
Evaluation of Clumping Effects on the Estimation of Global Terrestrial Evapotranspiration

 ,
,  , , , , , and
, , , , , and
Abstract
:
1. Introduction
2. Modeling Methodology
2.1. ET Modeling
2.2. The Two-Leaf Model for ET Modeling
3. Input Data
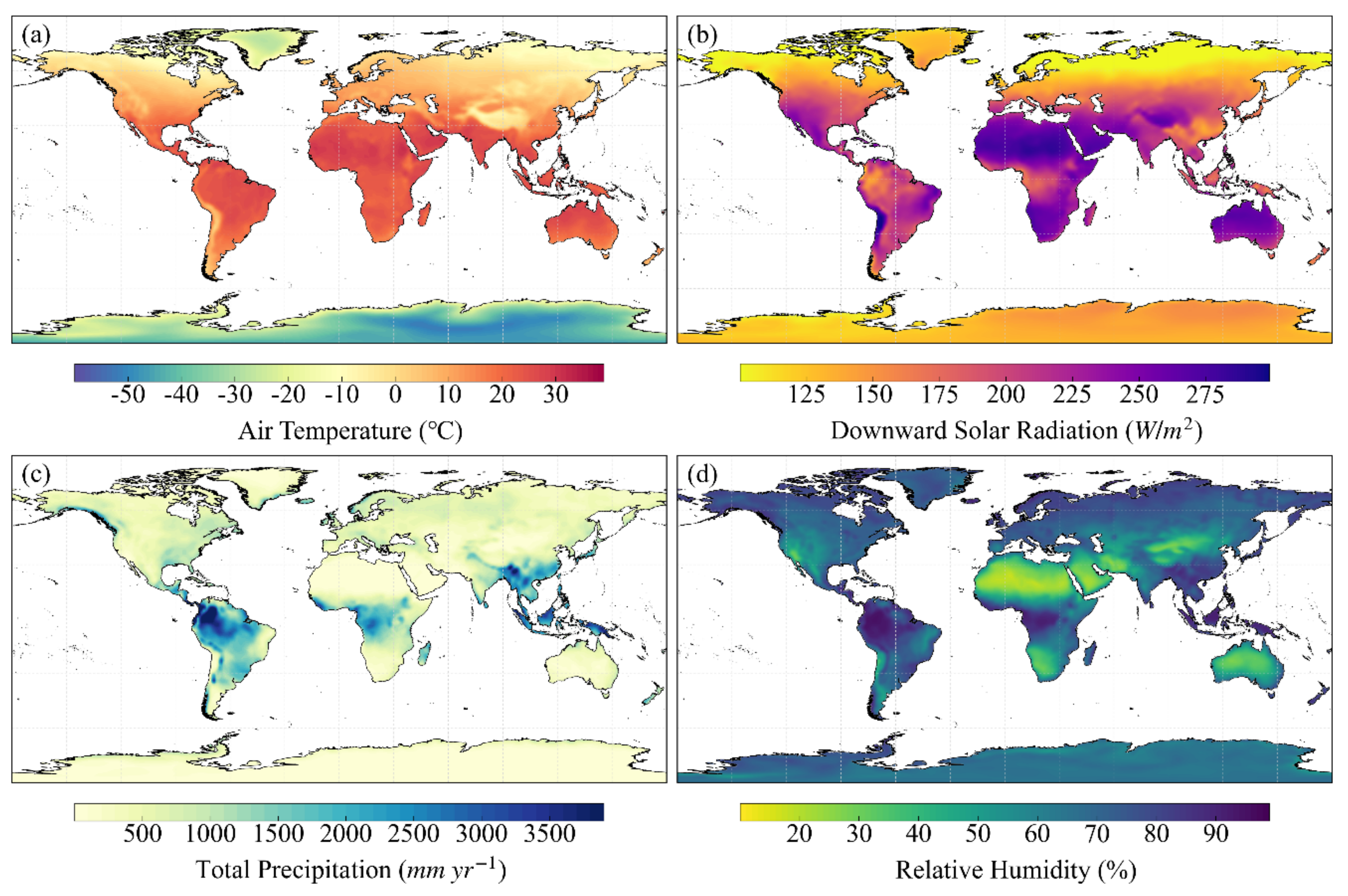
3.1. Meteorological Data
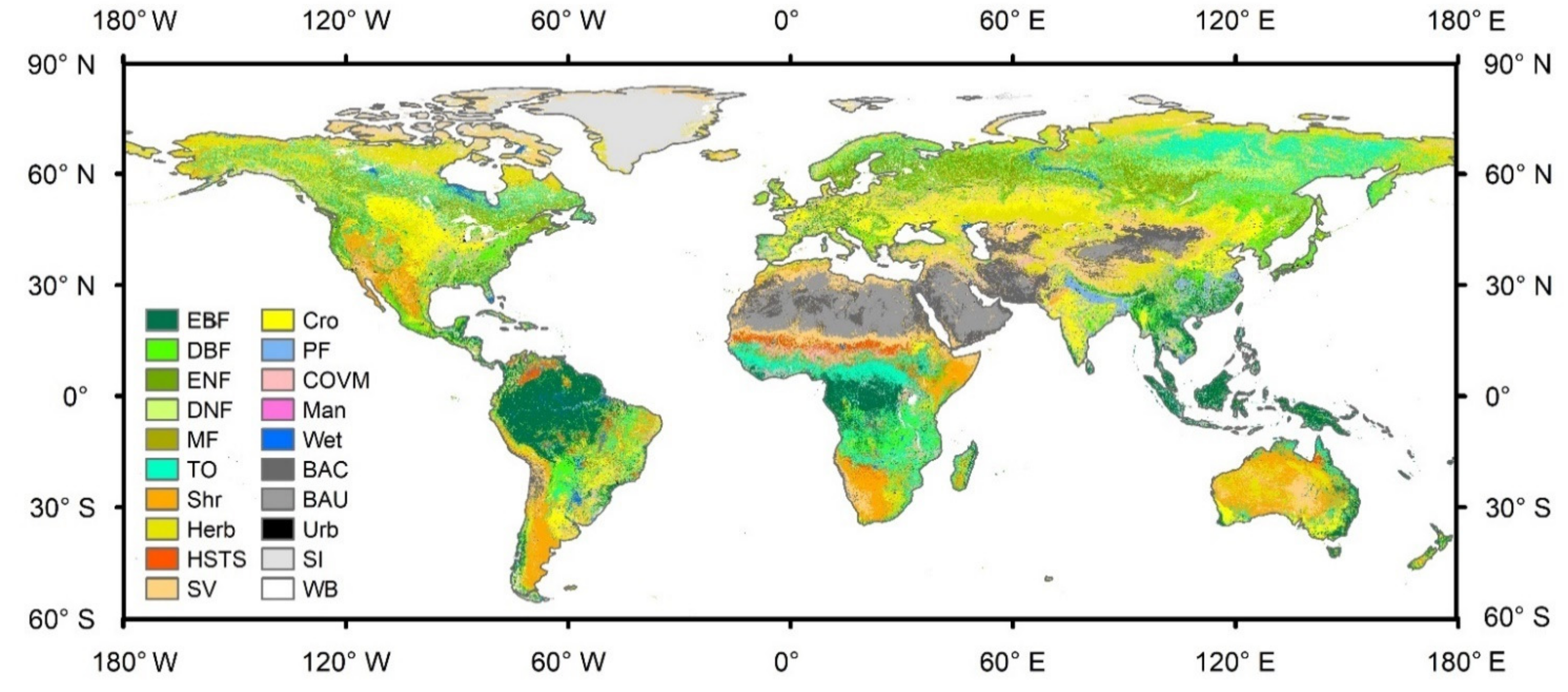
3.2. Land Cover Map
3.3. Global LAI Product
3.4. Global Clumping Index Map
3.5. Soil Texture Data
4. Simulation Cases and Model Validation
5. Results
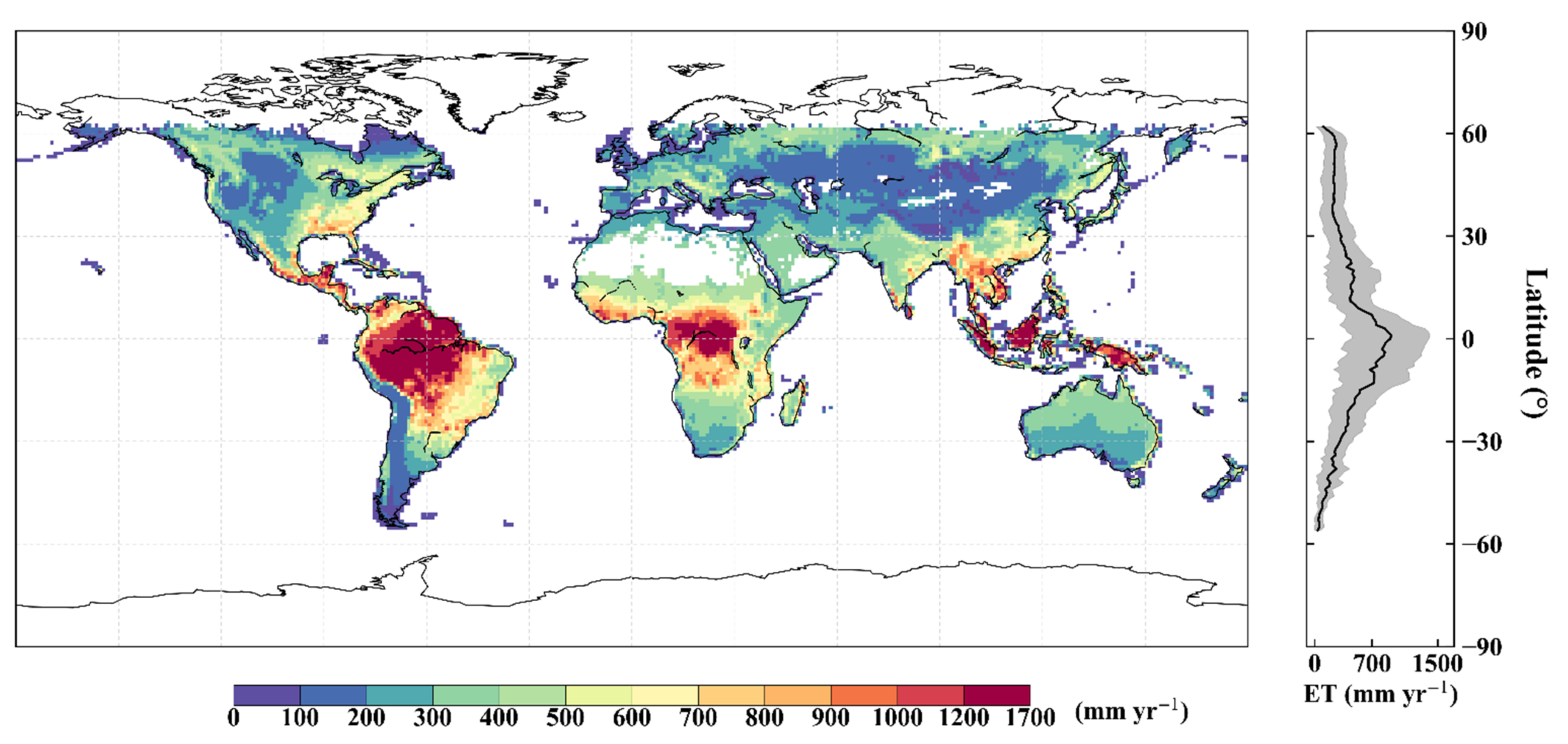
5.1. Simulations of Global Terrestrial ET
5.2. Spatial Patterns of Leaf Clumping Effects on ET Estimation
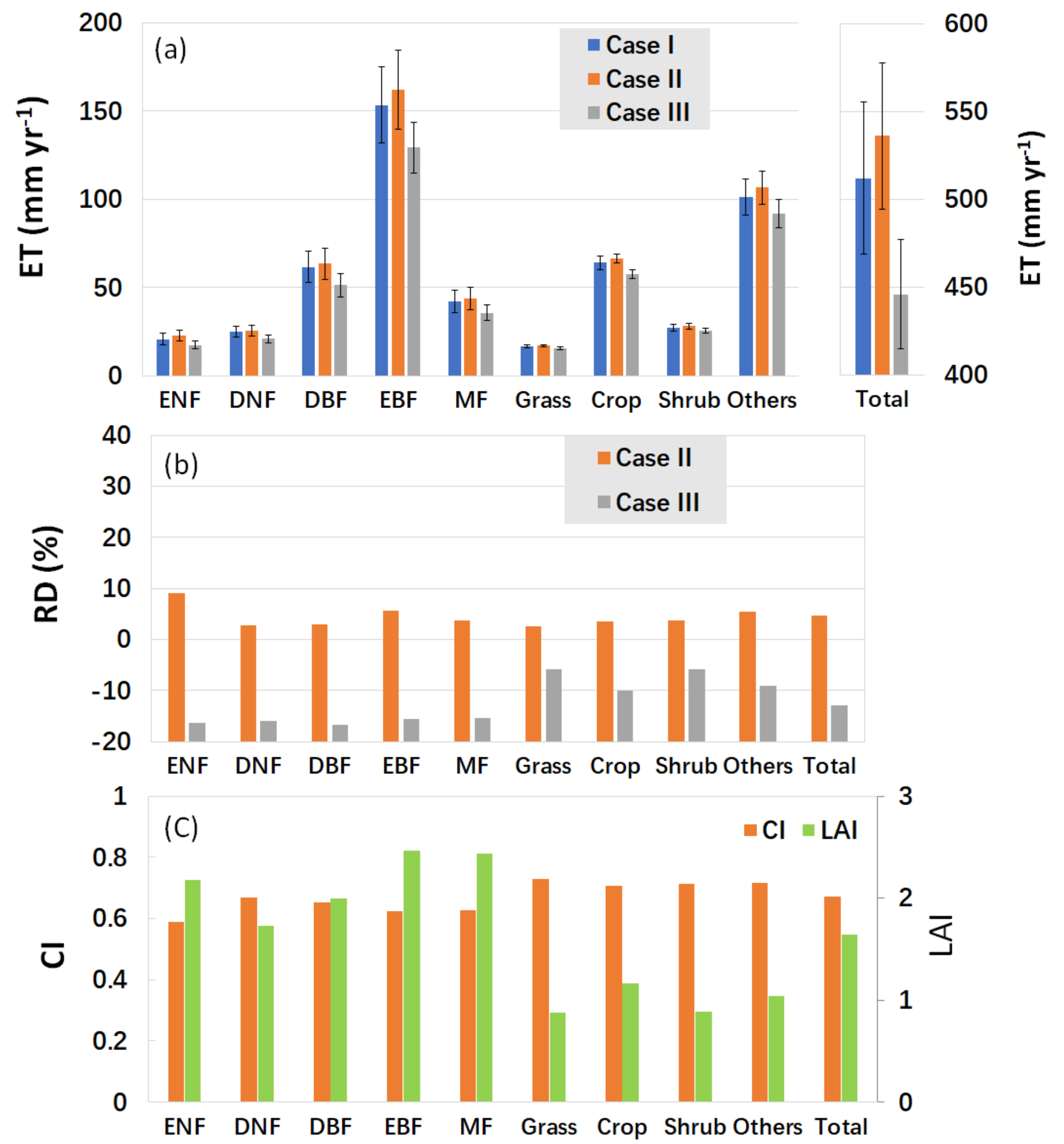
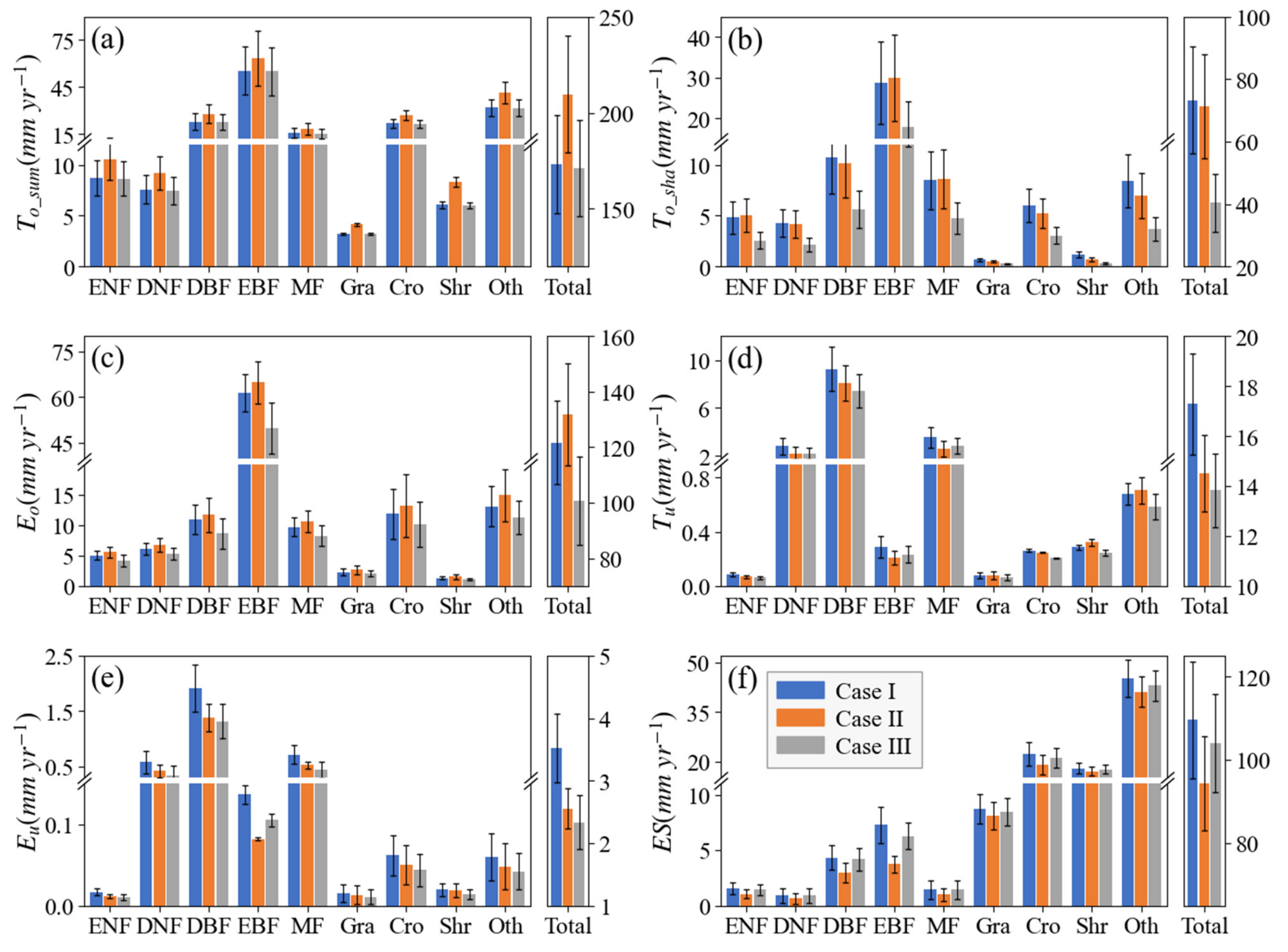
5.3. Leaf Clumping Effects on ET Estimation and Its Components for Various PFTs
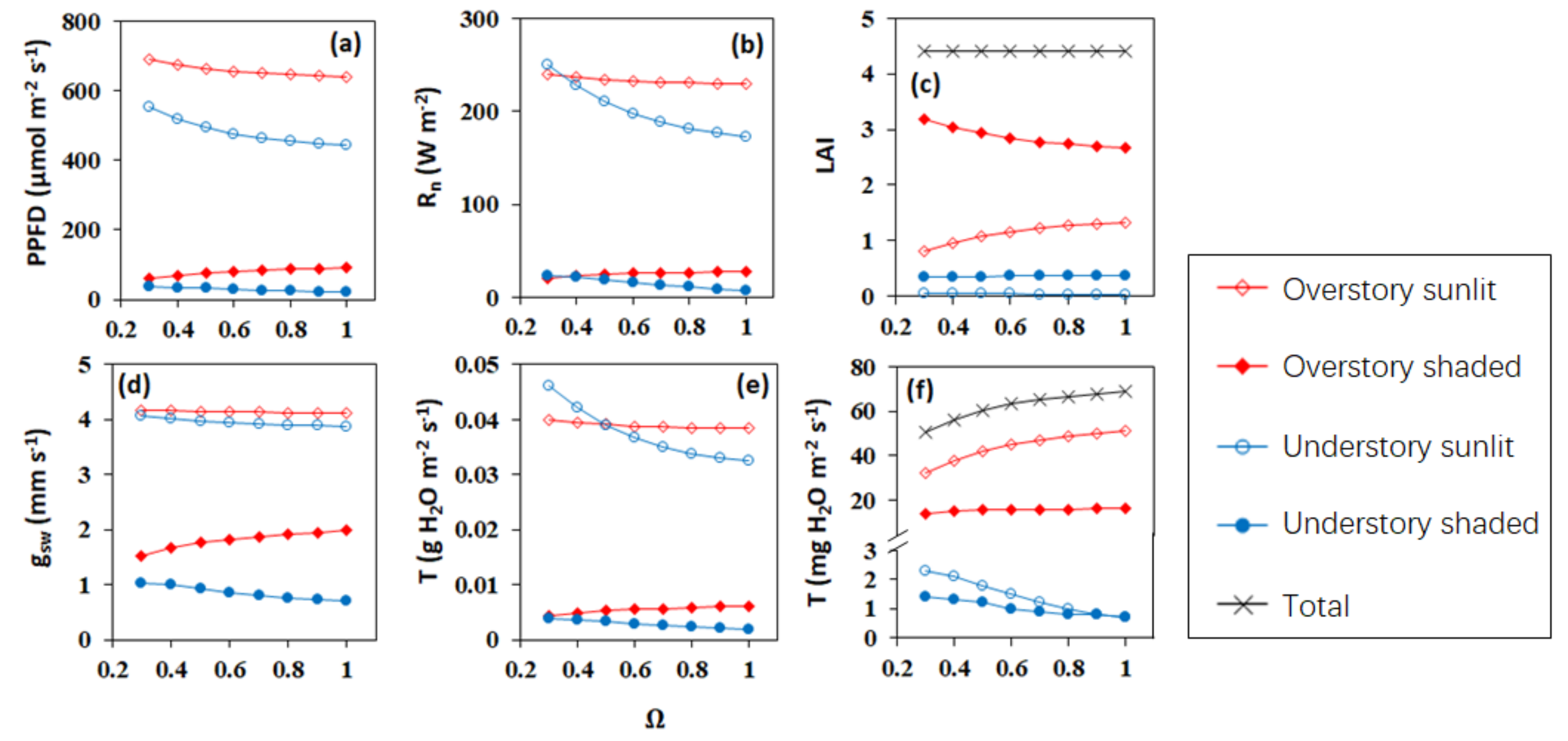
5.4. Leaf Clumping Effects on the Calculation of Key Biophysical Parameters Controlling Transpiration Simulations
6. Discussion
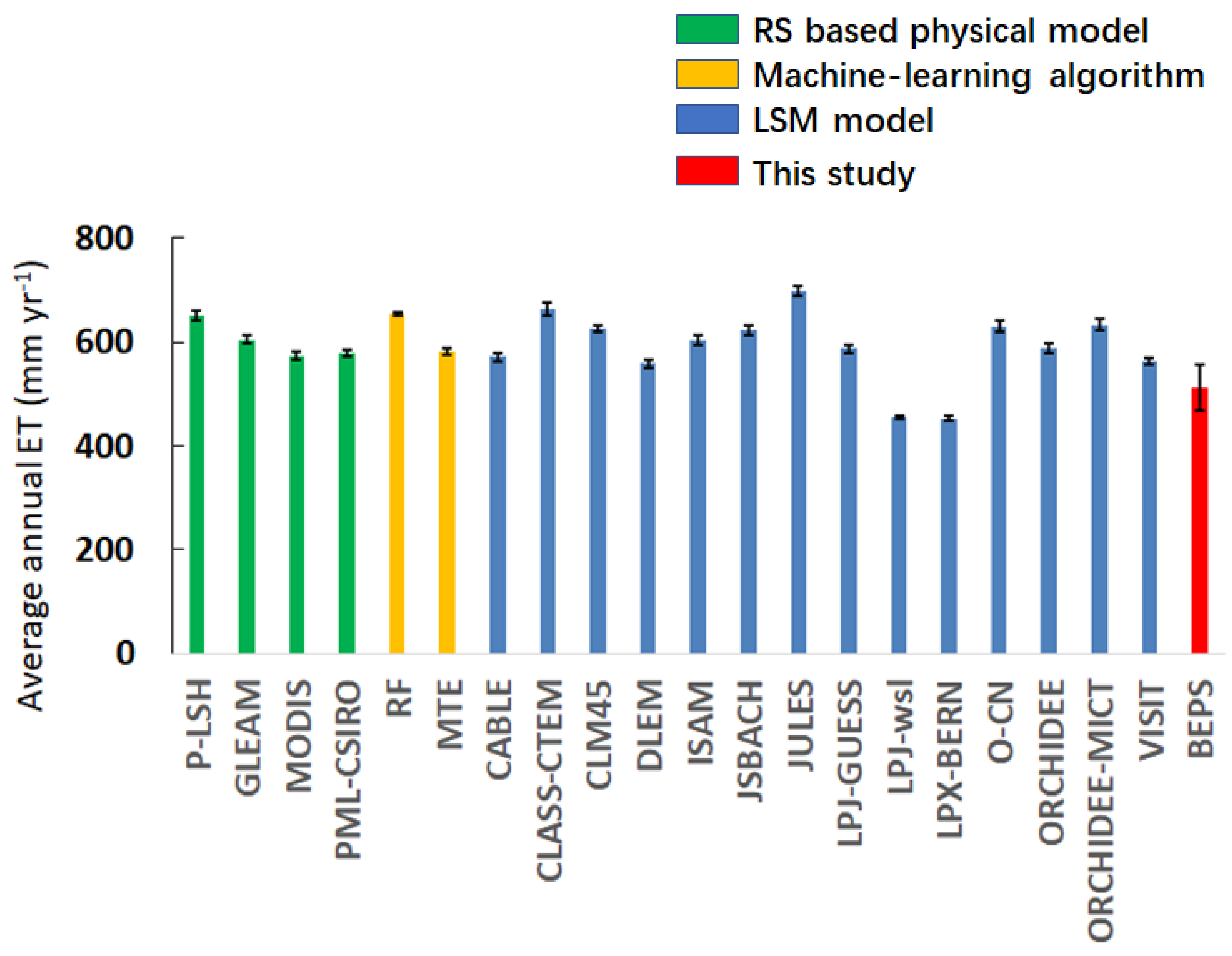
6.1. Estimations of Global Terrestrial ET
6.2. Sensitivities of ET to Errors in Key Parameters of the BEPS Model
6.3. Implications and Uncertainties of This Study
7. Conclusions
- Even though accurate global LAI data is used, neglecting leaf clumping will overestimate global terrestrial ET by about 4.7%. The reason is that leaf clumping reduces sunlit LAI while increases shaded LAI, resulting in an overall reduction in ET.
- If Le is used, neglecting leaf clumping will underestimate global terrestrial ET by 13.0%. The reason is that Le is lower than true LAI for a clumped canopy. If Le instead of true LAI is used in ET simulation ignoring leaf clumping impacts, shaded LAI would be substantially negatively biased while sunlit LAI is invariant, causing negative bias in ET.
- Although the accuracy of global terrestrial ET simulation still needs improvements, leaf clumping impacts quantified by the RDs between the model simulation cases considering or ignoring clumping are still robust since errors of key model parameters will move in the same directions for all cases.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Methods of Estimating Net Radiation for Canopies and the Ground
References
- Pan, S.; Pan, N.; Tian, H.; Friedlingstein, P.; Sitch, S.; Shi, H.; Arora, V.K.; Haverd, V.; Jain, A.K.; Kato, E.; et al. Evaluation of global terrestrial evapotranspiration using state-of-the-art approaches in remote sensing, machine learning and land surface modeling. Hydrol Earth Syst Sci. 2020, 24, 1485–1509. [Google Scholar] [CrossRef] [Green Version]
