

Using Satellite NDVI Time-Series to Monitor Grazing Effects on Vegetation Productivity and Phenology in Heterogeneous Mediterranean Forests



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Design
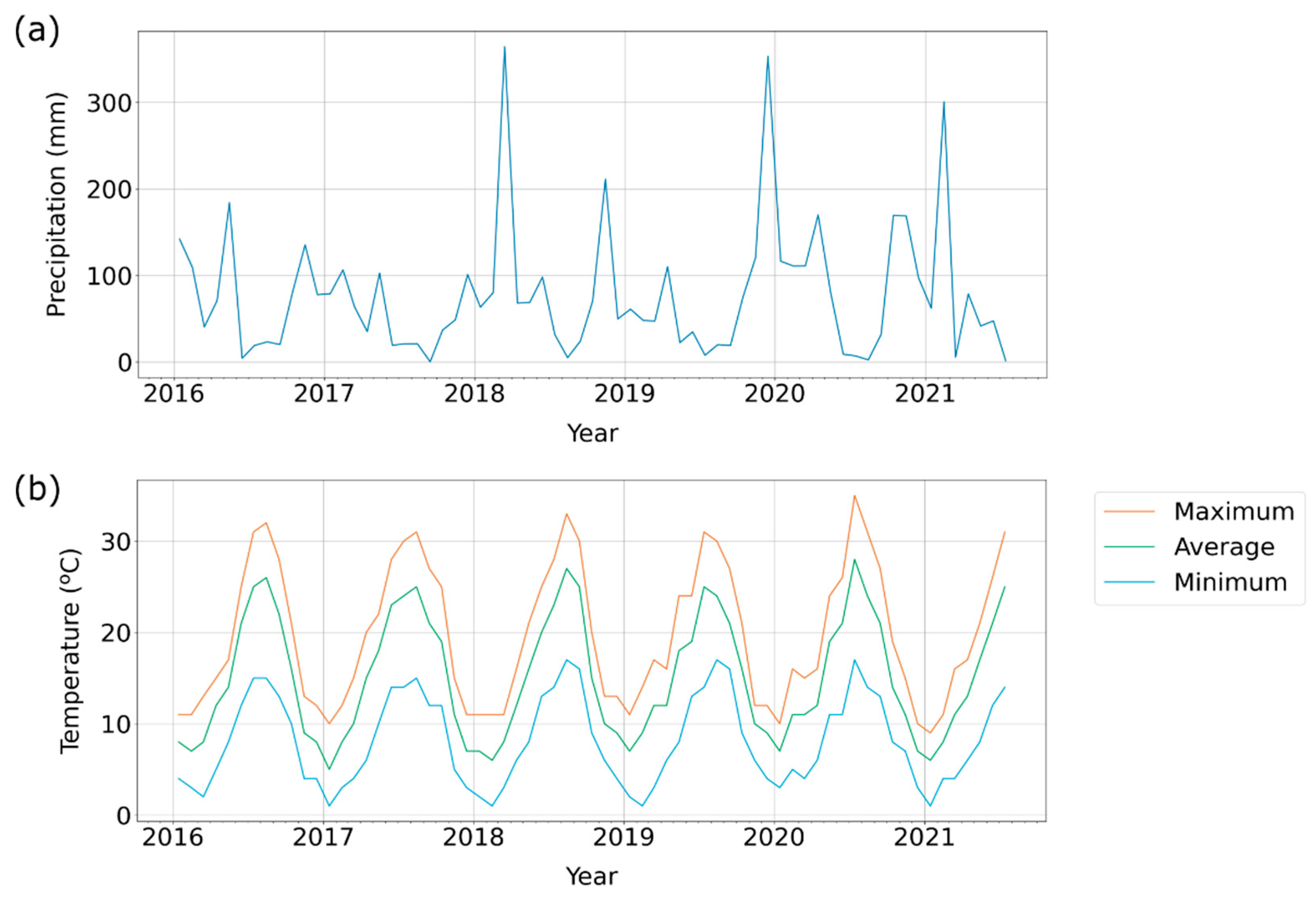
2.3. Satellite Data
2.4. Data Analysis
3. Results
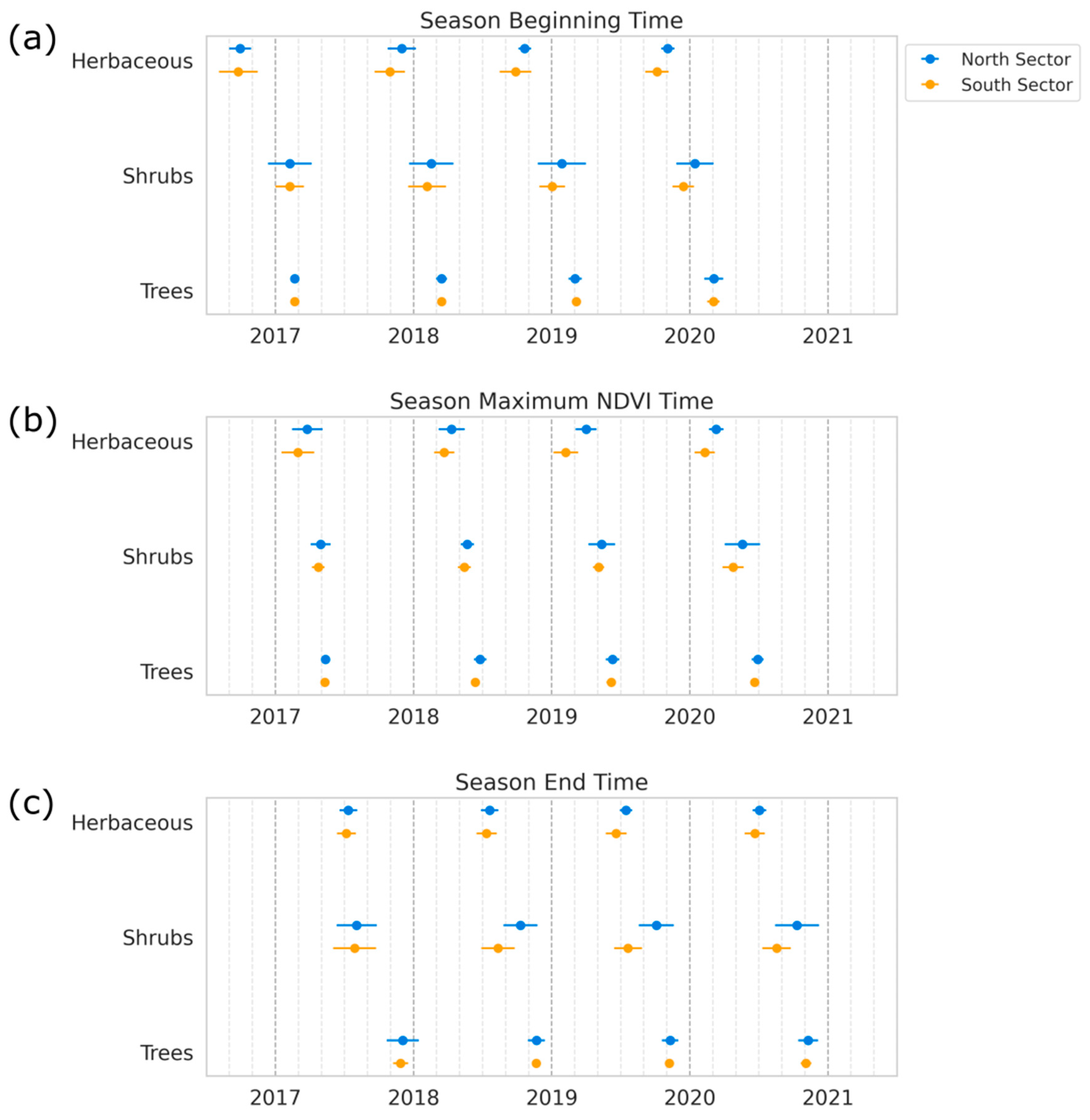
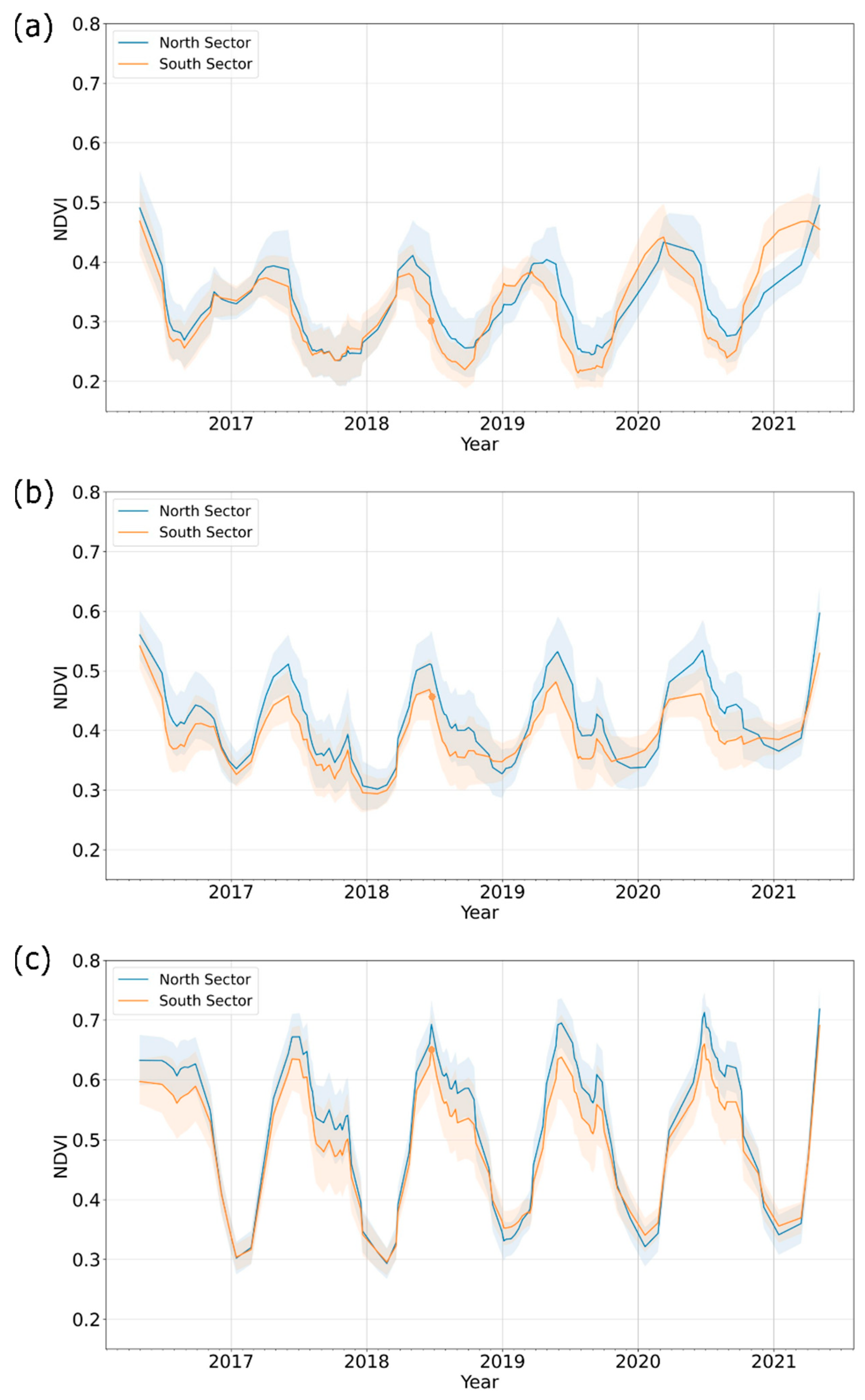
3.1. Phenology Patterns of Vegetation Types
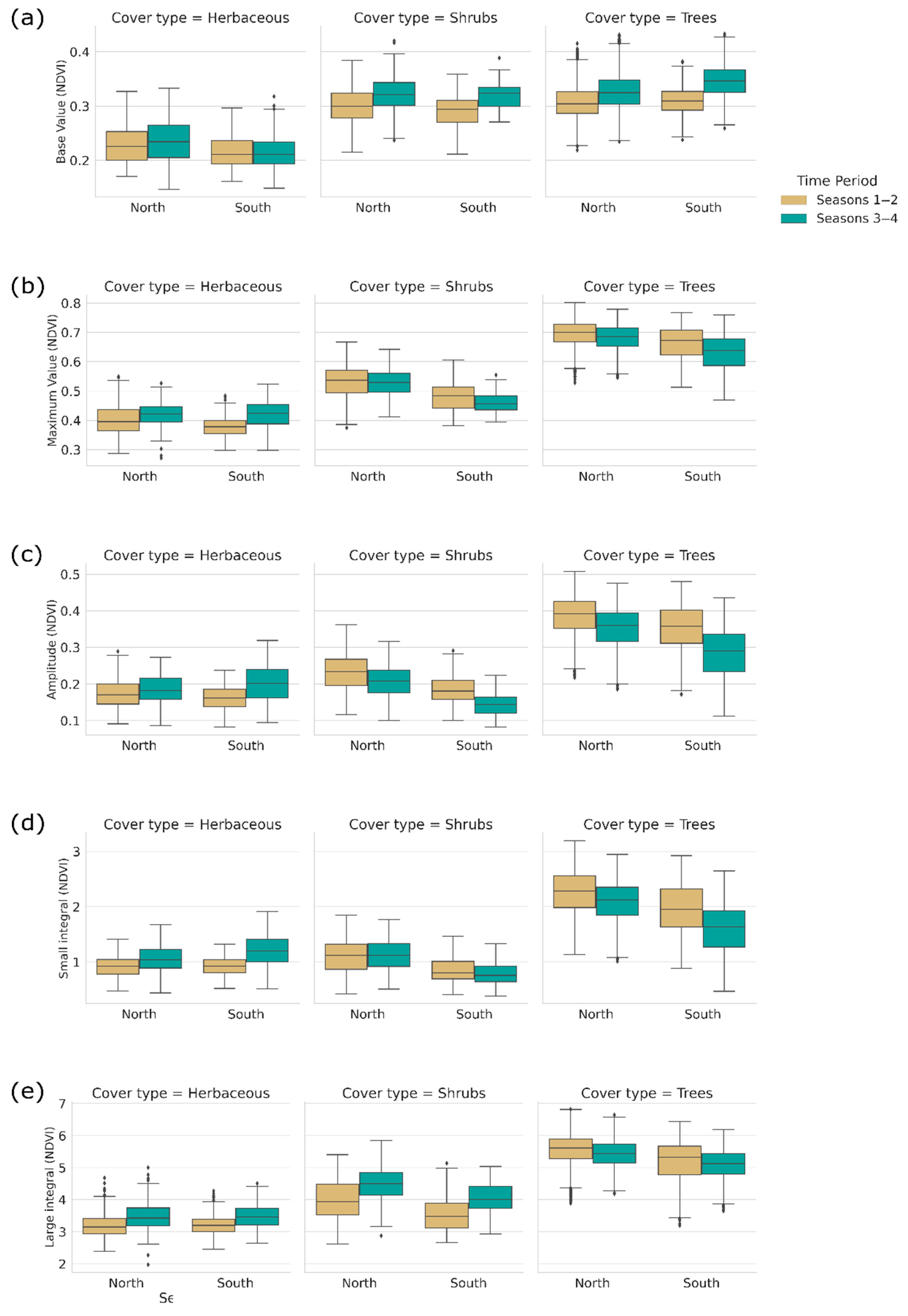
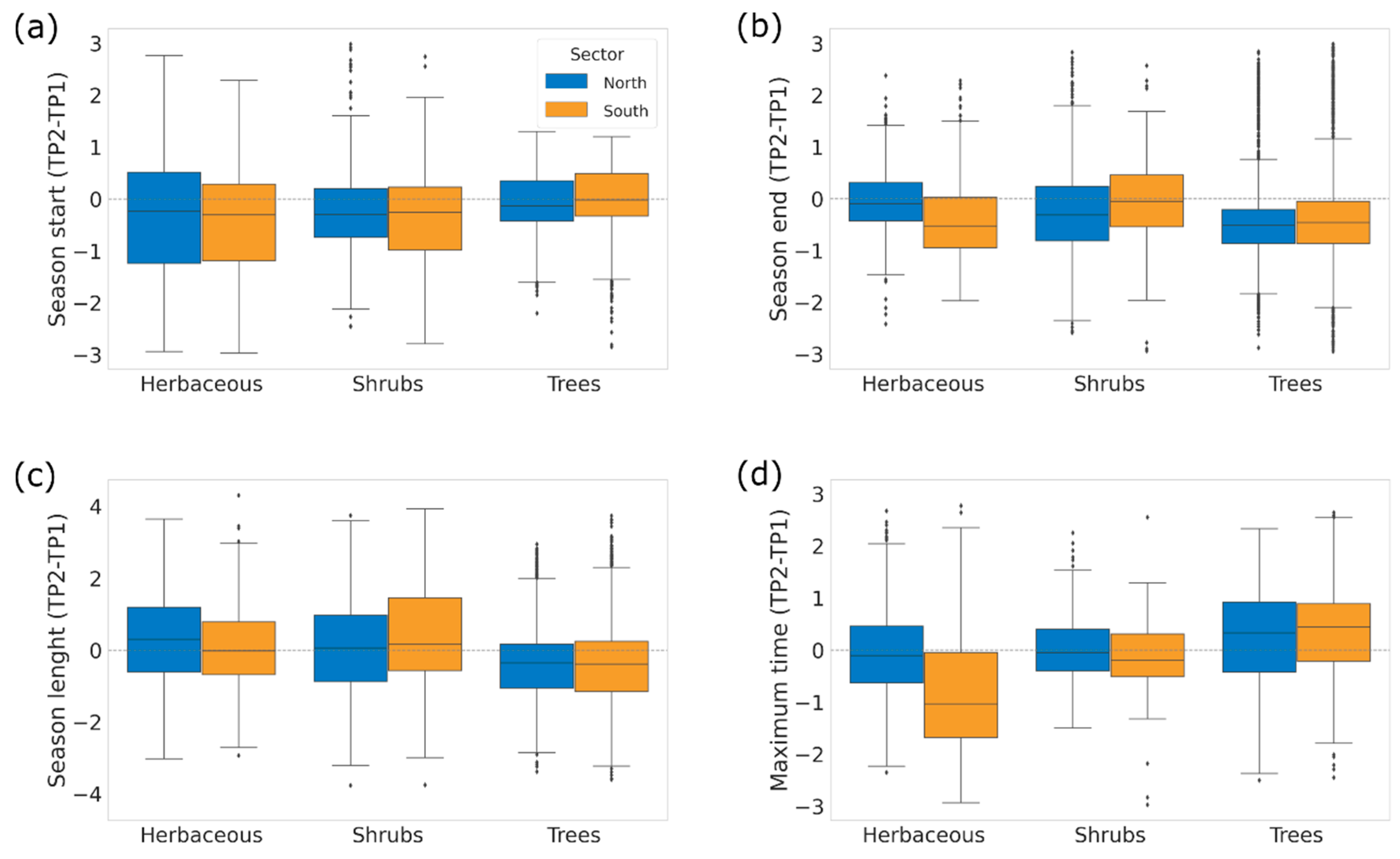
3.2. Changes in Phenology Metrics
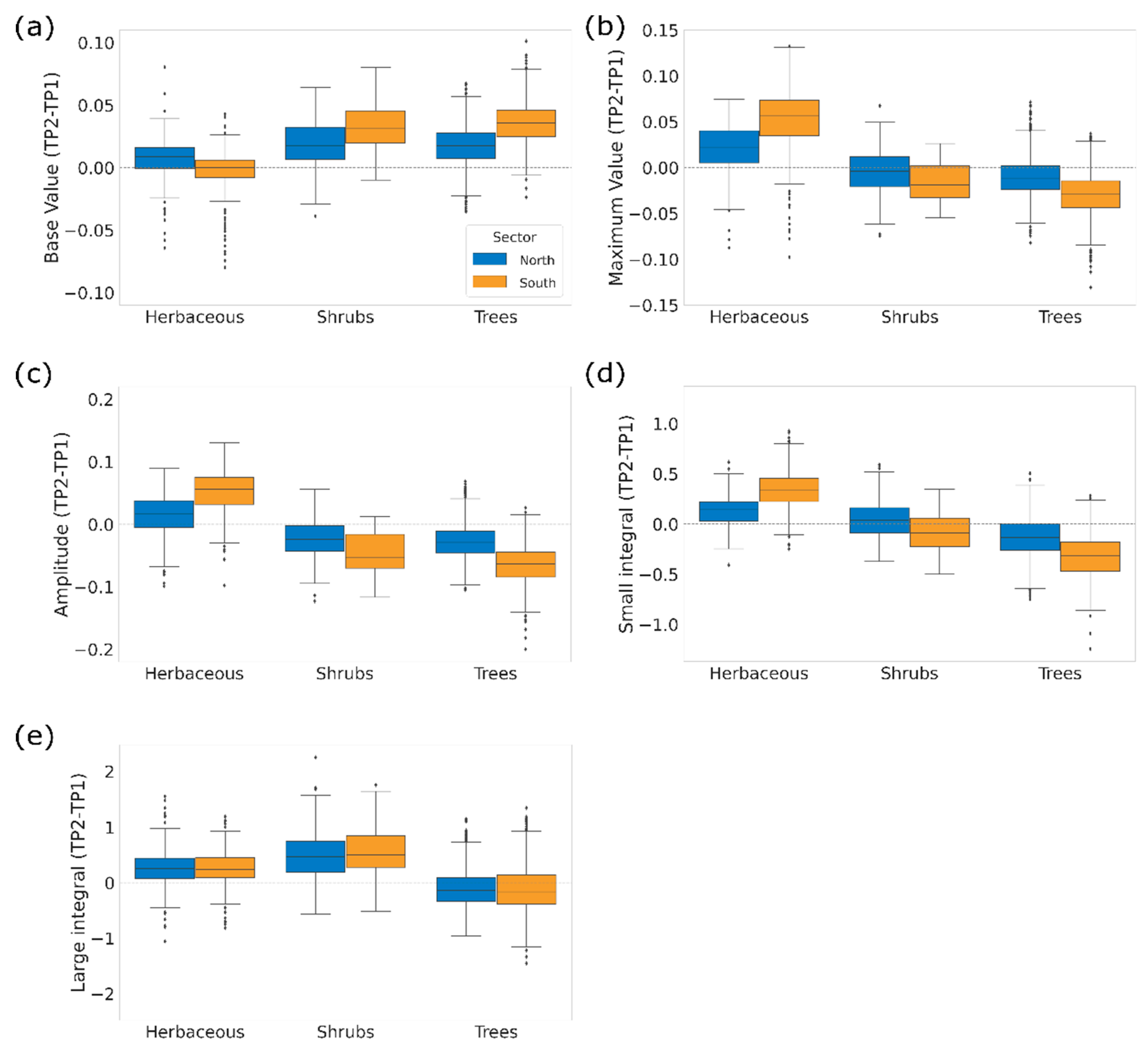
3.3. Changes in Productivity Metrics
4. Discussion
4.1. Vegetation Seasonal Dynamics and Response to Grazing
4.2. Use of NDVI Time-Series to Monitor Grazing
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Ripple, W.J.; Newsome, T.M.; Wolf, C.; Dirzo, R.; Everatt, K.T.; Galetti, M.; Hayward, M.W.; Kerley, G.I.H.; Levi, T.; Lindsey, P.A.; et al. Collapse of the World’s Largest Herbivores. Sci. Adv. 2015, 1, e1400103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, C.; Putman, R. Large Herbivores as ‘Environmental Engineers’. In Ungulate Management in Europe: Problems and Practices; Apollonio, M., Andersen, R., Putman, R., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 260–283. ISBN 978-0-521-76059-1. [Google Scholar]
- Teillard, F.; de Souza, D.M.; Thoma, G.; Gerber, P.J.; Finn, J.A. What Does Life-Cycle Assessment of Agricultural Products Need for More Meaningful Inclusion of Biodiversity? J. Appl. Ecol. 2016, 53, 1422–1429. [Google Scholar] [CrossRef] [Green Version]
- Bignal, E.; McCracken, D. The Conservation Value of European Traditional Farming Systems. Environ. Rev. 2000, 8, 149–171. [Google Scholar] [CrossRef]
- Navarro, L.M.; Proença, V.; Kaplan, J.O.; Pereira, H.M. Maintaining Disturbance-Dependent Habitats. In Rewilding European Landscapes; Pereira, H.M., Navarro, L.M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 143–167. ISBN 978-3-319-12039-3. [Google Scholar]
- Fernández-Manjarrés, J.F.; Ruiz-Benito, P.; Zavala, M.A.; Camarero, J.J.; Pulido, F.; Proença, V.; Navarro, L.; Sansilvestri, R.; Granda, E.; Marqués, L.; et al. Forest Adaptation to Climate Change along Steep Ecological Gradients: The Case of the Mediterranean-Temperate Transition in South-Western Europe. Sustainability 2018, 10, 3065. [Google Scholar] [CrossRef] [Green Version]
