
Evaluating the Impact of Grazing Cessation and Reintroduction in Mixed Prairie Using Raster Time Series Analysis of Landsat Data


Abstract
:
1. Introduction
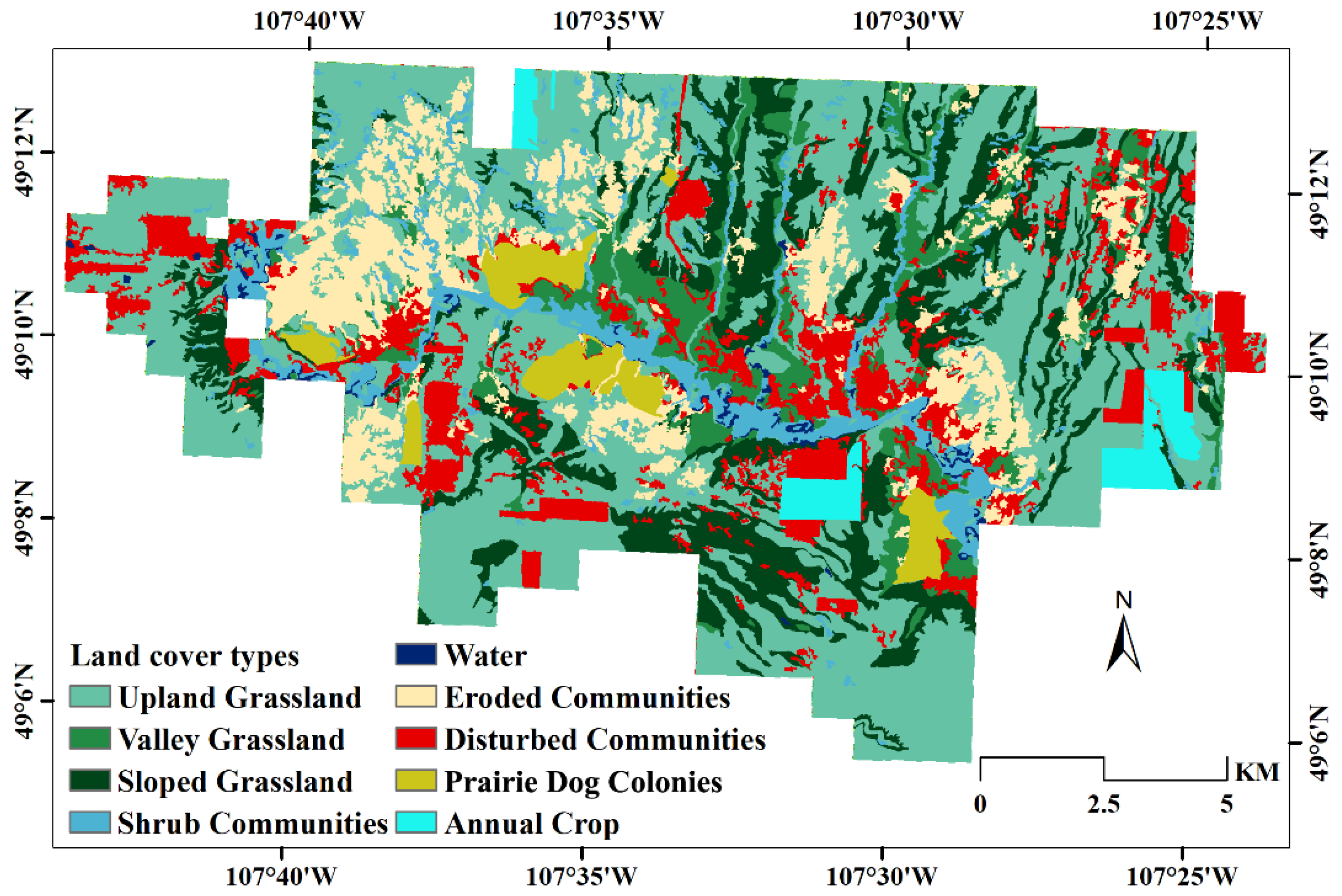
2. Study Area
3. Materials and Methods
3.1. Datasets
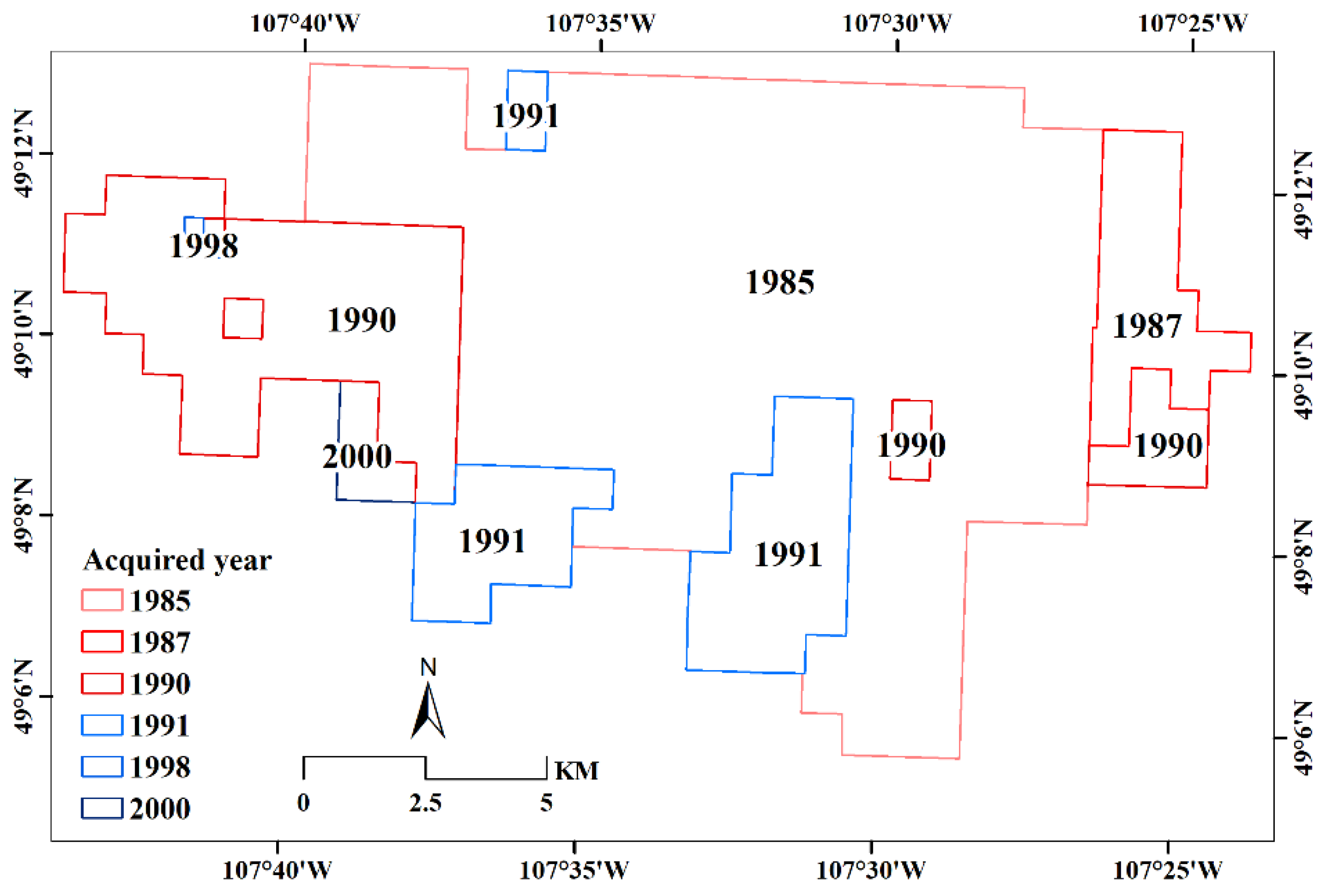
3.1.1. Landsat Images for Temporal Analysis
