
Dynamic Microclimate Boundaries across a Sharp Tropical Rainforest–Clearing Edge

 ,
,
Abstract
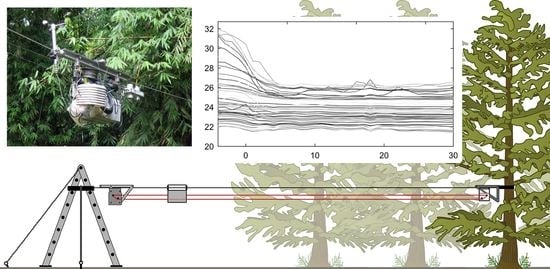
1. Introduction
2. Materials and Methods
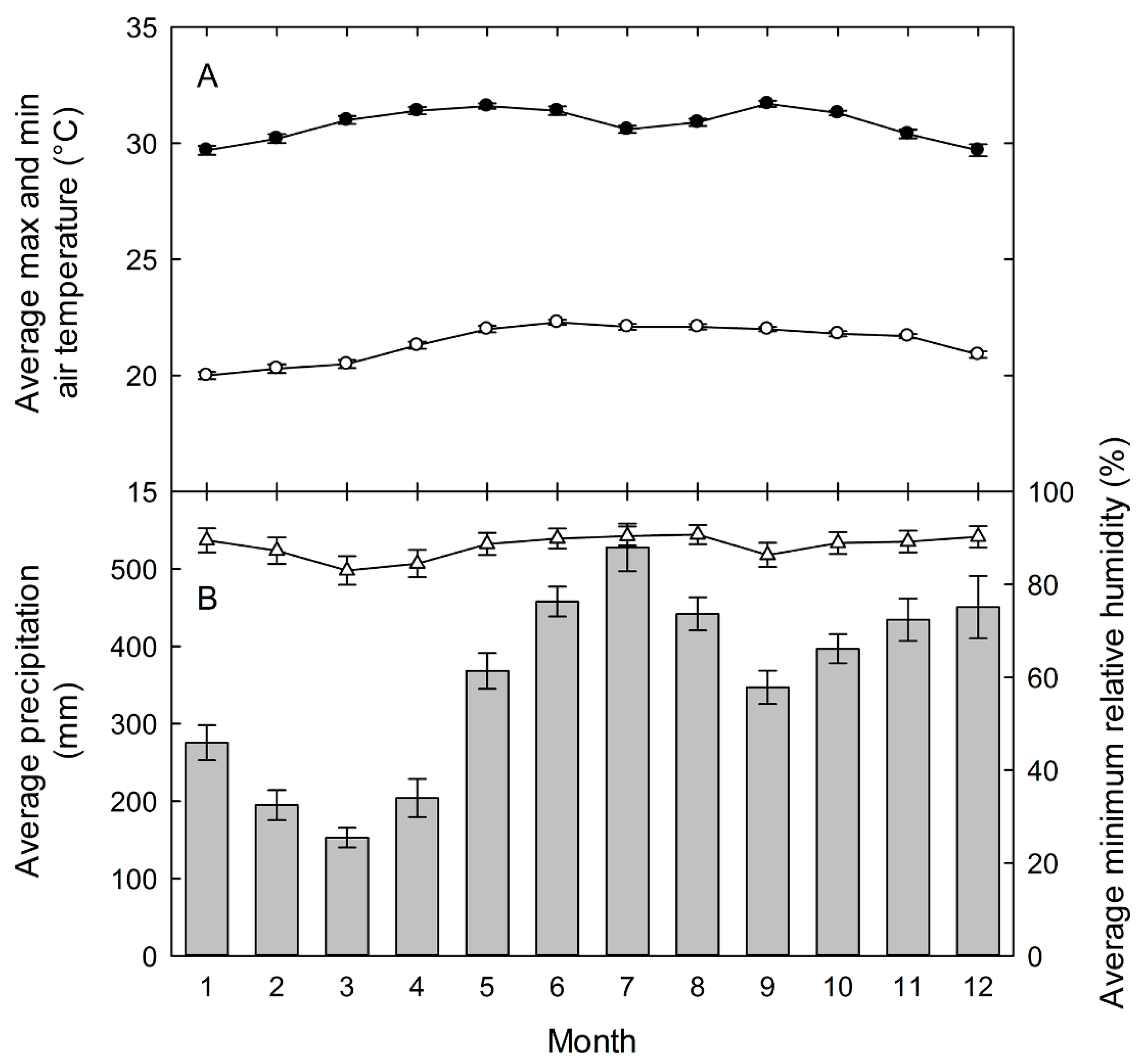
2.1. Field Site
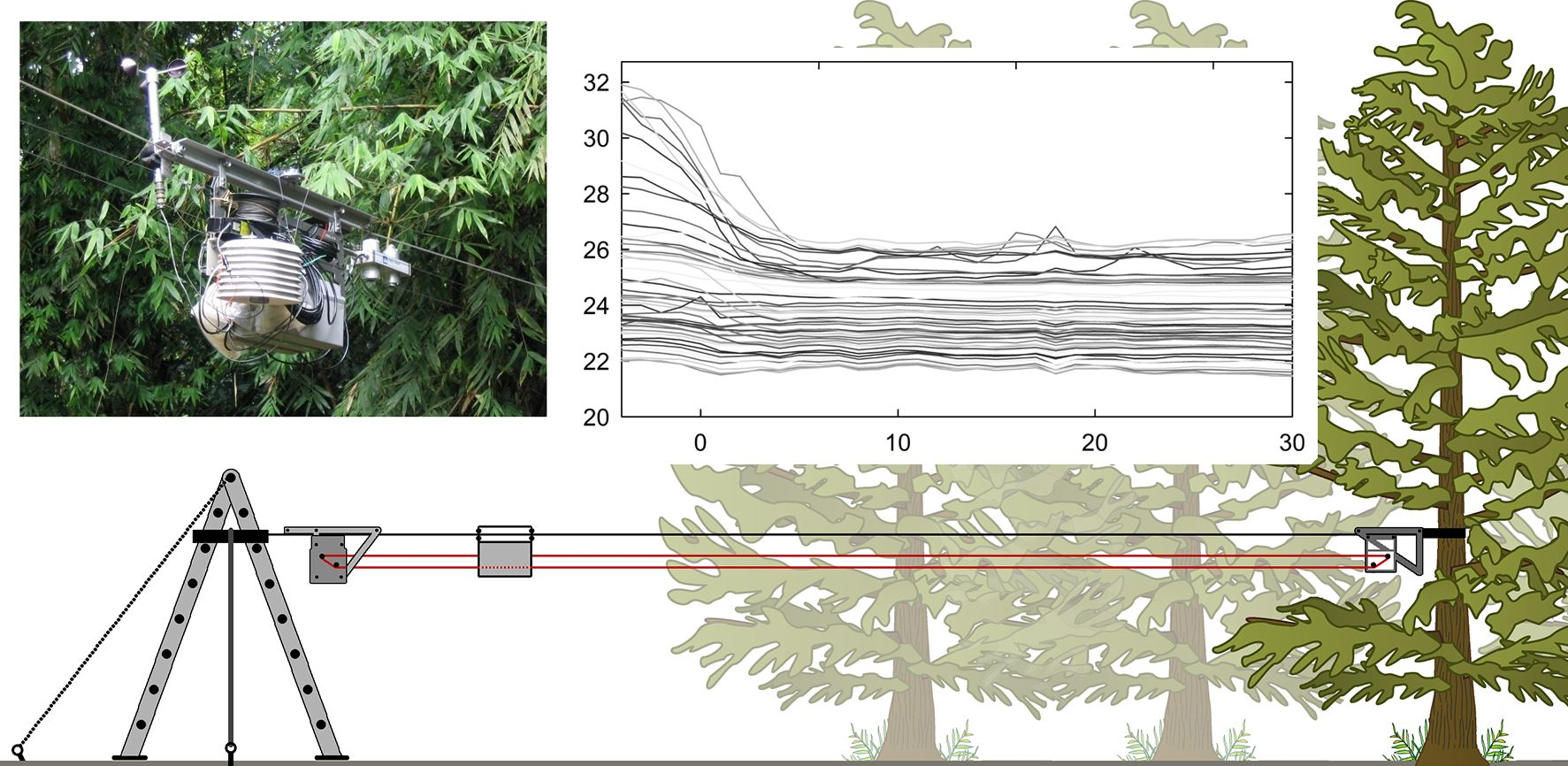
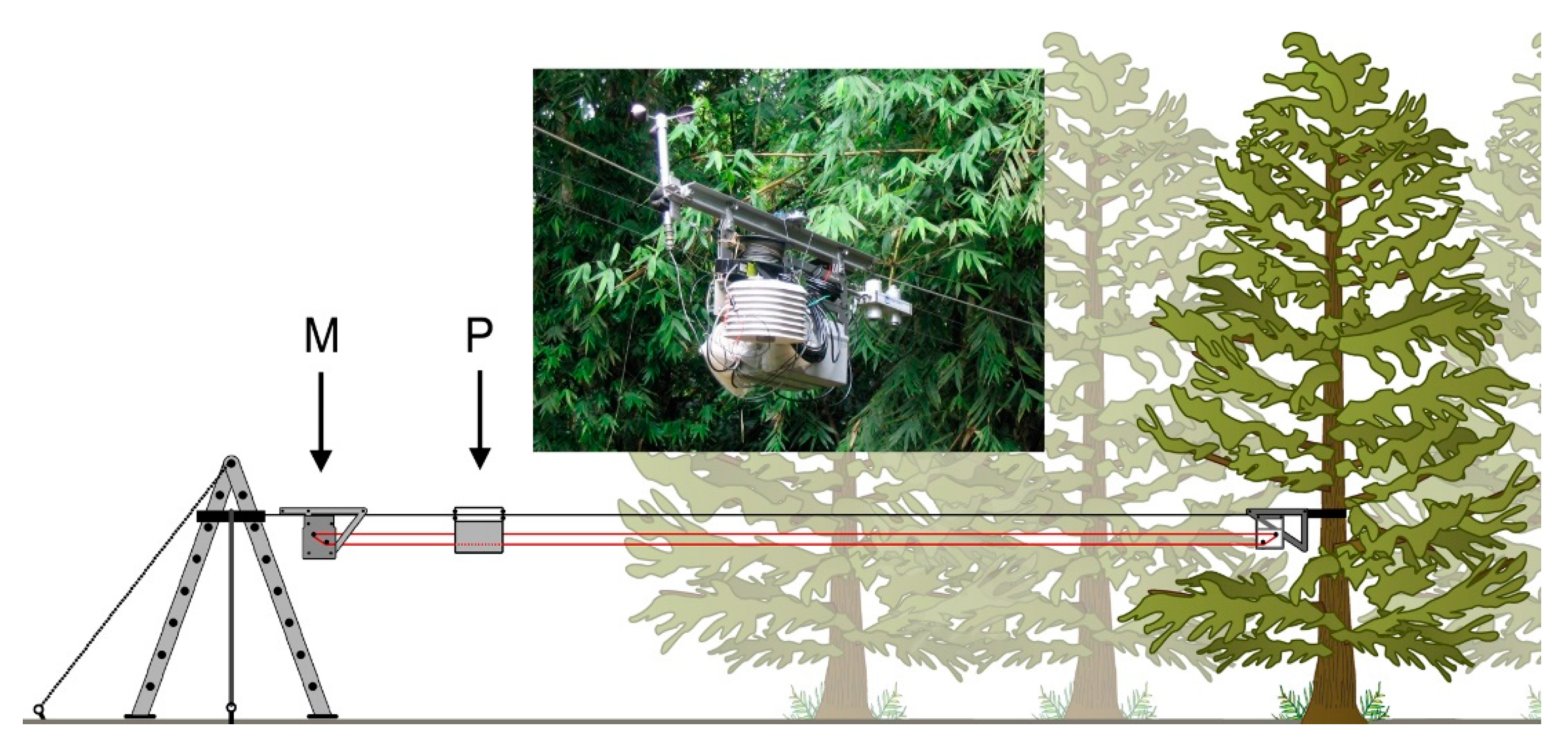
2.2. Mobile Sensing Platform
2.3. Data Analysis
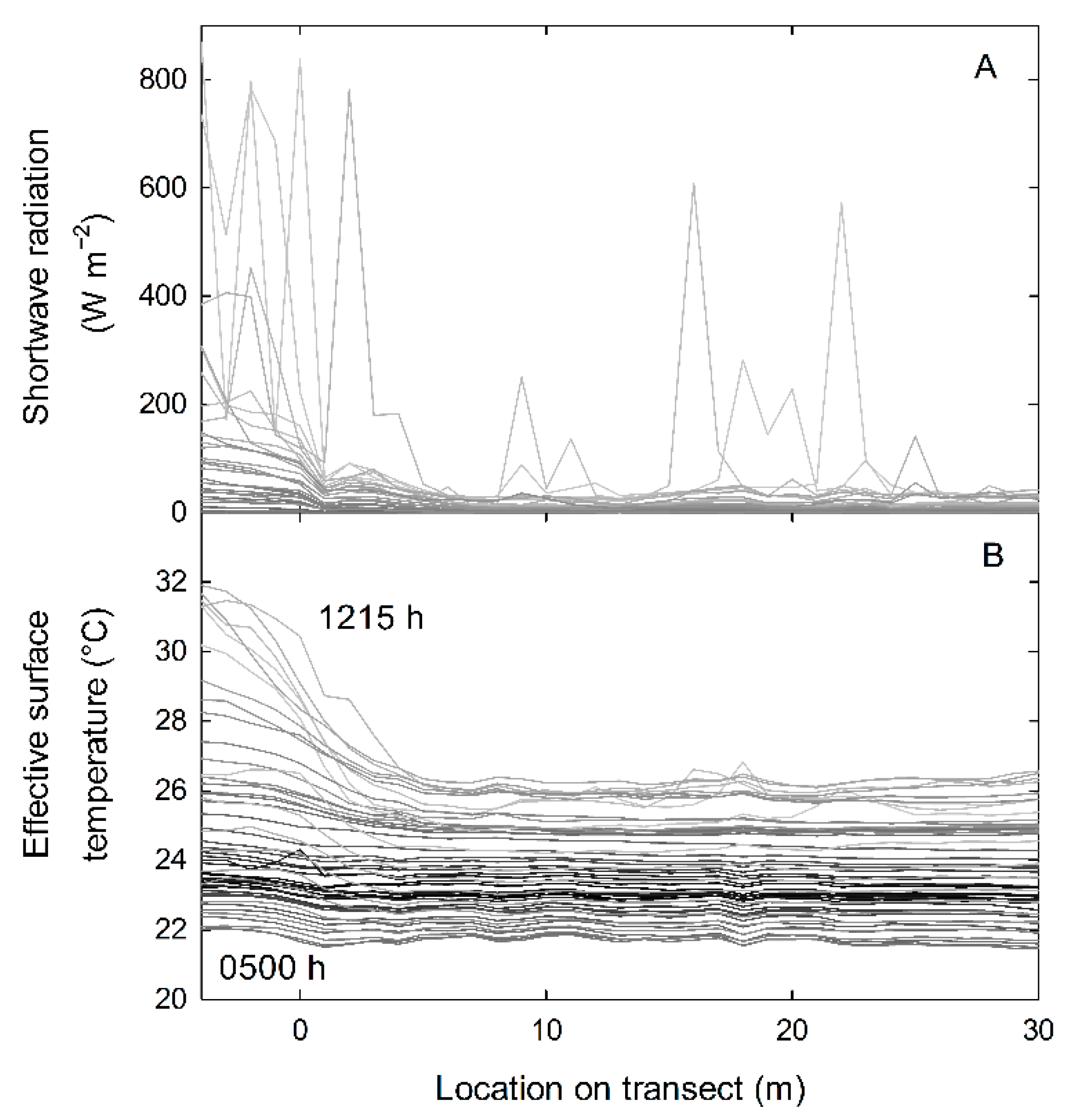
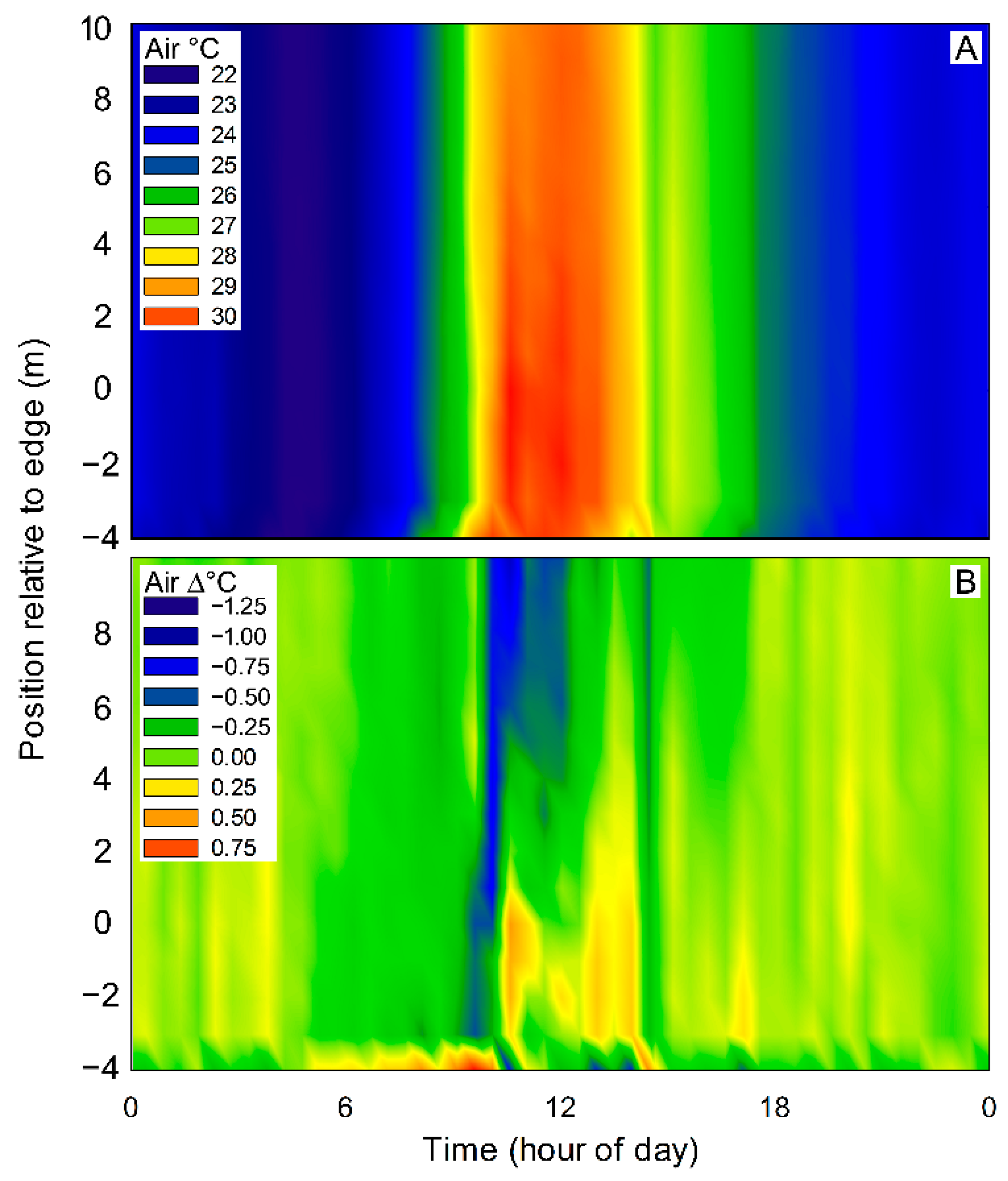
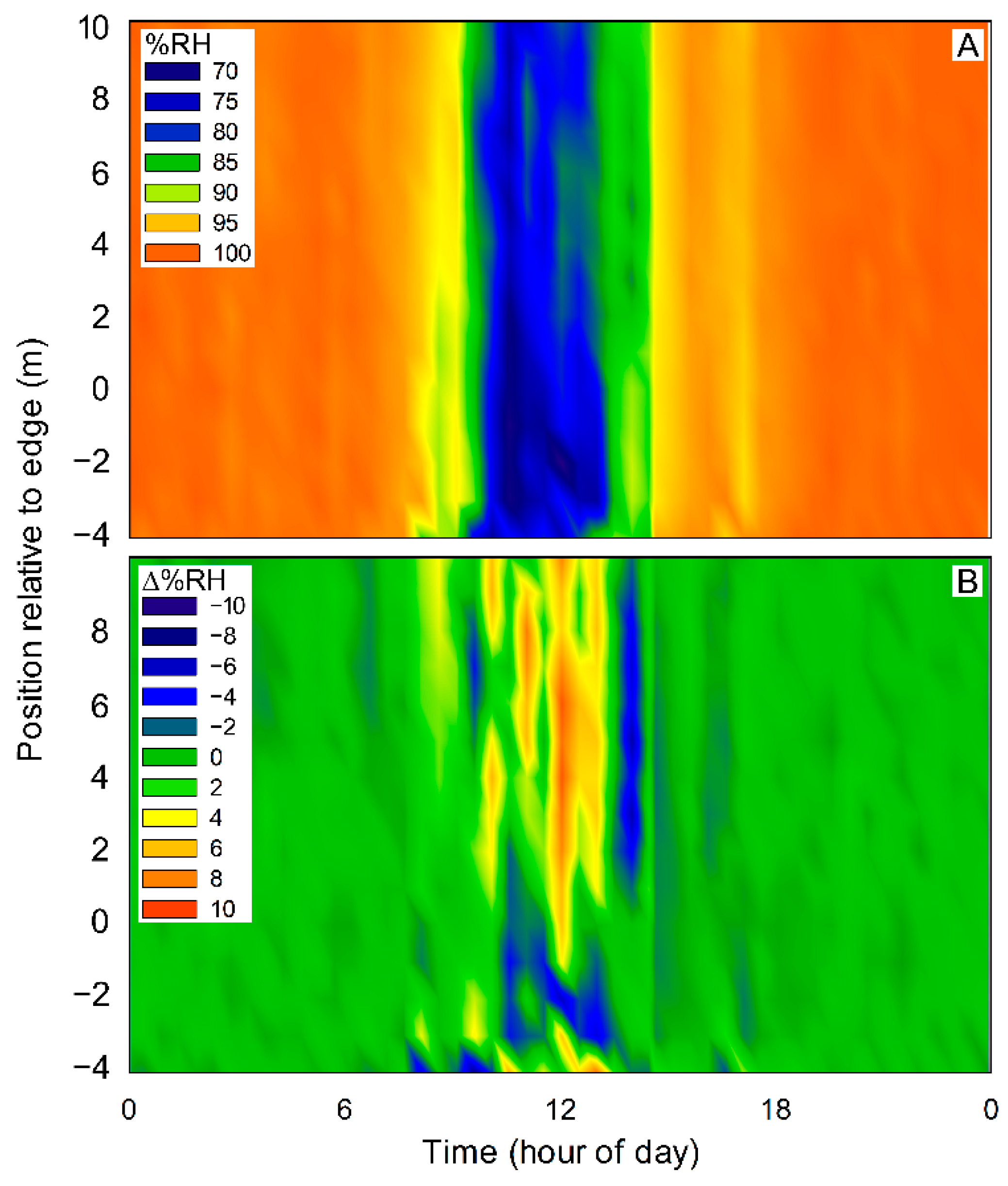
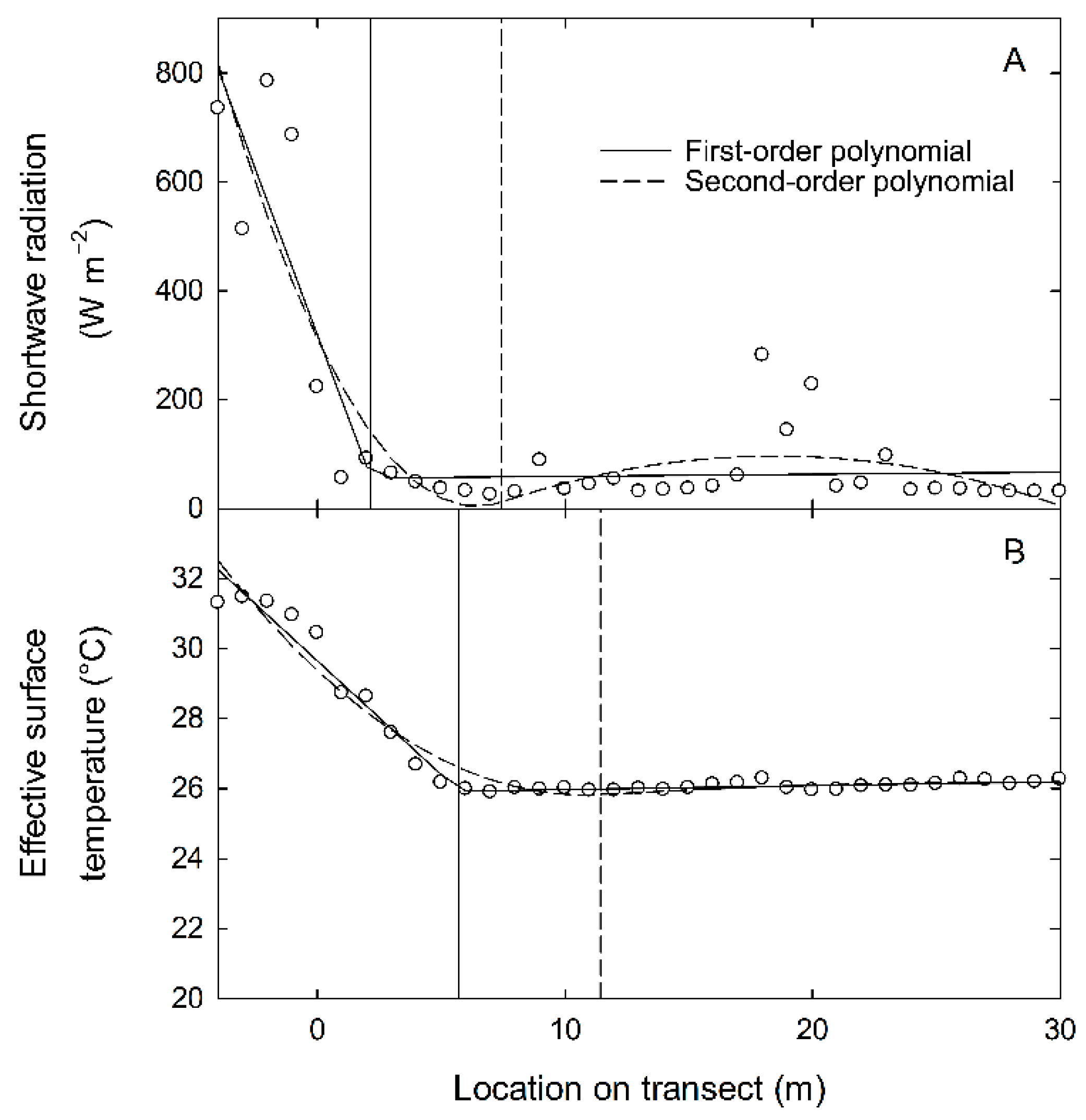
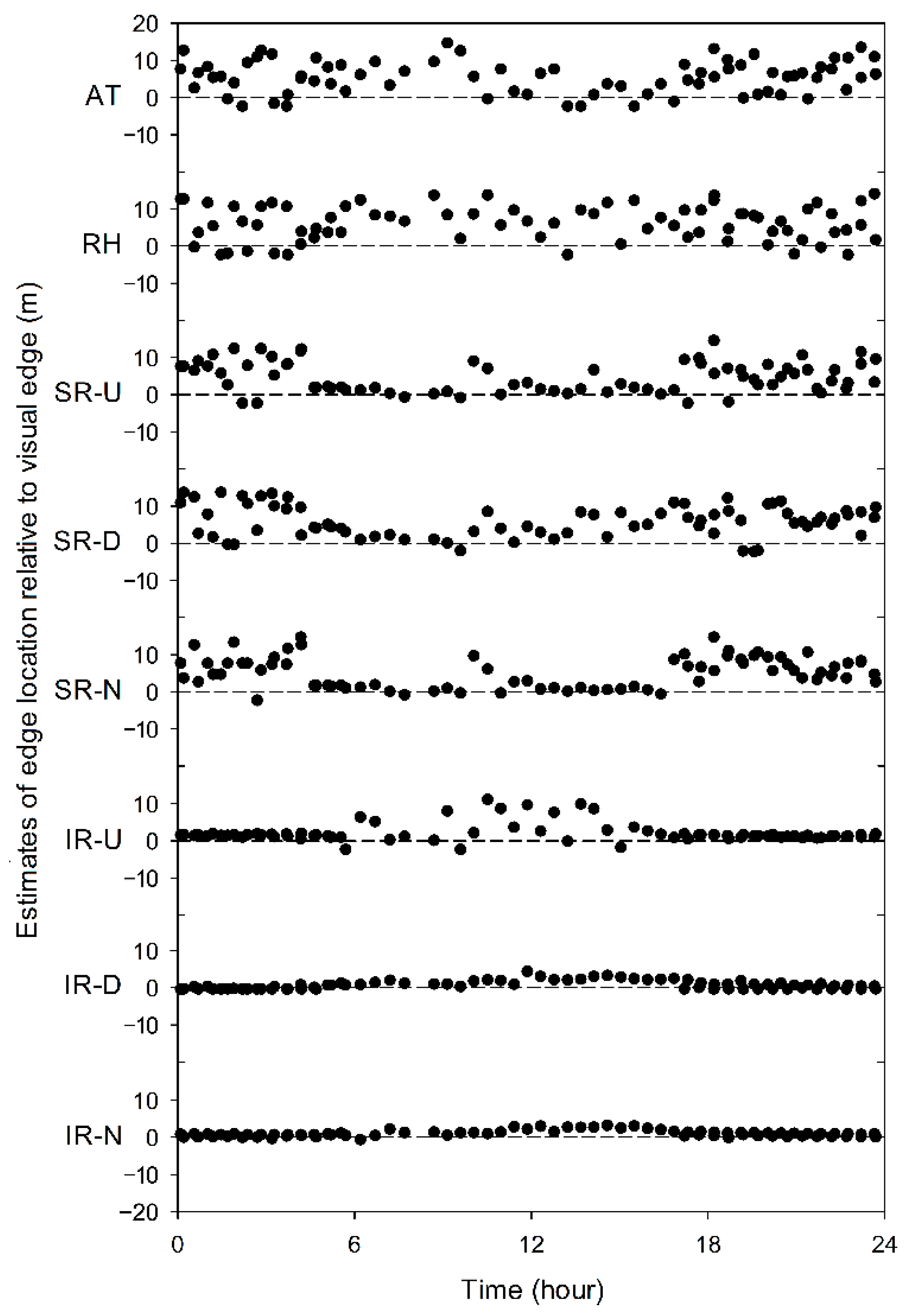
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmidt, M.; Jochheim, H.; Kersebaum, K.-C.; Lischeid, G.; Nende, L.C. Gradients of microclimate, carbon and nitrogen in transition zones of fragmented landscapes—A review. Agric. For. Meteorol. 2017, 232, 659–671. [Google Scholar] [CrossRef]
- Jucker, T.; Hardwick, S.R.; Both, S.; Elias, D.M.; Ewers, R.M.; Milodowski, D.T.; Coomes, D.A. Canopy structure and topography jointly constrain the microclimate of human-modified tropical landscapes. Glob. Chang. Biol. 2018, 24, 5243–5258. [Google Scholar] [CrossRef]
- Jucker, T.; Jackson, T.D.; Zellweger, F.; Swinfield, T.; Gregory, N.; Williamson, J.; Slade, E.M.; Phillips, J.W.; Bittencourt, P.R.; Blonder, B.; et al. A research agenda for microclimate ecology in human-modified tropical forests. Front. For. Glob. Chang. 2020, 2, 92. [Google Scholar] [CrossRef]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.A.R.; Jaiteh, M.S.; Esseen, P.A. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Laurance, W.F. Do edge effects occur over large spatial scales? Trends Ecol. Evol. 2000, 15, 134–135. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Nijs, I.; Lenoir, J. Incorporating microclimate into species distribution models. Ecography 2019, 42, 1267–1279. [Google Scholar] [CrossRef]
- Fagan, W.F.; Fortin, M.-J.; Soykan, C. Integrating edge detection and dynamic modeling in quantitative analyses of ecological boundaries. BioScience 2003, 53, 730–738. [Google Scholar] [CrossRef]
- Ries, L.; Murphy, S.M.; Wimp, G.M.; Fletcher, R.J. Closing persistent gaps in knowledge about edge ecology. Curr. Lands Ecol. Rep. 2017, 2, 30–41. [Google Scholar] [CrossRef]
- Chen, J.; Franklin, J.F.; Spies, T.A. Growing season microclimatic gradients from clearcut edges into old growth Douglas-fir forest. Ecol. Appl. 1995, 5, 74–86. [Google Scholar] [CrossRef]
- Meiners, S.J.; Pickett, S.T.A.; Handel, S.N. Probability of tree seedling establishment changes across a forest-old field edge gradient. Am. J. Bot. 2002, 89, 466–471. [Google Scholar] [CrossRef]
- Benitez-Malvido, J.; Lemus-Albor, A. The seedling community of tropical rain forest edges and its interaction with herbivores and pathogens. Biotropica 2005, 37, 301–313. [Google Scholar] [CrossRef]
- Wirth, R.; Meyer, S.T.; Leal, I.R.; Tabarelli, M. Plant herbivore interactions at the forest edge. Prog. Bot. 2008, 69, 423–448. [Google Scholar]
