
Monitoring the Severity of Pantana phyllostachysae Chao on Bamboo Using Leaf Hyperspectral Data



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. The Criteria of Pest Severity
2.3. Measurements and Analysis of Leaf Physicochemical Parameters
2.3.1. Leaf Reflectance Measurements
2.3.2. Measurements and Analysis of Leaf Biochemical Factors
2.4. Indicators of Pest Severity
2.4.1. Hyperspectral Indices
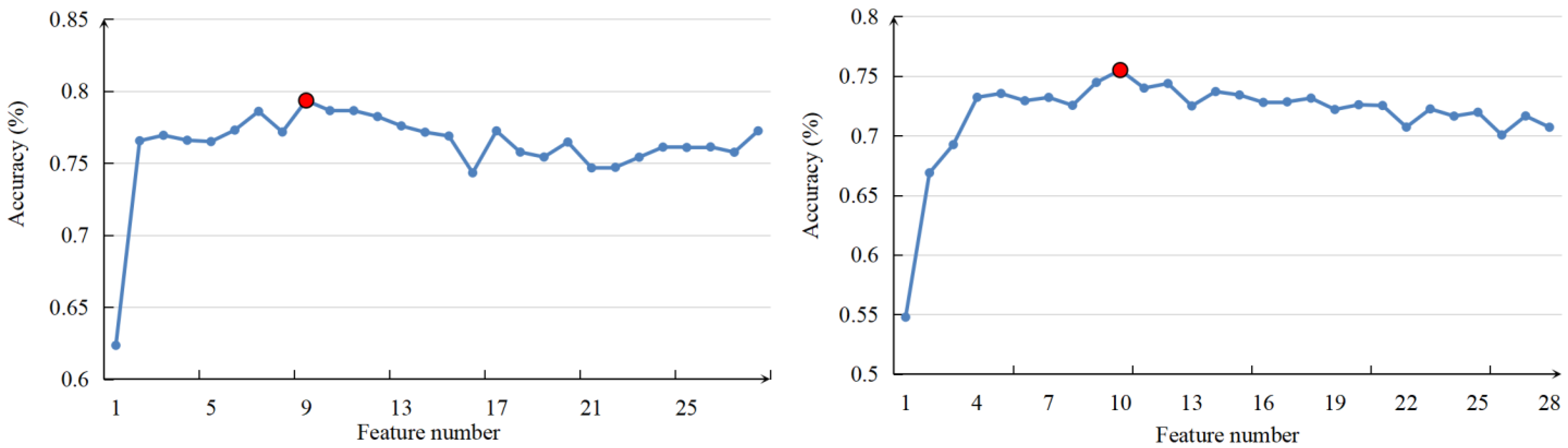
2.4.2. Feature Selection
2.5. Development of Severity Identification Model
2.6. Model Accuracy Evaluation
