
Estimation of the Conifer-Broadleaf Ratio in Mixed Forests Based on Time-Series Data



Abstract
:
1. Introduction
2. Materials and Methods
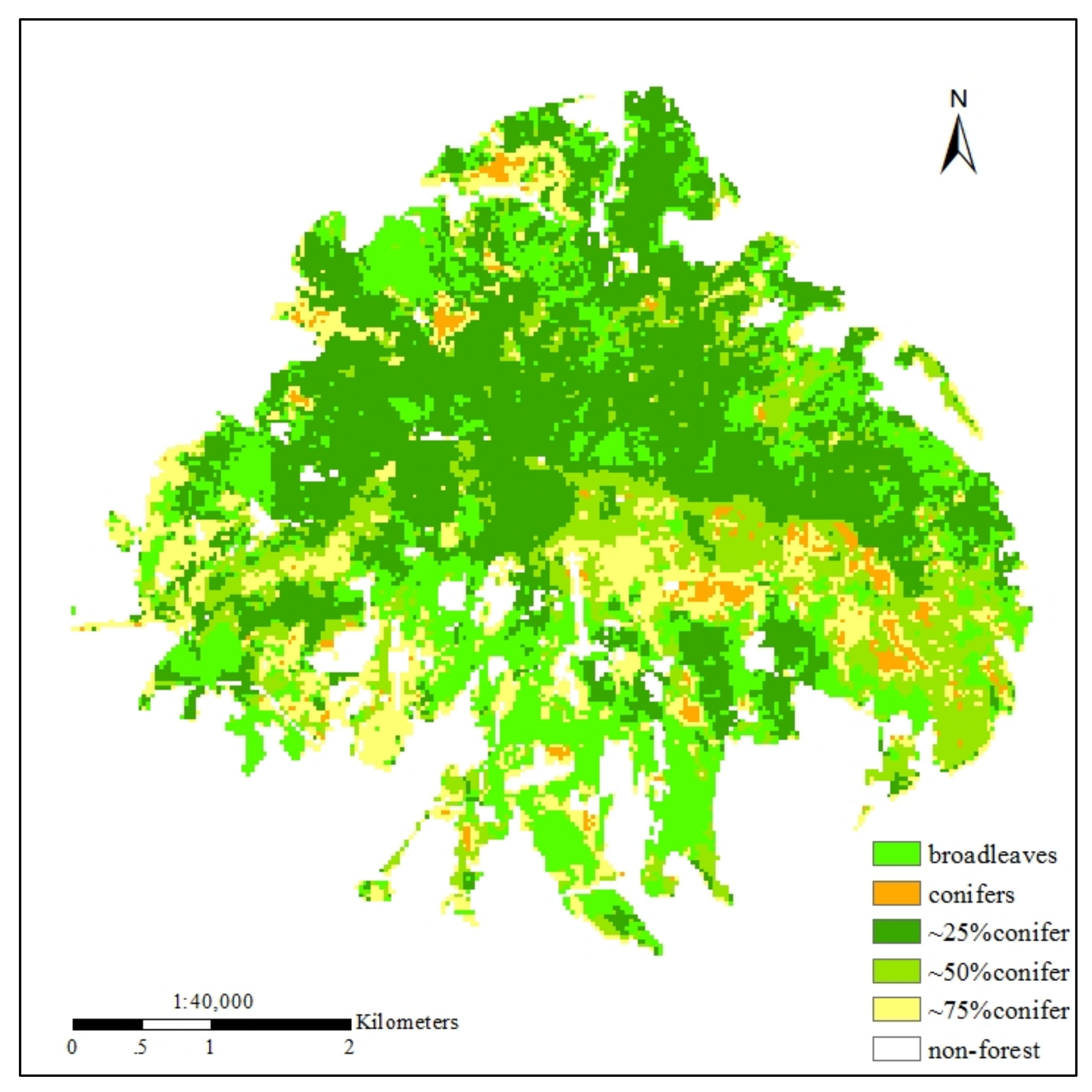
2.1. Study Area
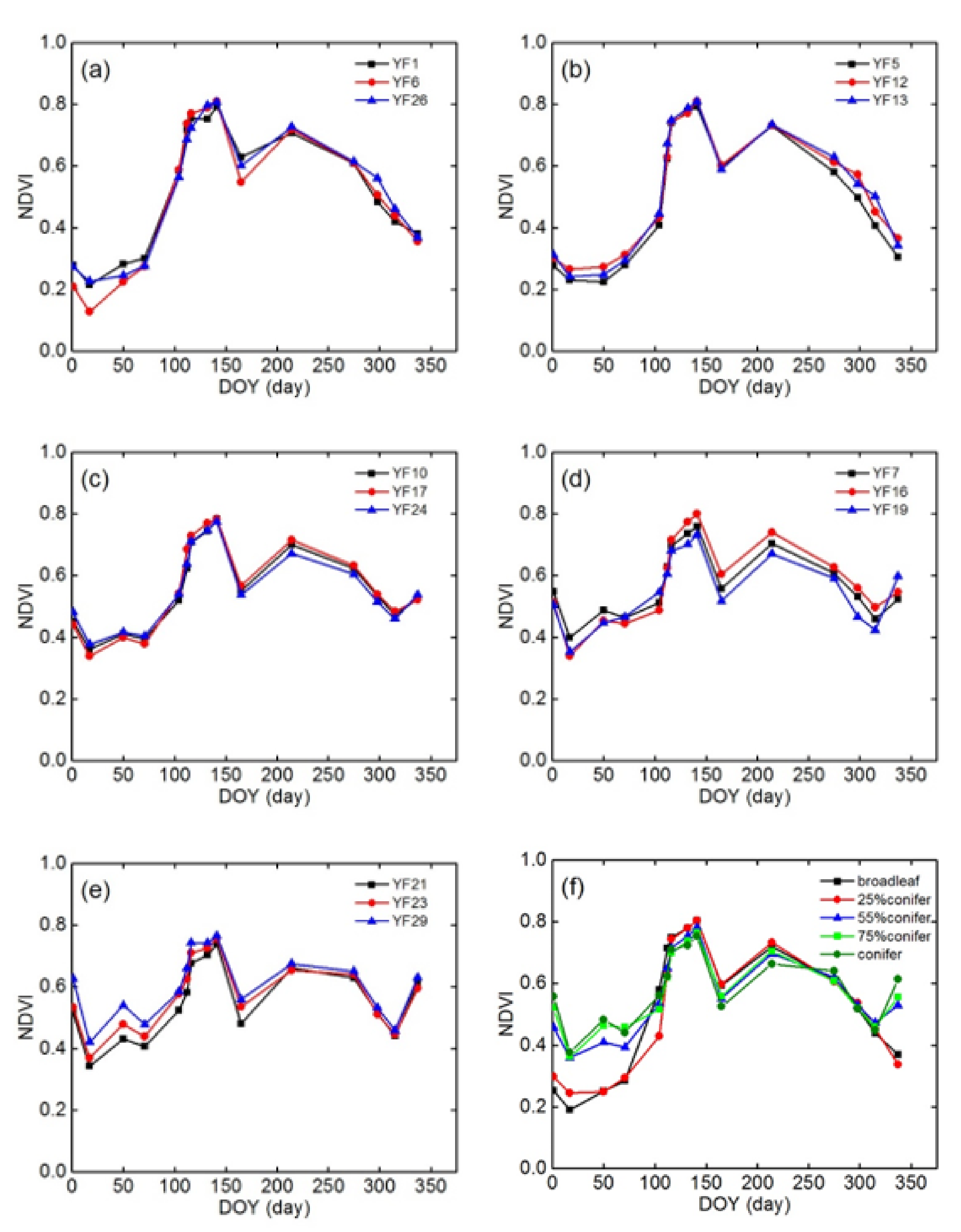
2.2. Field Measurements
2.2.1. Field Experiment Design
2.2.2. LAI Observations
2.2.3. Conifer-Broadleaf Ratio Observations
2.2.4. Model Parameter Input Measurements
2.3. Satellite Image Information and Pre-Processing
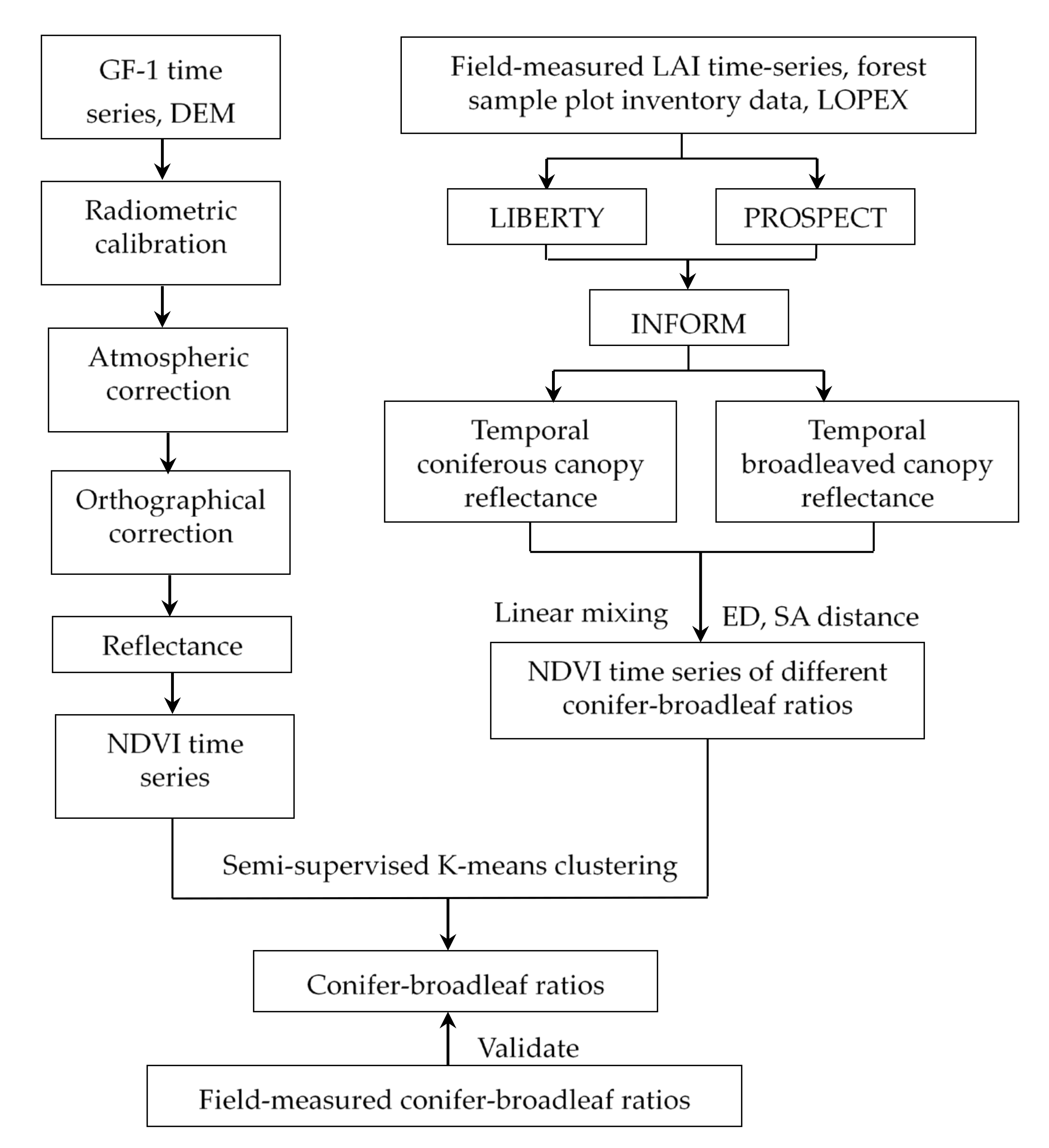
2.4. Methodology
- (a)
- Analyzing model parameter sensitivity and determining the value range of the parameter inputs according to the field measurements of the LAI time-series data, sample plot inventory, and other reference materials.
- (b)
- Simulating the visible and near-infrared bi-directional spectral reflectance of coniferous and broad-leaved forest canopies using INFORM and calculating the NDVI time-series under different conifer-broadleaf ratios based on the principles of linear mixing.
- (c)
- Calculation of the Euclidean distance and spectral angle distance between any two NDVI time-series curves of different conifer-broadleaf ratios, analyzing the separability and determining the typical separable ratios.
- (d)
- Constructing a GF-1 NDVI time-series dataset, from which curves with typical separable ratios were extracted as prior sample data.
- (e)
- Using the semi-supervised K-means clustering method, obtain the conifer-broadleaf ratio of the study area at the pixel level based on the GF-I NDVI time-series data and evaluate the overall estimation accuracy.
2.4.1. Model Description
2.4.2. NDVI Time-Series
2.4.3. Similarity Measures for Time-Series
2.4.4. Semi-Supervised K-Means Clustering of NDVI Time-Series
- Step 1.