3.1.2. Sentinel-2 Images and DEM Image for Classification
3.2. Methodology
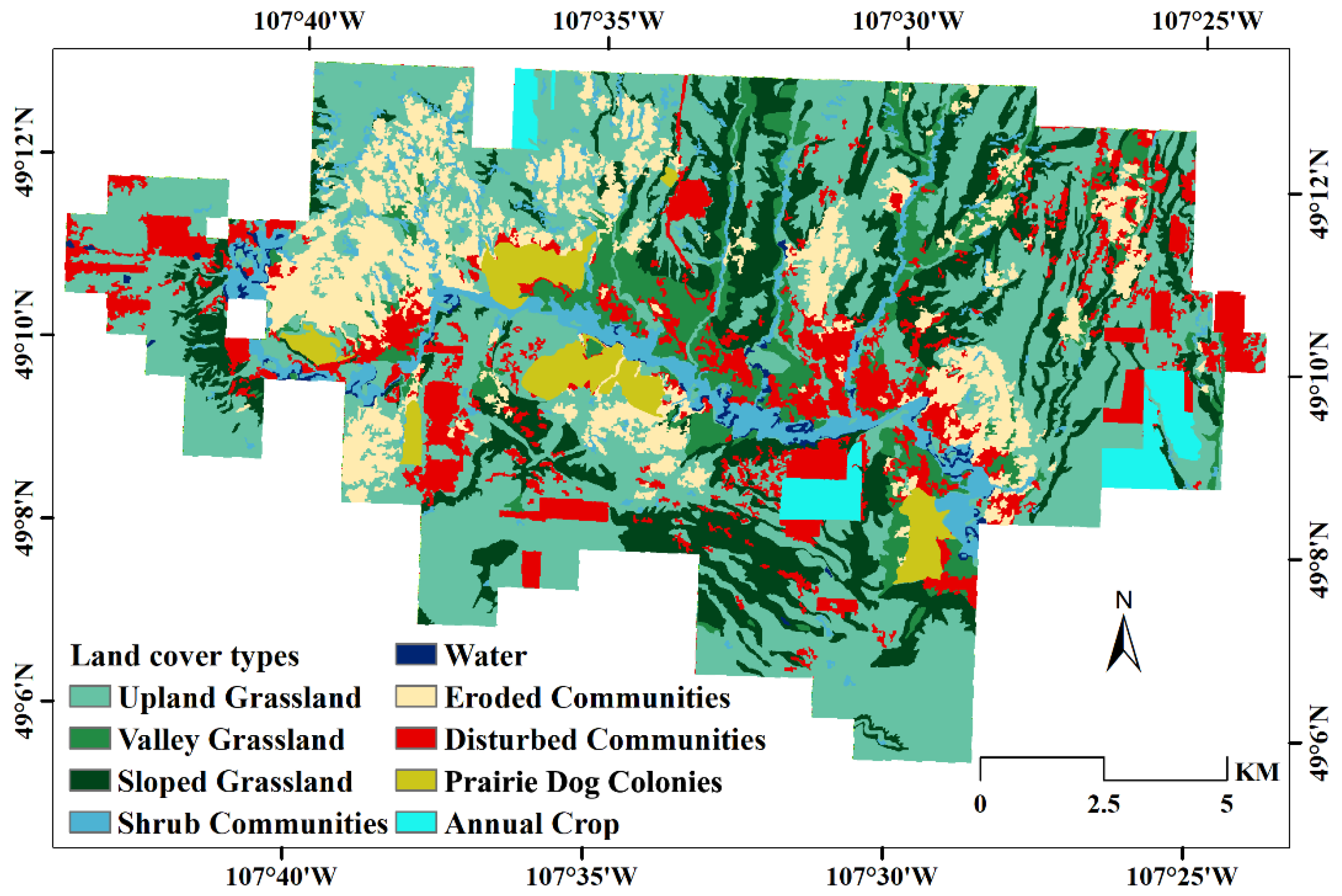
3.2.1. Classification of Vegetation Communities
3.2.2. Links between Surface