- Yahner, R.H. Changes in wildlife communities near edges. Conserv. Biol. 1988, 2, 333–339. [Google Scholar] [CrossRef]
- Kareiva, P. Habitat fragmentation and the stability of predator-prey interactions. Nature 1987, 326, 388–390. [Google Scholar] [CrossRef]
- Fagan, W.E.; Cantrell, R.S.; Cosner, C. How habitat edges change species interactions. Am. Nat. 1999, 153, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Foggo, A.; Ozanne, C.M.P.; Speight, M.R.; Hambler, C. Edge effects and tropical forest canopy invertebrates. Plant Ecol. 2001, 153, 347–359. [Google Scholar] [CrossRef]
- Asquith, N.M.; Mejia-Chang, M. Mammals, edge effects, and the loss of tropical forest diversity. Ecology 2005, 86, 379–390. [Google Scholar] [CrossRef]
- Ewers, R.M.; Boyle, M.J.; Gleave, R.A.; Plowman, N.S.; Benedick, S.; Bernard, H.; Turner, E.C. Logging cuts the functional importance of invertebrates in tropical rainforest. Nat. Commun. 2015, 6, 6836. [Google Scholar] [CrossRef]
- Chen, J.Q.; Saunders, S.C.; Crow, T.R.; Naiman, R.J.; Brosofske, K.D.; Mroz, G.D.; Brookshire, B.L.; Franklin, J.F. Microclimate in forest ecosystem and landscape ecology. BioScience 1999, 49, 288–297. [Google Scholar] [CrossRef]
- Dodonov, P.; Menezes, G.S.C.; Caitano, B.; Cazetta, E.; Mielke, M.S. Air and soil temperature across fire-created edges in a Neotropical rainforest. Agric. For. Meteorol. 2019, 276, 107606. [Google Scholar] [CrossRef]