- Jasechko, S.; Sharp, Z.D.; Gibson, J.J.; Birks, S.J.; Yi, Y.; Fawcett, P.J. Terrestrial water fluxes dominated by transpiration. Nature 2013, 496, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Oki, T.; Kanae, S. Global hydrological cycles and world water resources. Science 2006, 313, 1068–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trenberth, K.E.; Fasullo, J.T.; Kiehl, J. Earth’s global energy budget. Bull Am. Meteorol. Soc. 2009, 90, 311–312. [Google Scholar] [CrossRef]
- Lian, X.; Piao, S.; Huntingford, C.; Li, Y.; Zeng, Z.; Wang, X.; Ciais, P.; McVicar, T.R.; Peng, S.; Ottlé, C.; et al. Partitioning global land evapotranspiration using CMIP5 models constrained by observations. Nat. Clim. Chang. 2018, 8, 640–646. [Google Scholar] [CrossRef]
- Sellers, P.J.; Dickinson, R.E.; Randall, D.A.; Betts, A.K.; Hall, F.G.; Berry, J.A.; Collatz, G.J.; Denning, A.S.; Mooney, H.A.; Nobre, C.A.; et al. Modeling the Exchanges of Energy, Water, and Carbon Between Continents and the Atmosphere. Science 1997, 275, 502–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Wang, S.Q.; Billesbach, D.; Oechel, W.; Zhang, J.H.; Meyers, T.; Martin, T.A.; Matamala, R.; Baldocchi, D.; Bohrer, G. Global estimation of evapotranspiration using a leaf area index-based surface energy and water balance model. Remote Sens Environ. 2012, 124, 581–595. [Google Scholar] [CrossRef] [Green Version]
- Dirmeyer, P.A.; Gao, X.; Zhao, M.; Guo, Z.; Oki, T.; Hanasaki, N. GSWP-2: Multimodel Analysis and Implications for Our Perception of the Land Surface. Bull Am Meteorol Soc. 2006, 87, 1381–1397. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Piao, S.; Lin, X.; Yin, G.; Peng, S.; Ciais, P.; Myneni, R.B. Global evapotranspiration over the past three decades: Estimation based on the water balance equation combined with empirical models. Environ. Res. Lett. 2012, 7, 014026. [Google Scholar] [CrossRef]
- Mu, Q.; Zhao, M.; Running, S.W. Improvements to a MODIS global terrestrial evapotranspiration algorithm. Remote Sens. Environ. 2011, 115, 1781–1800. [Google Scholar] [CrossRef]
- Luo, X.; Chen, J.M.; Liu, J.; Andrew Black, T.; Croft, H.; Staebler, R.; He, L.; Altaf Arain, M.; Chen, B.; Mo, G. Comparison of big-leaf, two-big-leaf and two-leaf upscaling schemes for evapotranspiration estimation using coupled carbon-water modelling. J. Geophys. Res. Biogeosci. 2018, 123, 207–225. [Google Scholar] [CrossRef]
- Penman, H.L. Natural evaporation from open water, bare soil and grass. Proceedings of the Royal Society of London A: Mathematical, Physical and Engineering Sciences. R. Soc. 1948, 193, 120–145. [Google Scholar]
