Predicting the Presence of Leptospires in Rodents from Environmental Indicators Opens Up Opportunities for Environmental Monitoring of Human Leptospirosis

 , ,
, ,  , , and
, , and
Abstract
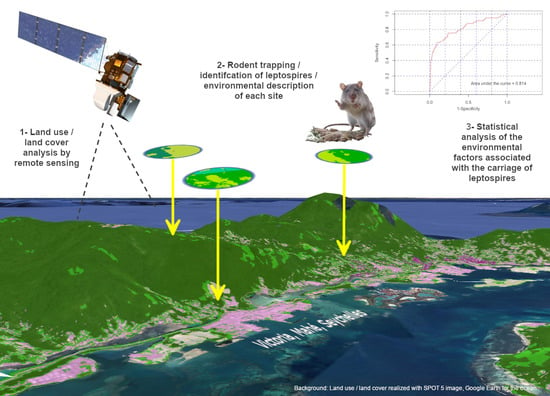
1. Introduction
2. Materials and Methods
2.1. Sampling Permit and Ethics Approval
2.2. Animal Sampling and Leptospira Detection
2.3. Landscape Analysis
2.4. Meteorological Data
2.5. Cartography
2.6. Statistical Analyses
3. Results
3.1. Trapping Success
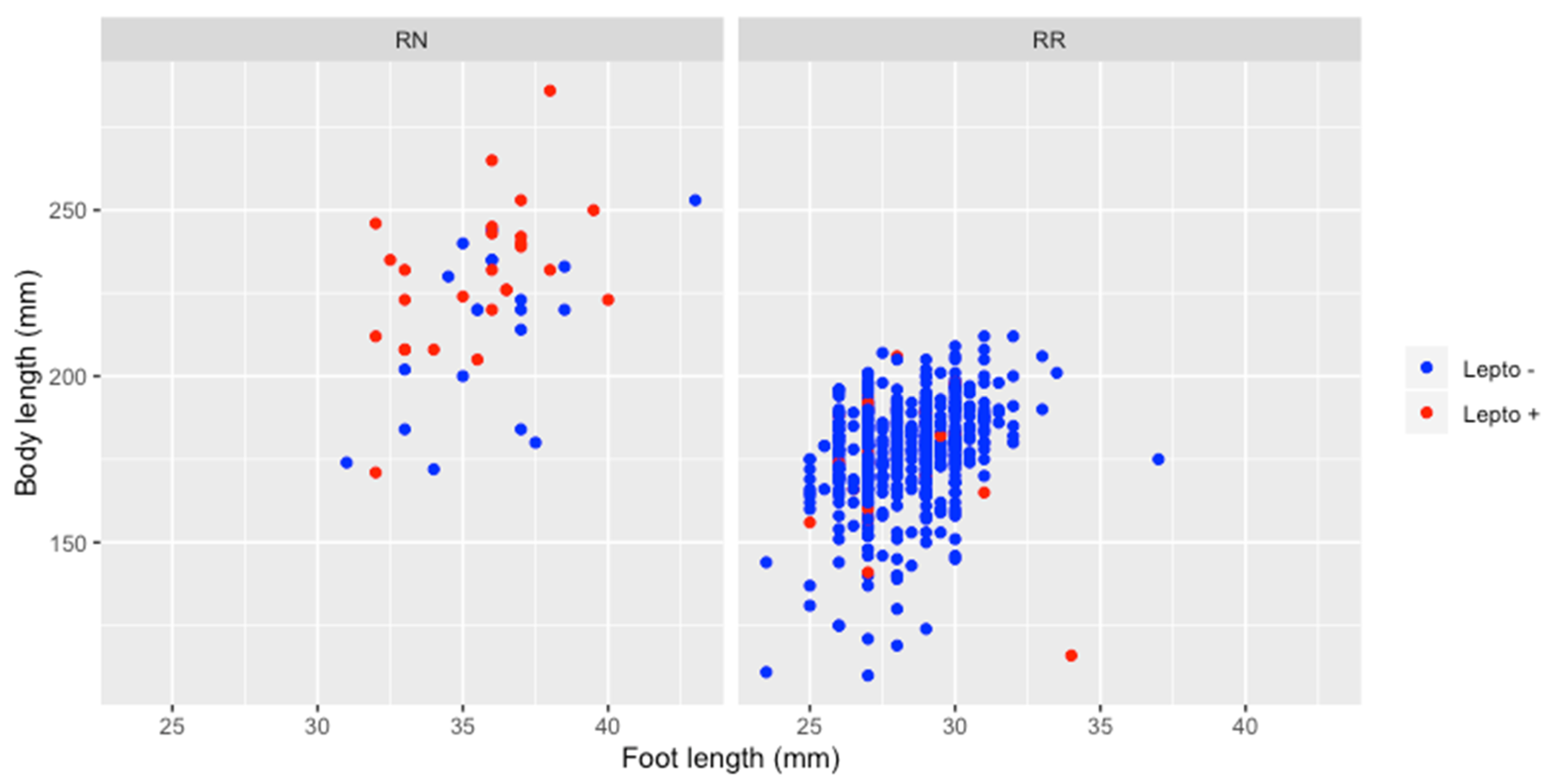
3.2. Rat Characteristics
3.3. Renal Carriage of Leptospira spp.
3.4. Ecological Pattern of Rat Distribution
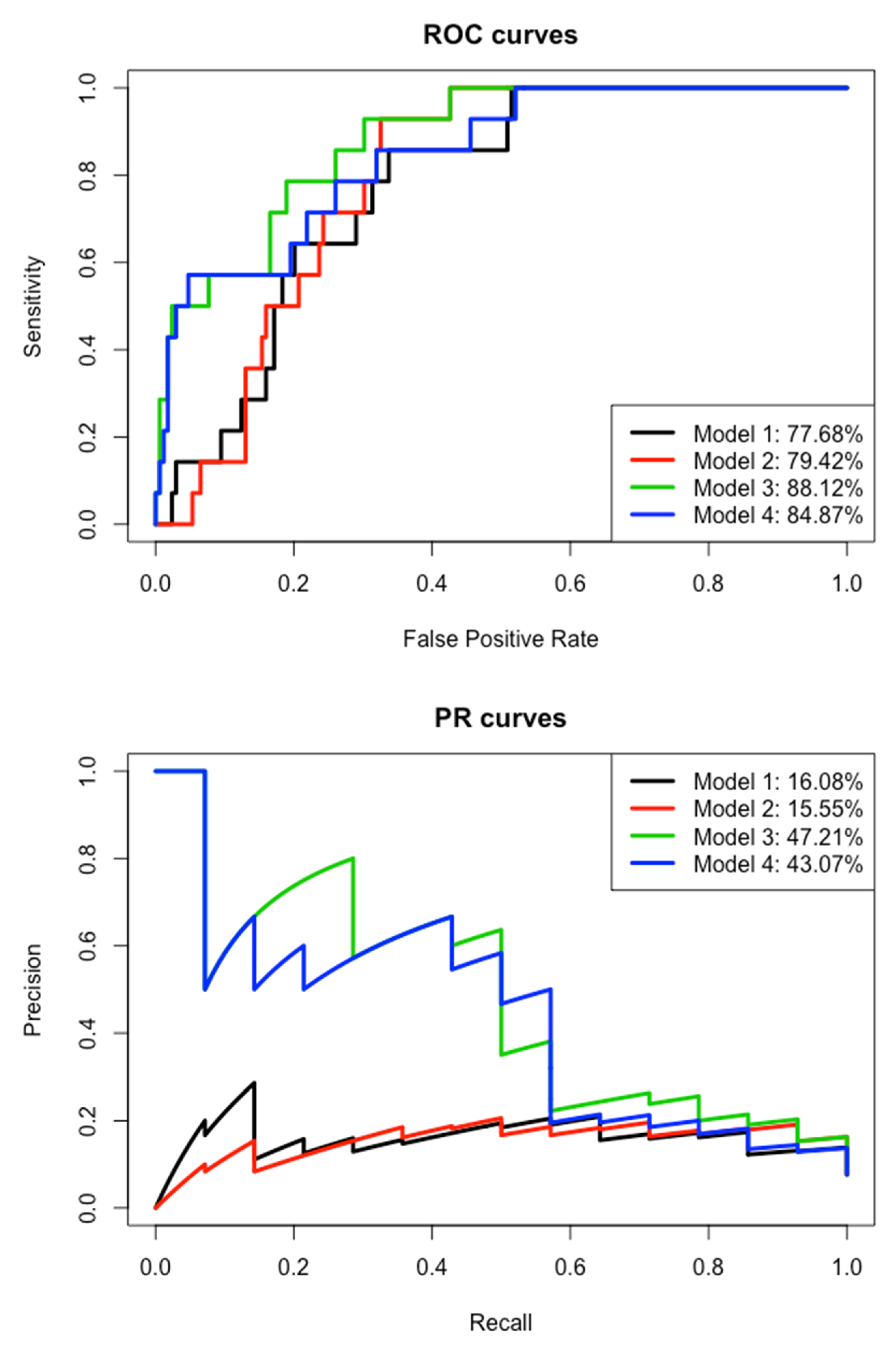
3.5. Environmental Prediction of Leptospires Carriage
4. Discussion
4.1. Rattus Diversity and Abundance
4.2. Leptospira-Laden Rattus spp.
4.3. Rattus Characteristics in Relation to Infection Rates
4.4. Environmental Factors in Relation to Rat Distribution and Infection Rates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ko, A.I.; Goarant, C.; Picardeau, M. Leptospira: The dawn of the molecular genetics era for an emerging zoonotic pathogen. Nat. Rev. Microbiol. 2009, 7, 736–747. [Google Scholar] [CrossRef]
- Fouts, D.E.; Matthias, M.A.; Adhikarla, H.; Adler, B.; Amorim-Santos, L.; Berg, D.E.; Bulach, D.; Buschiazzo, A.; Chang, Y.-F.; Galloway, R.L.; et al. What Makes a Bacterial Species Pathogenic?: Comparative Genomic Analysis of the Genus Leptospira. PLoS Negl. Trop. Dis. 2016, 10, e0004403. [Google Scholar] [CrossRef]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E.; et al. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Desvars, A.; Cardinale, E.; Michault, A. Animal leptospirosis in small tropical areas. Epidemiol. Infect. 2010, 139, 1–22. [Google Scholar] [CrossRef]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef] [PubMed]
- Picardeau, M. Leptospirosis: Updating the Global Picture of an Emerging Neglected Disease. PLoS Negl. Trop. Dis. 2015, 9, e0004039. [Google Scholar] [CrossRef]
- Durski, K.N.; Jancloes, M.; Chowdhary, T.; Bertherat, E. A Global, Multi-Disciplinary, Multi-Sectorial Initiative to Combat Leptospirosis: Global Leptospirosis Environmental Action Network (GLEAN). Int. J. Environ. Res. Public Health 2014, 11, 6000–6008. [Google Scholar] [CrossRef] [PubMed]
