
Quantifying Biochemical Traits over the Patagonian Sub-Antarctic Forests and Their Relation to Multispectral Vegetation Indices

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Study Area and Methods
2.1. Sample Collection and Laboratory Analysis
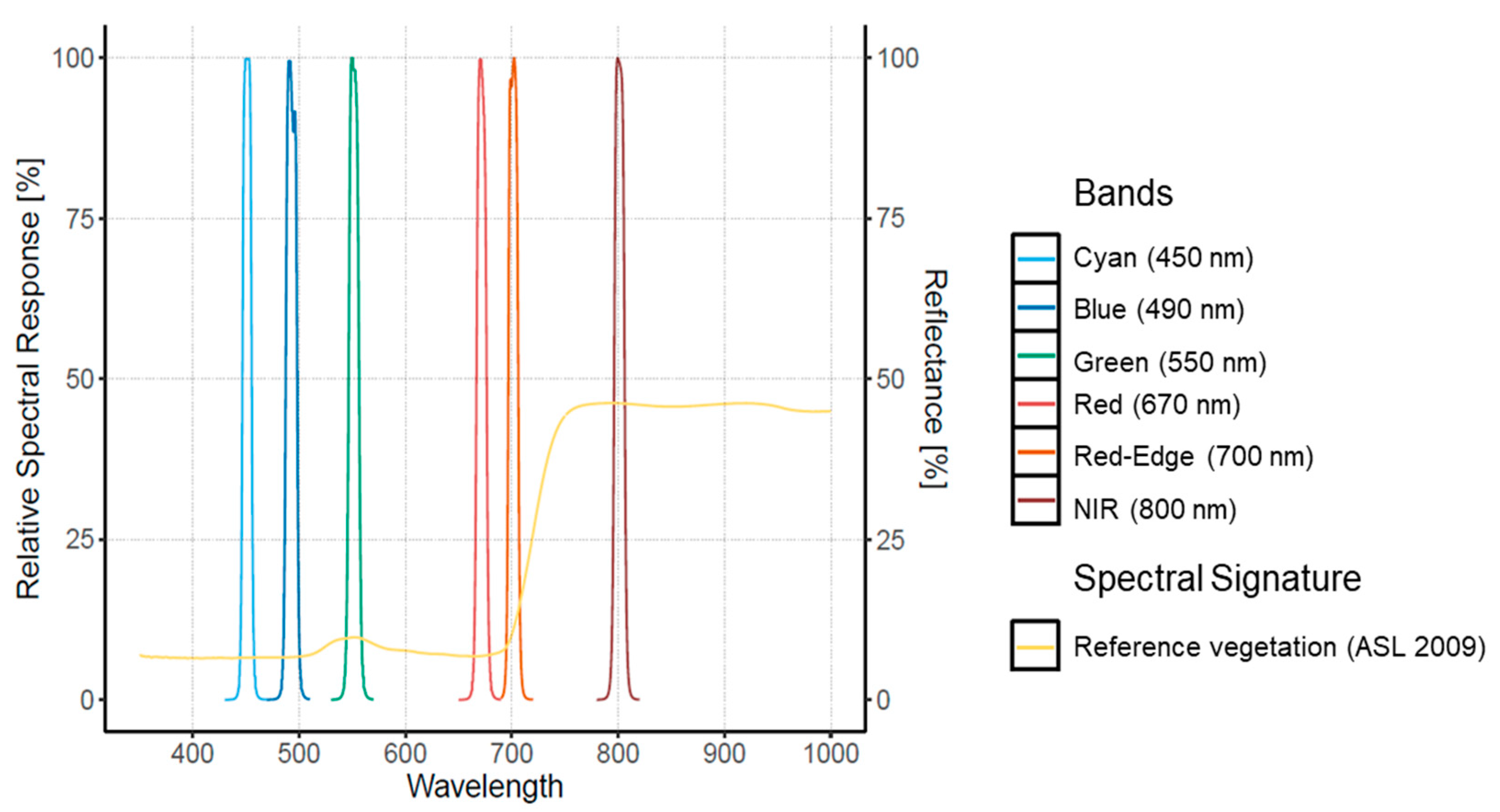
2.2. UAV Data Acquisition
2.3. Imagery Radiometric Calibration and Orthomosaic
2.4. Vegetation Indices
2.5. Extracting Values from Sites
2.6. Statistical Analysis
2.7. Transect and Vertical Profiles
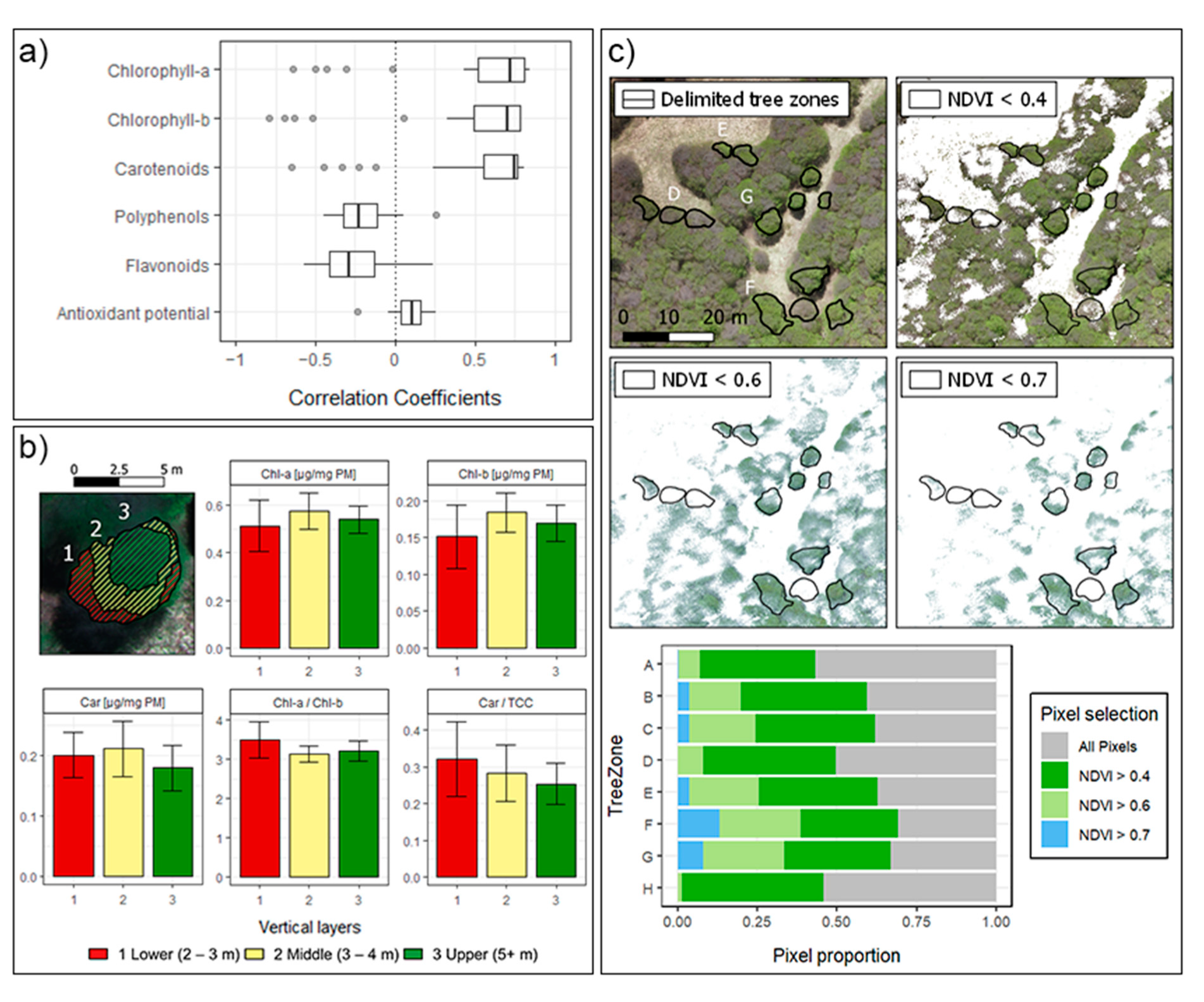
3. Results and Discussion
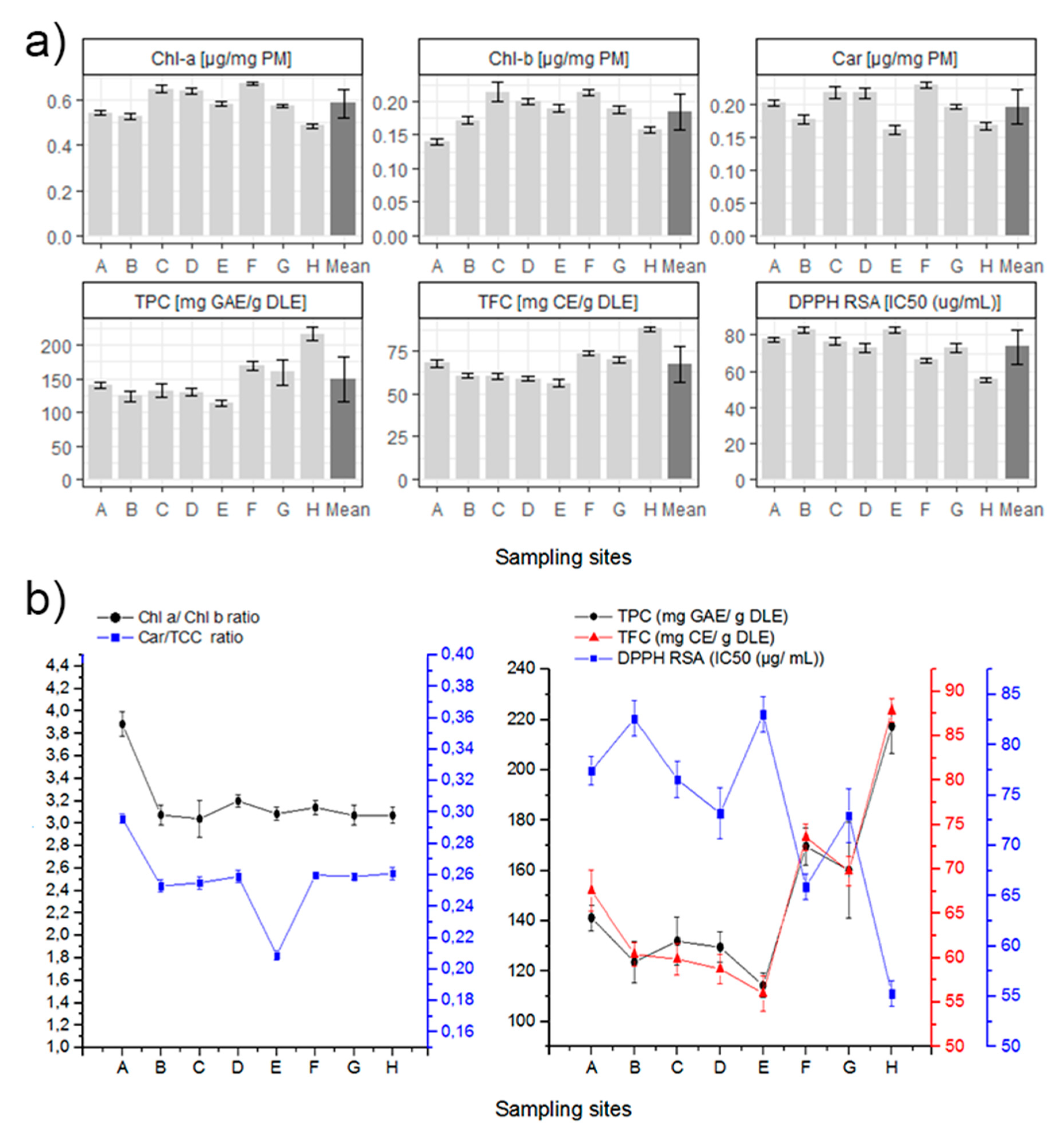
3.1. Biochemical Analysis
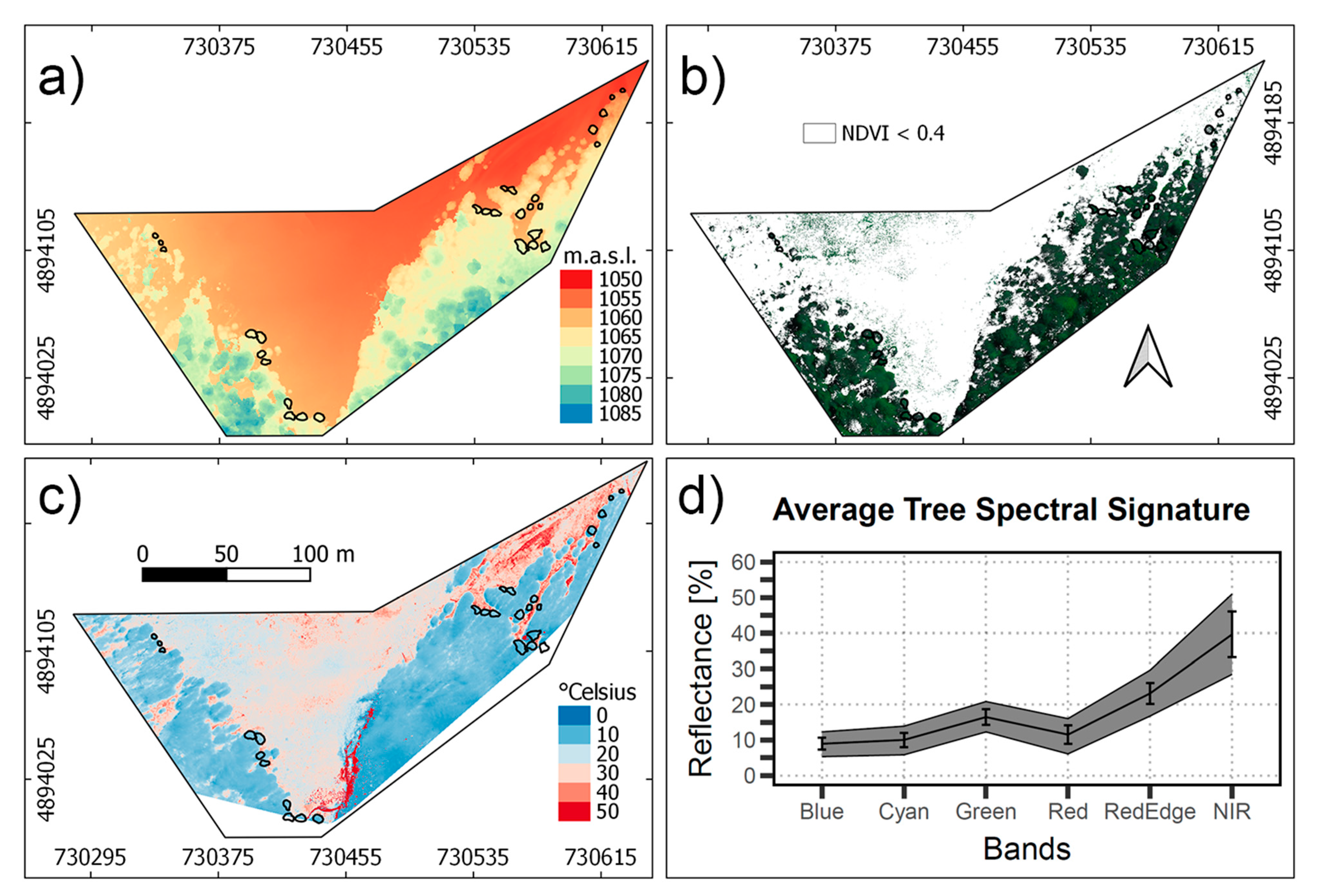
3.2. UAV Data Acquisition
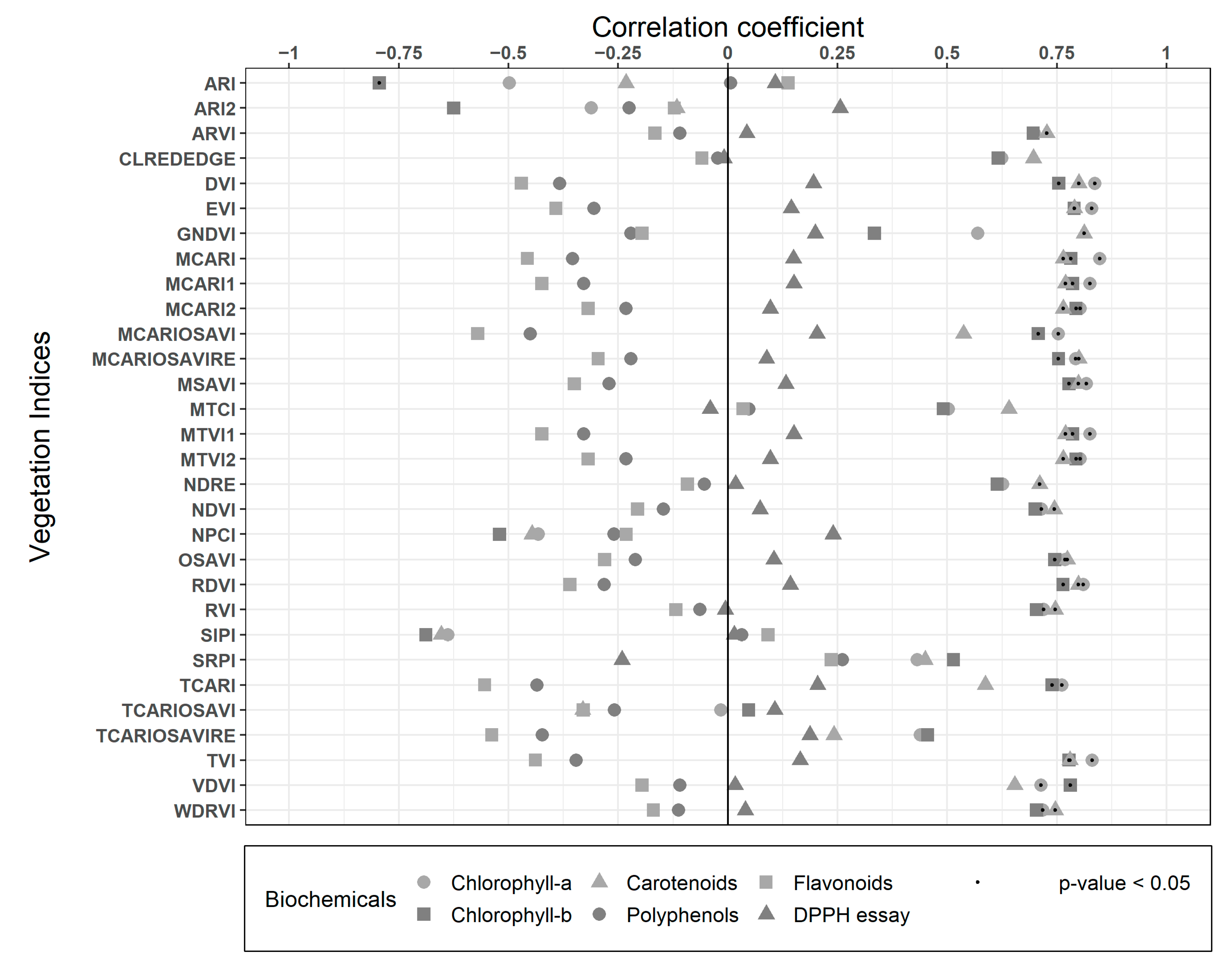
3.3. Statistical Analysis
3.4. Linear Regressions and Spatialization
3.5. Transect
3.6. Study Limitations and Final Remarks
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Acronym | Equation | Source |
|---|---|---|---|
| Normalized Difference Vegetation Index | NDVI | Rouse Jr. et al. [90] | |
| Green NDVI | GNDVI | Gitelson et al. [91] | |
| Normalized Difference Red Edge | NDRE | Gitelson and Merzlyak [92] | |
| Modified Chlorophyll Absorption in Reflectance Index | MCARI | Daughtry et al. [84] | |
| Red Edge Chlorophyll Index | ClRedEdge | Gitelson et al. [93] | |
| Structural Independent Pigment Index | SIPI | J. Peñuelas et al. [94] | |
| Transformed Chlorophyll Absorption in ReflectanceIndex | TCARI | Haboudane et al. [24] | |
| MERIS terrestrial chlorophyll index | MTCI | Dash and Curran [95] | |
| ARVI | where RB = R − (B − R) = | Kaufman and Tanré [96] | |
| Modified Soil-Adjusted Vegetation Index | MSAVI | 0.5[(2 NIR + 1) − Sqrt((2 NIR + 1)2 − 8 (NIR − R))] | Qi et al. [97] |
| Optimized Soil Adjusted Vegetation Index | OSAVI | Rondeaux et al. [98] | |
| Transformed Chlorophyll Absorption in Reflectance Index/Optimized Soil-Adjusted Vegetation Index | TCARI/OSAVI | Haboudane et al. [24] | |
| Modified Chlorophyll Absorption in Reflectance Index/Optimized Soil-Adjusted Vegetation Index | MCARI/OSAVI | Zarco-Tejada et al. [99] | |
| Red Edge-based Transformed Chlorophyll Absorption in Reflectance Index/Optimized Soil-Adjusted Vegetation Index | TCARI/OSAVI_RE | Wu et al. [100] | |
| Red Edge-based Modified Chlorophyll Absorption in Reflectance Index/Optimized Soil-Adjusted Vegetation Index | MCARI/OSAVI_RE | Wu et al. [100] | |
| Ratio Vegetation Index | RVI | Jordan [101] | |
| Difference Vegetation Index | DVI | Tucker [102] | |
| Anthocyanin Reflectance Index | ARI | Gitelson et al. [103] | |
| Anthocyanin Reflectance Index 2 | ARI2 | Gitelson et al. [103] | |
| Simple Ratio Pigment Index | SRPI | Peñuelas et al. [104] | |
| Normalized Pigment Chlorophyll Index | NPCI | Peñuelas et al. [105] | |
| Enhanced Vegetation Index | EVI | Justice et. al. [106] | |
| Visible-Band Difference Vegetation Index | VDVI | Wang et al. [107] | |
| Wide Dynamic Range Vegetation Index | WDRVI | Gitelson [108] | |
| Triangular Vegetation Index | TVI | 0.5 (120 (NIR − G) − 200 (R – G)) | Broge and Leblanc [109] |
| Modified Triangular Vegetation Index 1 | MTVI1 | 1.2 (1.2 (NIR – G) − 2.5 (R − G)) | Haboudane et al. [86] |
| Modified Triangular Vegetation Index 2 | MTVI2 | Haboudane et al. [86] | |
| Modified Chlorophyll Absorption in Reflectance Index 1 | MCARI1 | 1.2 (2.5 (NIR − R) − 1.3 (NIR − G)) | Haboudane et al. [86] |
| Modified Chlorophyll Absorption in Reflectance Index 2 | MCARI2 | Haboudane et al. [86] | |
| Renormalized Difference Vegetation Index | RDVI | Roujean and Breon [110] |
References
