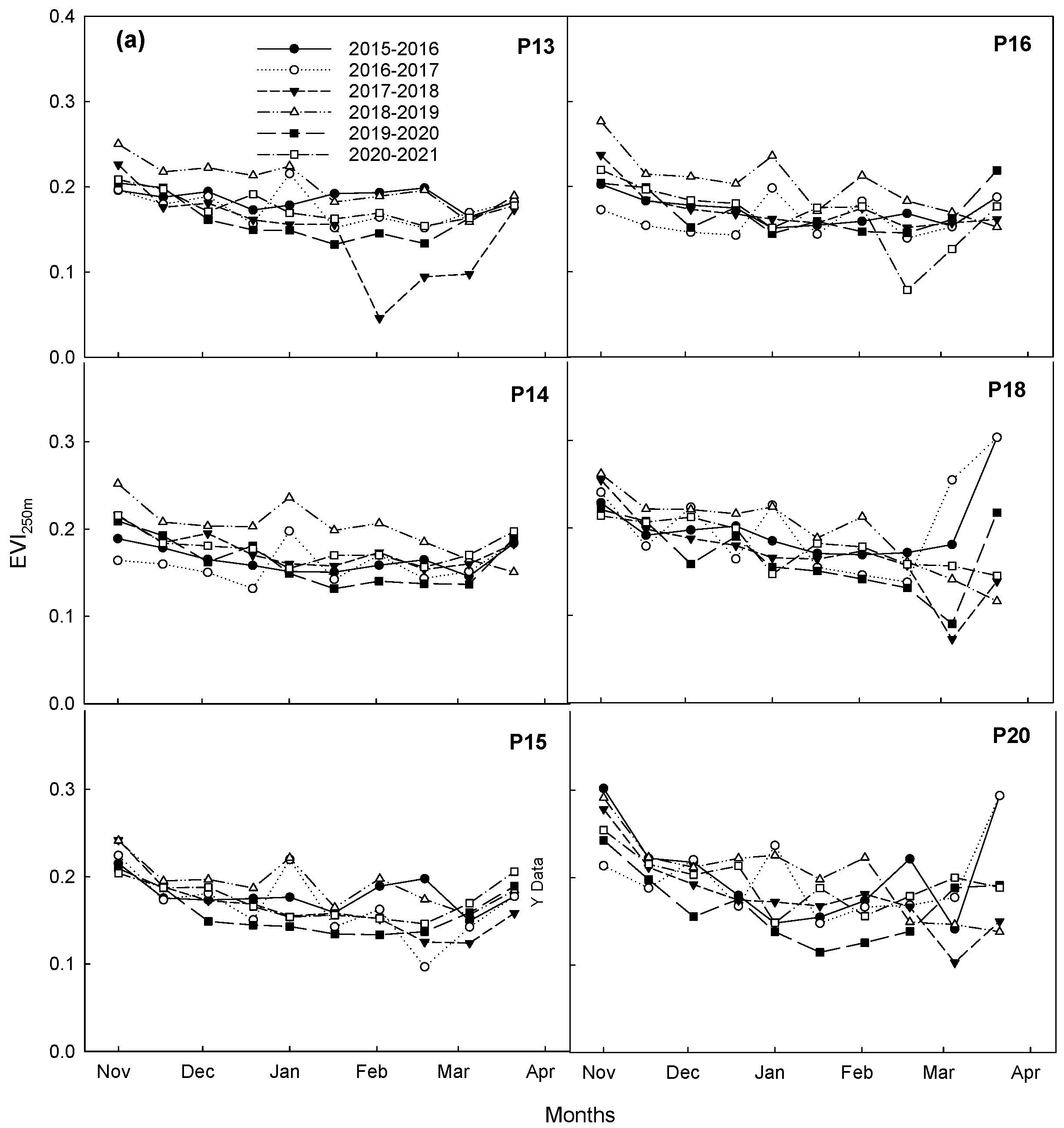
3.2. Interannual Variability in Vegetation Phenology during Dormant Seasons
3.3. Comparison of Vegetation Phenology during Dormant Season among Pastures
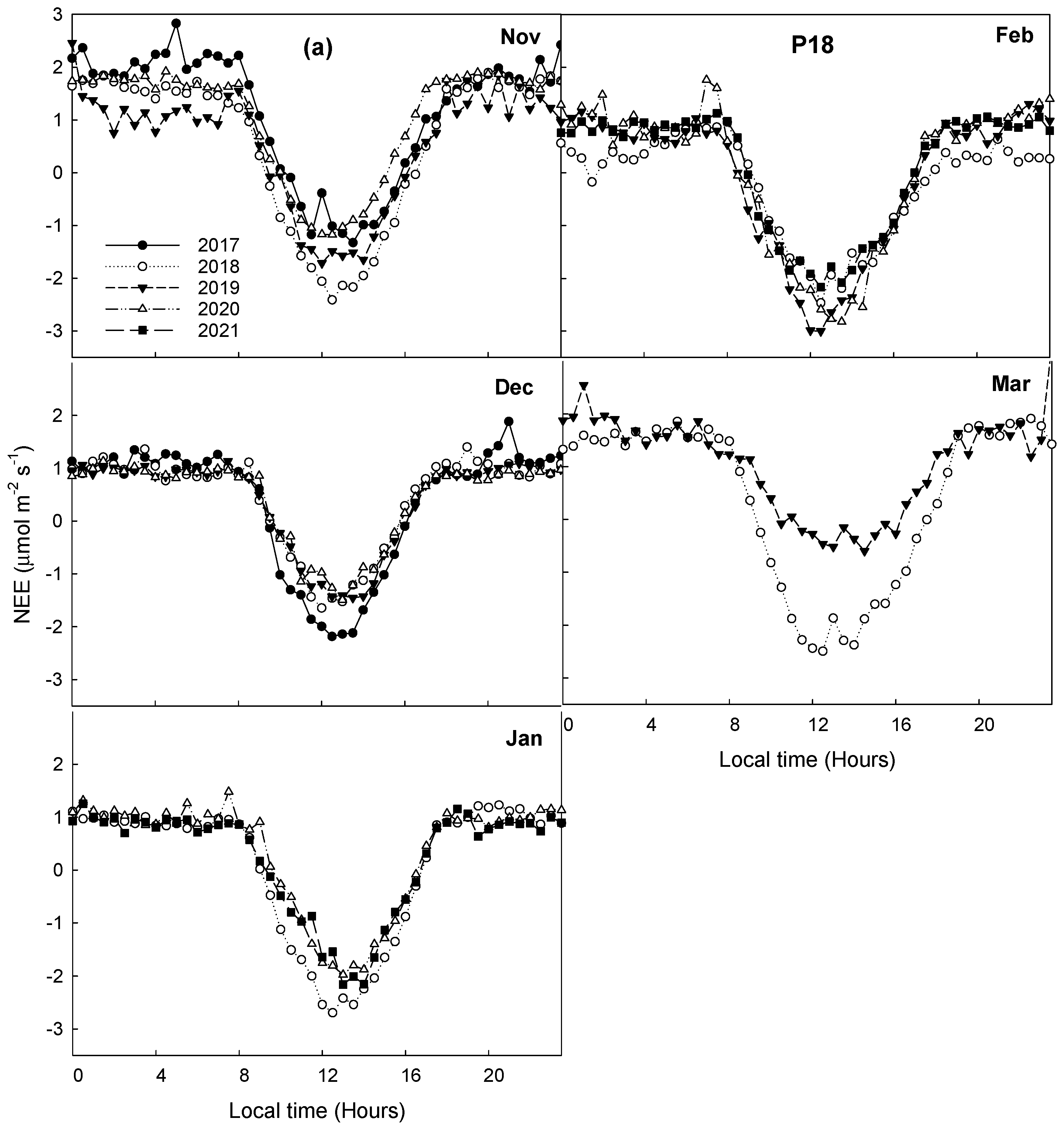
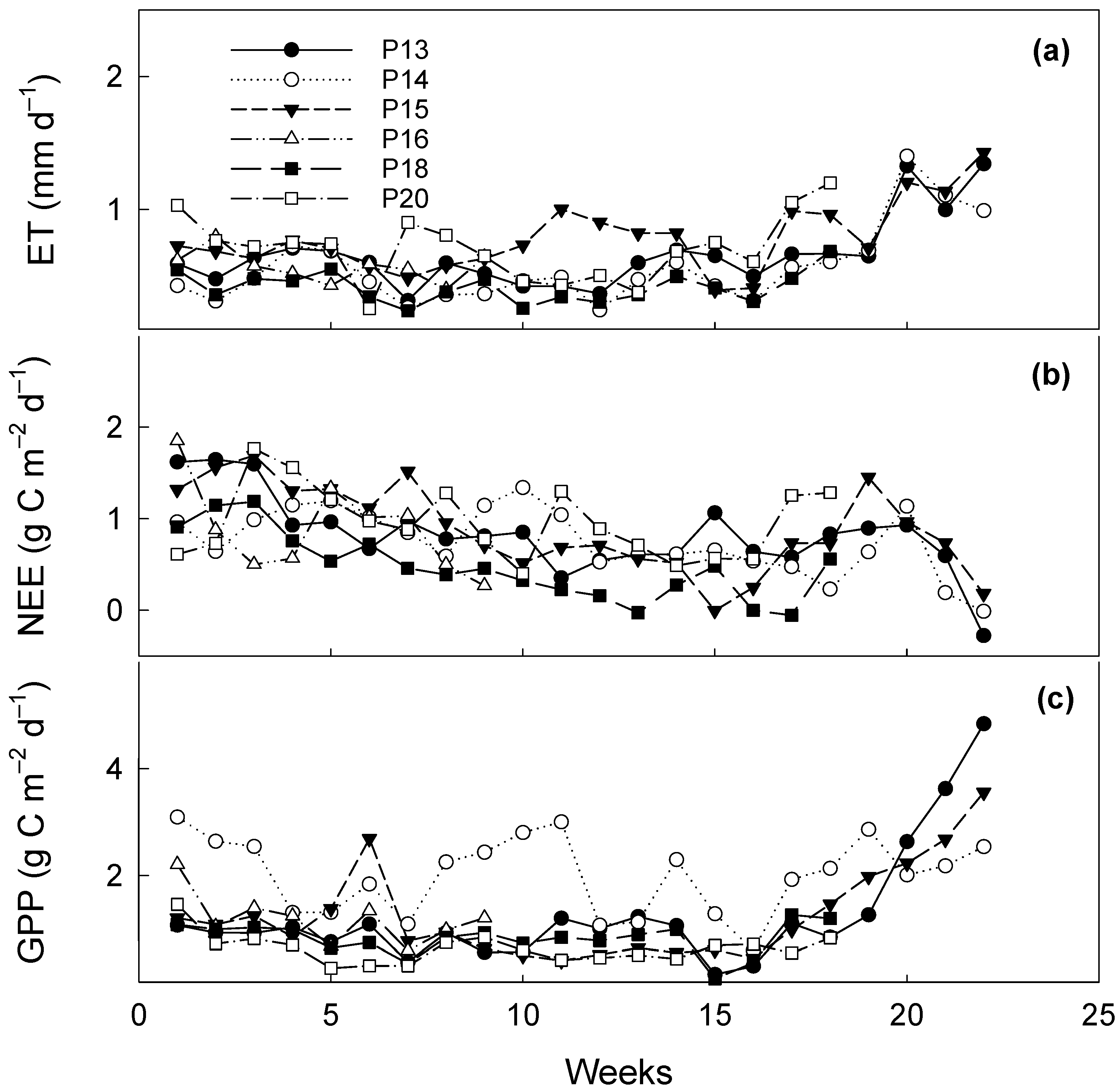
3.4. Interannual Variability in CO2 Fluxes and ET during Dormant Seasons
3.5. Comparison of CO2 Fluxes and ET during Dormant Seasons among Pastures
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Suttie, J.M.; Reynolds, S.G.; Batello, C. Grasslands of the World; Food & Agriculture Organization: Rome, Italy, 2005; Volume 34. [Google Scholar]
- Turner, B.L.; Meyer, W.B. Global land-use and land-cover change: An overview. In Changes in Land Use and Land Cover: A Global Perspective; University of Cambridge: Cambridge, UK, 1994; Volume 4. [Google Scholar]
- Samson, F.; Knopf, F. Prairie Conservation in North America. BioScience 1994, 44, 418–421. [Google Scholar] [CrossRef] [Green Version]
- Briske, D.; Derner, J.; Brown, J.; Fuhlendorf, S.; Teague, W.; Havstad, K.; Gillen, R.; Ash, A.; Willms, W. Rotational Grazing on Rangelands: Reconciliation of Perception and Experimental Evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Owensby, C.E.; Auen, L.M.; Berns, H.F.; Dhuyvetter, K.C. Grazing Systems for Yearling Cattle on Tallgrass Prairie. Rangel. Ecol. Manag. 2008, 61, 204–210. [Google Scholar] [CrossRef]
- Collins, S.L. Interaction of disturbances in tallgrass prairie: A field experiment. Ecology 1987, 68, 1243–1250. [Google Scholar] [CrossRef]
- Knapp, A.K.; Seastedt, T.R. Detritus Accumulation Limits Productivity of Tallgrass Prairie. BioScience 1986, 36, 662–668. [Google Scholar] [CrossRef]
- Anderson, K.L. Time of Burning as It Affects Soil Moisture in an Ordinary Upland Bluestem Prairie in the Flint Hills. Rangel. Ecol. Manag. 1965, 18, 311. [Google Scholar] [CrossRef]
- Wagle, P.; Gowda, P.H.; Northup, B.K.; Starks, P.J.; Neel, J.P. Response of Tallgrass Prairie to Management in the US Southern Great Plains: Site Descriptions, Management Practices, and Eddy Covariance Instrumentation for a Long-Term Experiment. Remote Sens. 2019, 11, 1988. [Google Scholar] [CrossRef] [Green Version]