- Monteith, J.L. Evaporation and environment. Symp. Soc. Exp. Biol. 1965, 19, 4. [Google Scholar]
- Chen, B.; Liu, J.; Chen, J.M.; Croft, H.; Gonsamo, A.; He, L.; Luo, X. Assessment of foliage clumping effects on evapotranspiration estimates in forested ecosystems. Agric. For. Meteorol. 2016, 216, 82–92. [Google Scholar] [CrossRef]
- Raupach, M.R.; Finnigan, J.J. “Single-layer models of evaporation from plant canopies are incorrect but useful, whereas multilayer models are correct but useless”: Discuss. Funct Plant Biol. 1988, 15, 705–716. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Murphy, C.E.; Knoerr, K.R. Development and Evaluation of Simplified Models for Simulating Canopy Photosynthesis and Transpiration. J. Appl. Ecol. 1976, 13, 813–829. [Google Scholar] [CrossRef]
- Baldocchi, D.; Meyers, T. On using eco-physiological, micrometeorological and biogeochemical theory to evaluate carbon dioxide, water vapor and trace gas fluxes over vegetation: A perspective. Agric. For. Meteorol. 1998, 90, 1–25. [Google Scholar] [CrossRef]
- Chen, J.M.; Liu, J.; Cihlar, J.; Goulden, M.L. Daily canopy photosynthesis model through temporal and spatial scaling for remote sensing applications. Ecol. Modell. 1999, 124, 99–119. [Google Scholar] [CrossRef] [Green Version]
- Sprintsin, M.; Chen, J.M.; Desai, A.; Gough, C.M. Evaluation of leaf-to-canopy upscaling methodologies against carbon flux data in North America. J. Geophys. Res. 2012, 117, G01023. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.M.; Black, T.A. Defining leaf area index for non-flat leaves. Plant Cell Environ. 1992, 15, 421–429. [Google Scholar] [CrossRef]
- Chen, J.M.; Black, T.A. Measuring leaf area index of plant canopies with branch architecture. Agric. For. Meteorol. 1991, 57, 1–12. [Google Scholar] [CrossRef]
- Chen, J.M.; Govind, A.; Sonnentag, O.; Zhang, Y.; Barr, A.; Amiro, B. Leaf area index measurements at Fluxnet-Canada forest sites. Agric. For. Meteorol. 2006, 140, 257–268. [Google Scholar] [CrossRef]
- Chen, J.M.; Mo, G.; Pisek, J.; Liu, J.; Deng, F.; Ishizawa, M.; Chan, D. Effects of foliage clumping on the estimation of global terrestrial gross primary productivity. Global Biogeochem. Cycles. 2012, 26, GB1019. [Google Scholar] [CrossRef]
- Chen, B.; Chen, J.M.; Ju, W. Remote sensing-based ecosystem–atmosphere simulation scheme (EASS)—Model formulation and test with multiple-year data. Ecol. Modell. 2007, 209, 277–300. [Google Scholar] [CrossRef]