- Ramírez, C.; Ortiz, I.; San Matín, C.; Vidal, O.; Álvarez, M.; Pérez, Y.; Solis, J.L.; Alvarez, I. Estudio preliminar de la biodiversidad vegetal terrestre en el Estero Walker (Región de Aysén, Chile): Utilizando líneas base de proyectos de inversión. Gayana Botánica 2014, 71, 227–245. [Google Scholar] [CrossRef][Green Version]
- Bonan, G. Forest and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef]
- Armesto, J.J.; Rozzi, R.; Smith-Ramírez, C.; Arroyo, M.T.K. Conservation targets in South American temperate forests. Science 1998, 282, 1271–1272. [Google Scholar] [CrossRef]
- Olivares-Contreras, V.A.; Mattar, C.; Gutiérrez, A.G.; Jiménez, J.C. Warming trends in Patagonian subantartic forest. Int. J. Appl. Earth Obs. Geoinf. 2019, 76, 51–65. [Google Scholar] [CrossRef]
- Olson, M.; Soriano, D.; Rosell, J.; Anfodillo, T.; Donoghue, M.; Edwards, E.; León-Gómez, C.; Dawson, T.; Camarero Martínez, J.; Castorena, M.; et al. Plant height and hydraulic vulnerability to drought and cold. Proc. Natl. Acad. Sci. USA 2018, 115, 7551–7556. [Google Scholar] [CrossRef]
- Fajardo, A. Are trait-scaling relationships invariant across contrasting elevations in the widely distributed treeline species Nothofagus pumilio? Am. J. Bot. 2016, 103, 821–829. [Google Scholar] [CrossRef]
- Piper, F.; Gundale, M.; Fajardo, A. Extreme defoliation reduces tree growth but not C and N storage in a winter-deciduous species. Ann. Botany 2015, 115, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Piper, F.I.; Fajardo, A.; Cavieres, L.A. Simulated warming does not impair seedling survival and growth of Nothofagus pumilio in the southern Andes. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 97–105. [Google Scholar] [CrossRef]
- Fajardo, A.; Piper, F.; Cavieres, L.A. Distinguishing local from global climate influences in the variation of carbon status with altitude in a tree line species. Global. Ecol. Biogeogr. 2011, 20, 307–318. [Google Scholar] [CrossRef]
- Aldunce, P.; Vicuña, S. Adaptación al cambio climático en Chile: Brechas y recomendaciones. In Informe de las Mesas Adaptación y Agua. Comité Científico COP25; Ministerio de Ciencia, Tecnología, Conocimiento e Innovación: Santiago, Chile, 2019. [Google Scholar]
- Belnap, J.; Phillips, S.L.; Miller, M.E. Response of desert biological soil crusts to alterations in precipitation frequency. Oecologia 2004, 141, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Brand, J.D.; Tang, C.; Graham, R.D. The effect of soil moisture on the tolerance of Lupinus pilosus genotypes to a calcareous soil. Plant Soil. 2000, 219, 263–271. [Google Scholar] [CrossRef]
- Chappelle, E.W.; Kim, M.S.; McMurtrey, J.E. Ratio analysis of reflectance spectra (RARS): An algorithm for the remote estimation of the concentrations of chlorophyll a, chlorophyll B, and carotenoids in soybean leaves. Remote Sens. Environ. 1992, 39, 239–247. [Google Scholar] [CrossRef]
- Zhang, D.; Zhou, X.; Zhang, J.; Lan, Y.; Xu, C.; Liang, D. Detection of rice sheath blight using an unmanned aerial system with high resolution color and multispectral imaging. PLoS ONE 2018, 13, e0187470. [Google Scholar] [CrossRef]
- Gago, J.; Douthe, C.; Coopman, R.E.; Gallego, P.P.; Ribas-Carbo, M.; Flexas, J.; Escalona, J.; Medrano, H. UAVs challenge to assess water stress for sustainable agriculture. Agric. Water Manag. 2015, 153, 9–19. [Google Scholar] [CrossRef]
- Sankaran, S.; Khot, L.R.; Espinoza, C.Z.; Jarolmasjed, S.; Sathuvalli, V.R.; Vandemark, G.J.; Miklas, P.N.; Carter, A.H.; Pumphrey, M.O.; Knowles, R.R.N.; et al. Low-altitude, high-resolution aerial imaging systems for row and field crop phenotyping: A review. Eur. J. Agron. 2015, 70, 112–123. [Google Scholar] [CrossRef]