- Mwachui, M.A.; Crump, L.; Hartskeerl, R.; Zinsstag, J.; Hattendorf, J. Environmental and Behavioural Determinants of Leptospirosis Transmission: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003843. [Google Scholar] [CrossRef]
- Allan, K.J.; Biggs, H.M.; Halliday, J.E.B.; Kazwala, R.R.; Maro, V.P.; Cleaveland, S.; Crump, J.A. Epidemiology of Leptospirosis in Africa: A Systematic Review of a Neglected Zoonosis and a Paradigm for ‘One Health’ in Africa. PLoS Negl. Trop. Dis. 2015, 9, e0003899. [Google Scholar] [CrossRef]
- Hartskeerl, R.A.; Collares-Pereira, M.; Ellis, W.A. Emergence, control and re-emerging leptospirosis: Dynamics of infection in the changing world. Clin. Microbiol. Infect. 2011, 17, 494–501. [Google Scholar] [CrossRef]
- Andre-Fontaine, G.; Aviat, F.; Thorin, C. Waterborne Leptospirosis: Survival and Preservation of the Virulence of Pathogenic Leptospira spp. in Fresh Water. Curr. Microbiol. 2015, 71, 136–142. [Google Scholar] [CrossRef]
- Thibeaux, R.; Geroult, S.; Benezech, C.; Chabaud, S.; Soupé-Gilbert, M.-E.; Girault, D.; Bierque, E.; Goarant, C. Seeking the environmental source of Leptospirosis reveals durable bacterial viability in river soils. PLoS Negl. Trop. Dis. 2017, 11. [Google Scholar] [CrossRef]
- Gonzalez-Astudillo, V.; Bustamante-Rengifo, J.A.; Bonilla, Á.; Lehmicke, A.J.J.; Castillo, A.; Astudillo-Hernández, M. Synanthropic Cockroaches (Blattidae: Periplaneta spp.) Harbor Pathogenic Leptospira in Colombia. J. Med. Entomol. 2016, 53, 177–182. [Google Scholar] [CrossRef]
- Biscornet, L.; Dellagi, K.; Pagès, F.; Bibi, J.; de Comarmond, J.; Mélade, J.; Govinden, G.; Tirant, M.; Gomard, Y.; Guernier, V.; et al. Human leptospirosis in Seychelles: A prospective study confirms the heavy burden of the disease but suggests that rats are not the main reservoir. PLoS Negl. Trop. Dis. 2017, 11, e0005831. [Google Scholar] [CrossRef]
