
Enhancing Animal Movement Analyses: Spatiotemporal Matching of Animal Positions with Remotely Sensed Data Using Google Earth Engine and R



Abstract
:
1. Introduction
2. Materials and Methods
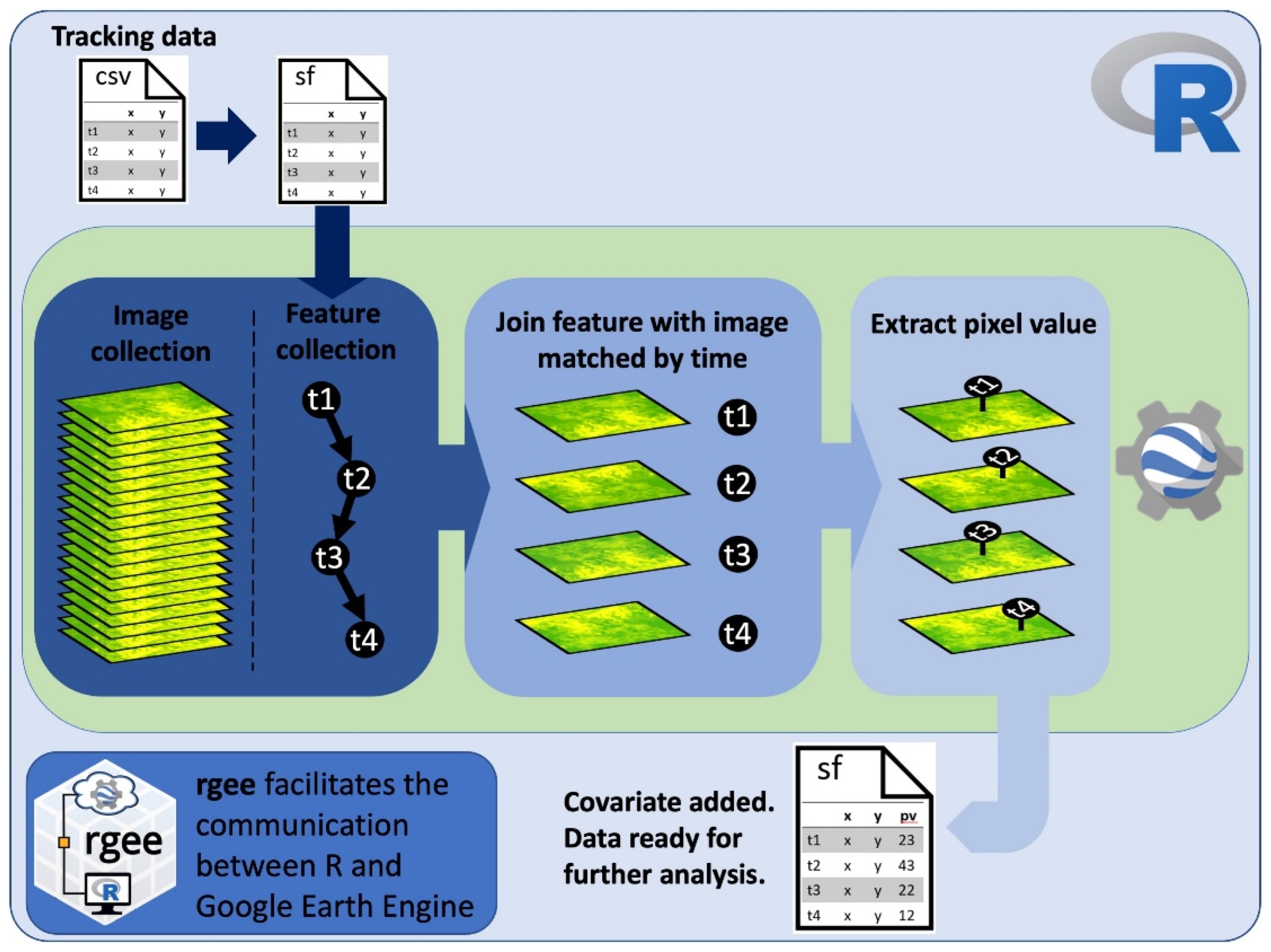
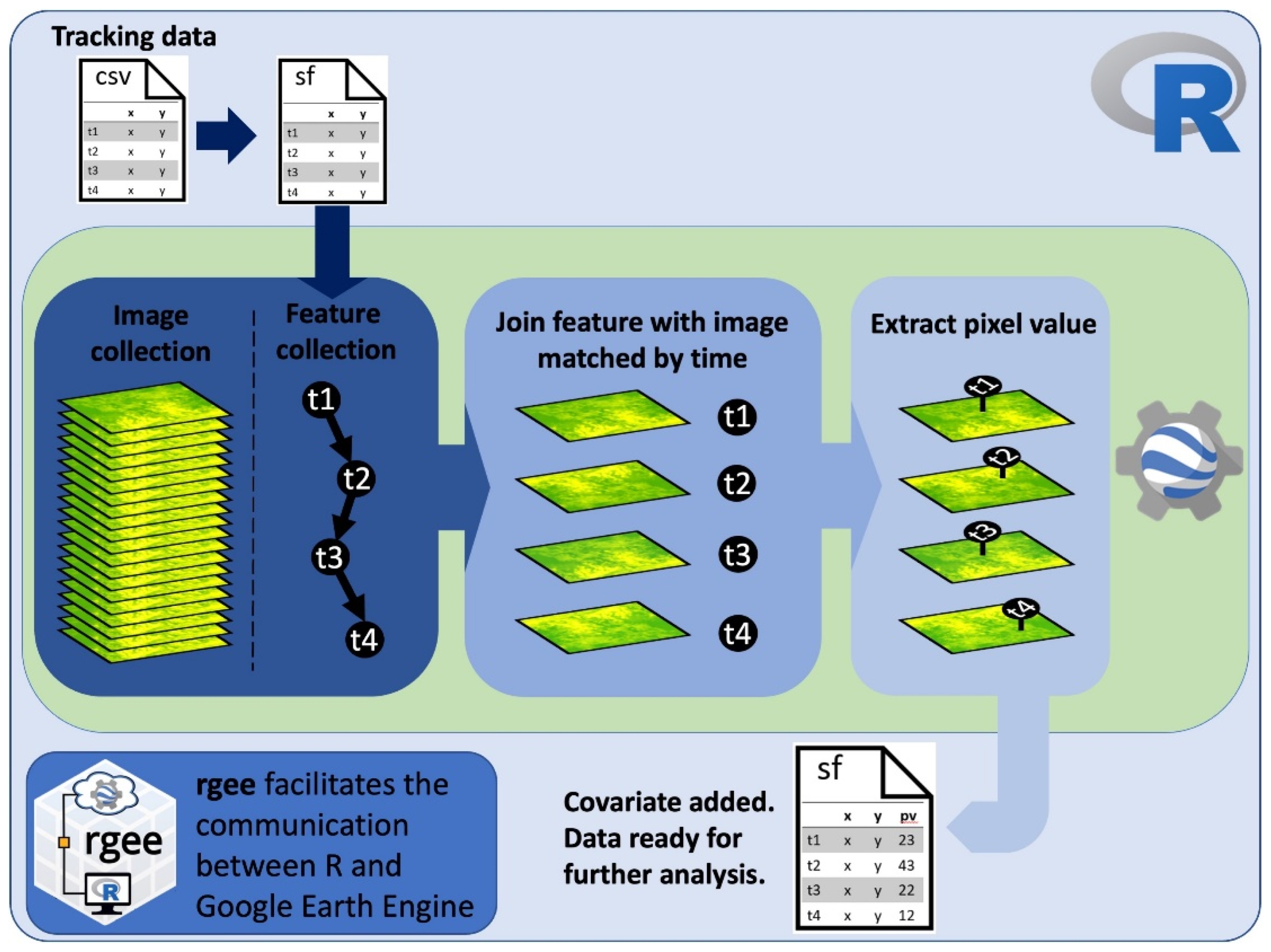
2.1. Code Workflow
2.2. Animal-Tracking Data for Case Studies
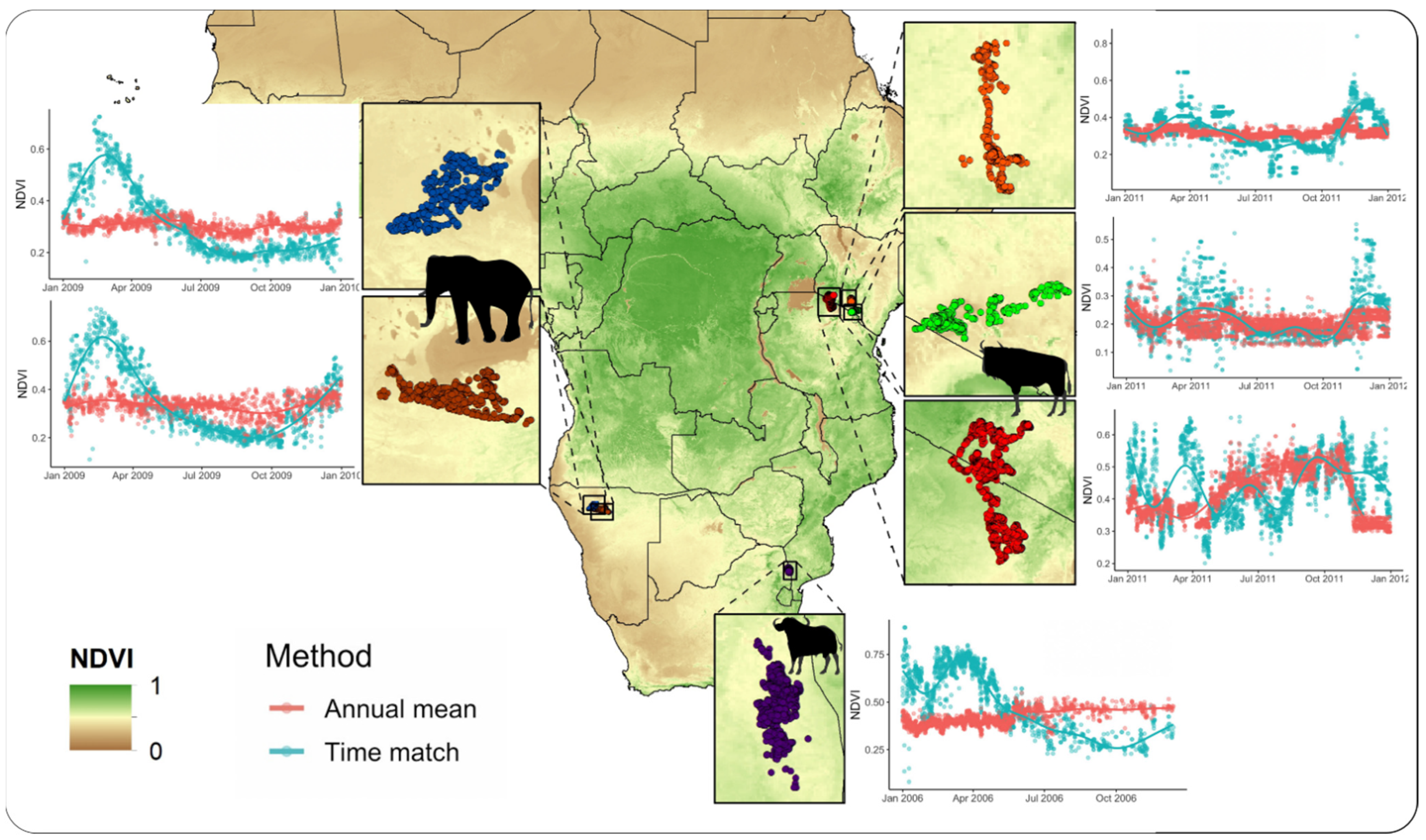
2.3. Case Study 1—NDVI Extraction
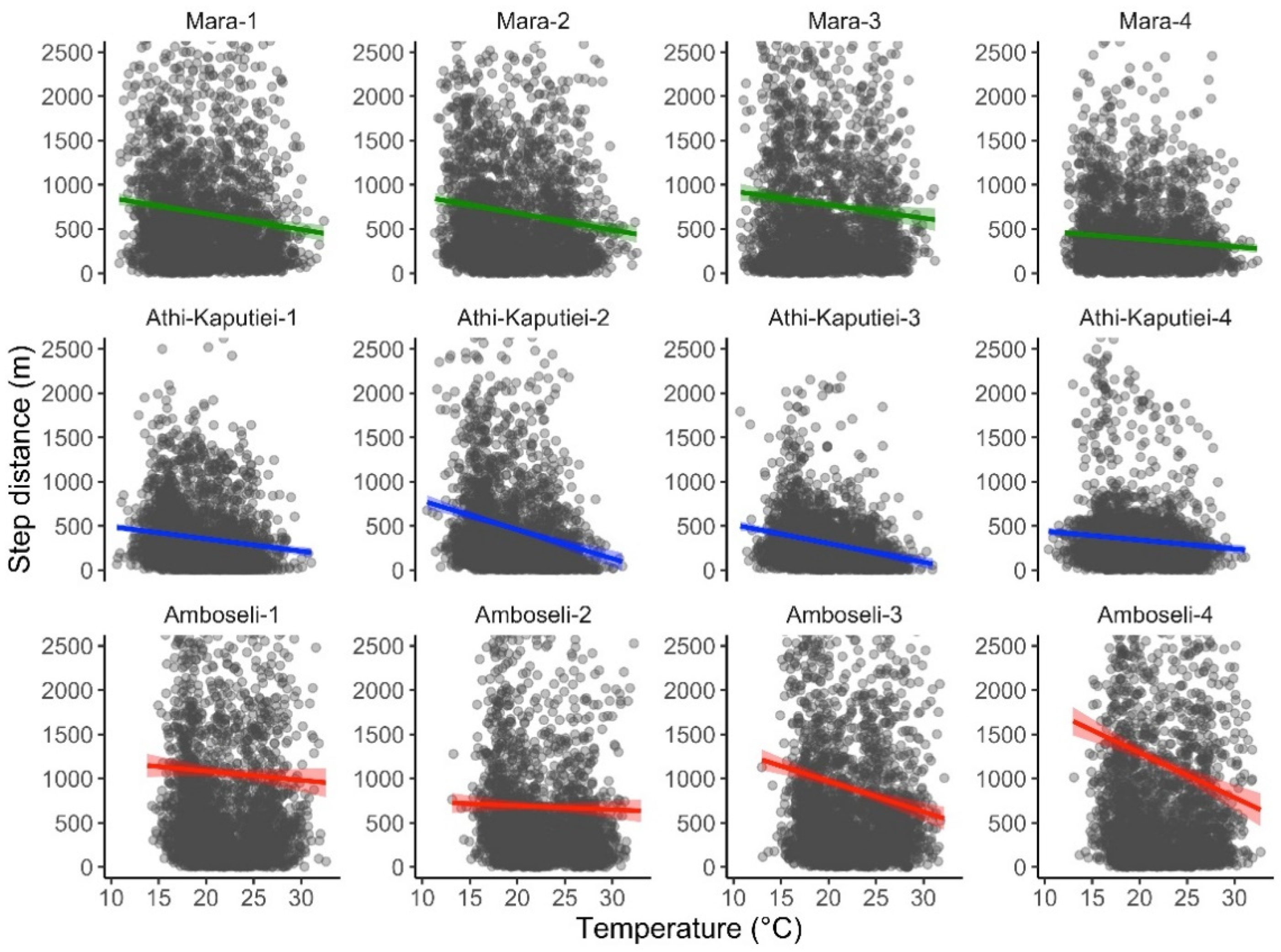
2.4. Case Study 2—Temperature Extraction
3. Results
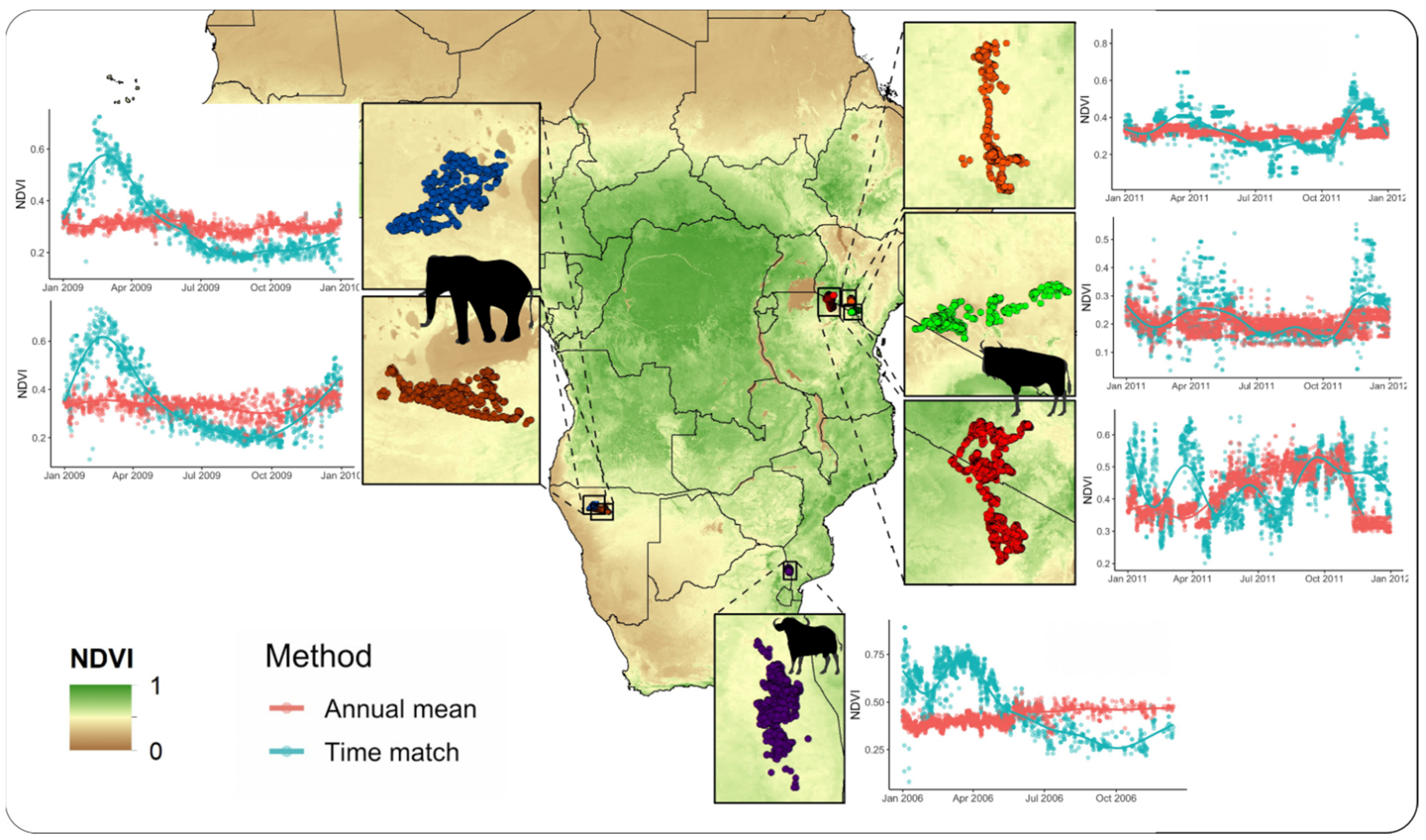
3.1. Case Study 1—NDVI Extraction
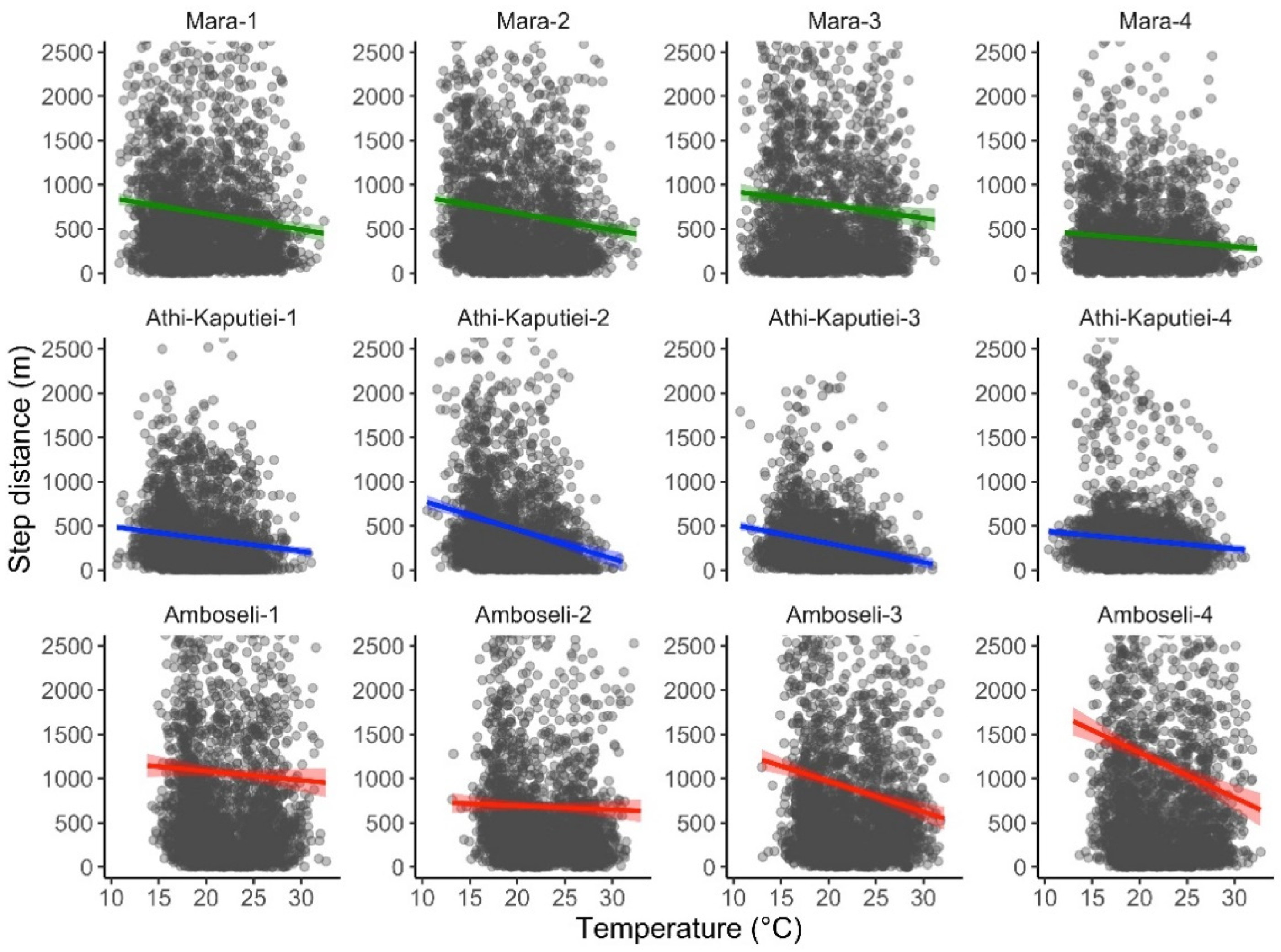