- Lama, G.F.C.; Rillo Migliorini Giovannini, M.; Errico, A.; Mirzaei, S.; Padulano, R.; Chirico, G.B.; Preti, F. Hydraulic Efficiency of Green-Blue Flood Control Scenarios for Vegetated Rivers: 1D and 2D Unsteady Simulations. Water 2021, 13, 2620. [Google Scholar] [CrossRef]
- Box, W.; Järvelä, J.; Västilä, K. Flow resistance of floodplain vegetation mixtures for modelling river flows. J. Hydrol. 2021, 601, 126593. [Google Scholar] [CrossRef]
- Wei, S.; Fang, H.; Schaaf, C.B.; He, L.; Chen, J.M. Global 500 m clumping index product derived from MODIS BRDF data (2001–2017). Remote Sens Environ. 2019, 232, 111296. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.M.; Cihlar, J.; Chen, W. Net primary productivity mapped for Canada at 1-km resolution. Glob. Ecol. Biogeogr. 2002, 11, 115–129. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chen, J.M.; Cihlar, J.; Park, W.M. A process-based boreal ecosystem productivity simulator using remote sensing inputs. Remote Sens Environ. 1997, 62, 158–175. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.M.; Cihlar, J.; Chen, W. Net primary productivity distribution in the BOREAS region from a process model using satellite and surface data. J. Geophys. Res. 1999, 104, 27735–27754. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.M.; Cihlar, J. Mapping evapotranspiration based on remote sensing: An application to Canada’s landmass. Water Resour. Res. 2003, 39, 1189. [Google Scholar] [CrossRef]
- Matsushita, B.; Tamura, M. Integrating remotely sensed data with an ecosystem model to estimate net primary productivity in East Asia. Remote Sens. Environ. 2002, 81, 58–66. [Google Scholar] [CrossRef]
- Feng, X.; Liu, G.; Chen, J.M.; Chen, M.; Liu, J.; Ju, W.M.; Sun, R.; Zhou, W. Net primary productivity of China’s terrestrial ecosystems from a process model driven by remote sensing. J. Environ. Manag. 2007, 85, 563–573. [Google Scholar] [CrossRef]
- He, L.; Chen, J.M.; Liu, J.; Zheng, T.; Wang, R.; Joiner, J.; Chou, S.; Chen, B.; Liu, Y.; Liu, R.; et al. Diverse photosynthetic capacity of global ecosystems mapped by satellite chlorophyll fluorescence measurements. Remote Sens. Environ. 2019, 232, 111344. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef]
- Chen, B.; Chen, J.M.; Baldocchi, D.D.; Liu, Y.; Wang, S.; Zheng, T.; Black, T.A.; Croft, H. Including soil water stress in process-based ecosystem models by scaling down maximum carboxylation rate using accumulated soil water deficit. Agric. For. Meteorol. 2019, 276–277. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, R.; Chen, J.M. Retrospective retrieval of long-term consistent global leaf area index (1981–2011) from combined AVHRR and MODIS data. J. Geophys. Res. Biogeosc. 