- Define the number of clusters, k, and calculated the initial cluster centers, .
- Step 2.
- Calculate the ED of objects from .
- Step 3.
- Obtain the closest cluster to the object and assign the object to that cluster.
- Step 4.
- Recalculate the cluster means and update the cluster centers.
- Step 5.
- Recalculate the ED of the objects from the updated cluster centers.
- Step 6.
- Repeat steps 2–5 until the cluster means do not update.
3. Results
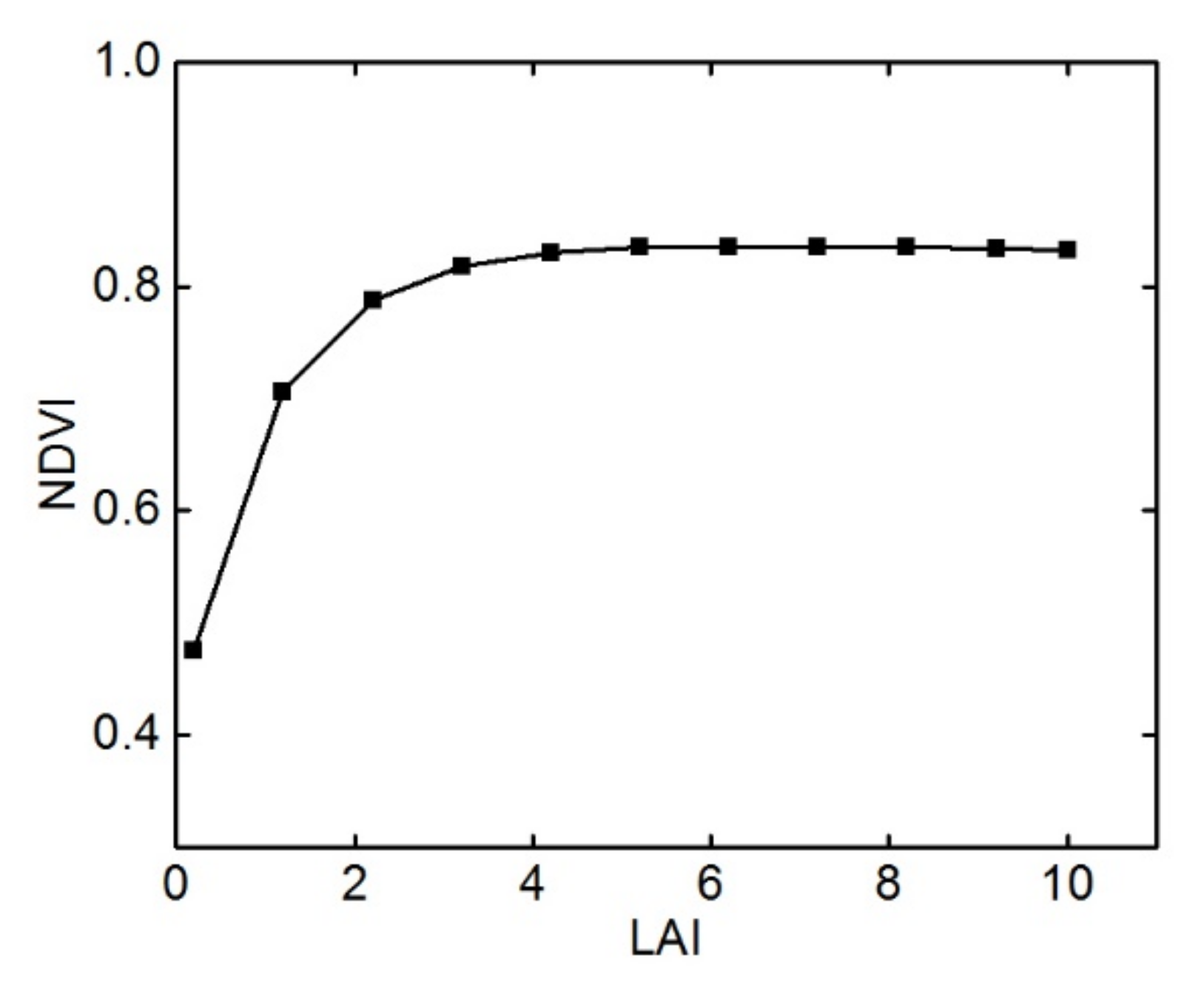
3.1. Sensitivity Analysis and Determination of Model Parameters in Different Growth Periods
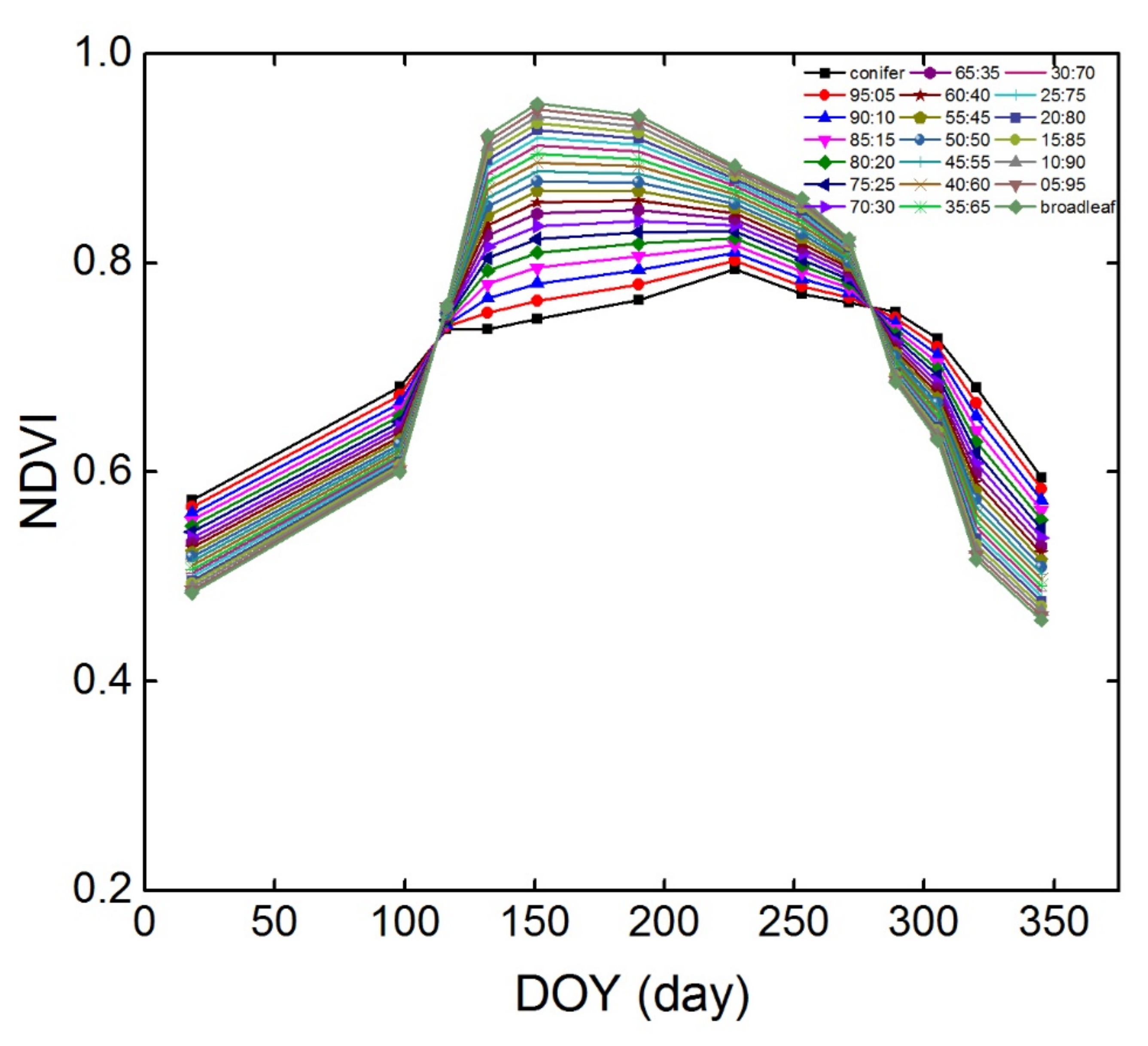
3.2. NDVI Time-Series of Different Conifer-Broadleaf Ratios
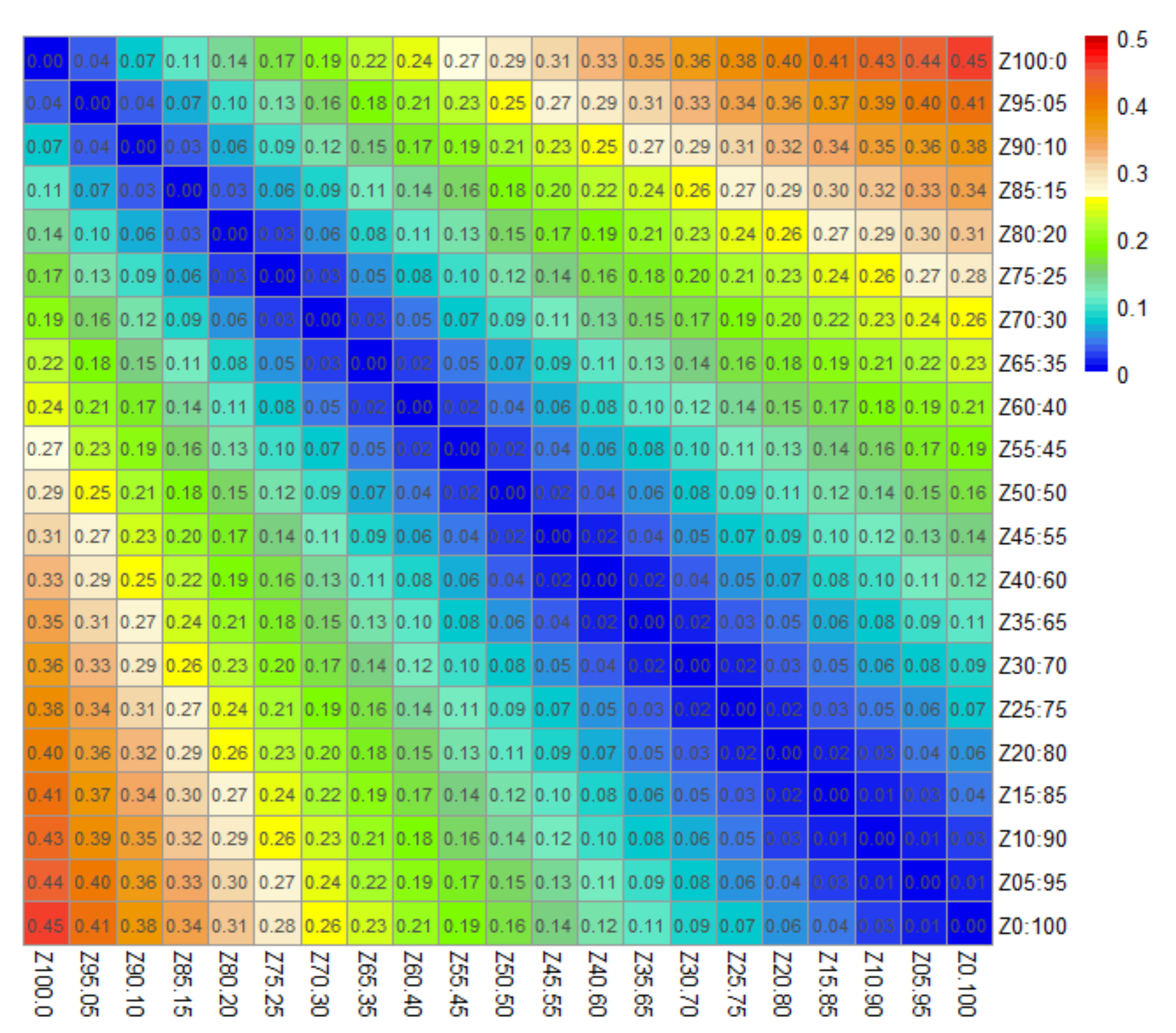
3.3. Calculation of ED and SA Distance
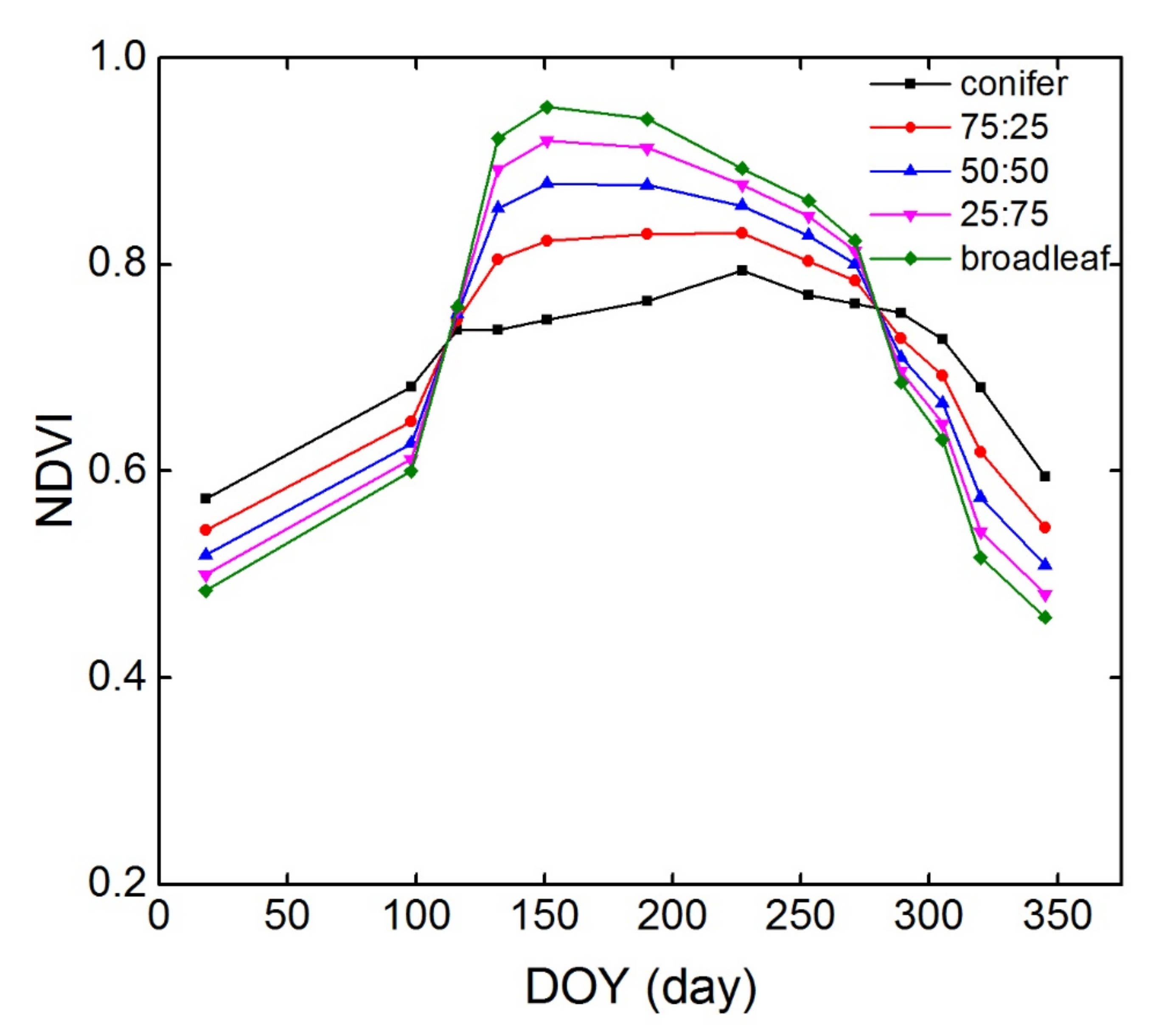
3.4. NDVI Time-Series of Typical Conifer-Broadleaf Ratios
3.5. Conifer-Broadleaf Ratio Estimation Based on Time-Series
4. Discussion
4.1. Parameter Sensitivity Analysis
4.2. The NDVI Time-Series
4.3. Conifer-Broadleaf Ratios
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Canadian Council of Forest Ministers. Criteria and Indicators of Sustainable Forest Management in Canada: National Status. 2000. Available online: https://d1ied5g1xfgpx8.cloudfront.net/pdfs/18104.pdf (accessed on 6 April 2020).
- Brown, S. Measuring carbon in forests: Current status and future challenges. Environ. Pollut. 2002, 116, 363–372. [Google Scholar] [CrossRef]
- Ustin, L.S. Remote sensing of canopy chemistry. Proc. Natl. Acad. Sci. USA 2013, 110, 804–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzystek, P.; Serebryanyk, A.; Schnörr, C.; Červenka, J.; Heurich, M. Large-scale mapping of tree species and dead trees in šumava national park and bavarian forest national park using lidar and multispectral imagery. Remote Sens. 2020, 12, 661. [Google Scholar] [CrossRef] [Green Version]
- Porté, A.; Bartelink, H. Modelling mixed forest growth: A review of models for forest management. Ecol. Model. 2002, 150, 141–188. [Google Scholar] [CrossRef]
- Ollinger, S.V.; Reich, P.B.; Frolking, S.; Lucie, C.; Lepinea, D.Y.; Hollingerd, A.D.; Richardsone, A.D. Nitrogen cycling, forest canopy reflectance, and emergent properties of ecosystems. Proc. Natl. Acad. Sci. USA 2013, 110, 2437. [Google Scholar] [CrossRef] [Green Version]
- Asner, G.P.; Martin, R.E. Spectral and chemical analysis of tropical forests: Scaling from leaf to canopy levels. Remote Sens. Environ. 2008, 112, 3958–3970. [Google Scholar] [CrossRef]
- Rautiainen, M.; Stenberg, P.; Nilson, T.; Kuusk, A. The effect of crown shape on the reflectance of coniferous stands. Remote Sens. Environ. 2004, 89, 41–52. [Google Scholar] [CrossRef]
- Knyazikhin, Y.; Schull, M.A.; Stenberg, P.; Mõttusd, M.; Rautiainenc, M.; Yanga, Y.; Marshake, A.; Carmonaf, P.L.; Kaufmanna, R.K.; Lewisg, P.; et al. Hyperspectral remote sensing of foliar nitrogen content. Proc. Natl. Acad. Sci. USA 2013, 110, E185–E192. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ding, X.; Tong, X.; Atkinson, M.P. Spatio-temporal spectral unmixing of time-series images. Remote Sens. Environ. 2021, 259, 112407. [Google Scholar] [CrossRef]