- Van Der Meij, B.; Kooistra, L.; Suomalainen, J.; Barel, J.M.; De Deyn, G.B. Remote sensing of plant trait responses to field-based plant-soil feedback using UAV-based optical sensors. Biogeosciences 2017, 14, 733–749. [Google Scholar] [CrossRef]
- Verger, A.; Vigneau, N.; Chéron, C.; Gilliot, J.M.; Comar, A.; Baret, F. Green area index from an unmanned aerial system over wheat and rapeseed crops. Remote Sens. Environ. 2014, 152, 654–664. [Google Scholar] [CrossRef]
- Zaman-Allah, M.; Vergara, O.; Araus, J.L.; Tarekegne, A.; Magorokosho, C.; Zarco-Tejada, P.J.; Hornero, A.; Albà, A.H.; Das, B.; Craufurd, P.; et al. Unmanned aerial platform-based multi-spectral imaging for field phenotyping of maize. Plant Methods 2015, 11, 35. [Google Scholar] [CrossRef] [PubMed]
- Zarco-Tejada, P.J.; Guillén-Climent, M.L.; Hernández-Clemente, R.; Catalina, A.; González, M.R.; Martín, P. Estimating leaf carotenoid content in vineyards using high resolution hyperspectral imagery acquired from an unmanned aerial vehicle (UAV). Agric. For. Meteorol. 2013, 171, 281–294. [Google Scholar] [CrossRef]
- Durán, S.; Martin, R.E.; Díaz, S.; Maitner, B.; Malhi, Y.; Salinas, N.; Shenkin, A.; Silman, M.; Wieczynski, D.J.; Asner, G.P.; et al. Informing trait based ecology by assessing remotely sensed functional diversity across broad tropical tefajardomperature gradient. Science Adv. 2019, 5, eaaw8114. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Knapp, D.E.; Tupayachi, R.; Anderson, C.B.; Sinca, F.; Vaughn, N.R.; Llactayo, W. Airborne laser-guided imaging spectroscopy to map forest trait diversity and guide conservation. Science 2017, 355, 385–389. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Anderson, C.; Kryston, K.; Vaughn, N.; Knapp, D.; Bentley, L.; Shenkin, A.; Salinas, N.; Sinca, F.; et al. Scale dependence of canopy trait distributions along a tropical forest elevation gradient. New Phytol. 2016, 214, 973–988. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Tremblay, N.; Zarco-Tejada, P.J.; Dextraze, L. Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote Sens. Environ. 2002, 81, 416–426. [Google Scholar] [CrossRef]
- Le Maire, G.; Francois, C.; Dufrene, E. Towards universal broad leaf chlorophyll indices using PROSPECT simulated database and hyperspectral reflectance measurements. Remote Sens. Environ. 2004, 89, 1–28. [Google Scholar] [CrossRef]
- Malenovský, Z.; Homolová, L.; Zurita-Milla, R.; Lukeš, P.; Kaplan, V.; Hanuš, J.; Gastellu-Etchegorry, J.P.; Schaepman, M.E. Retrieval of spruce leaf chlorophyll content from airborne image data using continuum removal and radiative transfer. Remote Sens. Environ. 2013, 131, 85–102. [Google Scholar] [CrossRef]
- Schlerf, M.; Atzberger, C.; Hill, J.; Buddenbaum, H.; Werner, W.; Schuler, G. Retrieval of chlorophyll and nitrogen in Norway spruce (Picea abies L. Karst.) using imaging spectroscopy. Int. J. Appl. Earth Obs. Geoinf. 2010, 12, 17–26. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Harron, J.; Hu, B.; Noland, T.L.; Goel, N.; Mohammed, G.H.; Sampson, P. Needle chlorophyll content estimation through model inversion using hyperspectral data from boreal conifer forest canopies. Remote Sens. Environ. 2004, 89, 189–199. [Google Scholar] [CrossRef]
- Gitelson, A.A. Assessing carotenoid content in plant leaves with reflectance spectroscopy. Photochem. Photobiol. 2002, 75, 272–281. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Keydan, G.P.; Merzlyak, M.N. Three-band model for noninvasive estimation of chlorophyll, carotenoids, and anthocyanin contents in higher plant leaves. Geophys. Res. Lett. 2006, 33, L11402. [Google Scholar] [CrossRef]