2012, 117. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, R.; Pisek, J.; Chen, J.M. Separating overstory and understory leaf area indices for global needleleaf and deciduous broadleaf forests by fusion of MODIS and MISR data. Biogeosciences 2017, 14, 1093–1110. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Fang, H. Estimation of canopy clumping index from MISR and MODIS sensors using the normalized difference hotspot and darkspot (NDHD) method: The influence of BRDF models and solar zenith angle. Remote Sens. Environ. 2016, 187, 476–491. [Google Scholar] [CrossRef]
- Murray, F.W. On the Computation of Saturation Vapor Pressure. J. Appl. Meteorol. 1967, 6, 203–204. [Google Scholar] [CrossRef]
- Ball, J.T.; Woodrow, I.E.; Berry, J.A. A Model Predicting Stomatal Conductance and its Contribution to the Control of Photosynthesis under Different Environmental Conditions. In Progress in Photosynthesis Research; Springer: Dordrecht, The Netherlands, 1987. [Google Scholar] [CrossRef]
- Miner, G.L.; Bauerle, W.L.; Baldocchi, D.D. Estimating the sensitivity of stomatal conductance to photosynthesis: A review. Plant Cell Environ. 2017, 40, 1214–1238. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Caemmerer, S.V.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C-3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Chen, J.M.; Liu, J.; Mo, G.; Joiner, J. Angular normalization of GOME-2 Sun-induced chlorophyll fluorescence observation as a better proxy of vegetation productivity. Geophys. Res. Lett. 2017, 44, 5691–5699. [Google Scholar] [CrossRef]
- Sellers, P.; Randall, D.; Collatz, G.; Berry, J.; Field, C.; Dazlich, D.; Zhang, C.; Collelo, G.; Bounoua, L. A Revised Land Surface Parameterization (SiB2) for Atmospheric GCMS. Part I: Model Formulation. J. Clim. 1996, 9, 676–705. [Google Scholar] [CrossRef]
- Norman, J.M. Simulation of microclimates. In Biometeorology in Integrated Pest Management; Hatfield, J.L., Thomason, I.J., Eds.; Academic Press: New York, NY, USA, 1982; pp. 65–99. [Google Scholar] [CrossRef]
- Dee, D.P.; Uppala, S.M.; Simmons, A.J.; Berrisford, P.; Poli, P.; Kobayashi, S.; Andrae, U.; Balmaseda, M.A.; Balsamo, G.; Bauer, P.; et al. The ERA-Interim reanalysis: Configuration and performance of the data assimilation system. Q. J. R. Meteorol. Soc. 2011, 137, 553–597. [Google Scholar] [CrossRef]