- Herbreteau, V. When geography of health meets health ecology. In The Biogeography of Host-Parasite Interaction; Morand, S., Krasnov, B., Eds.; Oxford University Press: Oxford, NY, USA, 2010; pp. 247–265. ISBN 978-0-19-956135-3. [Google Scholar]
- Soberon, J.; Peterson, A.T. Interpretation of Models of Fundamental Ecological Niches and Species’ Distributional Areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef]
- Lau, C.L.; Smythe, L.D.; Craig, S.B.; Weinstein, P. Climate change, flooding, urbanisation and leptospirosis: Fuelling the fire? Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 631–638. [Google Scholar] [CrossRef]
- Ledien, J.; Sorn, S.; Hem, S.; Huy, R.; Buchy, P.; Tarantola, A.; Cappelle, J. Assessing the performance of remotely-sensed flooding indicators and their potential contribution to early warning for leptospirosis in Cambodia. PLoS ONE 2017, 12, e0181044. [Google Scholar] [CrossRef]
- Weinberger, D.; Baroux, N.; Grangeon, J.-P.; Ko, A.I.; Goarant, C. El Niño Southern Oscillation and Leptospirosis Outbreaks in New Caledonia. PLoS Negl. Trop. Dis. 2014, 8, e2798. [Google Scholar] [CrossRef]
- Schneider, M.C.; Najera, P.; Pereira, M.M.; Machado, G.; dos Anjos, C.B.; Rodrigues, R.O.; Cavagni, G.M.; Muñoz-Zanzi, C.; Corbellini, L.G.; Leone, M.; et al. Leptospirosis in Rio Grande do Sul, Brazil: An Ecosystem Approach in the Animal-Human Interface. PLoS Negl. Trop. Dis. 2015, 9, e0004095. [Google Scholar] [CrossRef]
- Hagan, J.E.; Moraga, P.; Costa, F.; Capian, N.; Ribeiro, G.S.; Wunder, E.A., Jr.; Felzemburgh, R.D.M.; Reis, R.B.; Nery, N.; Santana, F.S.; et al. Spatiotemporal Determinants of Urban Leptospirosis Transmission: Four-Year Prospective Cohort Study of Slum Residents in Brazil. PLoS Negl. Trop. Dis. 2016, 10, e0004275. [Google Scholar] [CrossRef]
- Cosson, J.-F.; Picardeau, M.; Mielcarek, M.; Tatard, C.; Chaval, Y.; Suputtamongkol, Y.; Buchy, P.; Jittapalapong, S.; Herbreteau, V.; Morand, S. Epidemiology of Leptospira Transmitted by Rodents in Southeast Asia. PLoS Negl. Trop. Dis. 2014, 8, e2902. [Google Scholar] [CrossRef] [PubMed]