- Hernandez-Clemente, R.; Navarro-Cerrillo, R.M.; Zarco-Tejada, P.J. Carotenoid content estimation in a heterogeneous conifer forest using narrow-band indices and PROSPECT + DART simulations. Remote Sens. Environ. 2012, 127, 298–315. [Google Scholar] [CrossRef]
- Kokaly, R.F.; Skidmore, A.K. Plant phenolics and absorption features in vegetation reflectance spectra near 1.66 microns. Int. J. Appl. Earth Obs. Geoinf. (JAG) 2015, 43, 55–83. [Google Scholar] [CrossRef]
- Clevers, J.G.P.W.; Kooistra, L.; Schaepman, M.E. Estimating canopy water content using hyperspectral remote sensing data. Int. J. Appl. Earth Obs. Geoinf. 2010, 12, 119–125. [Google Scholar] [CrossRef]
- Colombo, R.; Meroni, M.; Marchesi, A.; Busetto, L.; Rossini, M.; Giardino, C.; Panigada, C. Estimation of leaf and canopy water content in poplar plantations by means of hyperspectral indices and inverse modeling. Remote Sens. Environ. 2008, 112, 1820–1834. [Google Scholar] [CrossRef]
- Kokaly, R.F.; Asner, G.P.; Ollinger, S.V.; Martin, M.E.; Wessman, C.A. Characterizing canopy biochemistry from imaging spectroscopy and its application to ecosystem studies. Remote Sens. Environ. 2009, 113, S78–S91. [Google Scholar] [CrossRef]
- Mutanga, O.; Skidmore, A.K.; Prins, H.H.T. Predicting in situ pasture quality in the Kruger National Park, South Africa, using continuum-removed absorption features. Remote Sens. Environ. 2004, 89, 393–408. [Google Scholar] [CrossRef]
- Pimstein, A.; Karnieli, A.; Bansal, S.K.; Bonfil, D.J. Exploring remotely sensed technologies for monitoring wheat potassium and phosphorus using field spectroscopy. Field Crop. Res. 2011, 121, 125–135. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant remote sensing vegetation indices: A review of developments and applications. J. Sens. 2017, 2017, 17. [Google Scholar] [CrossRef]
- Sylvester, G.E. Agriculture in Action: Drones for Agriculture; FAO (Food and Agriculture Organization of the United Nations) and ITU (International Telecommunication Union): Bangkok, Thailand, 2018; p. 126. [Google Scholar]
- Houborg, R.; Fisher, J.; Skidmore, A. Advances in remote sensing of vegetation function and traits. Int. J. Appl. Earth Obs. Geoinf. 2015, 43, 1–6. [Google Scholar] [CrossRef]
- Almeyda, E.; Sáez, F. Recopilación de Datos Climáticos de Chile y Mapas Sinópticos Respectivos; Ministerio de Agricultura: Santiago, Chile, 1958. [Google Scholar]
- Amigo, J.; Ramírez, C. A bioclimatic classification of Chile: Woodland communities in the temperate zone. Plant Ecol. 1998, 136, 9–26. [Google Scholar] [CrossRef]
- DGA (Dirección General de Aguas). Informe Meteorológico de Chile; Dirección General de Agua: Santiago, Chile, 2008. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A.; Richardson, K.; WorldClim. Global Climate Data. 2005. Available online: http://www.worldclim.org (accessed on 25 March 2021).
- Veblen, T.T.; Donoso, C.; Kitzberger, T.; Rebertus, A. Ecology of Southern Chilean and Argentinean Nothofagus forests. In The Ecology and Biogeography of Nothofagus Forests; Veblen, T.T., Hill, R.S., Read, J., Eds.; Yale University Press: New Haven, CT, USA, 1996; pp. 293–353. [Google Scholar]
- Premoli, A.C.; Raffaele, E.; Mathiassen, P. Morphological and phenological differences in Nothofagus pumilio from contrasting elevations: Evidence from a common garden. Aust. Ecol. 2007, 32, 515–523. [Google Scholar] [CrossRef]