- Kobayashi, T.; Tateishi, R.; Alsaaideh, B.; Sharma, R.C.; Wakaizumi, T.; Miyamoto, D.; Bai, X.; Long, B.D.; Gegentana, G.; Maitiniyazi, A.; et al. Production of Global Land Cover Data—GLCNMO2013. J. Geogr. Geol. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Chen, J.M.; Mo, G.; Yuen, C.-W.; Margolis, H.; Higuchi, K.; Chan, D. Modeling and Scaling Coupled Energy, Water, and Carbon Fluxes Based on Remote Sensing: An Application to Canada’s Landmass. J. Hydrometeorol. 2007, 8, 123–143. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Liu, Y. Generation of new cloud masks from MODIS land surface reflectance products. Remote Sens. Environ. 2013, 133, 21–37. [Google Scholar] [CrossRef]
- Chen, J.M.; Feng, D.; Mingzhen, C. Locally adjusted cubic-spline capping for reconstructing seasonal trajectories of a satellite-derived surface parameter. Geosci Remote Sens. IEEE Trans. 2006, 44, 2230–2238. [Google Scholar] [CrossRef]
- He, L.; Chen, J.M.; Pisek, J.; Schaaf, C.B.; Strahler, A.H. Global clumping index map derived from the MODIS BRDF product. Remote Sens Environ. 2012, 119, 118–130. [Google Scholar] [CrossRef]
- Chen, J.M.; Menges, C.H.; Leblanc, S.G. Global mapping of foliage clumping index using multi-angular satellite data. Remote Sens. Environ. 2005, 97, 447–457. [Google Scholar] [CrossRef]
- Marambe, Y.; Simic Milas, A. Modeling Evapotranspiration for C4 and C3 CROPS in the Western lake Erie basin using remote sensing data. In International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences—ISPRS Archives; Copernicus Publications: Göttingen, Germany, 2020. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Ge, Q.; Zhan, X.; Guan, F.; Yao, S. Drought monitoring based on simulated surface evapotranspiration by BEPS model. J Nat Disasters. 2014, 23, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.S.; Liu, Y.G.; Hu, Z.H.; Liu, Y.B.; Zhang, F.C.; Han, X.M. Evaluating the applicability of ecological model for simulating evapotranspiration and soil water content in winter wheat farmland. Chin. J. Ecol. 2017, 36. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Y.; Ju, W.; Chen, J.; Wang, S.; He, H.; Wang, H.; Guan, D.; Zhao, F.; Li, Y.; et al. Evapotranspiration and water yield over China’s landmass from 2000 to 2010. Hydrol. Earth Syst. Sci. 2013, 17, 4957–4980. [Google Scholar] [CrossRef] [Green Version]
- Ju, W.; Gao, P.; Wang, J.; Zhou, Y.; Zhang, X. Combining an ecological model with remote sensing and GIS techniques to monitor soil water content of croplands with a monsoon climate. Agric. Water Manag. 2010, 97, 1221–1231. [Google Scholar] [CrossRef]
- Chen, J.; Chen, B.; Black, T.A.; Innes, J.L.; Wang, G.; Kiely, G.; Hirano, T.; Wohlfahrt, G. Comparison of terrestrial evapotranspiration estimates using the mass transfer and Penman-Monteith equations in land surface models. J. Geophys. Res. Biogeosc. 2013, 118, 1715–1731. [Google Scholar] [CrossRef]