- Abdollahnejad, A.; Panagiotidis, D. Tree species classification and health status assessment for a mixed broadleaf-conifer forest with UAS multispectral imaging. Remote Sens. 2020, 12, 3722. [Google Scholar] [CrossRef]
- Burchard-Levine, V.; Nieto, H.; Riañoa, D.; Migliavacca, M.; El-Madany, T.S.; Guzinskie, R.; Carrara, A.; Martín, P.M. The effect of pixel heterogeneity for remote sensing based retrievals of evapotranspiration in a semi-arid tree-grass ecosystem. Remote Sens. Environ. 2021, 260, 112440. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, L.; Chen, J.; Rao, H.Y.; Zhou, Y.; Chen, X.H. Assessing the impact of endmember variability on linear spectral mixture analysis (LSMA): A theoretical and simulation analysis. Remote Sens. Environ. 2019, 235, 111471. [Google Scholar] [CrossRef]
- Chen, J.M.; Black, T.A.; Adams, R.S. Evaluation of hemispherical photography for determining plant area index and geometry of a forest stand. Agric. For. Meteorol. 1991, 56, 129–143. [Google Scholar] [CrossRef]
- Zou, T.Y.; Zhang, J. A New fluorescence quantum yield efficiency retrieval method to simulate chlorophyll fluorescence under natural conditions. Remote Sens. 2020, 12, 4053. [Google Scholar] [CrossRef]
- Chen, J.M. Spatial scaling of a remotely sensed surface parameter by contexture. Remote Sens. Environ. 1999, 69, 30–42. [Google Scholar] [CrossRef]
- Li, X.W.; Strahler, A.H.; Woodcock, C.A. Hybrid geometric optical-radiative transfer approach for modeling albedo and directional reflectance of discontinuous canopies. IEEE Trans. Geosci. Remote Sens. 1995, 33, 466–480. [Google Scholar] [CrossRef]
- Liang, L.; Di, G.; Yan, J.; Qiu, S.; Di, L.; Wang, S.; Xu, L.; Wang, L.; Kang, J.; Li, L. Estimating crop LAI using spectral feature extraction and the hybrid inversion method. Remote Sens. 2020, 12, 3534. [Google Scholar] [CrossRef]
- Atzberger, C. Development of an invertible forest reflectance model: The INFOR-Model. A Decade of Trans-European Remote Sensing Cooperation. In Proceedings of the 20th EARSeL Symposium, Dresden, Germany, 14–16 June 2000. [Google Scholar]
- Schlerf, M.; Atzberger, C. Inversion of a forest reflectance model to estimate structural canopy variables from hyperspectral remote sensing data. Remote Sens. Environ. 2006, 100, 281–294. [Google Scholar] [CrossRef]
- Rosema, A.; Verhoef, W.; Noorbergen, H.; Borgesius, J. A new forest light interaction model in support of forest monitoring. Remote Sens. Environ. 1992, 42, 23–41. [Google Scholar] [CrossRef]
- Verhoef, W. Light scattering by leaf layers with application to canopy reflectance modeling: The SAIL model. Remote Sens. Environ. 1984, 16, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Verhoef, W. Earth observation modeling based on layer scattering matrices. Remote Sens. Environ. 1985, 17, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Jacquemoud, S.; Baret, F. PROSPECT: A model of leaf optical properties spectra. Remote Sens. Environ. 1990, 34, 75–91. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Ustin, S.L.; Verdebout, J.; Schmuck, G.; Andreoli, G.; Hosgood, B. Estimating leaf biochemistry using the prospect leaf optical properties model. Remote Sens. Environ. 1996, 56, 194–202. [Google Scholar] [CrossRef]