- Derner, J.D.; Hickman, K.R.; Polley, H.W. Decreasing Precipitation Variability Does Not Elicit Major Aboveground Biomass or Plant Diversity Responses in a Mesic Rangeland. Rangel. Ecol. Manag. 2011, 64, 352–357. [Google Scholar] [CrossRef]
- Baldocchi, D.D. How eddy covariance flux measurements have contributed to our understanding of Global Change Biology. Glob. Chang. Biol. 2020, 26, 242–260. [Google Scholar] [CrossRef]
- Burba, G. Illustrative Maps of Past and Present Eddy Covariance Measurement Locations: I. Early Update; LI-COR Biosciences: Lincoln, NE, USA, 2019. [Google Scholar]
- Bajgain, R.; Xiao, X.; Basara, J.; Wagle, P.; Zhou, Y.; Mahan, H.; Gowda, P.; McCarthy, H.R.; Northup, B.; Neel, J.; et al. Carbon dioxide and water vapor fluxes in winter wheat and tallgrass prairie in central Oklahoma. Sci. Total Environ. 2018, 644, 1511–1524. [Google Scholar] [CrossRef]
- Williams, M.A.; Rice, C.W.; Owensby, C.E. Carbon dynamics and microbial activity in tallgrass prairie exposed to elevated CO2 for 8 years. Plant Soil 2000, 227, 127–137. [Google Scholar] [CrossRef]
- Owensby, C.E.; Ham, J.M.; Auen, L.M. Fluxes of CO2 From Grazed and Ungrazed Tallgrass Prairie. Rangel. Ecol. Manag. 2006, 59, 111–127. [Google Scholar] [CrossRef]
- Suyker, A.E.; Verma, S.B.; Burba, G.G. Interannual variability in net CO2 exchange of a native tallgrass prairie. Glob. Chang. Biol. 2003, 9, 255–265. [Google Scholar] [CrossRef]
- Fischer, M.L.; Torn, M.S.; Billesbach, D.P.; Doyle, G.; Northup, B.; Biraud, S.C. Carbon, water, and heat flux responses to experimental burning and drought in a tallgrass prairie. Agric. For. Meteorol. 2012, 166, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Wagle, P.; Gowda, P.H.; Neel, J.P.; Northup, B.K.; Zhou, Y. Integrating eddy fluxes and remote sensing products in a rotational grazing native tallgrass prairie pasture. Sci. Total Environ. 2020, 712, 136407. [Google Scholar] [CrossRef]