3.2. Case Study 2—Temperature Extraction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joo, R.; Picardi, S.; Boone, M.E.; Clay, T.A.; Patrick, S.C.; Romero-Romero, V.S.; Basille, M. A decade of movement ecology. arXiv 2020, arXiv:2006.00110. [Google Scholar]
- Kays, R.; Crofoot, M.C.; Jetz, W.; Wikelski, M. Terrestrial animal tracking as an eye on life and planet. Science 2015, 348, 2478. [Google Scholar] [CrossRef] [Green Version]
- Neumann, W.; Martinuzzi, S.; Estes, A.B.; Pidgeon, A.M.; Dettki, H.; Ericsson, G.; Radeloff, V.C. Opportunities for the application of advanced remotely-sensed data in ecological studies of terrestrial animal movement. Mov. Ecol. 2015, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Farr, T.G.T.G.; Rosen, P.A.P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The shuttle radar topography mission. Rev. Geophys. 2007, 45, RG2004. [Google Scholar] [CrossRef] [Green Version]
- Pettorelli, N.; Ryan, S.; Mueller, T.; Bunnefeld, N.; Jedrzejewska, B.; Lima, M.; Kausrud, K. The Normalized Difference Vegetation Index (NDVI): Unforeseen successes in animal ecology. Clim. Res. 2011, 46, 15–27. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 6 May 2021).