Reflectance of Landsat Images and Grassland Biophysical Parameters
3.2.3. Formation of Time Series Dataset by “Zoo Objects” and Missing Value Interpolation
3.2.4. Pixel Based Time Series Analysis
3.2.5. Segmented Linear Regression
4. Results
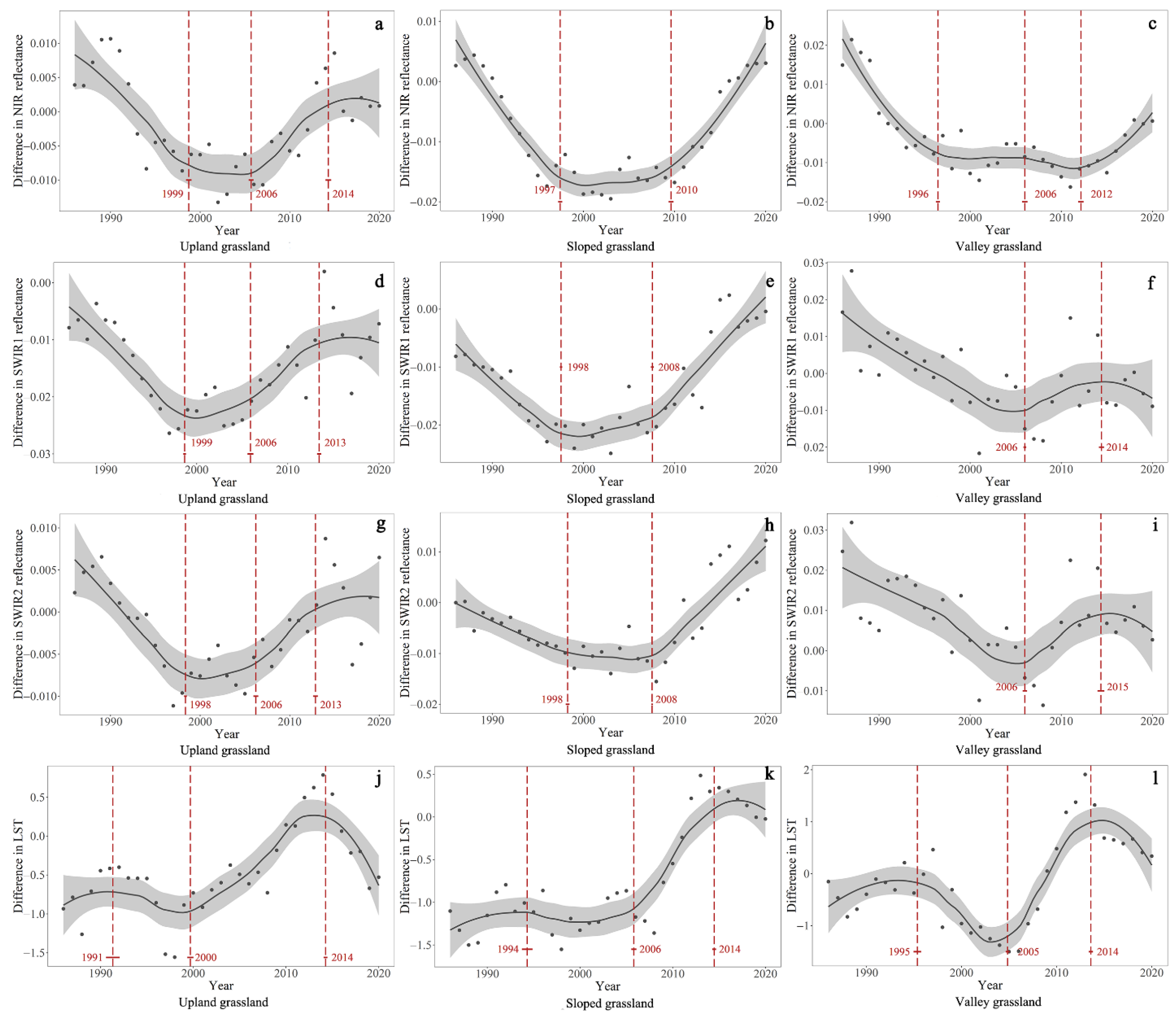
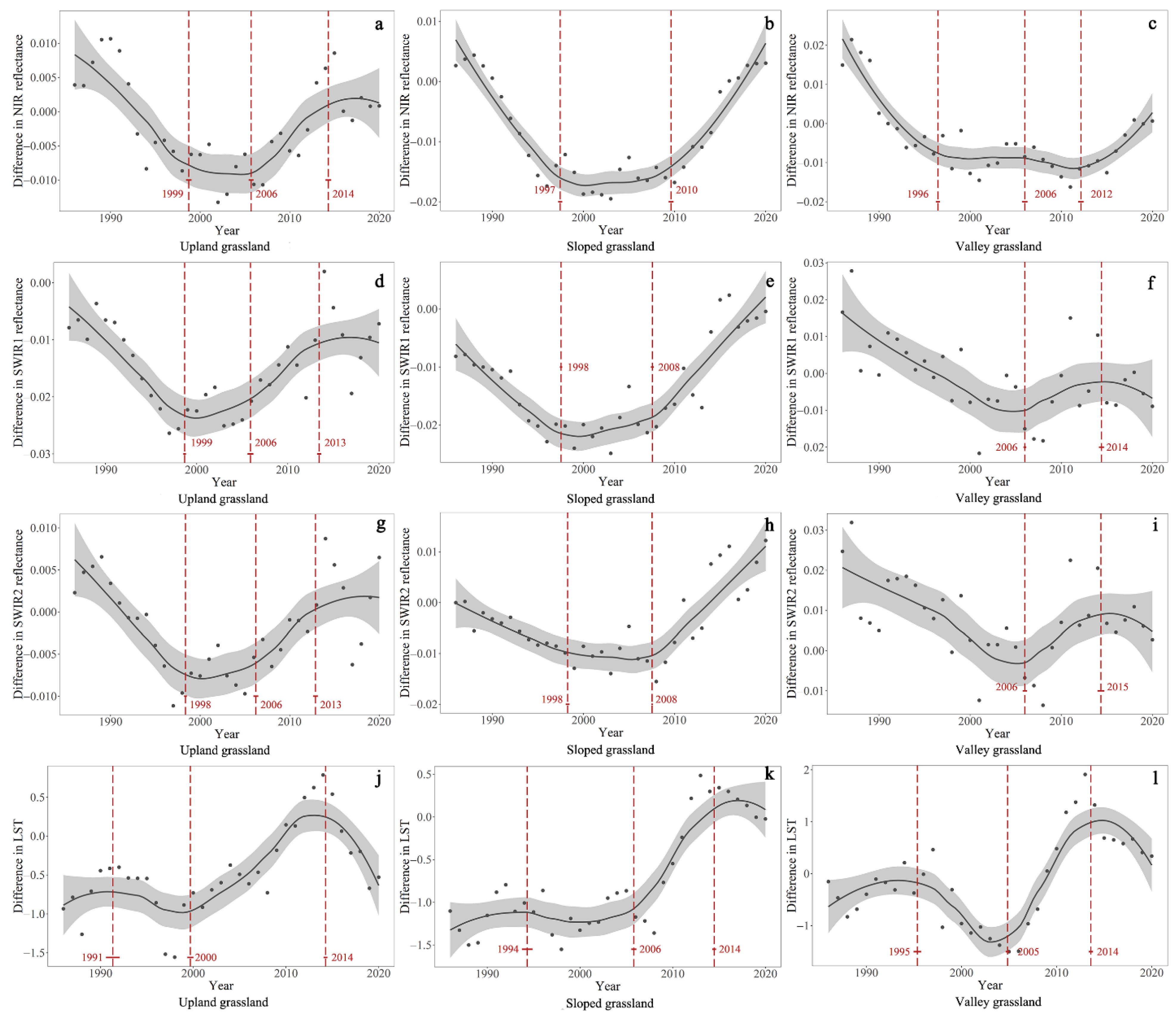
4.1. Temporal Trends of the Difference