- Jung, M.; Reichstein, M.; Ciais, P.; Seneviratne, S.I.; Sheffield, J.; Goulden, M.L.; Bonan, G.; Cescatti, A.; Chen, J.; De Jeu, R.; et al. Recent decline in the global land evapotranspiration trend due to limited moisture supply. Nature 2010, 467, 951–954. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Bodesheim, P.; Jung, M.; Gans, F.; Mahecha, M.D.; Reichstein, M. Upscaled diurnal cycles of land-Atmosphere fluxes: A new global half-hourly data product. Earth Syst. Sci. Data. 2018, 10, 1327–1365. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Baret, F.; Plummer, S.; Schaepman-Strub, G. An Overview of Global Leaf Area Index (LAI): Methods, Products, Validation, and Applications. Rev. Geophys. 2019, 57, 739–799. [Google Scholar] [CrossRef]
- Pinty, B.; Andredakis, I.; Clerici, M.; Kaminski, T.; Taberner, M.; Verstraete, M.M.; Gobron, N.; Plummer, S.; Widlowski, J.L. Exploiting the MODIS albedos with the Two-Stream Inversion Package (JRC-TIP): 1. Effective leaf area index, vegetation, and soil properties. J. Geophys. Res. Atmos. 2011, 116. [Google Scholar] [CrossRef]
- Disney, M.; Muller, J.P.; Kharbouche, S.; Kaminski, T.; Voßbeck, M.; Lewis, P.; Pinty, B. A new global fAPAR and LAI dataset derived from optimal albedo estimates: Comparison with MODIS products. Remote Sens. 2016, 8, 275. [Google Scholar] [CrossRef] [Green Version]
- García-Haro, F.J.; Campos-Taberner, M.; Muñoz-Marí, J.; Laparra, V.; Camacho, F.; Sánchez-Zapero, J.; Camps-Valls, G. Derivation of global vegetation biophysical parameters from EUMETSAT Polar System. ISPRS J. Photogramm. Remote Sens. 2018, 139, 57–74. [Google Scholar] [CrossRef]
- Pisek, J.; Govind, A.; Arndt, S.K.; Hocking, D.; Wardlaw, T.J.; Fang, H.; Matteucci, G.; Longdoz, B. Intercomparison of clumping index estimates from POLDER, MODIS, and MISR satellite data over reference sites. ISPRS J. Photogramm. Remote Sens. 2015, 101, 47–56. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters a | ENF b | DNF b | DBF b | EBF b | MFb | Grass | Crop | Shrub | Others | References |
|---|---|---|---|---|---|---|---|---|---|---|
| () | 48.2 ± 5.7 | 44.6 ± 2.7 | 72.7 ± 14.7 | 53.6 ± 14.7 | 60.5 ± 10.2 | 88.7 ± 20.9 | 82.7 ± 15.2 | 57.4 ± 31.0 | 90.0 ± 89.5 | [23,34] |
| N0 (g m−2) | 4.45 | 2.45 | 2.45 | 2.97 | 3.5 | 2.38 | 2.38 | 2.70 | 2.38 | [23] |
| 0.33 | 0.56 | 0.59 | 0.48 | 0.47 | 0.60 | 0.60 | 0.57 | 0.60 | [23] | |
| m | 8 | 8 | 8 | 8 | 8 | 4 | 4 | 8 | 8 | [23] |