- Honrado, J.P.; Lomba, A.; Alves, P.J.M.; Aguiar, C.; Monteiro-Henriques, T.; Cerqueira, Y.; Monteiro, P.; Caldas, F.B. Conservation Management of EU Priority Habitats after Collapse of Traditional Pastoralism: Navigating Socioecological Transitions in Mountain Rangeland. Rural Sociol. 2017, 82, 101–128. [Google Scholar] [CrossRef] [Green Version]
- Lomba, A.; Moreira, F.; Klimek, S.; Jongman, R.H.; Sullivan, C.; Moran, J.; Poux, X.; Honrado, J.P.; Pinto-Correia, T.; Plieninger, T.; et al. Back to the Future: Rethinking Socioecological Systems Underlying High Nature Value Farmlands. Front. Ecol. Environ. 2020, 18, 36–42. [Google Scholar] [CrossRef]
- Navarro, L.M.; Marques, A.; Proença, V.; Ceauşu, S.; Gonçalves, B.; Capinha, C.; Fernandez, M.; Geldmann, J.; Pereira, H.M. Restoring Degraded Land: Contributing to Aichi Targets 14, 15, and Beyond. Curr. Opin. Environ. Sustain. 2017, 29, 207–214. [Google Scholar] [CrossRef]
- Gordon, I.J.; Manning, A.D.; Navarro, L.M.; Rouet-Leduc, J. Domestic Livestock and Rewilding: Are They Mutually Exclusive? Front. Sustain. Food Syst. 2021, 5, 550410. [Google Scholar] [CrossRef]
- Ruiz-Mirazo, J.; Robles, A.; González-Rebollar, J. Two-Year Evaluation of Fuelbreaks Grazed by Livestock in the Wildfire Prevention Program in Andalusia (Spain). Agric. Ecosyst. Environ. 2011, 141, 13–22. [Google Scholar] [CrossRef]
- Vermeulen, R. (Ed.) Natural Grazing—Practices in the Rewilding of Cattle and Horses; Rewilding Europe: Nijmegen, The Netherlands, 2015; p. 40. [Google Scholar]
- Franzluebbers, A.J.; Paine, L.K.; Winsten, J.R.; Krome, M.; Sanderson, M.A.; Ogles, K.; Thompson, D. Well-Managed Grazing Systems: A Forgotten Hero of Conservation. J. Soil Water Conserv. 2012, 67, 100A–104A. [Google Scholar] [CrossRef] [Green Version]
- Lunt, I.D.; Eldridge, D.J.; Morgan, J.W.; Witt, G.B.; Lunt, I.D.; Eldridge, D.J.; Morgan, J.W.; Witt, G.B. A Framework to Predict the Effects of Livestock Grazing and Grazing Exclusion on Conservation Values in Natural Ecosystems in Australia. Aust. J. Bot. 2007, 55, 401–415. [Google Scholar] [CrossRef]
- Kristensen, J.A.; Svenning, J.-C.; Georgiou, K.; Malhi, Y. Can Large Herbivores Enhance Ecosystem Carbon Persistence? Trends Ecol. Evol. 2022, 37, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Papanastasis, V. Restoration of Degraded Grazing Lands through Grazing Management: Can It Work? Restor. Ecol. 2009, 17, 441–445. [Google Scholar] [CrossRef]