- Natali, S.M.; Watts, J.D.; Rogers, B.M.; Potter, S.; Ludwig, S.M.; Selbmann, A.K.; Sullivan, P.F.; Abbott, B.W.; Arndt, K.A.; Birch, L.; et al. Large loss of CO2 in winter observed across the northern permafrost region. Nat. Clim. Chang. 2019, 9, 852–857. [Google Scholar] [CrossRef]
- Winston, G.; Sundquist, E.T.; Stephens, B.B.; Trumbore, S.E. Winter CO2 fluxes in a boreal forest. J. Geophys. Res. Atmos. 1997, 102, 28795–28804. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.; Song, C.; Wang, X.; Sun, X.; Meng, H.; Sun, L. Greenhouse gas emissions from different wetlands during the snow-covered season in Northeast China. Atmospheric Environ. 2012, 62, 328–335. [Google Scholar] [CrossRef]
- Rafat, A.; Rezanezhad, F.; Quinton, W.L.; Humphreys, E.R.; Webster, K.; Van Cappellen, P. Non-growing season carbon emissions in a northern peatland are projected to increase under global warming. Commun. Earth Environ. 2021, 2, 111. [Google Scholar] [CrossRef]
- McPherson, R.A.; Stensrud, D.J.; Crawford, K.C. The impact of Oklahoma’s winter wheat belt on the mesoscale environment. Mon. Weather Rev. 2004, 132, 405–421. [Google Scholar] [CrossRef]
- Reed, B.C.; Schwartz, M.D.; Xiao, X. Remote sensing phenology. In Phenology of Ecosystem Processes; Springer: Berlin/Heidelberg, Germany, 2009; pp. 231–246. [Google Scholar]
- Wagle, P.; Xiao, X.; Torn, M.S.; Cook, D.R.; Matamala, R.; Fischer, M.L.; Jin, C.; Dong, J.; Biradar, C. Sensitivity of vegetation indices and gross primary production of tallgrass prairie to severe drought. Remote Sens. Environ. 2014, 152, 1–14. [Google Scholar] [CrossRef]
- Zhou, Y.; Xiao, X.; Wagle, P.; Bajgain, R.; Mahan, H.; Basara, J.; Dong, J.; Qin, Y.; Zhang, G.; Luo, Y.; et al. Examining the short-term impacts of diverse management practices on plant phenology and carbon fluxes of Old World bluestems pasture. Agric. For. Meteorol. 2017, 237, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Bajgain, R.; Xiao, X.; Wagle, P.; Basara, J.; Zhou, Y. Sensitivity analysis of vegetation indices to drought over two tallgrass prairie sites. ISPRS J. Photogramm. Remote Sens. 2015, 108, 151–160. [Google Scholar] [CrossRef]