3. Results
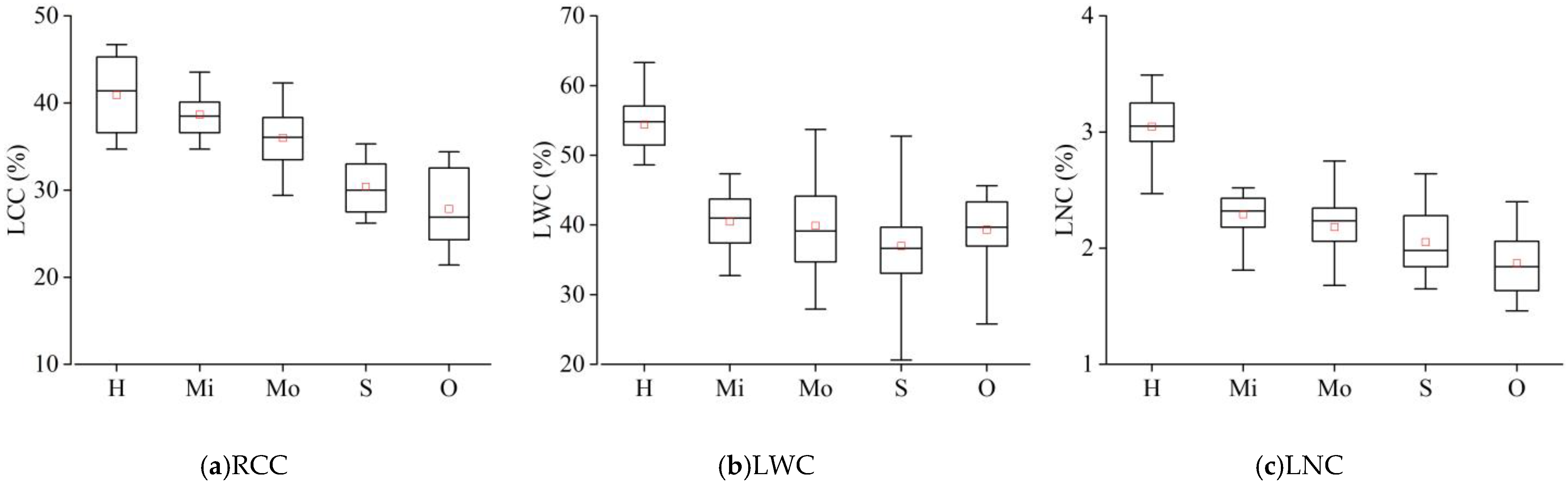
3.1. Physicochemical Differences among Healthy, Damaged, and Off-Year Leaves
3.1.1. Differences of Biochemical Factors among Healthy, Damaged, and Off-Year Leaves
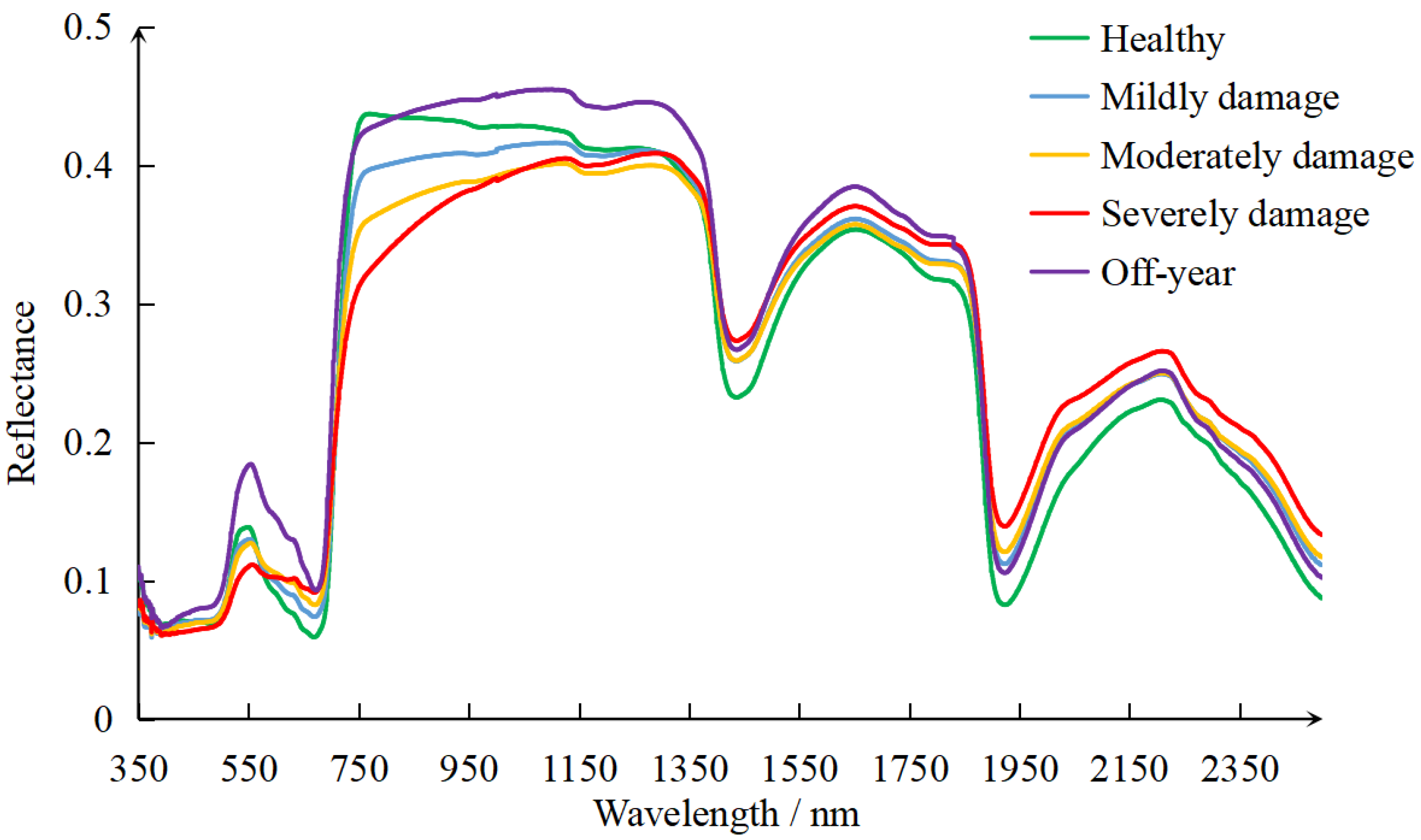
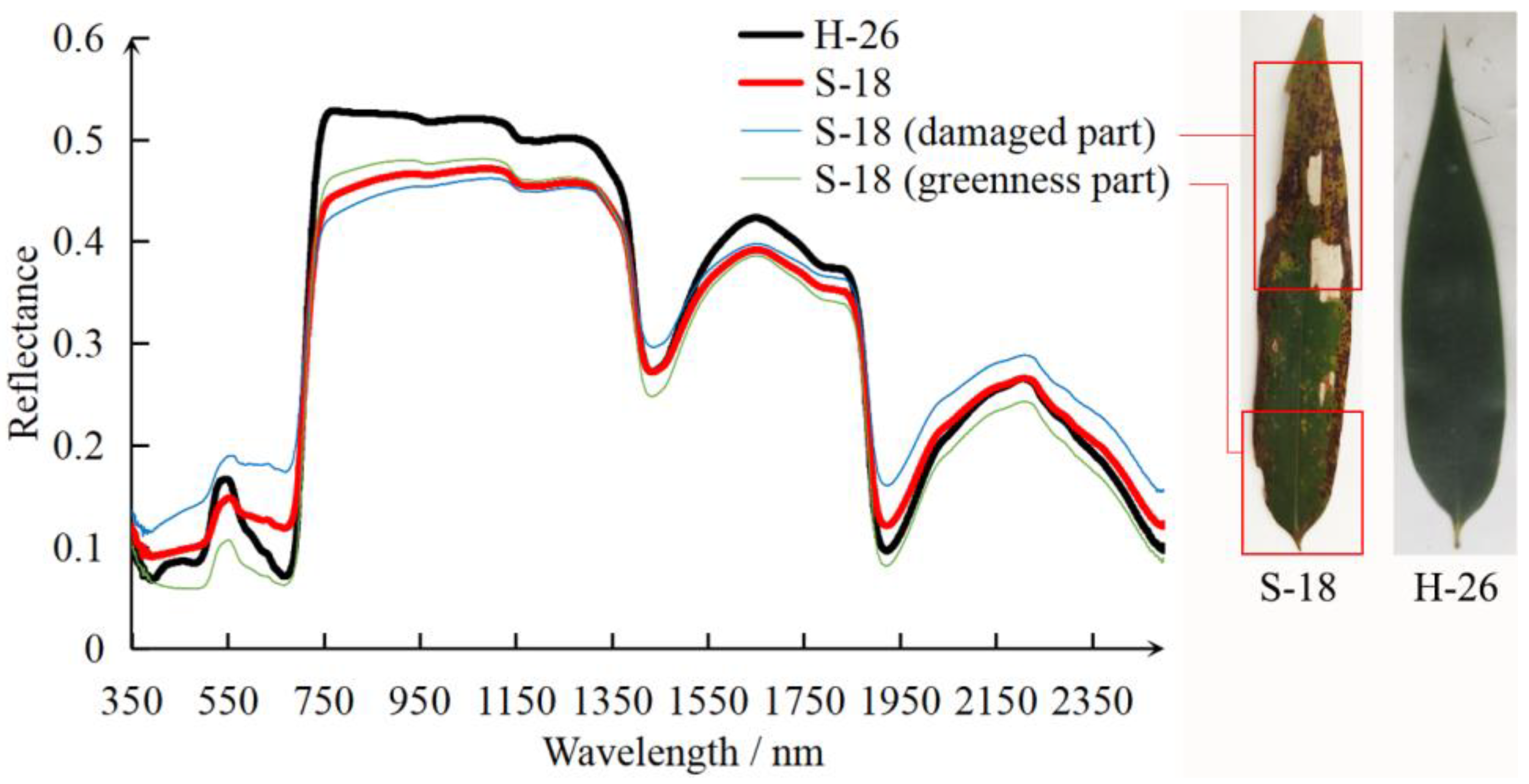
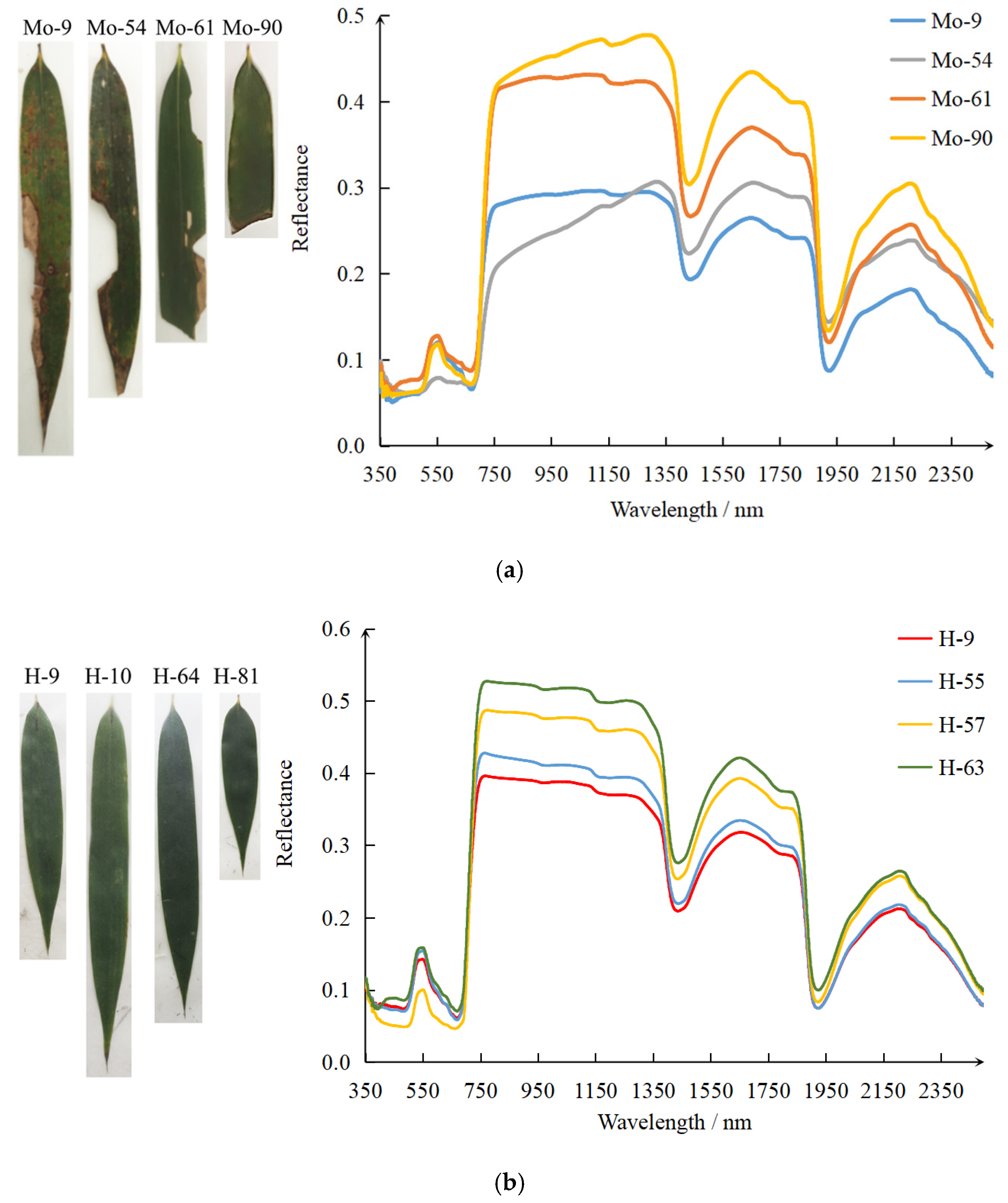
3.1.2. Spectra of Healthy, Damaged, and Off-Year Leaves
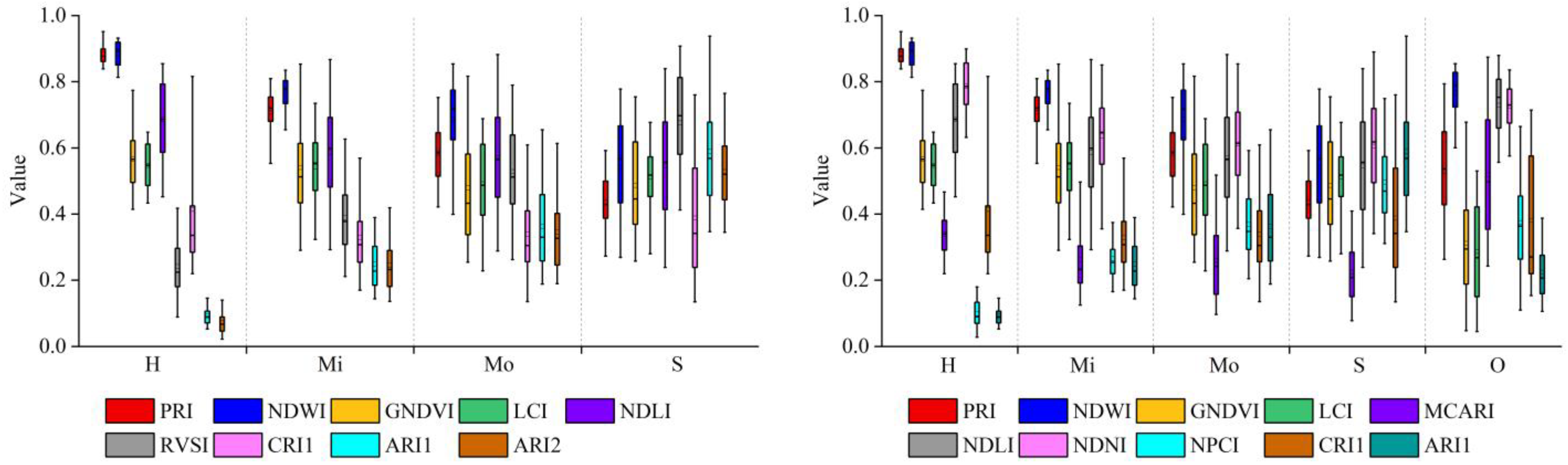
3.2. Features Selected as Model Inputs
3.3. Model Performance
4. Discussion
4.1. The Effects of PPC on Host Bamboo Leaf Spectrum
4.2. The Uncertainty in Pest Severity Identification
4.3. Weaknesses and Prospects of the Proposed Approach
5. Conclusions
- (1)
- The RCC, the LWC, and the LNC of damaged leaves were lower than those of healthy leaves, and the decreasing magnitude increased with the aggravation of damaged severity. The differences in LWC among different damaged groups were not significant. The nutrient content of off-year leaves was significantly lower than that of healthy leaves, but its difference in LWC with damaged leaves was not obvious.