- Giralt-Rueda, J.M.; Santamaria, L. Complementary Differences in Primary Production and Phenology among Vegetation Types Increase Ecosystem Resilience to Climate Change and Grazing Pressure in an Iconic Mediterranean Ecosystem. Remote Sens. 2021, 13, 3920. [Google Scholar] [CrossRef]
- Sánchez-Cuesta, R.; Ruiz-Gómez, F.J.; Duque-Lazo, J.; González-Moreno, P.; Navarro-Cerrillo, R.M. The Environmental Drivers Influencing Spatio-Temporal Dynamics of Oak Defoliation and Mortality in Dehesas of Southern Spain. For. Ecol. Manag. 2021, 485, 118946. [Google Scholar] [CrossRef]
- von Keyserlingk, J.; de Hoop, M.; Mayor, A.G.; Dekker, S.C.; Rietkerk, M.; Foerster, S. Resilience of Vegetation to Drought: Studying the Effect of Grazing in a Mediterranean Rangeland Using Satellite Time Series. Remote Sens. Environ. 2021, 255, 112270. [Google Scholar] [CrossRef]
- Bégué, A.; Arvor, D.; Lelong, C.; Vintrou, E.; Simoes, M. Agricultural Systems Studies Using Remote Sensing. In Remote Sensing Handbook, Vol. 2: Land Resources Monitoring, Modeling, and Mapping with Remote Sensing; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Skidmore, A. Remote Sensing of EBVs for Global Change Monitoring. In Proceedings of the Global Change and Biodiversity: Integrating Mechanisms of Interactions, Feedbacks and Scale: URPP Global Change and Biodiversity Conference: Program and Abstract Book, Ascona, Switzerland, 28 August–1 September 2016. [Google Scholar]
- Tian, X.; Minunno, F.; Schiestl-Aalto, P.; Chi, J.; Zhao, P.; Peichl, M.; Marshall, J.; Näsholm, T.; Lim, H.; Peltoniemi, M.; et al. Disaggregating the Effects of Nitrogen Addition on Gross Primary Production in a Boreal Scots Pine Forest. Agric. For. Meteorol. 2021, 301–302, 108337. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the Satellite-Derived NDVI to Assess Ecological Responses to Environmental Change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. TIMESAT—A Program for Analyzing Time-Series of Satellite Sensor Data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.L.; Hoffman, M.T.; Roberts, W. Long-term trends in vegetation phenology and productivity over Namaqualand using the GIMMS AVHRR NDVI3g data from 1982 to 2011. S. Afr. J. Bot. 2017, 111, 76–85. [Google Scholar] [CrossRef]
- Knapp, A.K.; Hoover, D.L.; Blair, J.M.; Buis, G.; Burkepile, D.E.; Chamberlain, A.; Collins, S.L.; Fynn, R.W.S.; Kirkman, K.P.; Smith, M.D.; et al. A Test of Two Mechanisms Proposed to Optimize Grassland Aboveground Primary Productivity in Response to Grazing. J. Plant Ecol. 2012, 5, 357–365. [Google Scholar] [CrossRef] [Green Version]