- Wagle, P.; Xiao, X.; Scott, R.L.; Kolb, T.E.; Cook, D.R.; Brunsell, N.; Baldocchi, D.D.; Basara, J.; Matamala, R.; Zhou, Y.; et al. Biophysical controls on carbon and water vapor fluxes across a grassland climatic gradient in the United States. Agric. For. Meteorol. 2015, 214, 293–305. [Google Scholar] [CrossRef] [Green Version]
- Steiner, J.L.; Starks, P.J.; Neel, J.P.; Northup, B.; Turner, K.E.; Gowda, P.; Coleman, S.; Brown, M. Managing Tallgrass Prairies for Productivity and Ecological Function: A Long-Term Grazing Experiment in the Southern Great Plains, USA. Agronomy 2019, 9, 699. [Google Scholar] [CrossRef] [Green Version]
- McPherson, R.A.; Fiebrich, C.A.; Crawford, K.C.; Kilby, J.R.; Grimsley, D.L.; Martinez, J.E.; Basara, J.; Illston, B.G.; Morris, D.A.; Kloesel, K.A.; et al. Statewide Monitoring of the Mesoscale Environment: A Technical Update on the Oklahoma Mesonet. J. Atmos. Ocean. Technol. 2007, 24, 301–321. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.; Noormets, A.; Gavazzi, M.; McNulty, S.; Chen, J.; Domec, J.-C.; King, J.; Amatya, D.; Skaggs, R. Energy and water balance of two contrasting loblolly pine plantations on the lower coastal plain of North Carolina, USA. For. Ecol. Manag. 2010, 259, 1299–1310. [Google Scholar] [CrossRef]
- Zeeman, M.J.; Hiller, R.; Gilgen, A.K.; Michna, P.; Plüss, P.; Buchmann, N.; Eugster, W. Management and climate impacts on net CO2 fluxes and carbon budgets of three grasslands along an elevational gradient in Switzerland. Agric. For. Meteorol. 2010, 150, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Wagle, P.; Kakani, V.G. Growing season variability in evapotranspiration, ecosystem water use efficiency, and energy partitioning in switchgrass. Ecohydrology 2012, 7, 64–72. [Google Scholar] [CrossRef]
- Wagle, P.; Kakani, V.G. Seasonal variability in net ecosystem carbon dioxide exchange over a young Switchgrass stand. GCB Bioenergy 2013, 6, 339–350. [Google Scholar] [CrossRef]