- Menezes, G.S.C.; Cazetta, E.; Dodonov, P. Vegetation structure across fire edges in a Neotropical rain forest. For. Ecol. Manag. 2019, 453, 117587. [Google Scholar] [CrossRef]
- Grimmond, C.S.B.; Robeson, S.M.; Schoof, J.T. Spatial variability of micro-climatic conditions within a mid-latitude deciduous forest. Clim. Res. 2000, 15, 137–149. [Google Scholar] [CrossRef]
- Camargo, J.C.L.; Kapos, V. Complex edge effects on soil moisture and microclimate in central Amazonian forest. J. Trop. Ecol. 1995, 11, 205–221. [Google Scholar] [CrossRef]
- Kapos, V.; Wandelli, E.E.; Camargo, J.L.; Ganade, G. Edge-related changes in environment and plant responses due to forest fragmentation in central Amazonia. In Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities; Laurance, W.F., Bierregaard, R.O., Eds.; University of Chicago Press: Chicago, IL, USA, 1997; pp. 33–44. [Google Scholar]
- Williams-Linera, G.; Dominguez-Gastelu, V.; Garcia-Zurita, M.E. Microenvironment and floristics of different edges in a fragmented tropical rainforest. Conserv. Biol. 1998, 12, 1091–1102. [Google Scholar] [CrossRef]
- Dignan, P.; Bren, L. Modelling light penetration edge effects for stream buffer design in mountain ash forest in southeastern. Aust. For. Ecol. Manag. 2003, 175, 95–106. [Google Scholar] [CrossRef]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological consequences of ecosystem fragmentation: A review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Li, Y.; Kang, W.; Han, Y.; Song, Y. Spatial and temporal patterns of microclimates at an urban forest edge and their management implications. Environ. Monit. Assess. 2018, 190, 93. [Google Scholar] [CrossRef] [PubMed]