- Papoutsa, C.; Kouhartsiouk, D.; Themistocleous, K.; Christoforou, M.; Hadjimitsis, D.G. Monitoring of Land Degradation from Overgrazing Using Space-Borne Radar and Optical Imagery: A Case Study in Randi Forest, Cyprus; Michel, U., Schulz, K., Ehlers, M., Nikolakopoulos, K.G., Civco, D., Eds.; SPIE: Edinburgh, UK, 2016; p. 1000516. [Google Scholar]
- Charles, G.K.; Porensky, L.M.; Riginos, C.; Veblen, K.E.; Young, T.P. Herbivore Effects on Productivity Vary by Guild: Cattle Increase Mean Productivity While Wildlife Reduce Variability. Ecol. Appl. 2017, 27, 143–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, A.P.; Amano, T.; Martin, P.A.; Shackelford, G.E.; Simmons, B.I.; Sutherland, W.J. Simple Study Designs in Ecology Produce Inaccurate Estimates of Biodiversity Responses. J. Appl. Ecol. 2019, 56, 2742–2754. [Google Scholar] [CrossRef] [Green Version]
- Ockendon, N.; Amano, T.; Cadotte, M.; Downey, H.; Hancock, M.H.; Thornton, A.; Tinsley-Marshall, P.; Sutherland, W.J. Effectively Integrating Experiments into Conservation Practice. Ecol. Solut. Evid. 2021, 2, e12069. [Google Scholar] [CrossRef]
- D’Odorico, P.; Gonsamo, A.; Damm, A.; Schaepman, M.E. Experimental Evaluation of Sentinel-2 Spectral Response Functions for NDVI Time-Series Continuity. IEEE Trans. Geosci. Remote Sens. 2013, 51, 1336–1348. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and Photographic Infrared Linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Huete, A.R.; Chen, J.; Chen, Y.; Li, J.; Yan, G.; Zhang, X. Analysis of NDVI and Scaled Difference Vegetation Index Retrievals of Vegetation Fraction. Remote Sens. Environ. 2006, 101, 366–378. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.-Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; Granier, A.; et al. Evaluation of the Potential of MODIS Satellite Data to Predict Vegetation Phenology in Different Biomes: An Investigation Using Ground-Based NDVI Measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Pan, Z.; Huang, J.; Zhou, Q.; Wang, L.; Cheng, Y.; Zhang, H.; Blackburn, G.A.; Yan, J.; Liu, J. Mapping Crop Phenology Using NDVI Time-Series Derived from HJ-1 A/B Data. Int. J. Appl. Earth Obs. Geoinf. 2015, 34, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.; Schaepman-Strub, G.; Ecker, K. Predicting Habitat Quality of Protected Dry Grasslands Using Landsat NDVI Phenology. Ecol. Indic. 2018, 91, 447–460. [Google Scholar] [CrossRef]