- Dawson, T.P.; Curran, P.J.; Plummer, S.E. LIBERTY-Modeling the effects of leaf biochemical concentration on reflectance spectra. Remote Sens. Environ. 1998, 65, 50–60. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, R.; Atzberger, C.; Li, F.; Loiselle, S.A.; Luo, J. Estimating forest fapar from multispectral landsat-8 data using the invertible forest reflectance model INFORM. Remote Sens. 2015, 7, 7425–7446. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.J.; Huang, W.J.; Wang, J.H.; Xing, Z.R. Inversion of forest leaf area index calculated from multi-source and multi-angle remote sensing data. Chin. Bull. Bot. 2010, 45, 566–578. [Google Scholar]
- Zhang, L.; Zhao, J.L.; Jia, K.; Li, X.S. Research on plant spectral recognition method based on phenological features. Spectrosc. Spectr. Analysis. 2015, 10, 2836–2840. [Google Scholar]
- Huang, X.; Zhu, W.; Wang, X.; Zhan, P.; Liu, Q.; Li, X.; Sun, L. A method for monitoring and forecasting the heading and flowering dates of winter wheat combining satellite-derived green-up dates and accumulated temperature. Remote Sens. 2020, 12, 3536. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS-1, Third Earth Resources Technology Satellite Symposium 1; NASA: Wasgington, DC, USA, 1974; pp. 309–317. [Google Scholar]
- Dobrinić, D.; Gašparović, M.; Medak, D. Sentinel-1 and 2 time-series for vegetation mapping using random forest classification: A case study of Northern Croatia. Remote Sens. 2021, 13, 2321. [Google Scholar] [CrossRef]
- López-Amoedo, A.; Álvarez, X.; Lorenzo, H.; Rodríguez, J.L. Multi-temporal Sentinel-2 data analysis for smallholding forest cut control. Remote Sens. 2021, 13, 2983. [Google Scholar] [CrossRef]
- Chamberlain, D.A.; Phinn, S.R.; Possingham, H.P. Mangrove forest cover and phenology with landsat dense time series in central queensland, Australia. Remote Sens. 2021, 13, 3032. [Google Scholar] [CrossRef]
- Lhermitte, S.; Verbesselt, J.; Verstraeten, W.W.; Coppin, P. A comparison of time series similarity measures for classification and change detection of ecosystem dynamics. Remote Sens. Environ. 2011, 115, 3129–3152. [Google Scholar] [CrossRef]
- Lambin, E.F.; Strahlers, A.H. Change-vector analysis in multitemporal space: A tool to detect and categorize land-cover change processes using high temporal-resolution satellite data. Remote Sens. Environ. 1994, 48, 231–244. [Google Scholar] [CrossRef]
- Gong, X.; Richman, M.B. On the application of cluster analysis to growing season precipitation data in North America east of the rockies. J. Clim. 1995, 8, 897–931. [Google Scholar] [CrossRef] [Green Version]
- Kruse, F.A.; Lefkoff, A.B.; Boardman, J.W.; Heidebrecht, K.B.; Shapiro, A.T.; Barloon, P.J.; Goetz, A.F.H. The spectral image processing system (SIPS)—Interactive visualization and analysis of imaging spectrometer data. Remote Sens. Environ. 1993, 44, 145–163. [Google Scholar] [CrossRef]
- Diao, C.; Wang, L. Landsat time series-based multiyear spectral angle clustering (MSAC) model to monitor the inter-annual leaf senescence of exotic saltcedar. Remote Sens. Environ. 2018, 209, 581–593. [Google Scholar] [CrossRef]
- Aghabozorgi, S.; Shirkhorshidi, A.S.; Wah, T.Y. Time-series clustering—A decade review. Inf. Syst. 2015, 53, 16–38. [Google Scholar] [CrossRef]