- Della Rossa, P.; Tantrakarnapa, K.; Sutdan, D.; Kasetsinsombat, K.; Cosson, J.-F.; Supputamongkol, Y.; Chaisiri, K.; Tran, A.; Supputamongkol, S.; Binot, A.; et al. Environmental factors and public health policy associated with human and rodent infection by leptospirosis: A land cover-based study in Nan province, Thailand. Epidemiol. Infect. 2016, 144, 1550–1562. [Google Scholar] [CrossRef] [PubMed]
- Catry, T.; Li, Z.; Roux, E.; Herbreteau, V.; Gurgel, H.; Mangeas, M.; Seyler, F.; Dessay, N. Wetlands and Malaria in the Amazon: Guidelines for the Use of Synthetic Aperture Radar Remote-Sensing. Int. J. Environ. Res. Public Health 2018, 15, 468. [Google Scholar] [CrossRef] [PubMed]
- Suwanpakdee, S.; Kaewkungwal, J.; White, L.J.; Asensio, N.; Ratanakorn, P.; Singhasivanon, P.; Day, N.P.J.; Pan-Ngum, W. Spatio-temporal patterns of leptospirosis in Thailand: Is flooding a risk factor? Epidemiol. Infect. 2015, 143, 2106–2115. [Google Scholar] [CrossRef]
- Rakwatin, P.; Sansena, T.; Marjang, N.; Rungsipanich, A. Using multi-temporal remote-sensing data to estimate 2011 flood area and volume over Chao Phraya River basin, Thailand. Remote Sens. Lett. 2013, 4, 243–250. [Google Scholar] [CrossRef]
- Pinn, T.G. Leptospirosis in the Seychelles. Med. J. Aust. 1992, 156, 163–167. [Google Scholar] [CrossRef]
- Yersin, C.; Bovet, P.; Mérien, F.; Wong, T.; Panowsky, J.; Perolat, P. Human leptospirosis in the Seychelles (Indian Ocean): A population-based study. Am. J. Trop. Med. Hyg. 1998, 59, 933–940. [Google Scholar] [CrossRef]
- Russell, J.C.; Cole, N.C.; Zuël, N.; Rocamora, G. Introduced mammals on Western Indian Ocean islands. Glob. Ecol. Conserv. 2016, 6, 132–144. [Google Scholar] [CrossRef]
- Dietrich, M.; Gomard, Y.; Lagadec, E.; Ramasindrazana, B.; Le Minter, G.; Guernier, V.; Benlali, A.; Rocamora, G.; Markotter, W.; Goodman, S.M.; et al. Biogeography of Leptospira in wild animal communities inhabiting the insular ecosystem of the western Indian Ocean islands and neighboring Africa. Emerg. Microbes Infect. 2018, 7, 1–12. [Google Scholar] [CrossRef]
- Mills, J.N.; Childs, J.E.; Ksiazek, T.G.; Peters, C.J.; Velleca, W.M. Methods for Trapping and Sampling Small Mammals for Virologic Testing; U.S. Department of Health and Human Services—Centers for Disease Control and Prevention: Atlanta, GA, USA, 1995.