- (2)
- The pest damage caused noticeable distortion of leaf spectrum. The reflectance of damaged leaves decreased in the green band and increased in the red band. The reflectance difference between healthy and damaged leaves in the near-infrared band was the greatest. Damaged leaves had much lower reflectance in the near-infrared and much higher reflectance in shortwave-infrared relative to healthy leaves. The reflectance of off-year leaves was noticeably higher than healthy and damaged leaves in visible and near-infrared regions.
- (3)
- Selected spectral indicators detectably differed among damage groups of leaves. However, their values also had great variations within the same severity group due to the physiological difference of individual leaves and the damaged timing on host bamboo, which imposes difficulties in identifying damage severities.
- (4)
- The proposed model can effectively distinguish the damaged leaves, and its identification accuracy for healthy and severely damaged leaves is the best. The identification ability of the model for moderately damaged leaves is poor, i.e., below 60%. Off-year leaves can noticeably affect the identification results of damaged leaves since they were easily classified as damaged ones.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mao, F.J.; Zhou, G.M.; Li, P.H.; Du, H.Q.; Xu, X.J.; Shi, Y.J.; Mo, L.F.; Zhou, Y.F.; Tu, G.Q. Optimizing selective cutting strategies for maximum carbon stocks and yield of Moso bamboo forest using BIOME-BGC model. J. Environ. Manag. 2017, 191, 126–135. [Google Scholar] [CrossRef] [PubMed]
- National Forestry and Grassland Administration. P.R.C. China Forest Resources Report (2014–2018); China Forestry Publishing House: Beijing, China, 2019; pp. 3–4. [Google Scholar]
- Yin, J.Y.; Ge, Z.P.; Deng, X.; Shi, Y.J.; Xu, L.; Zhou, G.M.; Ye, Y.Z.; Zhang, F.G.; Zhou, Y.F.; Li, C.; et al. Abandonment lead to structural degradation and changes in carbon allocation patterns in Moso bamboo forests. For. Ecol. Manag. 2019, 449, 117449. [Google Scholar] [CrossRef]
- Zhang, F.P.; Hou, Y.M.; You, M.S. Effects of different management measures on the composition and structure of arthropod community in Phyllostachys heterocycla cv. pubescens forest. Acta Entomol. Sin. 2005, 48, 928–934. [Google Scholar] [CrossRef]
- Chen, D.L.; Wang, B.Y. Life history of Pantana phyllostachysae and relationships between the insect and meteorological conditions. J. Zhejiang For. Coll. 1993, 10, 342–345. [Google Scholar]
- Lan, S.W.; Ye, X.Y.; Li, T.S.; Lu, S.Q.; Yang, Z.W.; Lu, M.Z. Type of distribution and method of density estimation for Pantana phyllostachysae. For. Res. 1993, 6, 579–582. [Google Scholar] [CrossRef]
- Rullan-Silva, C.D.; Olthoff, A.E.; Delgado de la Mata, J.A.; Pajares-Alonso, J.A. Remote Monitoring of Forest Insect Defoliation—A Review. For. Syst. 2013, 22, 377–391. [Google Scholar] [CrossRef]
- Su, J.; Zhang, F.P.; Huang, W.L.; Chen, D.L.; Chen, S.L. Influence of different types of Phyllostachys pubescens (Poales: Poaceae) leaves on population parameters of Pantana phyllostachysae (Lepidoptera: Lymantriidae) and parasitic effects of Beauveria bassiana (Moniliales: Moniliaceae). J. Insect Sci. 2015, 15, 39. [Google Scholar] [CrossRef]
- Cai, G.G. Screening of the superior strains of Beauveria bassiana of Pantana phyllostachysae and practical application. Sci. Silvae Sin. 2003, 39, 102–108. [Google Scholar]
- Zhou, Z.Y.; Hu, X.Y.; Lin, L.; Huang, X.Y.; Xu, Z.H.; Zhou, H.K.; Xu, R.N.; Wu, K.X. Preliminary study on the influence of topographic factors on the damage of Pantana phyllostachysae Chao. J. Agric. Catastrophol. 2020, 10, 61–64. [Google Scholar] [CrossRef]
- Wei, C.J. Plague division and application of Pantana phyllostachysae Chao in Fujian province. J. For. Environ. 2003, 23, 79–83. [Google Scholar]
- Lausch, A.; Heurich, M.; Gordalla, D.; Dobner, H.J.; Gwillym-Margianto, S.; Salbach, C. Forecasting potential bark beetle outbreaks based on spruce forest vitality using hyperspectral remote-sensing techniques at different scales. For. Ecol. Manag. 2013, 308, 76–89. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Latifi, H.; Ghosh, A.; Joshi, P.K.; Koch, B. Assessing the potential of hyperspectral imagery to map bark beetle-induced tree mortality. Remote Sens. Environ. 2014, 140, 533–548. [Google Scholar] [CrossRef]
- Liang, S.X.; Qian, S.S.; Hei, Z.H. Life histroy of the moth Pantana phyllostachysae and its control. Chin. J. Appl. Entomol. 2004, 41, 464–467. [Google Scholar]
- Zhang, S.L.; Qin, J.; Tang, X.D.; Wang, Y.J.; Huang, J.L.; Song, Q.L.; Min, J.Y. Spectral Characteristics and Evaluation Model of Pinus Massoniana Suffering from Bursaphelenchus xylophilus Disease. Spectrosc. Spectr. Anal. 2019, 39, 865–872. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, X.L.; Yang, G.J.; Zhu, C.H.; Huo, L.N.; Feng, H.K. Assessment of defoliation during the Dendrolimus tabulaeformis Tsai et Liu disaster outbreak using UAV-based hyperspectral images. Remote Sens. Environ. 2018, 217, 323–339. [Google Scholar] [CrossRef]