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Šigut, L.; Menzer, O.; Reichstein, M. Basic and extensible post-processing of eddy covariance flux data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On the Temperature Dependence of Soil Respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.V.; van Leeuwen, W. A comparison of vegetation indices over a global set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Wagle, P.; Gowda, P.H.; Northup, B.K.; Turner, K.E.; Neel, J.P.; Manjunatha, P.; Zhou, Y. Variability in carbon dioxide fluxes among six winter wheat paddocks managed under different tillage and grazing practices. Atmos. Environ. 2018, 185, 100–108. [Google Scholar] [CrossRef]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Knapp, A.K. Post-burn differences in solar radiation, leaf temperature and water stress influencing production in a lowland tallgrass prairie. Am. J. Bot. 1984, 71, 220–227. [Google Scholar] [CrossRef]
- Anderson, K.L.; Smith, E.F.; Owensby, C.E. Burning bluestem range. J. Range Manag. 1970, 23, 81–92. [Google Scholar] [CrossRef]
- Hickman, K.R.; Hartnett, D.C.; Cochran, R.C.; Owensby, C.E. Grazing management effects on plant species diversity in tallgrass prairie. Rangel. Ecol. Manag. 2004, 57, 58–65. [Google Scholar] [CrossRef]
- Campioli, M.; Vicca, S.; Luyssaert, S.; Bilcke, J.; Ceschia, E.; Iii, F.S.C.; Ciais, P.; Fernández-Martínez, M.; Malhi, Y.; Obersteiner, M.; et al. Biomass production efficiency controlled by management in temperate and boreal ecosystems. Nat. Geosci. 2015, 8, 843–846. [Google Scholar] [CrossRef]
- Singh, H.; Northup, B.K.; Baath, G.S.; Gowda, P.P.; Kakani, V.G. Greenhouse mitigation strategies for agronomic and grazing lands of the US Southern Great Plains. Mitig. Adapt. Strat. Glob. Chang. 2020, 25, 819–853. [Google Scholar] [CrossRef]
- Ilstedt, U.; Nordgren, A.; Malmer, A. Optimum soil water for soil respiration before and after amendment with glucose in humid tropical acrisols and a boreal mor layer. Soil Biol. Biochem. 2000, 32, 1591–1599. [Google Scholar] [CrossRef]