in NIR, SWIR1, SWIR2, and LST between GNP and Surrounding Pastures
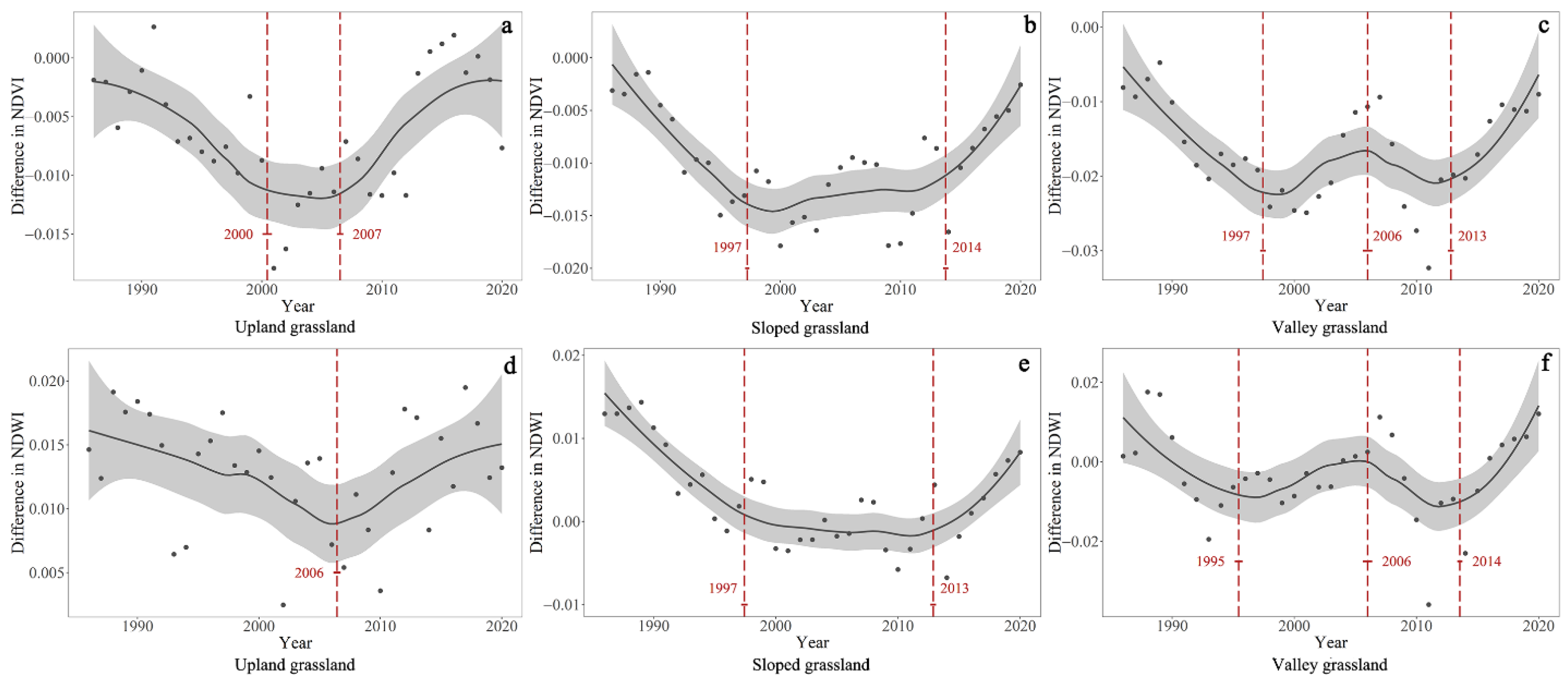
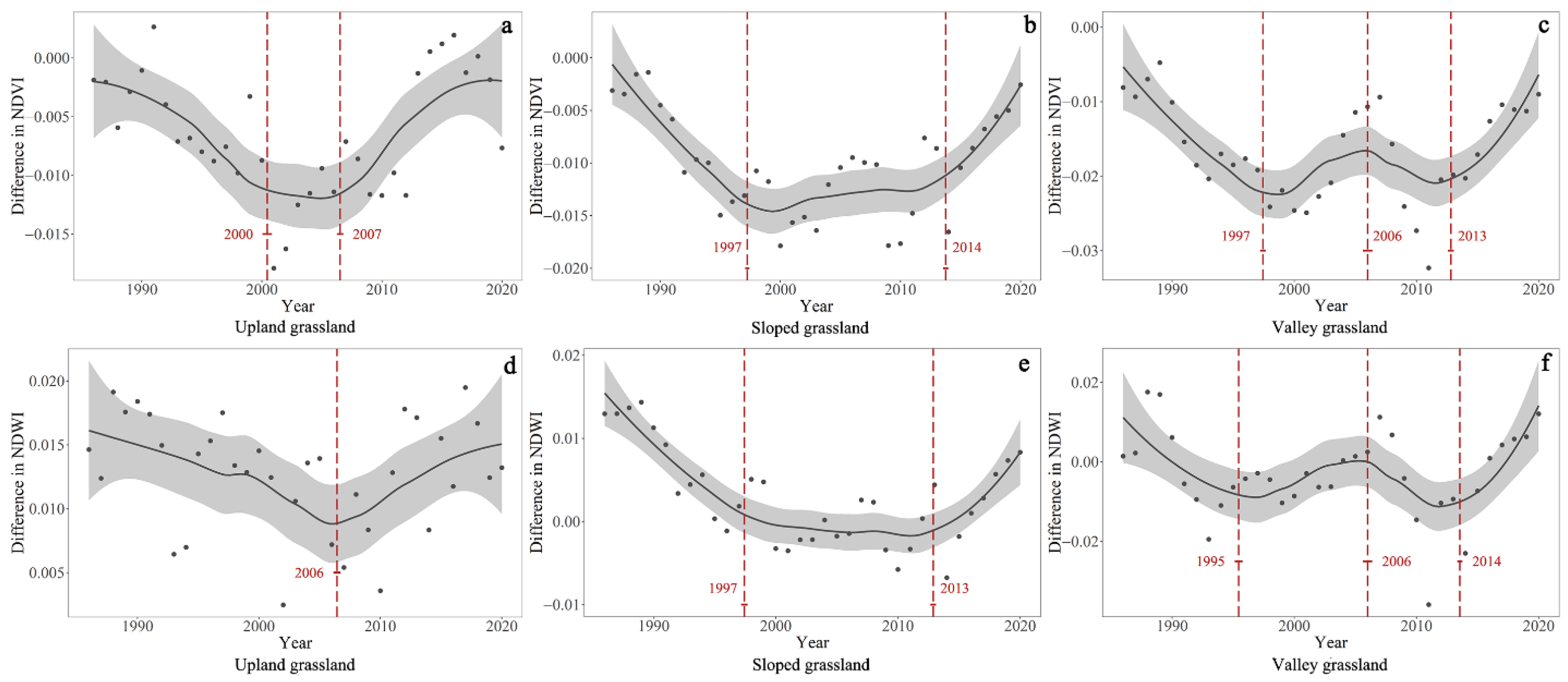
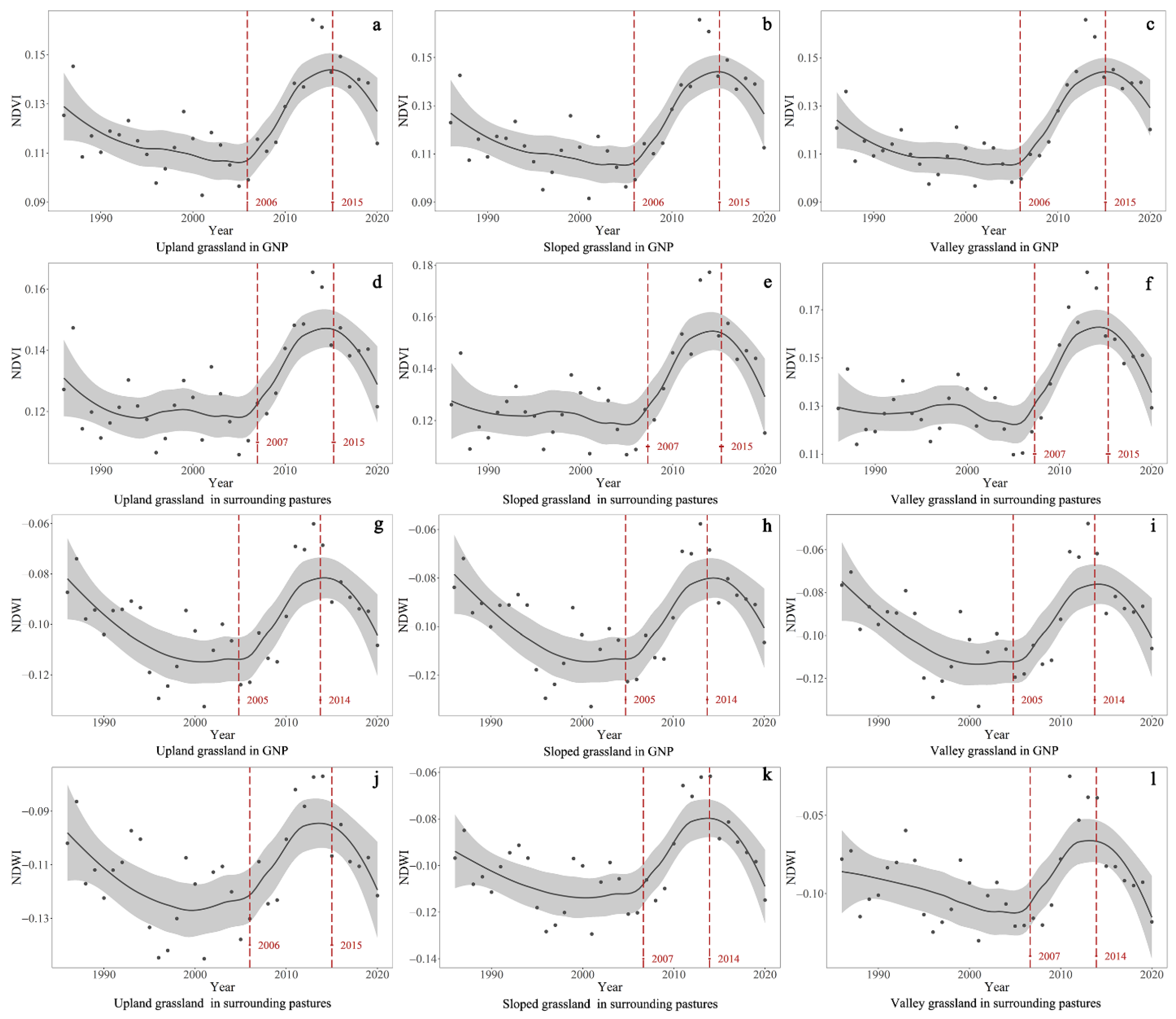