| 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | [23] | |
| 0.976 | 0.966 | 0.966 | 0.962 | 0.966 | 0.952 | 0.961 | 0.978 | 0.966 | [35] | |
| 0.6 | 0.6 | 0.885 | 0.575 | 0.6 | 0.495 | 0.6 | 0.6 | 0.6 | [36] | |
| a | −0.03 | −0.03 | −0.03 | −0.03 | −0.03 | −0.04 | −0.03 | −0.03 | −0.03 | [36] |
| LAI | 3.60 ± 2.14 | 3.43 ± 1.96 | 3.61 ± 2.01 | 3.98 ± 2.25 | 4.39 ± 2.01 | 1.64 ± 1.36 | 2.27 ± 1.60 | 1.71 ± 1.33 | 1.87 ± 1.64 | [37] |
| LAIu | 0.03 | 1.04 | 0.96 | 0.03 | 0.96 | 0.01 | 0.01 | 0.01 | 0.01 | [38] |
| CI | 0.58 ± 0.10 | 0.60 ± 0.11 | 0.60 ± 0.11 | 0.58 ± 0.12 | 0.57 ± 0.11 | 0.66 ± 0.14 | 0.68 ± 0.13 | 0.65 ± 0.14 | 0.65 ± 0.14 | [39] |
| Case I a | Case II | Case III | Case II–Case I b | Case III–Case I b | |
|---|---|---|---|---|---|
| Baseline c | 511.9 | 535.9 | 445.6 | 24.0 (4.7%) | −66.3 (−13.0%) |
| 1.2*LAI | 541.6 (5.8%) | 565.0 | 473.9 | 23.4 (4.3%) | −67.7 (−12.5%) |
| 0.8*LAI | 476.6 (−6.9%) | 501.1 | 413.3 | 24.6 (5.2%) | −63.3 (−13.3%) |
| 1.2*Ω | 521.6 (1.9%) | 535.9 | 476.4 | 14.3 (2.7%) | −45.3 (−8.7%) |
| 0.8*Ω | 499.1 (−2.5%) | 535.9 | 409.9 | 36.8 (7.4%) | −89.2 (−17.9%) |
| 525.2 (2.6%) | 551.6 | 458.7 | 26.4 (5.0%) | −66.5 (−12.7%) | |
| 494.5 (−3.4%) | 515.9 | 429.1 | 21.4 (4.3%) | −65.4 (−13.2%) | |
| 1.2*RASMth | 503.7 (−1.6%) | 526.0 | 437.5 | 22.3 (4.4%) | −66.2 (−13.1%) |
| 0.8*RASMth | 522.6 (2.1%) | 548.5 | 456.1 | 25.9 (5.0%) | −66.6 (−12.7%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.; Lu, X.; Wang, S.; Chen, J.M.; Liu, Y.; Fang, H.; Liu, Z.; Jiang, F.; Arain, M.A.; Chen, J.; et al. Evaluation of Clumping Effects on the Estimation of Global Terrestrial Evapotranspiration. Remote Sens. 2021, 13, 4075. https://doi.org/10.3390/rs13204075
Chen B, Lu X, Wang S, Chen JM, Liu Y, Fang H, Liu Z, Jiang F, Arain MA, Chen J, et al. Evaluation of Clumping Effects on the Estimation of Global Terrestrial Evapotranspiration. Remote Sensing. 2021; 13(20):4075. https://doi.org/10.3390/rs13204075
Chicago/Turabian StyleChen, Bin, Xuehe Lu, Shaoqiang Wang, Jing M. Chen, Yang Liu, Hongliang Fang, Zhenhai Liu, Fei Jiang, Muhammad Altaf Arain, Jinghua Chen, and et al. 2021. "Evaluation of Clumping Effects on the Estimation of Global Terrestrial Evapotranspiration" Remote Sensing 13, no. 20: 4075. https://doi.org/10.3390/rs13204075
APA StyleChen, B., Lu, X., Wang, S., Chen, J. M., Liu, Y., Fang, H., Liu, Z., Jiang, F., Arain, M. A., Chen, J., & Wang, X. (2021). Evaluation of Clumping Effects on the Estimation of Global Terrestrial Evapotranspiration. Remote Sensing, 13(20), 4075. https://doi.org/10.3390/rs13204075