- Fortin, M.-J.; Olson, R.J.; Ferson, S.; Iverson, L.; Hunsaker, C.; Edwards, G.; Levine, D.; Butera, K.; Klemas, V. Issues related to the detection of boundaries. Land Ecol. 2000, 15, 453–466. [Google Scholar] [CrossRef]
- Saunders, S.C.; Chen, J.Q.; Drummer, T.D.; Crow, T.R. Modeling temperature gradients across edges over time in a managed landscape. For. Ecol. Manag. 1999, 117, 17–31. [Google Scholar] [CrossRef]
- Newmark, W.D. Tanzanian forest edge microclimatic gradients: Dynamic patterns. Biotropica 2001, 33, 2–11. [Google Scholar] [CrossRef]
- Bramer, I.; Anderson, B.J.; Bennie, J.; Bladon, A.J.; De Frenne, P.; Hemming, D.; Gillingham, P.K. Advances in monitoring and modelling climate at ecologically relevant scales. Adv. Ecol. Res. 2018, 58, 101–161. [Google Scholar] [CrossRef]
- Wild, J.; Kopecký, M.; Macek, M.; Šanda, M.; Jankovec, J.; Haase, T. Climate at ecologically relevant scales: A new temperature and soil moisture logger for long-term microclimate measurement. Agric. For. Meteorol. 2019, 268, 40–47. [Google Scholar] [CrossRef]
- Zellweger, F.; Frenne, P.; De Lenoir, J.; Rocchini, D.; Coomes, D. Advances in microclimate ecology arising from remote sensing. Trends Ecol. Evol. 2019, 34, 327–341. [Google Scholar] [CrossRef]
- Toms, J.D.; Lesperance, M.L. Piecewise regression: A tool for identifying ecological thresholds. Ecology 2003, 84, 2034–2041. [Google Scholar] [CrossRef]