- Premoli, A.C.; Brewer, C.A. Environmental versus genetically driven variation in ecophysiological traits of Nothofagus pumilio from contrasting elevations. Aust. J. Bot. 2007, 55, 585–591. [Google Scholar] [CrossRef]
- Fajardo, A.; Piper, F. Intraspecific trait variation and covariation in a widespread trees species (Nothofagus pumilio) in southern Chile. New Phytol. 2011, 189, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Rebertus, A.; Veblen, T.T. Structure and tree-fall gap dynamics of old-growth Nothofagus forests in Tierra del Fuego, Argentina. J. Veg. Sci. 1993, 4, 461–654. [Google Scholar] [CrossRef]
- González, M.E. Fire History of Araucaria-Nothofagus Forests in the Andean Cordillera of South-Central Chile; University of Colorado: Boulder, CO, USA, 2002; p. 158. [Google Scholar]
- Parodi, L.R. Enciclopedia Argentina de Agricultura y Jardinería; Editorial ACME, S.A.C.I.: Buenos Aires, Argentina, 1987. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Bridi, R.; Atala, E.; Pizarro, P.N.; Montenegro, G. Honey bee pollen Load: Phenolic composition and antimicrobial activity and antioxidant capacity. J. Nat. Prod. 2019, 82, 559–565. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Ben Mrid, R.; Bouchmaa, N.; Bouargalne, Y.; Ramdan, B.; Karrouchi, K.; Kabach, I.; El Karbane, M.; Idir, A.; Zyad, A.; Nhiri, M. Phytochemical characterization, antioxidant and in vitro cytotoxic activity evaluation of Juniperus oxycedrus Subsp. oxycedrus needles and berries. Molecules 2019, 24, 502. [Google Scholar] [CrossRef]
- Hartwig, V.G.; Brumovsky, L.A.; Fretes, R.M.; Boado, L.S. A novel procedure to measure the antioxidant capacity of Yerba maté extracts. Food Sci. Technol. 2012, 32, 126–133. [Google Scholar] [CrossRef]
- Kumar, L.; Schmidt, K.; Dury, S.J.; Skidmore, A. Imaging spectrometry and vegetation sciences. In Imaging Spectrometry. Basic Principles and Prospective Applications; Van der Meer, F.D., de Jong, S.M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 111–155. [Google Scholar]
- Disney, M.; Lewis, P.; Saich, P. 3D modelling of forest canopy structure for remote sensing simulations in the optical and microwave domains. Remote Sens. Environ. 2006, 100, 114–132. [Google Scholar] [CrossRef]
- Homolová, L.; Malenovský, Z.; Clevers, J.; García-Santos, G.; Schaepman, M. Review of optical-based remote sensing for plant trait mapping. Ecol. Complex. 2013, 15, 1–16. [Google Scholar] [CrossRef]
- Fernandes, R.A.; Miller, J.R.; Chen, J.M.; Rubinstein, I.G. Evaluating image-based estimates of leaf area index in boreal conifer stands over a range of scales using high-resolution CASI imagery. Remote Sens. Environ. 2004, 89, 200–216. [Google Scholar] [CrossRef]
- Turner, D.P.; Cohen, W.B.; Kennedy, R.E.; Fassnacht, K.S.; Briggs, J.M. Relationships between leaf area index and Landsat TM spectral vegetation indices across three temperate zone sites. Remote Sens. Environ. 1999, 70, 52–68. [Google Scholar] [CrossRef]
- Xiao, X.; Hollinger, D.; Aber, J.; Goltz, M.; Davidson, E.A.; Zhang, Q.; Moore, B., III. Satellite-based modeling of gross primary production in an evergreen needleleaf forest. Remote Sens. Environ. 2004, 89, 519–534. [Google Scholar] [CrossRef]
- Yi, Q.X.; Huang, J.F.; Wang, F.M.; Wang, X.Z. Quantifying biochemical variables of corn by hyperspectral reflectance at leaf scale. J. Zhejiang Univ. Sci. B 2008, 9, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Peng-Sen, S.; Liu, S.-R. A review of plant spectral reflectance response to water physiological changes. Chin. J. Plant Ecol. 2016, 40, 80–91. [Google Scholar] [CrossRef]