- Aplin, K.P.; Brown, P.R.; Jacob, J.; Krebs, C.J.; Singleton, G.R. Field Methods for Rodent Studies in Asia and the Indo-Pacific; Monograph No.100; ACIAR: Canberra, Australia, 2003; ISBN 1-86320-393-1. [Google Scholar]
- Herbreteau, V.; Jittapalapong, S.; Rerkamnuaychoke, W.; Chaval, Y.; Cosson, J.-F.; Morand, S. Protocols for Field and Laboratory Rodent Studies; Kasetsart University Press: Bangkok, Thailand, 2011. [Google Scholar]
- Révillion, C.; Lagadec, E.; Le Minter, G.; Dessay, N.; Guernier, V.; Sand, A.; Tortosa, P.; Dellagi, K.; Herbreteau, V. Utilisation de la très haute résolution spatiale pour la caractérisation des habitats de rongeurs, vecteurs de zoonoses à la Réunion. Rev. Fr. Photogramm. Télédétect. 2015, 209, 65–71. [Google Scholar]
- Révillion, C.; Attoumane, A.; Herbreteau, V. Homisland-IO: Homogeneous Land Use/Land Cover over the Small Islands of the Indian Ocean. Data 2019, 4, 82. [Google Scholar] [CrossRef]
- Anderson, J.R.; Hardy, E.E.; Roach, J.T.; Witmer, R.E. A Land Use and Land Cover Classification System for Use with Remote Sensor Data; Geological Survey Professional Paper Series; U.S. Government Publishing Office: Washington, DC, USA, 1976. [Google Scholar]
- Foody, G.M. Status of land cover classification accuracy assessment. Remote Sens. Environ. 2002, 80, 185–201. [Google Scholar] [CrossRef]
- Morand, S.; Bordes, F.; Blasdell, K.; Pilosof, S.; Cornu, J.-F.; Chaisiri, K.; Chaval, Y.; Cosson, J.-F.; Claude, J.; Feyfant, T.; et al. Assessing the distribution of disease-bearing rodents in human-modified tropical landscapes. J. Appl. Ecol. 2015, 52, 784–794. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Brotons, L.; Thuiller, W.; Araujo, M.B.; Hirzel, A.H. Presence-absence versus presence-only modelling methods for predicting bird habitat suitability. Ecography 2004, 27, 437–448. [Google Scholar] [CrossRef]
- Chesson, P. Mechanisms of Maintenance of Species Diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Russell, J.C.; Caut, S.; Anderson, S.H.; Lee, M. Invasive rat interactions and over-invasion on a coral atoll. Biol. Conserv. 2015, 185, 59–65. [Google Scholar] [CrossRef]
- Carver, S.; Mills, J.N.; Kuenzi, A.; Flietstra, T.; Douglass, R.J. Sampling frequency differentially influences interpretation of zoonotic pathogen and host dynamics: Sin Nombre virus and deer mice. Vector Borne Zoonotic Dis. 2010, 10, 575–583. [Google Scholar] [CrossRef]
- Panti-May, J.A.; Carvalho-Pereira, T.S.A.; Serrano, S.; Pedra, G.G.; Taylor, J.; Pertile, A.C.; Minter, A.; Airam, V.; Carvalho, M.; Júnior, N.N.; et al. A Two-Year Ecological Study of Norway Rats (Rattus norvegicus) in a Brazilian Urban Slum. PLoS ONE 2016, 11, e0152511. [Google Scholar] [CrossRef]
- Vadell, M.V.; Cavia, R.; Suárez, O.V. Abundance, age structure and reproductive patterns of Rattus norvegicus and Mus musculus in two areas of the city of Buenos Aires. Int. J. Pest. Manag. 2010, 56, 327–336. [Google Scholar] [CrossRef]
- Harper, G.A.; Bunbury, N. Invasive rats on tropical islands: Their population biology and impacts on native species. Glob. Ecol. Conserv. 2015, 3, 607–627. [Google Scholar] [CrossRef]