- Nguyen, H.D.D.; Nansen, C. Hyperspectral remote sensing to detect leafminer-induced stress in bok choy and spinach according to fertilizer regime and timing. Pest Manag. Sci. 2020, 76, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Li, L.W.; Li, N.; Zang, Z.; Lu, D.S.; Wang, G.X.; Wang, N. Examining phenological variation of on-year and off-year bamboo forests based on the vegetation and environment monitoring on a New Micro-Satellite (VENµS) time-series data. Int. J. Remote Sens. 2020, 42, 2203–2219. [Google Scholar] [CrossRef]
- Feng, H.Y. Study on the Establishment of Health Evaluation System for Phyllostachys edulis Forest and its Application. Ph.D. Thesis, Chinese Academy of Forestry, Beijing, China, 2014. [Google Scholar]
- Ni, H.; Su, W.; Fan, S.; Chu, H. Effects of intensive management practices on rhizosphere soil properties, root growth, and nutrient uptake in Moso bamboo plantations in subtropical China. For. Ecol. Manag. 2021, 493, 119083. [Google Scholar] [CrossRef]
- Yang, C.; Wang, A.; Zhu, Z.; Lin, S.; Bi, Y.; Du, X. Impact of extensive management system on soil properties and carbon sequestration under an age chronosequence of Moso bamboo plantations in subtropical China. For. Ecol. Manag. 2021, 497, 119535. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Zhou, G.; Mao, F.; Du, H.; Xu, X.; Li, P.; Li, X. Diurnal and seasonal variations in carbon fluxes in bamboo forests during the growing season in Zhejiang province, China. J. For. Res. 2019, 30, 657–668. [Google Scholar] [CrossRef]
- Xu, C.; Wang, H.X.; Wen, G.S.; Hu, C.; Liang, X.E.; Huang, H.N.; Zhu, L.N.; Bai, S.B.; Zhang, R.M. Analysis on the reason of Phyllostachys edulis explosive growth by physiological and ecological. J. Cent. South Univ. For. Technol. 2018, 38, 64–69. [Google Scholar] [CrossRef]
- Li, X.T. Bionomics and comprehensive control measure of Pantana phyllostachysae. Contemp. Hortic. 2013, 2, 153–154. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Mohammed, G.H.; Noland, T.L. Chlorophyll fluorescence effects on vegetation apparent reflectance: I. Leaf-level measurements and model simulation. Remote Sens. Environ. 2000, 74, 582–595. [Google Scholar] [CrossRef]
- Sims, N.C.; De Barro, P.; Newnham, G.J.; Kalyebi, A.; Macfadyen, S.; Malthus, T.J. Spectral separability and mapping potential of cassava leaf damage symptoms caused by whiteflies (Bemisia tabaci). Pest Manag. Sci. 2018, 74, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Golhani, K.; Balasundram, S.K.; Vadamalai, G.; Pradhan, B. Estimating chlorophyll content at leaf scale in viroid-inoculated oil palm seedlings (Elaeis guineensis Jacq.) using reflectance spectra (400 nm–1050 nm). Int. J. Remote Sens. 2019, 40, 7647–7662. [Google Scholar] [CrossRef]
- Bu, Q.; Hou, Y.; Fang, H.; Zhang, Z.; Bai, X. Eco-Physiological Characteristics of the Exotic Plant Rhus typhina in Comparison with Its Indigenous Congeners Rhus chinensis. Sci. Silvae Sin. 2017, 53, 1–7. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, X.; Hou, Y.; Dong, Z.; Bu, Q. Comparison on the competitiveness of the invaded pokeweed with its accompanying species in the coastal protection forest of Shandong Peninsula. Sci. Silvae Sin. 2016, 52, 24–29. [Google Scholar] [CrossRef]
- Liu, B.; Dong, Y.; He, T.; Chen, Y.; Yang, H.; Wang, X.; Chen, H. Component change law of char during bamboo pyrolysis process. Transations Chin. Soc. Agric. Eng. 2016, 32, 232–239. [Google Scholar]
- Oumar, Z.; Mutanga, O.; Ismail, R. Predicting Thaumastocoris peregrinus damage using narrow band normalized indices and hyperspectral indices using field spectra resampled to the Hyperion sensor. Int. J. Appl. Earth Obs. 2013, 21, 113–121. [Google Scholar] [CrossRef]
- Abdullah, H.; Darvishzadeh, R.; Skidmore, A.K.; Groen, T.A.; Heurich, M. European spruce bark beetle (Ips typographus, L.) green attack affects foliar reflectance and biochemical properties. Int. J. Appl. Earth Obs. 2018, 64, 199–209. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Estimation of vegetation water content and photosynthetic tissue area from spectral reflectance: A comparison of indices based on liquid water and chlorophyll absorption features. Remote Sens. Environ. 2003, 84, 526–537. [Google Scholar] [CrossRef]
- Vogelmann, J.E.; Rock, B.N.; Moss, D.M. Red edge spectral measurements from sugar maple leaves. Int. J. Remote Sens. 1993, 14, 1563–1575. [Google Scholar] [CrossRef]
- Gamon, J.A.; Penuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Rakitin, V.Y. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef]