- Joo, R.; Boone, M.E.; Clay, T.A.; Patrick, S.C.; Clusella-Trullas, S.; Basille, M. Navigating through the r packages for movement. J. Anim. Ecol. 2020, 89, 248–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, J.; Lortie, C.J.; Muenchen, R.A.; Yang, J.; Ma, K. Evaluating the popularity of R in ecology. Ecosphere 2019, 10, e02567. [Google Scholar] [CrossRef] [Green Version]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Aybar, C.; Wu, Q.; Bautista, L.; Yali, R.; Barja, A. rgee: An R package for interacting with Google Earth Engine. J. Open Source Softw. 2020, 5, 2272. [Google Scholar] [CrossRef]
- Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Cross, P.; Bowers, J.; Hay, C.; Wolhuter, J.; Buss, P.; Hofmeyr, M.; du Toit, J.; Getz, W. Data from: Nonparameteric Kernel Methods for Constructing Home Ranges and Utilization Distributions. Movebank Data Repository. 2016. Available online: https://www.doi.org/10.5441/001/1.j900f88t (accessed on 6 May 2021).
- Tsalyuk, M.; Kilian, W.; Reineking, B.; Getz, W.M. Temporal variation in resource selection of African elephants follows long-term variability in resource availability. Ecol. Monogr. 2019, 89, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Kilian, W.; Getz, W.; Zidon, R.; Tsalyuk, M. Data from: Temporal variation in resource selection of African elephants follows long term variability in resource availability. Movebank Data Repository. 2018. Available online: https://www.doi.org/10.5441/001/1.3nj3qj45 (accessed on 6 May 2021).
- Stabach, J.A. Movement, Resource Selection, And The Physiological Stress Response of White-Bearded Wildebeest. Ph.D. Thesis, Colorado State University, Fort Collins, CO, USA, 2015. [Google Scholar]
- Stabach, J.; Hughey, L.; Reid, R.; Worden, J.; Leimgruber, P.; Boone, R. Data from: Comparison of Movement Strategies of Three Populations of White-Bearded Wildebeest. Movebank Data Repository. 2020. Available online: https://www.doi.org/10.5441/001/1.h0t27719 (accessed on 6 May 2021).