- Rocamora, G.; Henriette, E. Invasive Alien Species in Seychelles: Why and How to Eliminate Them? Identification and Management of Priority Species; Biotope, Muséum National d’Histoire Naturelle: Paris, France, 2015. [Google Scholar]
- Hill, M.J.; Vel, T.; Shah, N.J. The morphology, distribution and conservation implications of introduced rats, Rattus spp. in the granitic Seychelles. Afr. J. Ecol. 2003, 41, 179–186. [Google Scholar] [CrossRef]
- Herbreteau, V.; Bordes, F.; Jittapalapong, S.; Supputamongkol, Y.; Morand, S. Rodent-borne diseases in Thailand: Targeting rodent carriers and risky habitats. Infect. Ecol. Epidemiol. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Andersen-Ranberg, E.U.; Pipper, C.; Jensen, P.M. Global patterns of leptospira prevalence in vertebrate reservoir hosts. J. Wildl. Dis. 2016, 52, 468–477. [Google Scholar] [CrossRef]
- Krøjgaard, L.H.; Villumsen, S.; Markussen, M.D.K.; Jensen, J.S.; Leirs, H.; Heiberg, A.-C. High prevalence of Leptospira spp. in sewer rats (Rattus norvegicus). Epidemiol. Infect. 2009, 137, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.; Wunder, E.A.; De Oliveira, D.; Bisht, V.; Rodrigues, G.; Reis, M.G.; Ko, A.I.; Begon, M.; Childs, J.E. Patterns in Leptospira Shedding in Norway Rats (Rattus norvegicus) from Brazilian Slum Communities at High Risk of Disease Transmission. PLoS Negl. Trop. Dis. 2015, 9, e0003819. [Google Scholar] [CrossRef]
- Muñoz-Zanzi, C.; Mason, M.; Encina, C.; Gonzalez, M.; Berg, S. Household Characteristics Associated with Rodent Presence and Leptospira Infection in Rural and Urban Communities from Southern Chile. Am. J. Trop. Med. Hyg. 2014, 90, 497–506. [Google Scholar] [CrossRef]
- Van Der Geer, A.A.E. Changing invaders: Trends of gigantism in insular introduced rats. Environ. Conserv. 2018, 45, 203–211. [Google Scholar] [CrossRef]
- Berdoy, M.; Drickamer, L.C. Comparative social organization and life history of Rattus and Mus. In Rodent Societies: An Ecological and Evolutionary Perspective; University of Chicago Press: Chicago, IL, USA, 2007; pp. 380–392. [Google Scholar]
- Desvars-Larrive, A.; Baldi, M.; Walter, T.; Zink, R.; Walzer, C. Brown rats (Rattus norvegicus) in urban ecosystems: Are the constraints related to fieldwork a limit to their study? Urban Ecosyst. 2018, 21, 951–964. [Google Scholar] [CrossRef]
- Guiry, E.; Buckley, M. Urban rats have less variable, higher protein diets. Proc. R. Soc. B Biol. Sci. 2018, 285, 20181441. [Google Scholar] [CrossRef]
- Aplin, K.P.; Suzuki, H.; Chinen, A.A.; Chesser, R.T.; ten Have, J.; Donnellan, S.C.; Austin, J.; Frost, A.; Gonzalez, J.P.; Herbreteau, V.; et al. Multiple Geographic Origins of Commensalism and Complex Dispersal History of Black Rats. PLoS ONE 2011, 6, e26357. [Google Scholar] [CrossRef]
- Feng, A.Y.T.; Himsworth, C.G. The secret life of the city rat: A review of the ecology of urban Norway and black rats (Rattus norvegicus and Rattus rattus). Urban Ecosyst. 2014, 17, 149–162. [Google Scholar] [CrossRef]
- Byers, K.A.; Lee, M.J.; Patrick, D.M.; Himsworth, C.G. Rats About Town: A Systematic Review of Rat Movement in Urban Ecosystems. Front. Ecol. Evol. 2019, 7, 13. [Google Scholar] [CrossRef]
- Cheke, A. The timing of arrival of humans and their commensal animals on Western Indian Ocean oceanic islands. Phelsuma 2010, 18, 38–69. [Google Scholar]