- Orchared, V. Field and laboratory studies on the relationships between respiration and moisture for tow soils of contrasting fertility status. Pedobiologia 1992, 36, 21–33. [Google Scholar]
- Cook, F.J.; Orchard, V.A.; Corderoy, D.M. Effects of lime and water content on soil respiration. N. Z. J. Agric. Res. 1985, 28, 517–523. [Google Scholar] [CrossRef]
- Austin, A.T.; Yahdjian, L.; Stark, J.M.; Belnap, J.; Porporato, A.; Norton, U.; Ravetta, D.A.; Schaeffer, S. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 2004, 141, 221–235. [Google Scholar] [CrossRef]
- Suyker, A.E.; Verma, S.B. Year-round observations of the net ecosystem exchange of carbon dioxide in a native tallgrass prairie. Glob. Chang. Biol. 2001, 7, 279–289. [Google Scholar] [CrossRef]
- Wagle, P.; Gowda, P.H. Tallgrass Prairie Responses to Management Practices and Disturbances: A Review. Agronomy 2018, 8, 300. [Google Scholar] [CrossRef]
- Collins, S.L.; Knapp, A.K.; Briggs, J.M.; Blair, J.M.; Steinauer, E.M. Modulation of Diversity by Grazing and Mowing in Native Tallgrass Prairie. Science 1998, 280, 745–747. [Google Scholar] [CrossRef]
- Ehleringer, J.; Björkman, O. Quantum yields for CO2 uptake in C3 and C4 plants: Dependence on temperature, CO2, and O2 concentration. Plant Physiol. 1977, 59, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.M.; Hu, F.S.; Tian, J.; Stefanova, I.; Brown, T.A. Response of C3 and C4 plants to middle-Holocene climatic variation near the prairie-forest ecotone of Minnesota. Proc. Natl. Acad. Sci. USA 2004, 101, 562–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Northup, B.K.; Starks, P.J.; Turner, K.E. Stocking Methods and Soil Macronutrient Distributions in Southern Tallgrass Paddocks: Are There Linkages? Agronomy 2019, 9, 281. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ET | NEE | GPP | |
|---|---|---|---|
| November | |||
| 2017 | 18 ± 0.4 | 34 ± 1.34 | 28 |
| 2018 | 18 ± 0.32 | 18 ± 0.81 | 35 |
| 2019 | 15 ± 0.53 | 31 ± 1.32 | 25 |
| 2020 | 14 ± 0.34 | 30 ± 0.89 | 29 |
| Average | 16 | 28 | 29 |
| SD | 2 | 7 | 4 |
| December | |||