- Jordan, B.L.; Batalin, M.A.; Kaiser, W.J. NIMS RD: A Rapidly Deployable Cable Based Robot. ICRA’07. In Proceedings of the 2007 IEEE International Conference on Robotics and Automation, Roma, Italy, 10–14 April 2007. [Google Scholar]
- Restrepo, C.; Gomez, N.; Heredia, S. Anthropogenic edges, treefall gaps, and fruit–frugivore interactions in a neotropical montane forest. Ecology 1999, 80, 668–685. [Google Scholar]
- Kim, Y.; Still, C.J.; Hanson, C.V.; Kwon, H.; Greer, B.T.; Law, B.E. Canopy skin temperature variations in relation to climate, soil temperature, and carbon flux at a ponderosa pine forest in central Oregon. Agric. For. Meteorol. 2016, 226–227, 161–173. [Google Scholar] [CrossRef]
- Cescatti, A.; Marcolla, B.; Santhana Vannan, S.K.; Yun Pan, J.; Román, M.O.; Yang, X.; Ciais, P.; Cook, R.B.; Law, B.E.; Matteucci, G.; et al. Intercomparison of MODIS albedo retrievals and in situ measurements across the global FLUXNET network. Remote Sens. Environ. 2012, 121, 323–334. [Google Scholar] [CrossRef]
- Cadenasso, M.L.; Traynor, M.M.; Pickett, S.T.A. Functional location of forest edges: Gradients of multiple physical factors. Can. J. For. Res. 1997, 27, 774–782. [Google Scholar] [CrossRef]
- Davies-Colley, R.J.; Payne, G.W.; van Elswijk, M. Microclimate gradients across a forest edge. N. Z. J. Ecol. 2000, 24, 111–121. [Google Scholar]