- Sun, J.; Shi, S.; Gong, W.; Yang, J.; Du, L.; Song, S.; Chen, B.; Zhang, Z. Evaluation of hyperspectral LiDAR for monitoring rice leaf nitrogen by comparison with multispectral LiDAR and passive spectrometer. Sci. Rep. 2017, 7, 40362. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.; Martin, R.E.; Knapp, D.; Tupayachi, R.; Anderson, C.; Carranza, L.; Martinez, P.; Houcheime, M.; Sinca, F.; Weiss, P. Spectroscopy of canopy chemicals in humid tropical forests. Remote Sens. Environ. 2011, 115, 3587–3598. [Google Scholar] [CrossRef]
- Jay, S.; Baret, F.; Dutartre, D.; Malatesta, G.; Héno, S.; Comar, A.; Weiss, M.; Maupas, F. Exploiting the centimeter resolution of UAV multispectral imagery to improve remote-sensing estimates of canopy structure and biochemistry in sugar beet crops. Rem. Sens. Environ. 2019, 231, 110898. [Google Scholar] [CrossRef]
- Ollinger, S.V.; Smith, M.L.; Martin, M.E.; Hallett, R.A.; Goodale, C.L.; Aber, J.D. Regional variation in foliar chemistry and N cycling among forests of diverse history and composition. Ecology 2002, 83, 339–355. [Google Scholar]
- Smith, M.L.; Ollinger, S.V.; Martin, M.E.; Aber, J.D.; Hallett, R.A.; Goodale, C.L. Direct estimation of aboveground forest productivity through hyperspectral remote sensing of canopy nitrogen. Ecol. Appl. 2002, 12, 1286–1302. [Google Scholar] [CrossRef]
- Smith, M.L.; Martin, M.E.; Plourde, L.; Ollinger, S.V. Analysis of hyperspectral data for estimation of temperate forest canopy nitrogen concentration: Comparison between airborne (AVIRIS) and spaceborne (Hyperion) sensor. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1332–1337. [Google Scholar] [CrossRef]
- Asner, G.; Martin, R.E. Spectral and chemical analysis of tropical forests: Scaling from leaf to canopy levels. Remote Sens. Environ. 2008, 112, 3958–3970. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C.; Döll, M.; Fietz, H.J.; Bach, T.; Kozel, U.; Meir, D.; Rahmsdorf, U. Photosynthetic activity, chloroplasts ultrastructure, and leaf characteristics of high-light and low-light plants and of sun and shade leaves. Photosynth. Res. 1981, 2, 115–141. [Google Scholar] [CrossRef]
- Kitajima, K.; Hogan, K.P. Increases of chlorophyll a/b ratios during acclimation of tropical woody seedlings to nitrogen limitation and high light. Plant Cell Environ. 2003, 26, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, G.C.; Stamatakis, K. Water and solute transport in cyanobacteria as probed by chlorophyll fluorescence. In Chlorophyll a Fluorescence: A Signature of Photosynthesis, Advances in Photosynthesis and Respiration; Sharkey, T.D., Ed.; Springer: Dordrecht, Switzerland, 2004; Volume 19, pp. 663–678. [Google Scholar]
- Yamazaki, J.; Suzuki, T.; Maruta, E.; Kamimura, Y. The stoichiometry and antenna size of the two photosystems in marine green algae, Bryopsis maxima and Ulva pertusa, in relation to the light environment of their natural habitat. J. Exp. Bot. 2005, 56, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T. The Effect of Light Exposure on the Total Chlorophyll Content, Chl a/b ratio, and Car/chl Ratio in the Barks of Fraxinus latifolia Seedlings. University Honors, Bachelor of Science Thesis, Portland State University, Porland, OR, USA, 2018. [Google Scholar]
- Levizou, E.; Petropoulou, Y.; Manetas, Y. Carotenoid composition of peridermal twigs does not fully conform to a shade acclimation hypothesis. Photosynthetica 2004, 42, 591–596. [Google Scholar] [CrossRef]