- Lin, M.; Lucas, H.C.; Shmueli, G. Research Commentary—Too Big to Fail: Large Samples and the p-Value Problem. Inf. Syst. Res. 2013, 24, 906–917. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, G.M.; Feinn, R. Using Effect Size—or Why the P Value Is Not Enough. J. Grad. Med. Educ. 2012, 4, 279–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cliff, N. Answering Ordinal Questions with Ordinal Data Using Ordinal Statistics. Multivar. Behav. Res. 1996, 31, 331–350. [Google Scholar] [CrossRef] [PubMed]
- Romano, J.; Kromrey, J. Appropriate Statistics for Ordinal Level Data: Should We Really Be Using t-Test and Cohen’s d for Evaluating Group Differences on the NSSE and Other Surveys? In Proceedings of the Annual Meeting of the Florida Association of Institutional Research; Florida Association for Institutional Research: Cocoa Beach, FL, USA, 2006. [Google Scholar]
- Alcaraz-Segura, D.; Cabello, J.; Paruelo, J. Baseline Characterization of Major Iberian Vegetation Types Based on the NDVI Dynamics. Plant Ecol. 2009, 202, 13–29. [Google Scholar] [CrossRef]
- Olofsson, P.; Foody, G.M.; Herold, M.; Stehman, S.V.; Woodcock, C.E.; Wulder, M.A. Good practices for estimating area and assessing accuracy of land change. Remote Sens. Environ. 2014, 148, 42–57. [Google Scholar] [CrossRef]
- Kuemmerle, T.; Röder, A.; Hill, J. Separating Grassland and Shrub Vegetation by Multidate Pixel-adaptive Spectral Mixture Analysis. Int. J. Remote Sens. 2006, 27, 3251–3271. [Google Scholar] [CrossRef]
- Walker, J.J.; Soulard, C.E. Phenology Patterns Indicate Recovery Trajectories of Ponderosa Pine Forests After High-Severity Fires. Remote Sens. 2019, 11, 2782. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, I.; Proença, V.; Serra, P.; Palma, J.; Domingo-Marimon, C.; Pons, X.; Domingos, T. Remotely Sensed Indicators and Open-Access Biodiversity Data to Assess Bird Diversity Patterns in Mediterranean Rural Landscapes. Sci. Rep. 2019, 9, 6826. [Google Scholar] [CrossRef] [Green Version]
- Forbes, E.S.; Cushman, J.H.; Burkepile, D.E.; Young, T.P.; Klope, M.; Young, H.S. Synthesizing the effects of large, wild herbivore exclusion on ecosystem function. Funct. Ecol. 2019, 33, 1597–1610. [Google Scholar] [CrossRef] [Green Version]