- Bordes, F.; Morand, S.; Pilosof, S.; Claude, J.; Krasnov, B.R.; Cosson, J.-F.; Chaval, Y.; Ribas, A.; Chaisiri, K.; Blasdell, K.; et al. Habitat fragmentation alters the properties of a host–parasite network: Rodents and their helminths in South-East Asia. J. Anim. Ecol. 2015, 84, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, N.; Ng, C.F.S.; Kim, Y.; Suzuki, M.; Saito, N.; Ariyoshi, K.; Salva, E.P.; Dimaano, E.M.; Villarama, J.B.; Go, W.S.; et al. The non-linear and lagged short-term relationship between rainfall and leptospirosis and the intermediate role of floods in the Philippines. PLoS Negl. Trop. Dis. 2018, 12, e0006331. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, S.; Herbreteau, V.; Blasdell, K.; Chaval, Y.; Buchy, P.; Guillard, B.; Morand, S. Leptospira and Rodents in Cambodia: Environmental Determinants of Infection. Am. J. Trop. Med. Hyg. 2012, 86, 1032–1038. [Google Scholar] [CrossRef]
- Cavia, R.; Cueto, G.R.; Suárez, O.V. Changes in rodent communities according to the landscape structure in an urban ecosystem. Landsc. Urban Plan. 2009, 90, 11–19. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
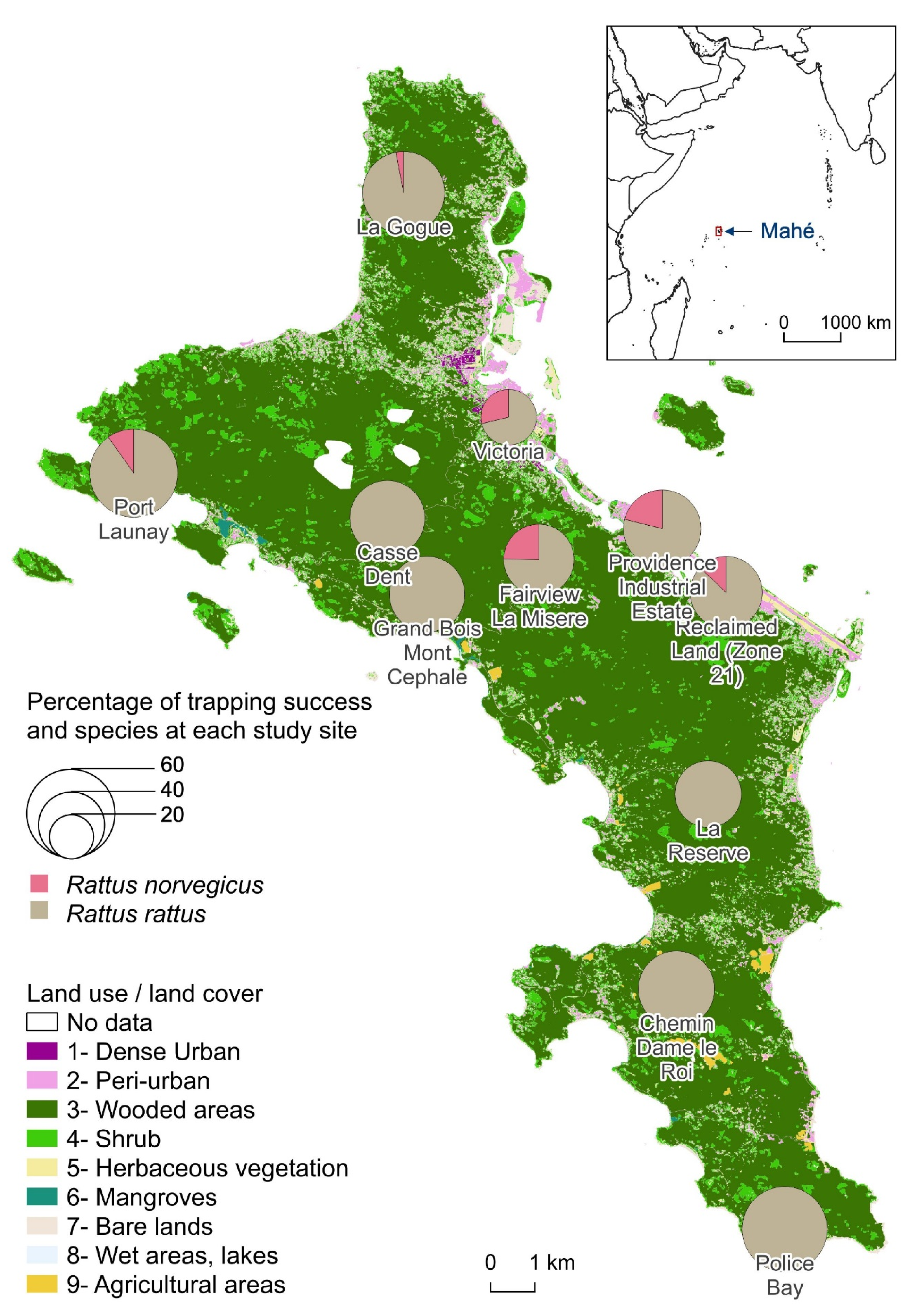
| Site | Trap-Nights | Environment | Trapping Success | Leptospira Detection (RT-qPCR) Positive/Total Sampled (% of Positive) | |||
|---|---|---|---|---|---|---|---|
| Rattus rattus | Rattus norvegicus | Rattus rattus | Rattus norvegicus | All | |||
| Casse Dent | 149 | N | 46.9 | 0.0 | 2/65 (3.1) | 0/2 (0.0) | 2/67 (3.0) |
| Chemin Dame le Roi | 179 | R | 47.9 | 0.0 | 2/75 (2.7) | - | 2/75 (2.7) |
| Fairview La Misère | 250 | P | 31.2 | 10.3 | 8/69 (11.6) | 8/15 (53.3) | 16/84 (19.0) |
| Grand Bois Mont Cephale | 140 | N | 47.2 | 0.0 | 3/58 (5.2) | - | 3/58 (5.2) |
| La Gogue | 133 | R | 54.0 | 1.7 | 1/66 (1.5) | 0/1 (0.0) | 1/67 (1.5) |
| La Reserve | 172 | N | 36.6 | 0.0 | 1/60 (1.7) | - | 1/60 (1.7) |
| Police Bay | 168 | N | 60.0 | 0.0 | 0/98 (0.0) | - | 0/98 (0.0) |
| Port Launay | 148 | R | 57.9 | 6.3 | 2/78 (2.6) | 3/5 (60.0) | 5/83 (6.0) |
| Providence Industrial Estate | 33 | U | 39.3 | 10.5 | 0/6 (0.0) | 2/2 100.0 | 2/8 (25.0) |
| Reclaimed Land (Zone 21) | 142 | P | 37.3 | 5.6 | 7/50 (14.0) | 5/5 (100.0) | 12/55 (21.8) |
| Victoria | 342 | U | 18.4 | 7.4 | 4/60 (6.7) | 9/20 (45.0) | 13/80 (16.3) |
| Total | 1859 | - | 43.3 | 3.8 | 30/685 (4.4) | 27/50 (54.0) | 57/735 (7.8) |
| Land Use/Land Cover Classes | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | Total | Producer Accuracy |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1—Dense Urban | 39 | 2 | 3 | 44 | 0.89 | ||||||
| 2—Peri-urban | 56 | 3 | 2 | 1 | 4 | 66 | 0.85 | ||||
| 3—Wooded areas | 79 | 2 | 81 | 0.98 | |||||||
| 4—Shrub | 4 | 28 | 1 | 33 | 0.85 | ||||||
| 5—Herbaceous vegetation | 1 | 2 | 1 | 25 | 5 | 34 | 0.74 | ||||
| 6—Mangroves | 2 | 2 | 26 | 32 | 0.81 | ||||||
| 7—Bare lands | 2 | 1 | 33 | 3 | 39 | 0.85 | |||||
| 8—Wet areas, lakes | 1 | 29 | 30 | 0.97 | |||||||
| 9—Agricultural areas | 1 | 1 | 31 | 33 | 0.94 | ||||||
| Total | 39 | 58 | 90 | 35 | 29 | 30 | 45 | 33 | 31 | 390 | |
| User accuracy | 1 | 0.97 | 0.88 | 0.80 | 0.86 | 0.87 | 0.73 | 0.88 | 1 |
| Model | Variables | Estimate | Std. Error | z Value | p-Value |
|---|---|---|---|---|---|