- Molyneux, P. The use of stable free radical Diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Jacquemoud, S.; Verhoef, W.; Baret, F.; Bacour, C.; Zarco-Tejada, P.J.; Asner, G.P.; Francois, C.; Ustin, S.L. PROSPECT + SAIL models: A review of use for vegetation characterization. Remote Sens. Environ. 2009, 113, S56–S66. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; Brown de Colstoun, E.; McMurtrey, J.E. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Asner, G.; Anderson, C.; Martin, R.; Tupayachi, R.; Knapp, D.E.; Sinca, F. Landscape biogeochemistry reflected in shifting distributions of chemical traits in the Amazon Forest canopy. Nat. Geosci. 2015, 8, 567–573. [Google Scholar] [CrossRef]
- Potgieter, A.; George-Jaeggli, B.; Chapman, S.; Laws, K.; Suárez, L.; Wixted, J.; Watson, J.; Eldridge, M.; Jordan, D.; Hammer, G. Multi-spectral imaging from an unmanned aerial vehicle enables the assessment of seasonal leaf area dynamics of sorghum breeding lines. Front. Plant Sci. 2017, 8, 1532. [Google Scholar] [CrossRef]
- Shen, X.; Cao, L.; Coops, N.; Fan, H.; Wu, X.; Liu, H.; Wang, G.; Cao, F. Quantifying vertical profiles of biochemical traits for forest plantation species using advanced remote sensing approaches. Remote Sens. Environ. 2020, 250, 112041. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. In Proceedings of the Third ERTS Symposium, Washington, DC, USA, 10–14 December 1973; NASA Special Publication-351. US. Government Printing Office: Washington, DC, USA, 1973; Volume 1, pp. 309–317. [Google Scholar]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Remote estimation of chlorophyll content in higher plant leaves. Int. J. Remote Sens. 1997, 18, 2691–2697. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Viña, A.; Ciganda, V.; Rundquist, D.C.; Arkebauer, T.J. Remote estimation of canopy chlorophyll content in crops. Geophys. Res. Lett. 2005, 32, L08403. [Google Scholar] [CrossRef]
- Peñuelas, J.; Baret, F.; Filella, I. Semi-empirical indices to assess carotenoids/chlorophyll-a ratio from leaf spectral reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Dash, J.; Curran, P.J. The MERIS terrestrial chlorophyll index. Int. J. Remote Sens. 2004, 25, 5403–5413. [Google Scholar] [CrossRef]
- Kaufman, Y.J.; Tanré, D. Atmospherically Resistant Vegetation Index (ARVI) for EOS-MODIS. IEEE Trans. Geosci. Remote Sens. 1992, 30, 261–270. [Google Scholar] [CrossRef]
- Qi, J.; Chehbouni, A.; Huete, A.R.; Kerr, Y.H.; Sorooshian, S. A modified soil adjusted vegetation index. Remote Sens. Environ. 1994, 48, 119–126. [Google Scholar] [CrossRef]
- Rondeaux, G.; Steven, M.; Baret, F. Optimization of soil-adjusted vegetation indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Berjón, A.; López-Lozano, R.; Miller, J.R.; Martín, P.; Cachorro, P.; González, M.R.; de Frutos, A. Assessing vineyard condition with hyperspectral indices: Leaf and canopy reflectance simulation in a row-structured discontinuous canopy. Remote Sens. Environ. 2005, 99, 271–287. [Google Scholar] [CrossRef]
- Wu, C.; Niu, Z.; Tang, Q.; Huang, W. Estimating chlorophyll content from hyperspectral vegetation indices: Modeling and validation. Agric. For. Meteorol. 2008, 148, 1230–1241. [Google Scholar] [CrossRef]
- Jordan, C.F. Derivation of leaf-area index from quality of light on the forest floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N.; Chivkunova, O.B. Optical properties and nondestructive estimation of anthocyanin content in plant leaves. Photochem. Photobiol. 2001, 74, 38–45. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Lloret, P.; Muñoz, F.; Vilajeliu, M. Reflectance assessment of mite effects on apple trees. Int. J. Remote Sens. 1995, 16, 2727–2733. [Google Scholar] [CrossRef]