- Gehlhausen, S.M.; Schwartz, M.W.; Augspurger, C.K. Vegetation and microclimatic edge effects in two mixed mesophytic forest fragments. Plant Ecol. 2000, 147, 21–35. [Google Scholar] [CrossRef]
- Chen, J.; Franklin, J.F.; Spies, T.A. Contrasting microclimates among clearcut, edge, and interior of old-growth Douglas-fir forest. Agric. For. Meteorol. 1993, 63, 219–237. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transect | Total Transect Length (m) | Relative Location of Base in Clearing from Forest Edge (m from Edge) | Number of Measurement Repetitions (“Runs”) in Approximately 24 h | Total Number of Data Points Collected per Sensor |
|---|---|---|---|---|
| 1 | 37 | −6 | 42 | 1065 |
| 2 | 27 | −6 | 77 | 1848 |
| 3 | 35 | −4 | 52 | 1820 |
| Variable | Time | In Clearing | Under Canopy |
|---|---|---|---|
| Air temperature (°C) | 11.8 ± 0.9 | 28.9 ± 1.0 | 27.7 ± 0.7 |
| Relative humidity (%) | 13.4 ± 1.9 | 79.2 ± 6.4 | 88.2 ± 5.7 |
| Solar radiation, upward-facing (W m−2) | 11.1 ± 0.6 | 558.5 ± 291.3 | 33.6 ± 25.1 |
| Solar radiation, downward-facing (W m−2) | 10.6 ± 0.8 | 51.7 ± 26.9 | 4.5 ± 2.9 |
| Net solar radiation (W m−2) | 11.1 ± 0.6 | 511.1 ± 268.2 | 29.6 ± 21.6 |
| IR radiation, upward-facing (W m−2) | 13.4 ± 8.8 | 434.8 ± 27.2 | 437.3 ± 15.2 |
| IR radiation, downward-facing (W m−2) | 12.3 ± 1.4 | 483.4 ± 15.5 | 448.1 ± 6.8 |
| Net IR radiation (W m−2) | 12.3 ± 1.4 | −25.4 ± 11.8 | 8.5 ± 3.4 |
| Variable | Relative Location of Edge (m) | ||
|---|---|---|---|
| Transect 1 (n = 16) | Transect 2 (n = 21) | Transect 3 (n = 24) | |