- Gao, B.C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Zhang, H.W.; Zhang, F.; Zhang, X.L.; Li, Z.; Abduwasit, G.; Song, J. Inversion of vegetation leaf water content based on spectral index. Spectrosc. Spectr. Anal. 2018, 38, 1540–1546. [Google Scholar] [CrossRef]
- Song, B.W.; Liu, L.Y.; Zhang, B. A novel restoration approach for vegetation reflectance spectra at noisy bands using the principal component analysis method. Int. J. Remote Sens. 2020, 41, 2303–2325. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Spectral reflectance changes associated with autumn senescence of Aesculus hippocastanum L. and Acer platanoides L. leaves. spectral features and relation to chlorophyll estimation. J. Plant Physiol. 1994, 143, 286–292. [Google Scholar] [CrossRef]
- Peñuelas, J.; Baret, F.; Filella, I. Semiempirical indexes to assess carotenoids chlorophyll—A ratio from leaf spectral reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Apan, A.; Held, A.; Phinn, S.; Markley, J. Detecting sugarcane ‘orange rust’ disease using EO-1 Hyperion hyperspectral imagery. Int. J. Remote Sens. 2004, 25, 489–498. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; de Colstoun, E.B.; McMurtrey, J.E. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Serrano, L.; Peñuelas, J.; Ustin, S.L. Remote sensing of nitrogen and lignin in Mediterranean vegetation from AVIRIS data: Decomposing biochemical from structural signals. Remote Sens. Environ. 2002, 81, 355–364. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Keydan, G.P.; Merzlyak, M.N. Three-band model for noninvasive estimation of chlorophyll, carotenoids, and anthocyanin contents in higher plant leaves. Geophys. Res. Lett. 2006, 33, L11402. [Google Scholar] [CrossRef]
- Cao, Z.S.; Yao, X.; Liu, H.Y.; Liu, B.; Cheng, T.; Tian, Y.C.; Cao, W.X.; Zhu, Y. Comparison of the abilities of vegetation indices and photosynthetic parameters to detect heat stress in wheat. Agric. For. Meteorol. 2019, 265, 121–136. [Google Scholar] [CrossRef]
- Perry, E.M.; Davenport, J.R. Spectral and spatial differences in response of vegetation indices to nitrogen treatments on apple. Comput. Electron. Agric. 2007, 59, 56–65. [Google Scholar] [CrossRef]
- Peñuelas, J.; Gamon, J.A.; Fredeen, A.L.; Merino, J.; Field, C.B. Reflectance indices associated with physiological changes in nitrogen- and water-limited sunflower leaves. Remote Sens. Environ. 1994, 48, 135–146. [Google Scholar] [CrossRef]
- Augusto-Silva, P.; Ogashawara, I.; Barbosa, C.; de Carvalho, L.; Jorge, D.; Fornari, C.; Stech, J. Analysis of MERIS reflectance algorithms for estimating chlorophyll—A concentration in a Brazilian reservoir. Remote Sens. 2014, 6, 11689–11707. [Google Scholar] [CrossRef]
- Xu, Z.H.; Liu, J.; Yu, K.Y.; Gong, C.H.; Xie, W.J.; Tang, M.Y.; Lai, R.W.; Li, Z.L. Spectral features analysis of Pinus massoniana with pest of Dendrolimus punctatus walker and level detection. Spectrosc. Spectr. Anal. 2013, 33, 428–433. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, S.Y.; Yan, L.; Kuan, F.H.; Zhao, S.Y.; Zhao, H.Y. Improvement of hyperspectral estimation of nitrogen content in winter wheat by leaf surface polarized reflection measurement. Spectrosc. Spectr. Anal. 2020, 40, 1956–1964. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing carotenoid content in plant leaves with reflectance spectroscopy. Photochem. Photobiol. 2002, 75, 272–281. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N.; Chivkunova, O.B. Optical properties and nondestructive estimation of anthocyanin content in plant leaves. Photochem. Photobiol. 2001, 74, 38–45. [Google Scholar] [CrossRef]
- Yan, K.; Zhang, D. Feature selection and analysis on correlated gas sensor data with recursive feature elimination. Sens. Actuators B. 2015, 212, 353–363. [Google Scholar] [CrossRef]
- Ke, G.; Meng, Q.; Finley, T.; Wang, T.; Chen, W.; Ma, W.; Ye, Q.; Liu, T. LightGBM: A Highly Efficient Gradient Boosting Decision Tree. In Proceedings of the 31st Annual Conference on Neural Information Processing Systems (NIPS), Long Beach, CA, USA, 4–9 December 2017; Curran Associates Inc.: New York, NY, USA, 2017; pp. 3146–3154. [Google Scholar]
- Wang, N.; Zhang, G.; Pang, W.; Ren, L.; Wang, Y. Novel monitoring method for material removal rate considering quantitative wear of abrasive belts based on LightGBM learning algorithm. Int. J. Adv. Manuf. Technol. 2021, 114, 3241–3253. [Google Scholar] [CrossRef]
- Su, H.; Lu, X.; Chen, Z.; Zhang, H.; Lu, W.; Wu, W. Estimating Coastal Chlorophyll-A Concentration from Time-Series OLCI Data Based on Machine Learning. Remote Sens. 2021, 13, 576. [Google Scholar] [CrossRef]