- Campana, S.; Tognetti, P.M.; Yahdjian, L. Livestock exclusion reduces the temporal stability of grassland productivity regardless of eutrophication. Sci. Total Environ. 2022, 817, 152707. [Google Scholar] [CrossRef]
- Helman, D.; Lensky, I.M.; Tessler, N.; Osem, Y. A phenology-based method for monitoring woody and herbaceous vegetation in Mediterranean forests from NDVI time series. Remote Sens. 2015, 7, 12314–12335. [Google Scholar] [CrossRef] [Green Version]
- Nagendra, H.; Lucas, R.; Honrado, J.P.; Jongman, R.H.; Tarantino, C.; Adamo, M.; Mairota, P. Remote sensing for conservation monitoring: Assessing protected areas, habitat extent, habitat condition, species diversity, and threats. Ecol. Indic. 2013, 33, 45–59. [Google Scholar] [CrossRef]
- Pettorelli, N.; Wegmann, M.; Skidmore, A.; Mucher, S.; Dawson, T.P.; Fernandez, M.; Lucas, R.; Schaepman, M.E.; Wang, T.; O’Connor, B.; et al. Framing the concept of satellite remote sensing essential biodiversity variables: Challenges and future directions. Remote Sens. Ecol. Conserv. 2016, 2, 122–131. [Google Scholar] [CrossRef]
- Chaves, M.E.D.; Picoli, M.C.A.; Sanches, I.D. Recent Applications of Landsat 8/OLI and Sentinel-2/MSI for Land Use and Land. Cover Mapping: A Systematic Review. Remote Sens. 2020, 12, 3062. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
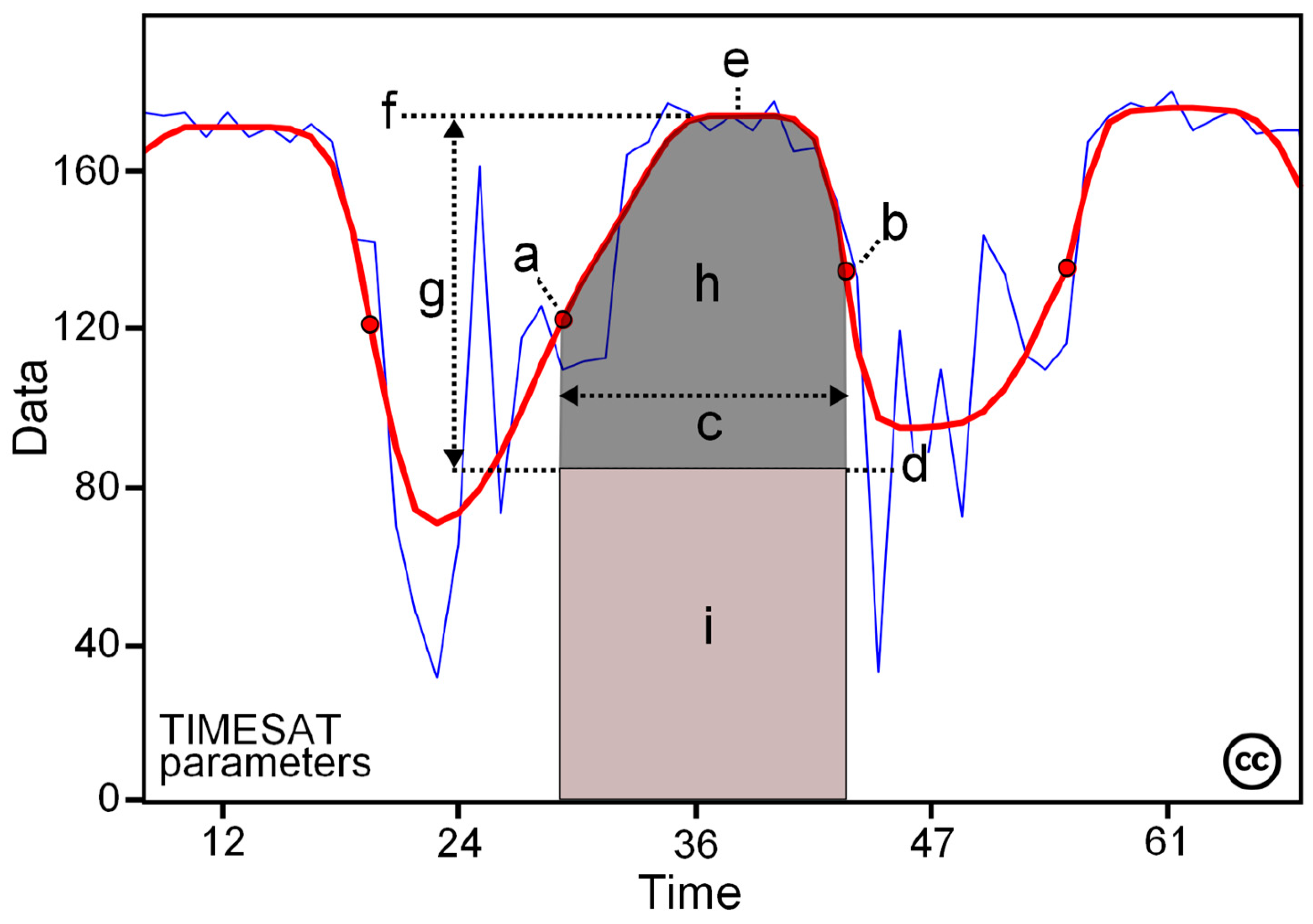
| Metric Definition | |
|---|---|
| Phenology metrics | |
| Season start | Point in time when NDVI reaches 20% of seasonal amplitude as measured from the left minimum level. It corresponds to the start of the growing season. |
| Season end | Point in time when NDVI reaches 20% of seasonal amplitude as measured from the right minimum level. It corresponds to the end of the growing season. |
| Season length | Length of time from the start to the end of the season. It corresponds to the duration of the growing season |
| Time of maximum NDVI | Date of the seasonal NDVI peak. |
| Productivity metrics | |
| Base value | Average of the left and right minimum values (between seasons). |
| Maximum value | Largest NDVI value for the fitted function during the season. A proxy of the maximum amount of vegetation for the sample point. |
| Seasonal amplitude | Difference between the maximum and base NDVI values for the season. |
| Small integral | Integral of the difference between the function describing the season and the base level from season start to end. Used as proxy for seasonal productivity between season start and season end. |