- Zhou, Z.H. Machine Learning; Tsinghua University Press: Beijing, China, 2016; pp. 307–309. ISBN 978-7-302-206853-6. [Google Scholar]
- Xing, X.; Yan, C.; Jia, Y.; Jia, H.; Lu, J.; Luo, G. An effective high spatiotemporal resolution ndvi fusion model based on histogram clustering. Remote Sens. 2020, 12, 3774. [Google Scholar] [CrossRef]
- Santos, L.A.; Ferreira, K.; Picoli, M.; Camara, G.; Zurita-Milla, R.; Augustijn, E.W. Identifying spatiotemporal patterns in land use and cover samples from satellite image time series. Remote Sens. 2021, 13, 974. [Google Scholar] [CrossRef]
- Yan, W.J.; Kong, F.H.; Yin, H.W.; Sun, C.F.; Xu, F.; Li, W.C.; Zhang, X.T. Factors affecting the cooling effect in zijin mountain forest park. Acta Ecol. Sin. 2014, 34, 3169–3178. [Google Scholar]
- Li, M.S.; Peng, S.K.; Zhong, H.Y. Landscape pattern and dynamic analysis of zijin mountain scenic area based on GIS. J. Nanjing For. Univ. 2004, 5, 67–70. [Google Scholar]
- Deng, S.; Katoh, M.; Guan, Q.; Yin, N.; Li, M. Interpretation of forest resources at the individual tree level at purple mountain, Nanjing City, China, using worldview-2 imagery by combining GPS, RS and GIS Technologies. Remote Sens. 2014, 6, 87–110. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Katoh, M.; Guan, Q.; Yin, N.; Li, M. Estimating forest aboveground biomass by combining ALOS PALSAR and WorldView-2 data: A case study at purple mountain national park, Nanjing, China. Remote Sens. 2014, 6, 7878–7910. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.M.; Black, T.A. Measuring leaf area index of plant canopies with branch architecture. Agric. For. Meteorol. 1991, 57, 1–12. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M.; Baret, F. Review of methods for in situ leaf area index determination: Part i. theories, sensors and hemispherical photography. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Welles, J.M.; Norman, J.M. Instrument for indirect measurement of canopy architecture. Agron. J. 1991, 83, 818–825. [Google Scholar] [CrossRef]
- Welles, J.M.; Cohen, S. Canopy structure measurement by gap fraction analysis using commercial instrumentation. J. Exp. Bot. 1996, 47, 1335–1342. [Google Scholar] [CrossRef]
- Huang, Y.; Tian, Q.J.; Wang, L.; Geng, J.; Lyu, C. Estimating canopy leaf area index in the late stages of wheat growth using continuous wavelet transform. J. Appl. Remote Sens. 2014, 8, 083517. [Google Scholar] [CrossRef]
- Bai, Z.G. Technical features of gaofen-1 satellite. Aerosp. China 2013, 8, 5–9. [Google Scholar]
- Jia, K.; Liang, S.; Gu, X.; Baret, F.; Wei, X.; Wang, X.; Yao, Y.; Yang, L.; Li, Y. Fractional vegetation cover estimation algorithm for Chinese GF-1 wide field view data. Remote Sens. Environ. 2016, 177, 184–191. [Google Scholar] [CrossRef]
- Yu, J.; Liu, Y.; Ren, Y.; Ma, H.; Wang, D.; Jing, Y.; Yu, L. Application study on double-constrained change detection for land use/land cover based on GF-6 WFV imageries. Remote Sens. 2020, 12, 2943. [Google Scholar] [CrossRef]