- Calenge, C. The package “adehabitat” for the R software: A tool for the analysis of space and habitat use by animals. Ecol. Model. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Pettorelli, N.; Laurance, W.F.; O’Brien, T.G.; Wegmann, M.; Nagendra, H.; Turner, W. Satellite remote sensing for applied ecologists: Opportunities and challenges. J. Appl. Ecol. 2014, 51, 839–848. [Google Scholar] [CrossRef]
- Boone, R.B.; Thirgood, S.J.; Hopcraft, J.G.C. Serengeti wildebeest migratory patterns modeled from rainfall and new vegetation growth. Ecology 2006, 87, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.J.; Grachev, I.A.; Bekenov, A.B.; Milner-Gulland, E.J. Saiga antelope calving site selection is increasingly driven by human disturbance. Biol. Conserv. 2010, 143, 1770–1779. [Google Scholar] [CrossRef]
- Merkle, J.A.; Monteith, K.L.; Aikens, E.O.; Hayes, M.M.; Hersey, K.R.; Middleton, A.D.; Oates, B.A.; Sawyer, H.; Scurlock, B.M.; Kauffman, M.J. Large herbivores surf waves of green-up during spring. Proc. R. Soc. B Biol. Sci. 2016, 283, 1–8. [Google Scholar] [CrossRef]
- Aikens, E.O.; Mysterud, A.; Merkle, J.A.; Cagnacci, F.; Rivrud, I.M.; Hebblewhite, M.; Hurley, M.A.; Peters, W.; Bergen, S.; De Groeve, J.; et al. Wave-like Patterns of Plant Phenology Determine Ungulate Movement Tactics. Curr. Biol. 2020, 30, 3444–3449. [Google Scholar] [CrossRef]
- Aikens, E.O.; Kauffman, M.J.; Merkle, J.A.; Dwinnell, S.P.H.; Fralick, G.L.; Monteith, K.L. The greenscape shapes surfing of resource waves in a large migratory herbivore. Ecol. Lett. 2017, 20, 741–750. [Google Scholar] [CrossRef]
- Geremia, C.; Merkle, J.A.; Eacker, D.R.; Wallen, R.L.; White, P.J.; Hebblewhite, M.; Kauffman, M.J. Migrating bison engineer the green wave. Proc. Natl. Acad. Sci. USA 2019, 116, 25707–25713. [Google Scholar] [CrossRef] [Green Version]
- Gibert, J.P.; Chelini, M.C.; Rosenthal, M.F.; Delong, J.P. Crossing regimes of temperature dependence in animal movement. Glob. Chang. Biol. 2016, 22, 1722–1736. [Google Scholar] [CrossRef] [PubMed]
- Bryce, C.M.; Wilmers, C.C.; Williams, T.M. Energetics and evasion dynamics of large predators and prey: Pumas vs. hounds. PeerJ 2017, 2017, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scantlebury, D.M.; Mills, M.G.L.; Wilson, R.P.; Wilson, J.W.; Mills, M.E.J.; Durant, S.M.; Bennett, N.C.; Bradford, P.; Marks, N.J.; Speakman, J.R. Flexible energetics of cheetah hunting strategies provide resistance against kleptoparasitism. Science 2014, 346, 79–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, D.; Snelling, E.P.; Hetem, R.S.; Maloney, S.K.; Strauss, W.M.; Fuller, A. Revisiting concepts of thermal physiology: Predicting responses of mammals to climate change. J. Anim. Ecol. 2018, 87, 956–973. [Google Scholar] [CrossRef]
- Alston, J.M.; Joyce, M.J.; Merkle, J.A.; Moen, R.A. Temperature shapes movement and habitat selection by a heat-sensitive ungulate. Landsc. Ecol. 2020, 35, 1961–1973. [Google Scholar] [CrossRef]
- Thaker, M.; Gupte, P.R.; Prins, H.H.T.; Slotow, R.; Vanak, A.T. Fine-scale tracking of ambient temperature and movement reveals shuttling behavior of elephants to water. Front. Ecol. Evol. 2019, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mole, M.A.; DÁraujo, S.R.; van Aarde, R.J.; Mitchell, D.; Fuller, A. Coping with heat: Behavioural and physiological responses of savanna elephants in their natural habitat. Conserv. Physiol. 2016, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, S.L.; Hundertmark, K.J.; Person, D.K.; Lindberg, M.S.; Boyce, M.S. Behavioral plasticity in a variable environment: Snow depth and habitat interactions drive deer movement in winter. J. Mammal. 2017, 98, 246–259. [Google Scholar] [CrossRef] [Green Version]
- Robinson, B.G.; Merrill, E.H. The influence of snow on the functional response of grazing ungulates. Oikos 2012, 121, 28–34. [Google Scholar] [CrossRef]
- Davidson, S.C.; Bohrer, G.; Gurarie, E.; LaPoint, S.; Mahoney, P.J.; Boelman, N.T.; Eitel, J.U.H.; Prugh, L.R.; Vierling, L.A.; Jennewein, J.; et al. Ecological insights from three decades of animal movement tracking across a changing Arctic. Science 2020, 370, 712–715. [Google Scholar] [CrossRef]
- Muñoz Sabater, J. ERA5-Land hourly data from 1981 to present. Copernicus Climate Change Service (C3S) Climate Data Store (CDS). 2019. Available online: https://doi.org/10.24381/cds.e2161bac (accessed on 15 September 2021).