- Xu, X.J.; Du, H.Q.; Zhou, G.M.; Mao, F.J.; Li, X.J.; Zhu, D.E.; Li, Y.G.; Cui, L. Remote estimation of canopy leaf area index and chlorophyll content in Moso bamboo (Phyllostachys edulis (Carrière) J. Houz.) forest using MODIS reflectance data. Ann. For. Sci. 2018, 75, 33. [Google Scholar] [CrossRef]
- Woodward, I. Plant science: Tall storeys. Nature 2004, 428, 807–808. [Google Scholar] [CrossRef]
- Nolan, R.H.; Mitchell, P.J.; Bradstock, R.A.; Lane, P.N. Structural adjustments in resprouting trees drive differences in post-fire transpiration. Tree Physiol. 2014, 34, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Barta, V.; Lukes, P.; Homolova, L. Early detection of bark beetle infestation in Norway spruce forests of Central Europe using Sentinel-2. Int. J. Appl. Earth Obs. Geoinf. 2021, 100, 102335. [Google Scholar] [CrossRef]
- Huo, L.N.; Persson, H.J.; Lindberg, E. Early detection of forest stress from European spruce bark beetle attack, and a new vegetation index: Normalized distance red & SWIR (NDRS). Remote Sens. Environ. 2021, 255, 112240. [Google Scholar] [CrossRef]
- Martinez-Vilalta, J.; Sauret, M.; Duro, A.; Pinol, J. Make your own transpiring tree. J. Biol. Educ. 2003, 38, 32–35. [Google Scholar] [CrossRef]
- Huang, L.; Ning, Z.Y.; Zhang, X.L. Impacts of caterpillar disturbance on forest net primary production estimation in China. Ecol. Indic. 2010, 10, 1144–1151. [Google Scholar] [CrossRef]
- Datt, B. Remote Sensing of Chlorophyll a, Chlorophyll b, Chlorophyll a + b, and Total Carotenoid Content in Eucalyptus Leaves. Remote Sens. Environ. 1998, 66, 111–121. [Google Scholar] [CrossRef]
- Zhou, X.H.; Li, Y.Q.; Huang, Z.; Luo, J.D.; Xiao, Z.Y.; Yan, S.B. Effects of defoliations on chlorophyll contents and growth of Sapium discolor seedlings. J. Cent. South Univ. For. Technol. 2017, 37, 39–43. [Google Scholar] [CrossRef]
- Barros, P.P.S.; Schutze, I.X.; Iost Filho, F.H.; Yamamoto, P.T.; Fiorio, P.R.; Dematte, J.A.M. Monitoring Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) Infestation in Soybean by Proximal Sensing. Insects 2021, 12, 47. [Google Scholar] [CrossRef] [PubMed]
- Nabity, P.D.; Hillstrom, M.L.; Lindroth, R.L.; DeLucia, E.H. Elevated CO2 interacts with herbivory to alter chlorophyll fluorescence and leaf temperature in Betula papyrifera and Populus tremuloides. Oecologia 2012, 169, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Donovan, S.D.; MacLean, D.A.; Zhang, Y.; Lavigne, M.B.; Kershaw, J.A. Evaluating annual spruce budworm defoliation using change detection of vegetation indices calculated from satellite hyperspectral imagery. Remote Sens. Environ. 2021, 253, 112204. [Google Scholar] [CrossRef]
- Li, K.; Chen, Y.Z.; Xu, Z.H.; Huang, X.Y.; Hu, X.Y.; Wang, X.Q. Hyperspectral Estimation Method of Chlorophyll Content in MOSO Bamboo under Pests Stress. Spectrosc. Spectr. Anal. 2020, 40, 2578–2583. [Google Scholar] [CrossRef]
- Liu, Y.; Zhan, Z.; Ren, L.; Ze, S.; Yu, L.; Jiang, Q.; Luo, Y. Hyperspectral evidence of early-stage pine shoot beetle attack in Yunnan pine. For. Ecol. Manag. 2021, 497, 119505. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R.; Hostert, P. Remote sensing of forest insect disturbances: Current state and future directions. Int. J. Appl. Earth Obs. Geoinf. 2017, 60, 49–60. [Google Scholar] [CrossRef]
- Meng, R.; Dennison, P.E.; Zhao, F.; Shendryk, I.; Rickert, A.; Hanavan, R.P.; Cook, B.D.; Serbin, S.P. Mapping canopy defoliation by herbivorous insects at the individual tree level using bi-temporal airborne imaging spectroscopy and LiDAR measurements. Remote Sens. Environ. 2018, 215, 170–183. [Google Scholar] [CrossRef]
- Huo, L.N.; Zhang, N.; Zhang, X.L.; Wu, Y.S. Tree defoliation classification based on point distribution features derived from single-scan terrestrial laser scanning data. Ecol. Indic. 2019, 103, 782–790. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Townsend, P.A. Estimating the effect of gypsy moth defoliation using MODIS. Remote Sens. Environ. 2008, 112, 3983–3990. [Google Scholar] [CrossRef]
- Lin, Q.; Huang, H.; Chen, L.; Wang, J.; Huang, K.; Liu, Y. Using the 3D model RAPID to invert the shoot dieback ratio of vertically heterogeneous Yunnan pine forests to detect beetle damage. Remote Sens. Environ. 2021, 260, 112475. [Google Scholar] [CrossRef]
- Spruce, J.P.; Sader, S.; Ryan, R.E.; Smoot, J.; Kuper, P.; Ross, K.; Prados, D.; Russell, J.; Gasser, G.; McKellip, R.; et al. Assessment of MODIS NDVI time series data products for detecting forest defoliation by gypsy moth outbreaks. Remote Sens. Environ. 2011, 115, 427–437. [Google Scholar] [CrossRef]
- Townsend, P.A.; Singh, A.; Foster, J.R.; Rehberg, N.J.; Kingdon, C.C.; Eshleman, K.N.; Seagle, S.W. A general Landsat model to predict canopy defoliation in broadleaf deciduous forests. Remote Sens. Environ. 2012, 119, 255–265. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Formula | References |
|---|---|---|