| Large integral | Integral of the function describing the season from start to end. Same as the small integral but with a cumulative start at zero NDVI. Used as a proxy for total productivity, including existing vegetation produced outside of growing season. |
| Zone | Season | S Start | S End | S Len | Max. T | Base V | Max V | Amp | S Int | L Int |
|---|---|---|---|---|---|---|---|---|---|---|
| Herb. North | 1–2 | 10.91 (2.39) | 7.46 (1.73) | 8.41 (1.06) | 3.91 (2.13) | 0.23 (0.03) | 0.40 (0.05) | 0.17 (0.04) | 0.93 (0.18) | 3.19 (0.43) |
| 3–4 | 10.88 (2.21) | 7.27 (1.94) | 8.42 (1.07) | 3.64 (2.01) | 0.24 (0.04) | 0.43 (0.04) | 0.19 (0.04) | 1.04 (0.23) | 3.54 (0.51) | |
| Herb. South | 1–2 | 10.40 (1.87) | 7.21 (1.71) | 8.56 (0.84) | 3.21 (2.15) | 0.22 (0.03) | 0.38 (0.04) | 0.16 (0.03) | 0.91 (0.16) | 3.24 (0.35) |
| 3–4 | 10.04 (1.44) | 6.60 (1.48) | 8.53 (0.57) | 2.25 (1.65) | 0.21 (0.03) | 0.42 (0.04) | 0.20 (0.05) | 1.16 (0.25) | 3.47 (0.32) | |
| Shrubs North | 1–2 | 2.43 (1.99) | 9.27 (2.58) | 7.11 (1.40) | 5.34 (1.36) | 0.30 (0.03) | 0.53 (0.05) | 0.23 (0.05) | 1.06 (0.28) | 3.83 (0.67) |
| 3–4 | 1.68 (1.92) | 10.26 (2.16) | 8.50 (0.84) | 5.44 (1.62) | 0.32 (0.03) | 0.53 (0.04) | 0.20 (0.04) | 1.09 (0.27) | 4.48 (0.52) | |
| Shrubs South | 1–2 | 2.34 (2.66) | 8.24 (2.61) | 6.93 (1.75) | 5.12 (1.51) | 0.29 (0.03) | 0.48 (0.05) | 0.18 (0.04) | 0.82 (0.19) | 3.48 (0.71) |
| 3–4 | 12.83 (2.70) | 8.47 (2.65) | 8.51 (1.21) | 4.96 (2.12) | 0.32 (0.03) | 0.46 (0.03) | 0.15 (0.03) | 0.79 (0.19) | 4.12 (0.56) | |
| Trees North | 1–2 | 3.05 (1.16) | 11.86 (1.47) | 8.83 (0.43) | 6.07 (1.25) | 0.31 (0.03) | 0.70 (0.04) | 0.39 (0.05) | 2.25 (0.36) | 5.60 (0.43) |
| 3–4 | 3.06 (1.45) | 11.28 (1.41) | 8.34 (0.46) | 6.61 (1.37) | 0.32 (0.03) | 0.68 (0.04) | 0.36 (0.05) | 2.11 (0.33) | 5.46 (0.40) | |
| Trees South | 1–2 | 3.06 (1.24) | 11.78 (1.93) | 8.73 (0.82) | 5.84 (1.30) | 0.31 (0.02) | 0.67 (0.05) | 0.35 (0.05) | 1.92 (0.42) | 5.34 (0.59) |
| 3–4 | 3.11 (1.64) | 11.15 (1.71) | 8.21 (0.56) | 6.40 (1.51) | 0.35 (0.03) | 0.63 (0.06) | 0.28 (0.03) | 1.59 (0.41) | 5.13 (0.45) |
| North Sector | Herbaceous | Shrubs | Trees |
|---|---|---|---|
| Phenology metrics | |||
| Season start | negligible | −0.47 *** | negligible |
| Season end | −0.25 ** | negligible | −0.66 *** |
| Season length | n.s. | 0.43 *** | −0.56 *** |
| Max. Time | −0.23 *** | 0.27 *** | 0.80 *** |
| Productivity metrics | |||