- Plummer, M. JAGS: A program for analysis of Bayesian graphical models using Gibbs sampling. (Version 4.2). In Proceedings of the 3rd International Workshop on Distributed Statistical Computing (DSC 2003), Vienna, Austria, 20–22 March 2003. [Google Scholar]
- Gelman, A.; Rubin, D. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–511. [Google Scholar] [CrossRef]
- Tucker, M.A.; Böhning-gaese, K.; Fagan, W.F.; Fryxell, J.M.; Van Moorter, B.; Alberts, S.C.; Ali, A.H.; Allen, A.M.; Attias, N.; Avgar, T.; et al. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 2018, 469, 466–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pekel, J.F.; Cottam, A.; Gorelick, N.; Belward, A.S. High-resolution mapping of global surface water and its long-term changes. Nature 2016, 540, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Wardlow, B.D.; Hu, S.; Zhang, X.; Zhou, G.; Peng, G.; Xiang, D.; Wang, R.; Meng, R.; Wu, W. A novel strategy to reconstruct NDVI time-series with high temporal resolution from MODIS multi-temporal composite products. Remote Sens. 2021, 13, 1397. [Google Scholar] [CrossRef]
- Oeser, J.; Heurich, M.; Senf, C.; Pflugmacher, D.; Belotti, E.; Kuemmerle, T. Habitat metrics based on multi-temporal Landsat imagery for mapping large mammal habitat. Remote Sens. Ecol. Conserv. 2020, 6, 52–69. [Google Scholar] [CrossRef] [Green Version]
- Manly, B.F.; McDonald, L.; Thomas, D.; McDonald, T.L.; Erickson, W.P. Resource Selection by Animals; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Thurfjell, H.; Ciuti, S.; Boyce, M.S. Applications of step-selection functions in ecology and conservation. Mov. Ecol. 2014, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avgar, T.; Potts, J.R.; Lewis, M.A.; Boyce, M.S. Integrated step selection analysis: Bridging the gap between resource selection and animal movement. Methods Ecol. Evol. 2016, 7, 619–630. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Number of Individuals | Year | Remote Sensing Product | Number of Images in the Filtered Collection | Number of GPS Locations | Processing Time * |
|---|---|---|---|---|---|---|
| African buffalo | 1 | 2006 | MOD13Q1 | 23 | 1545 | 16.84 s |
| African elephant | 2 | 2009 | MOD13Q1 | 23 | 2451 | 25.39 s |
| Wildebeest case study 1 | 3 | 2011 | MOD13Q1 | 23 | 8348 | 2.28 min |
| Wildebeest case study 2 | 12 | 2011 | ERA5_LAND/HOURLY | 8760 | 33,074 | 33.83 min |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crego, R.D.; Masolele, M.M.; Connette, G.; Stabach, J.A. Enhancing Animal Movement Analyses: Spatiotemporal Matching of Animal Positions with Remotely Sensed Data Using Google Earth Engine and R. Remote Sens. 2021, 13, 4154. https://doi.org/10.3390/rs13204154
Crego RD, Masolele MM, Connette G, Stabach JA. Enhancing Animal Movement Analyses: Spatiotemporal Matching of Animal Positions with Remotely Sensed Data Using Google Earth Engine and R. Remote Sensing. 2021; 13(20):4154. https://doi.org/10.3390/rs13204154
Chicago/Turabian StyleCrego, Ramiro D., Majaliwa M. Masolele, Grant Connette, and Jared A. Stabach. 2021. "Enhancing Animal Movement Analyses: Spatiotemporal Matching of Animal Positions with Remotely Sensed Data Using Google Earth Engine and R" Remote Sensing 13, no. 20: 4154. https://doi.org/10.3390/rs13204154
APA StyleCrego, R. D., Masolele, M. M., Connette, G., & Stabach, J. A. (2021). Enhancing Animal Movement Analyses: Spatiotemporal Matching of Animal Positions with Remotely Sensed Data Using Google Earth Engine and R. Remote Sensing, 13(20), 4154. https://doi.org/10.3390/rs13204154