| Air temperature | 13.2 ± 1.5 | 7.6 ± 1.1 | 10.4 ± 1.5 |
| Relative humidity | 13.1 ± 2.0 | 7.7 ± 0.8 | 8.6 ± 1.5 |
| Solar radiation, upward-facing | −1.4 ± 0.4 | 2.9 ± 0.8 | 0.2 ± 0. 5 |
| Solar radiation, downward-facing | 7.0 ± 1.1 | 3.3 ± 0.8 | 1.3 ± 0.3 |
| Net solar radiation | 1.1 ± 2.1 | 7.2 ± 1.1 | −0.2 ± 0.5 |
| IR radiation, upward-facing | 6.9 ± 3.1 | 7.1 ± 1.0 | 1.4 ± 1.4 |
| IR radiation, downward-facing | 1.8 ± 0.2 | 1.8 ± 0.2 | 0.8 ± 0.2 |
| Net IR radiation | 0.6 ± 0.2 | 1.7 ± 0.2 | −0.2 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graham, E.A.; Hansen, M.; Kaiser, W.J.; Lam, Y.; Yuen, E.; Rundel, P.W. Dynamic Microclimate Boundaries across a Sharp Tropical Rainforest–Clearing Edge. Remote Sens. 2021, 13, 1646. https://doi.org/10.3390/rs13091646
Graham EA, Hansen M, Kaiser WJ, Lam Y, Yuen E, Rundel PW. Dynamic Microclimate Boundaries across a Sharp Tropical Rainforest–Clearing Edge. Remote Sensing. 2021; 13(9):1646. https://doi.org/10.3390/rs13091646
Chicago/Turabian StyleGraham, Eric A., Mark Hansen, William J. Kaiser, Yeung Lam, Eric Yuen, and Philip W. Rundel. 2021. "Dynamic Microclimate Boundaries across a Sharp Tropical Rainforest–Clearing Edge" Remote Sensing 13, no. 9: 1646. https://doi.org/10.3390/rs13091646
APA StyleGraham, E. A., Hansen, M., Kaiser, W. J., Lam, Y., Yuen, E., & Rundel, P. W. (2021). Dynamic Microclimate Boundaries across a Sharp Tropical Rainforest–Clearing Edge. Remote Sensing, 13(9), 1646. https://doi.org/10.3390/rs13091646