- Schlerf, M.; Atzberger, C.; Hill, J. Remote sensing of forest biophysical variables using hymap imaging spectrometer data. Remote Sens. Environ. 2005, 95, 177–194. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.P.; Gao, X.; Lei, J.Q.; Zhou, N.; Wang, Y.D. Spatial and temporal changes in the normalized difference vegetation index and their driving factors in the desert/grassland biome transition zone of the sahel region of africa. Remote Sens. 2020, 12, 4119. [Google Scholar] [CrossRef]
- Jain, A.K. Data clustering: 50 years beyond K-means. Pattern Recognit. Lett. 2010, 31, 651–666. [Google Scholar] [CrossRef]
- Engelen, J.E.; Hoos, H.H. A survey on semi-supervised learning. Mach. Learn. 2020, 109, 373–440. [Google Scholar] [CrossRef] [Green Version]
- Somers, B.; Asner, G.P.; Tits, L.; Coppin, P. Endmember variability in spectral mixture analysis: A review. Remote Sens. Environ. 2011, 115, 1603–1616. [Google Scholar] [CrossRef]
- Basu, S.; Banerjee, A.; Mooney, R. Semi-supervised clustering by seeding. In Proceedings of the 19th International Conference on Machine Learning (ICML-2002), Sydney, NSW, Australia, 8–12 July 2002. [Google Scholar]
- Gara, T.W.; Darvishzadeh, R.; Skidmore, A.K.; Wang, T.; Heurich, M. Evaluating the performance of PROSPECT in the retrieval of leaf traits across canopy throughout the growing season. Int. J. Appl. Earth Obs. Geoinf. 2019, 83, 101919. [Google Scholar] [CrossRef]
- Zeng, L.; Wardlow, B.D.; Xiang, D.; Hu, S.; Li, D. A review of vegetation phenological metrics extraction using time-series, multispectral satellite data. Remote Sens. Environ. 2020, 237, 111511. [Google Scholar] [CrossRef]
- Marino, S.; Alvino, A. Vegetation indices data clustering for dynamic monitoring and classification of wheat yield crop traits. Remote Sens. 2021, 13, 541. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | 08/01/2015 | 08/04/2015 | 26/04/2015 | 12/05/2015 | 31/05/2015 | 09/07/2015 | 15/08/2015 | 10/09/2015 | 28/09/2015 | 16/10/2015 | 01/11/2015 | 16/11/2015 | 11/12/2015 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DOY | 18 | 98 | 116 | 132 | 151 | 190 | 227 | 253 | 271 | 289 | 305 | 320 | 345 |
| Launch date | 26/04/2013 | |
| Payloads | WFV | |
| Spatial resolution (m) | 16 | |
| Wavelength (nm) | Panchromatic: 450–890 nm | |
| Multispectral | Blue: 450–520 Green: 520–590 Red: 630–690 Near-infrared: 770–890 | |
| Swath width (km) | 800 (4 cameras combined) | |
| Average orbit height (km) | 645 | |
| Repetition cycle (day) | 4 | |
| Orbit inclination/Local time of descending node | 98°/11:10 am | |
| Orbit type | Sun-synchronous | |
| Designed lifetime (year) | 5–8 | |
| Number | Image Acquisition Time | Sensor | Forest Growth Stage |
|---|---|---|---|
| 1 | 01/01/2015 | WFV2 | dormancy |
| 2 | 17/01/2015 | WFV1 | dormancy |
| 3 | 19/02/2015 | WFV1 | germination |
| 4 | 12/03/2015 | WFV3 | frondescence |
| 5 | 14/04/2015 | WFV3 | frondescence |
| 6 | 22/04/2015 | WFV2 | frondescence |
| 7 | 26/04/2015 | WFV2 | frondescence |
| 8 | 12/05/2015 | WFV1 | frondescence |
| 9 | 21/05/2015 | WFV4 | frondescence |
| 10 | 14/06/2015 | WFV2 | leaf peak |
| 11 | 02/08/2015 | WFV4 | leaf peak |
| 12 | 02/10/2015 | WFV1 | leaf fall |
| 13 | 15/10/2015 | WFV1 | leaf fall |
| 14 | 01/11/2015 | WFV3 | leaf fall |
| 15 | 03/12/2015 | WFV2 | leaf fall |
| DOY | 18 | 98 | 116 | 132 | 151 | 190 | 227 | 253 | 271 | 289 | 305 | 320 | 345 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | ||||||||||||||