| Model 1 (Excluding variable Species) | (Intercept) | −2.943 | 0.238 | −12.382 | <2 × 10−16 |
| AIC = 272.74 | Edge density 100 | 0.591 | 0.160 | 3.683 | <3 × 10−4 |
| AUCROC = 77.68% | 20-year average rainfall | 0.643 | 0.283 | 2.272 | 0.023 |
| AUCPR = 16.08% | 3-month cumulated rainfall | 30.260 | 0.159 | 1.629 | 0.103 |
| Accuracy = 92.35% F1 = NA | Elevation | 0.219 | 0.144 | 1.526 | 0.127 |
| Model 2 (Excluding variable Species) | (Intercept) | −2.925 | 0.233 | −12.548 | <2 × 10−16 |
| AIC = 272.14 | Edge density 100 | 0.499 | 0.177 | 2.81 | 0.005 |
| AUCROC = 79.42% | Distances to dense urban areas | −0.738 | 0.317 | −2.332 | 0.020 |
| AUCPR = 15.55% | 3-month cumulated rainfall | 0.247 | 0.159 | 1.555 | 0.120 |
| Accuracy = 92.35% F1 = NA | |||||
| Model 3 (Including variable Species) | (Intercept) | −3.034 | 0.235 | −12.907 | <2 × 10−16 |
| AIC = 237.9 | Species RR | −0.648 | 0.107 | −6.044 | 1.5 × 10−9 |
| AUCROC = 88.12% | Edge density 100 | 0.380 | 0.195 | 1.947 | 0.052 |
| AUCPR = 47.21% Accuracy = 93.99% | Distances to dense urban areas | −0.395 | 0.305 | −1.294 | 0.195 |
| F1 = 42.11% | 3-month cumulated rainfall | 0.21 | 0.17 | 1.251 | 0.211 |
| Model 4 (Including variable Species) | (Intercept) | −3.1093 | 0.254 | −12.246 | <2 × 10−16 |
| AIC = 236.65 AUCROC = 84.87% | Species RR | −0.657 | 0.105 | −6.242 | 4.3 × 10−10 |
| AUCPR = 43.07% | Edge density 100 | 0.3835 | 0.180 | 2.132 | 0.033 |
| Accuracy = 92.35% | 20-year average rainfall | 0.5217 | 0.304 | 1.718 | 0.086 |
| F1 = 30% | 3-month cumulated rainfall | 0.2302 | 0.176 | 1.309 | 0.190 |
| Elevation | 0.2125 | 0.158 | 1.348 | 0.178 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biscornet, L.; Révillion, C.; Jégo, S.; Lagadec, E.; Gomard, Y.; Le Minter, G.; Rocamora, G.; Guernier-Cambert, V.; Mélade, J.; Dellagi, K.; et al. Predicting the Presence of Leptospires in Rodents from Environmental Indicators Opens Up Opportunities for Environmental Monitoring of Human Leptospirosis. Remote Sens. 2021, 13, 325. https://doi.org/10.3390/rs13020325
Biscornet L, Révillion C, Jégo S, Lagadec E, Gomard Y, Le Minter G, Rocamora G, Guernier-Cambert V, Mélade J, Dellagi K, et al. Predicting the Presence of Leptospires in Rodents from Environmental Indicators Opens Up Opportunities for Environmental Monitoring of Human Leptospirosis. Remote Sensing. 2021; 13(2):325. https://doi.org/10.3390/rs13020325
Chicago/Turabian StyleBiscornet, Leon, Christophe Révillion, Sylvaine Jégo, Erwan Lagadec, Yann Gomard, Gildas Le Minter, Gérard Rocamora, Vanina Guernier-Cambert, Julien Mélade, Koussay Dellagi, and et al. 2021. "Predicting the Presence of Leptospires in Rodents from Environmental Indicators Opens Up Opportunities for Environmental Monitoring of Human Leptospirosis" Remote Sensing 13, no. 2: 325. https://doi.org/10.3390/rs13020325
APA StyleBiscornet, L., Révillion, C., Jégo, S., Lagadec, E., Gomard, Y., Le Minter, G., Rocamora, G., Guernier-Cambert, V., Mélade, J., Dellagi, K., Tortosa, P., & Herbreteau, V. (2021). Predicting the Presence of Leptospires in Rodents from Environmental Indicators Opens Up Opportunities for Environmental Monitoring of Human Leptospirosis. Remote Sensing, 13(2), 325. https://doi.org/10.3390/rs13020325