- Peñuelas, J.; Gamon, J.A.; Fredeen, A.L.; Merino, J.; Field, C.B. Reflectance indices associated with physiological changes in nitrogen- and water-limited sunflower leaves. Remote Sens. Environ. 1994, 48, 135–146. [Google Scholar] [CrossRef]
- Justice, C.O.; Vermote, E.; Townshend, J.R.G.; Defries, R.; Roy, D.P.; Hall, D.K.; Salomonson, V.V.; Privette, J.L.; Riggs, G.; Strahler, A.; et al. The Moderate Resolution Imaging Spectroradiometer (MODIS): Land remote sensing for global change research. IEEE Trans. Geosci. Remote Sens. 1998, 36, 1228–1249. [Google Scholar] [CrossRef]
- Wang, X.; Wang, M.; Wang, S.; Wu, Y. Extraction of vegetation information from visible unmanned aerial vehicle images. Nongye Gongcheng Xuebao/Trans. Chin. Soc. Agric. Eng. 2015, 31, 152–159. [Google Scholar] [CrossRef]
- Gitelson, A.A. Wide dynamic range vegetation index for remote quantification of biophysical characteristics of vegetation. J. Plant Physiol. 2004, 161, 165–173. [Google Scholar] [CrossRef]
- Broge, N.H.; Leblanc, E. Comparing prediction power and stability of broadband and hyperspectral vegetation indices for estimation of green leaf area index and canopy chlorophyll density. Remote Sens. Environ. 2001, 76, 156–172. [Google Scholar] [CrossRef]
- Roujean, J.L.; Breon, F.M. Estimating PAR absorbed by vegetation from bidirectional reflectance measurements. Remote Sens. Environ. 1995, 51, 375–384. [Google Scholar] [CrossRef]









| Bands | Chlorophyll-a | Chlorophyll-b | Carotenoids | Polyphenols | Flavonoids | DPPH Essay |
|---|---|---|---|---|---|---|
| Blue [450 nm] | −0.50 | −0.79 ** | −0.23 | 0.01 | 0.14 | 0.11 |
| Cyan [490 nm] | −0.31 | −0.63 * | −0.12 | −0.22 | −0.12 | 0.26 |
| Green [550 nm] | 0.70 ** | 0.70 * | 0.73 ** | −0.11 | −0.17 | 0.04 |
| Red [670 nm] | 0.62 * | 0.62 * | 0.70 * | −0.02 | −0.06 | −0.01 |
| Red Edge [700 nm] | 0.84 ** | 0.75 ** | 0.80 ** | −0.38 | −0.47 | 0.20 |
| NIR [800 nm] | 0.83 ** | 0.79 ** | 0.79 ** | −0.31 | −0.39 | 0.14 |
| Biochemical Trait | VI Parameter and Coefficient | R2 | p-Value |
|---|---|---|---|
| Chl-a | 1.0649*MCARI + 0.3108 | 0.72 | <0.01 |
| Chl-b | 0.3295*EVI-0.003 | 0.62 | <0.01 |
| Car | 0.8723*GNDVI-0.1702 | 0.66 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taylor-Zavala, R.; Ramírez-Rodríguez, O.; de Armas-Ricard, M.; Sanhueza, H.; Higueras-Fredes, F.; Mattar, C. Quantifying Biochemical Traits over the Patagonian Sub-Antarctic Forests and Their Relation to Multispectral Vegetation Indices. Remote Sens. 2021, 13, 4232. https://doi.org/10.3390/rs13214232
Taylor-Zavala R, Ramírez-Rodríguez O, de Armas-Ricard M, Sanhueza H, Higueras-Fredes F, Mattar C. Quantifying Biochemical Traits over the Patagonian Sub-Antarctic Forests and Their Relation to Multispectral Vegetation Indices. Remote Sensing. 2021; 13(21):4232. https://doi.org/10.3390/rs13214232
Chicago/Turabian StyleTaylor-Zavala, Richard, Oney Ramírez-Rodríguez, Merly de Armas-Ricard, Harold Sanhueza, Francisca Higueras-Fredes, and Cristian Mattar. 2021. "Quantifying Biochemical Traits over the Patagonian Sub-Antarctic Forests and Their Relation to Multispectral Vegetation Indices" Remote Sensing 13, no. 21: 4232. https://doi.org/10.3390/rs13214232
APA StyleTaylor-Zavala, R., Ramírez-Rodríguez, O., de Armas-Ricard, M., Sanhueza, H., Higueras-Fredes, F., & Mattar, C. (2021). Quantifying Biochemical Traits over the Patagonian Sub-Antarctic Forests and Their Relation to Multispectral Vegetation Indices. Remote Sensing, 13(21), 4232. https://doi.org/10.3390/rs13214232