| Cab | 2 | 6 | 17 | 45 | 70 | 85 | 75 | 40 | 25 | 10 | 8 | 5 | 4 | |
| Car | 4 | 4 | 6 | 7 | 8 | 10 | 10 | 12 | 15 | 18 | 19 | 19 | 18 | |
| Cw | 0.005 | 0.007 | 0.01 | 0.01 | 0.02 | 0.04 | 0.03 | 0.02 | 0.015 | 0.01 | 0.008 | 0.007 | 0.006 | |
| Cm | 0.002 | 0.002 | 0.003 | 0.004 | 0.004 | 0.004 | 0.005 | 0.005 | 0.006 | 0.007 | 0.008 | 0.008 | 0.01 | |
| N | 1.5 | |||||||||||||
| Cbrown | 0 | |||||||||||||
| LAI | 0 | 1–1.5 | 1.5–2.5 | 4–5.5 | 6.5–10 | 6.5–9 | 6.5–8 | 5.5–7.5 | 4–5.5 | 3.5–5 | 2.5–3.5 | 1.5–2.5 | 1–2 | |
| ALA | 40 | 70 | 70 | 60 | 50 | 40 | 25 | 25 | 25 | 25 | 40 | 40 | 40 | |
| SZA | 35 | 60 | 68 | 71 | 76 | 75 | 62 | 56 | 54 | 50 | 43 | 40 | 35 | |
| RA | 59 | 126 | 47 | 39 | 137 | 52 | 52 | 52 | 60 | 63 | 106 | 52 | 69 | |
| OZA | 18 | 5 | 3 | 20 | 15 | 28 | 25 | 24 | 24 | 7 | 16 | 15 | 13 | |
| LAIU | 0.1–0.5 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | |
| SD | 400 | |||||||||||||
| H | 15 | |||||||||||||
| CD | 5 | |||||||||||||
| FDR | 0.34 | |||||||||||||
| DOY | 18 | 98 | 116 | 132 | 151 | 190 | 227 | 253 | 271 | 289 | 305 | 320 | 345 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameters | ||||||||||||||
| LT | 1.6 | 1.2 | 1.4 | 1.5 | 1.6 | 1.6 | 1.6 | 1.6 | 1.6 | 1.6 | 1.6 | 1.6 | 1.6 | |
| Cab | 40 | 45 | 50 | 55 | 60 | 70 | 90 | 75 | 70 | 65 | 60 | 55 | 50 | |
| Cw | 55 | 60 | 65 | 70 | 75 | 90 | 100 | 100 | 90 | 80 | 70 | 60 | 55 | |
| LC | 70 | 45 | 45 | 45 | 50 | 55 | 60 | 60 | 60 | 60 | 65 | 70 | 70 | |
| N | 1 | 1 | 1 | 1 | 1.05 | 1.15 | 1.15 | 1.15 | 1.15 | 1.15 | 1.15 | 1 | 1 | |
| CD | 45 | |||||||||||||
| IAS | 0.028 | |||||||||||||
| B | 0.0006 | |||||||||||||
| AA | 2 | |||||||||||||
| LAI | 2–4.5 | 2.5–4.5 | 3–4.5 | 3–5 | 3–5 | 3–5 | 3.5–5 | 3–5 | 3–5 | 3–5 | 3–5 | 3–5 | 2.5–4.5 | |
| LAIU | 0.1 | 0.5 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0.8 | 0.5 | 0.1 | |
| ALA | 40 | 70 | 65 | 55 | 45 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | |
| SZA | 35 | 60 | 68 | 71 | 76 | 75 | 62 | 56 | 54 | 50 | 43 | 40 | 35 | |
| RA | 59 | 126 | 47 | 39 | 137 | 52 | 52 | 52 | 60 | 63 | 106 | 52 | 69 | |
| OZA | 18 | 5 | 3 | 20 | 15 | 28 | 25 | 24 | 24 | 7 | 16 | 15 | 13 | |
| SD | 600 | |||||||||||||
| H | 20 | |||||||||||||
| CD | 4 | |||||||||||||
| FDR | 0.34 | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, R.; Wang, L.; Tian, Q.; Xu, N.; Yang, Y. Estimation of the Conifer-Broadleaf Ratio in Mixed Forests Based on Time-Series Data. Remote Sens. 2021, 13, 4426. https://doi.org/10.3390/rs13214426
Yang R, Wang L, Tian Q, Xu N, Yang Y. Estimation of the Conifer-Broadleaf Ratio in Mixed Forests Based on Time-Series Data. Remote Sensing. 2021; 13(21):4426. https://doi.org/10.3390/rs13214426
Chicago/Turabian StyleYang, Ranran, Lei Wang, Qingjiu Tian, Nianxu Xu, and Yanjun Yang. 2021. "Estimation of the Conifer-Broadleaf Ratio in Mixed Forests Based on Time-Series Data" Remote Sensing 13, no. 21: 4426. https://doi.org/10.3390/rs13214426
APA StyleYang, R., Wang, L., Tian, Q., Xu, N., & Yang, Y. (2021). Estimation of the Conifer-Broadleaf Ratio in Mixed Forests Based on Time-Series Data. Remote Sensing, 13(21), 4426. https://doi.org/10.3390/rs13214426