| Normalized Difference Vegetable Index, NDVI | (R800 − R670)/(R800 + R670) | [33] |
| Modified Red Edge Simple Ratio Index, mSR705 | (R750 − R445)/(R705 + R445) | [34] |
| Vogelmann Red Edge Index 1, VOG 1 | R740/R720 | [35] |
| Vogelmann Red Edge Index 2, VOG 2 | (R734 − R747)/(R715 + R726) | [35] |
| Photochemical reflectance index, PRI | (R531 − R570)/(R531 + R570) | [36] |
| Plant Senescence Reflectance Index, PSRI | (R680 − R500)/R750 | [37] |
| Normalized Difference Water Index, NDWI | (R857 − R1241)/(R857 + R1241) | [38] |
| Moisture Stress Index, MSI | R1599/R819 | [39] |
| Normalized Difference Infrared Index, NDII | (R819 − R1649)/(R819 + R1649) | [40] |
| Green Normalized Difference Vegetation Index, GNDVI | (R750 − R550)/(R750 + R550) | [41] |
| Leaf Chlorophyll Index, LCI | (R850 − R710)/(R850 − R680) | [42] |
| Disease Water Stress Index, DSWI | (R803 + R549)/(R1659 + R681) | [43] |
| Modified Chlorophyll Absorption Reflectance Index, MCARI | [(R700 − R670) − 0.2 × (R700 − R550)] × (R700/R670) | [44] |
| Normalized Difference Lignin Index, NDLI | [45] | |
| Normalized Difference Nitrogen Index, NDNI | [45] | |
| Green Chlorophyll Index, CIgreen | (R800/R550) − 1 | [46] |
| Red-edge Chlorophyll Index, CIrededge | (R800/R720) − 1 | [46] |
| Normalized Difference Red Edge Index, NDRE | (R790 − R720)/(R790 + R720) | [47] |
| Red-edge Vegetation Stress Index, RVSI | (R712 + R752) − R732 | [48] |
| Structure Insensitive Pigment Index, SIPI | (R800 − R445)/(R800 − R680) | [49] |
| Normalized Difference Chlorophyll Index, NDCI | (R762 − R527)/(R762 + R527) | [50] |
| Normalized Pigment Chlorophyll ratio Index, NPCI | (R680 − R430)/(R680 + R430) | [49] |
| Red Edge Normalized Difference Vegetation Index, RENDVI | (R760 − R680)/(R760 + R680) | [51] |
| Red Edge Ratio Vegetation Index, RERVI | R760/R680 | [51] |
| Simple Ratio, SR705 | R750/R705 | [52] |
| Carotenoid Reflectance Index 1, CRI1 | (1/R510) − (1/R550) | [53] |
| Carotenoid Reflectance Index 2, CRI2 | (1/R510) − (1/R700) | [53] |
| Anthocyanin Reflectance Index 1, ARI1 | (1/R550) − (1/R700) | [54] |
| Anthocyanin Reflectance Index 2, ARI2 | R800 × [(1/R550) − (1/R700)] | [54] |
| Significance (p-Value) | ||||
|---|---|---|---|---|
| RCC | LWC | LNC | ||
| H | Mi | 0.230 | 0.000 ** | 0.000 ** |
| Mo | 0.000 ** | 0.000 ** | 0.000 ** | |
| S | 0.000 ** | 0.000 ** | 0.000 ** | |
| O | 0.000 ** | 0.000 ** | 0.000 ** | |
| Mi | Mo | 0.047 * | 1.000 | 0.070 |
| S | 0.000 ** | 0.169 | 0.000 ** | |
| O | 0.000 ** | 0.950 | 0.000 ** | |
| Mo | S | 0.000 ** | 0.537 | 0.029 * |
| O | 0.000 ** | 1.000 | 0.000 ** | |
| S | O | 0.206 | 0.680 | 0.002 ** |
| Model | Hyper-Parameters | OA (%) | |||
|---|---|---|---|---|---|
| NL | MD | LR | NE | ||
| Scenario-A | 17 | −1 | 0.1 | 77 | 81.12 |
| Scenario-B | 17 | −1 | 0.04 | 80 | 76.54 |
| H | Mi | Mo | S | O | GPA (%) | OA (%) | ||
|---|---|---|---|---|---|---|---|---|
| Scenario-A | H | 31 | 1 | 0 | 0 | 96.88 | 81.51 | |
| Mi | 0 | 23 | 8 | 0 | 74.19 | |||
| Mo | 0 | 8 | 15 | 4 | 55.56 | |||
| S | 0 | 0 | 1 | 28 | 96.55 | |||
| Scenario-B | H | 31 | 0 | 0 | 0 | 1 | 96.88 | 75.71 |
| Mi | 0 | 23 | 7 | 1 | 0 | 74.19 | ||
| Mo | 0 | 8 | 14 | 6 | 0 | 50.00 | ||
| S | 0 | 0 | 3 | 28 | 0 | 90.32 | ||
| O | 1 | 2 | 4 | 1 | 10 | 55.56 |
| Leaf Code | RCC | LWC | LNC | Indentation Area Ratio | Disease Spots Area Ratio | LLR |
|---|---|---|---|---|---|---|
| Mo-9 | 31.90 | 36.59% | 1.71% | 3.69% | 36.53% | 40.22% |
| Mo-54 | 36.10 | 44.23% | 2.08% | 13.62% | 33.16% | 46.78% |
| Mo-61 | 45.00 | 40.94% | 2.45% | 22.49% | 7.41% | 29.90% |
| Mo-90 | 43.10 | 53.72% | 2.33% | 43.54% | 3.61% | 47.16% |
| H-9 | 36.6 | 63.31% | 2.97% | 0.00% | 0.00% | 0.00% |
| H-10 | 34.4 | 57.05% | 2.88% | 0.00% | 0.00% | 0.01% |
| H-64 | 46.9 | 53.55% | 3.43% | 0.00% | 0.45% | 0.45% |
| H-81 | 45.8 | 51.70% | 2.94% | 0.00% | 0.00% | 0.00% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Xu, Z.; Yang, X.; Shi, J.; Hu, X.; Ju, W. Monitoring the Severity of Pantana phyllostachysae Chao on Bamboo Using Leaf Hyperspectral Data. Remote Sens. 2021, 13, 4146. https://doi.org/10.3390/rs13204146
Huang X, Xu Z, Yang X, Shi J, Hu X, Ju W. Monitoring the Severity of Pantana phyllostachysae Chao on Bamboo Using Leaf Hyperspectral Data. Remote Sensing. 2021; 13(20):4146. https://doi.org/10.3390/rs13204146
Chicago/Turabian StyleHuang, Xuying, Zhanghua Xu, Xu Yang, Jingming Shi, Xinyu Hu, and Weimin Ju. 2021. "Monitoring the Severity of Pantana phyllostachysae Chao on Bamboo Using Leaf Hyperspectral Data" Remote Sensing 13, no. 20: 4146. https://doi.org/10.3390/rs13204146
APA StyleHuang, X., Xu, Z., Yang, X., Shi, J., Hu, X., & Ju, W. (2021). Monitoring the Severity of Pantana phyllostachysae Chao on Bamboo Using Leaf Hyperspectral Data. Remote Sensing, 13(20), 4146. https://doi.org/10.3390/rs13204146