| 2017 | 11 ± 0.24 | 11 ± 0.61 | 30 |
| 2018 | 14 ± 0.35 | 16 ± 0.8 | 28 |
| 2019 | 13 ± 0.43 | 16 ± 0.9 | 32 |
| 2020 | 12 ± 0.35 | 16 ± 1.1 | 22 |
| Average | 12 | 15 | 28 |
| SD | 1 | 3 | 5 |
| January | |||
| 2018 | 9 ± 0.31 | 4 ± 0.68 | 31 |
| 2019 | 17 ± 0.37 | 6 ± 0.54 | 18 |
| 2020 | 12 ± 0.47 | 16 ± 0.84 | 22 |
| 2021 | 11 ± 0.35 | 5 ± 0.85 | 26 |
| Average | 12 | 8 | 24 |
| SD | 3 | 5 | 6 |
| February | |||
| 2018 | 12 ± 0.4 | −6 ± 0.99 | 33 |
| 2019 | 15 ± 0.4 | 2 ± 0.59 | 20 |
| 2020 | 19 ± 0.46 | 1 ± 1.04 | 31 |
| 2021 | 12 ± 0.34 | 6 ± 0.76 | 19 |
| Average | 15 | 1 | 26 |
| SD | 3 | 5 | 7 |
| March | |||
| 2018 | 30 ± 0.48 | 19 ± 0.95 | 31 |
| 2019 | 48 ± 0.79 | 15 ± 1 | 24 |
| 2020 | |||
| 2021 | |||
| Average | 39 | 17 | 27 |
| SD | 13 | 3 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagle, P.; Kakani, V.G.; Gowda, P.H.; Xiao, X.; Northup, B.K.; Neel, J.P.S.; Starks, P.J.; Steiner, J.L.; Gunter, S.A. Dormant Season Vegetation Phenology and Eddy Fluxes in Native Tallgrass Prairies of the U.S. Southern Plains. Remote Sens. 2022, 14, 2620. https://doi.org/10.3390/rs14112620
Wagle P, Kakani VG, Gowda PH, Xiao X, Northup BK, Neel JPS, Starks PJ, Steiner JL, Gunter SA. Dormant Season Vegetation Phenology and Eddy Fluxes in Native Tallgrass Prairies of the U.S. Southern Plains. Remote Sensing. 2022; 14(11):2620. https://doi.org/10.3390/rs14112620
Chicago/Turabian StyleWagle, Pradeep, Vijaya G. Kakani, Prasanna H. Gowda, Xiangming Xiao, Brian K. Northup, James P. S. Neel, Patrick J. Starks, Jean L. Steiner, and Stacey A. Gunter. 2022. "Dormant Season Vegetation Phenology and Eddy Fluxes in Native Tallgrass Prairies of the U.S. Southern Plains" Remote Sensing 14, no. 11: 2620. https://doi.org/10.3390/rs14112620
APA StyleWagle, P., Kakani, V. G., Gowda, P. H., Xiao, X., Northup, B. K., Neel, J. P. S., Starks, P. J., Steiner, J. L., & Gunter, S. A. (2022). Dormant Season Vegetation Phenology and Eddy Fluxes in Native Tallgrass Prairies of the U.S. Southern Plains. Remote Sensing, 14(11), 2620. https://doi.org/10.3390/rs14112620