| Base Value | n.s. | 0.35 *** | 0.34 *** |
| Max Value | 0.26 *** | n.s. | −0.19 *** |
| Amplitude | 0.16 *** | −0.29 *** | −0.34 *** |
| Small int. | 0.32 *** | n.s. | −0.25 *** |
| Large int. | 0.40 *** | 0.45 *** | −0.18 *** |
| South Sector | Herbaceous | Shrubs | Trees |
|---|---|---|---|
| Phenology metrics | |||
| Season start | −0.2 *** | −0.40 *** | n.s. |
| Season end | −0.59 *** | negligible | −0.39 *** |
| Season length | negligible | 0.41 ** | −0.37 *** |
| Max. Time | −0.65 *** | negligible | 0.70 *** |
| Productivity metrics | |||
| Base Value | n.s. | 0.47 *** | 0.65 *** |
| Max Value | 0.50 *** | −0.22 *** | −0.32 *** |
| Amplitude | 0.27 *** | −0.60 *** | −0.55 *** |
| Small int. | 0.60 *** | negligible | −0.41 *** |
| Large int. | 0.40 *** | 0.49 *** | negligible |
| Herbaceous | Shrubs | Trees | |
|---|---|---|---|
| Phenology metrics | |||
| Season start | negligible | n.s. | negligible |
| Season end | 0.45 *** | n.s. | negligible |
| Season length | −0.18 *** | n.s. | negligible |
| Max. Time | 0.74 *** | 0.27 *** | negligible |
| Productivity metrics | |||
| Base Value | −0.20 *** | 0.33 *** | 0.58 *** |
| Max Value | 0.61 *** | 0.16 ** | 0.44 *** |
| Amplitude | 0.55 *** | 0.32 *** | 0.63 *** |
| Small int. | 0.60 *** | negligible | 0.46 *** |
| Large int. | n.s. | n.s. | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balata, D.; Gama, I.; Domingos, T.; Proença, V. Using Satellite NDVI Time-Series to Monitor Grazing Effects on Vegetation Productivity and Phenology in Heterogeneous Mediterranean Forests. Remote Sens. 2022, 14, 2322. https://doi.org/10.3390/rs14102322
Balata D, Gama I, Domingos T, Proença V. Using Satellite NDVI Time-Series to Monitor Grazing Effects on Vegetation Productivity and Phenology in Heterogeneous Mediterranean Forests. Remote Sensing. 2022; 14(10):2322. https://doi.org/10.3390/rs14102322
Chicago/Turabian StyleBalata, Duarte, Ivo Gama, Tiago Domingos, and Vânia Proença. 2022. "Using Satellite NDVI Time-Series to Monitor Grazing Effects on Vegetation Productivity and Phenology in Heterogeneous Mediterranean Forests" Remote Sensing 14, no. 10: 2322. https://doi.org/10.3390/rs14102322
APA StyleBalata, D., Gama, I., Domingos, T., & Proença, V. (2022). Using Satellite NDVI Time-Series to Monitor Grazing Effects on Vegetation Productivity and Phenology in Heterogeneous Mediterranean Forests. Remote Sensing, 14(10), 2322. https://doi.org/10.3390/rs14102322