4.2. Temporal Trends of the Differences in NDVI and NDWI between GNP and Surrounding Pastures
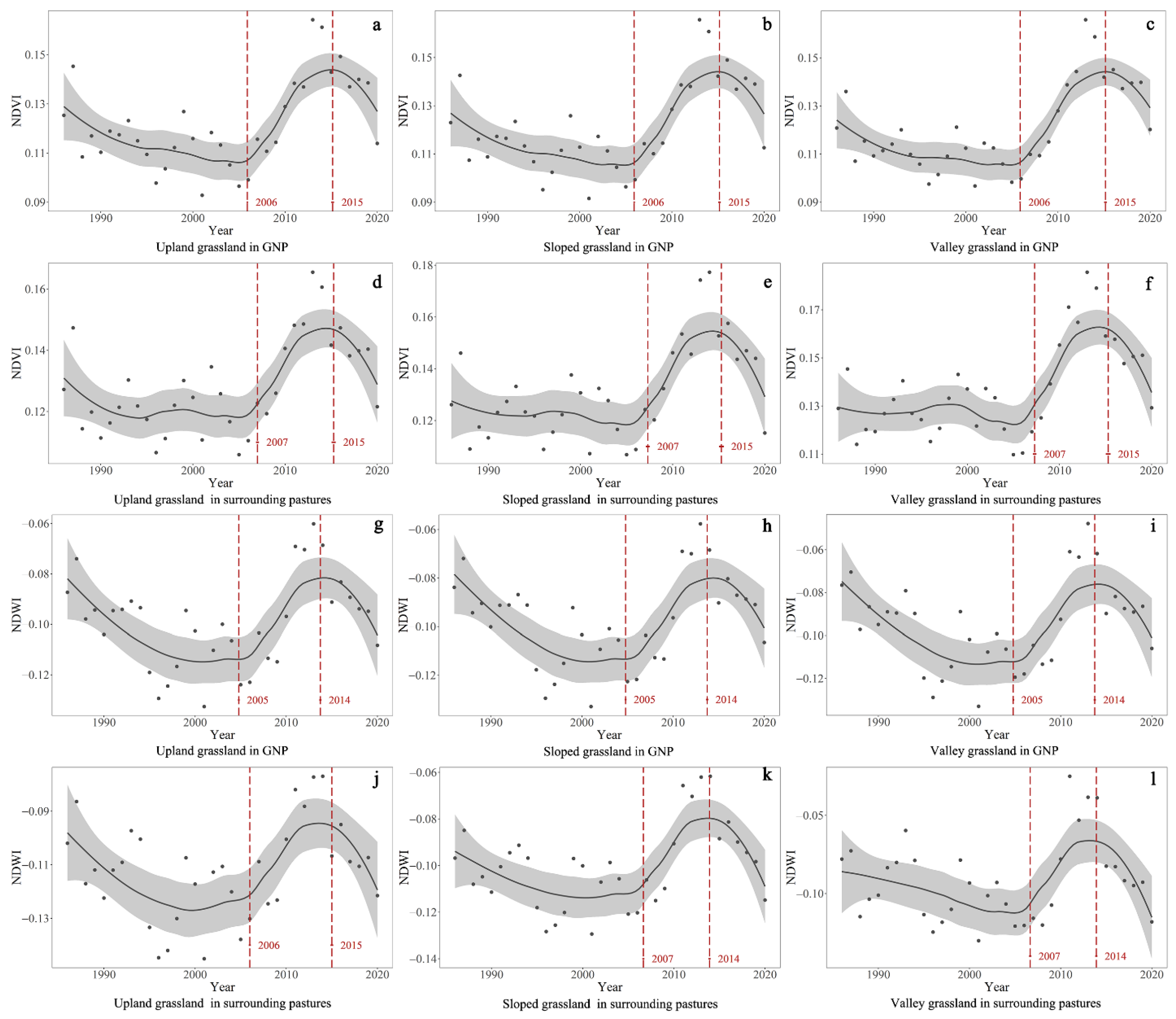
5. Discussion
5.1. Monitoring Prairie Management Effects Using a Landsat Imagery Raster Time Series Analysis
5.2. The Relationship between the Difference in Biophysical Parameters and Multispectral Reflectance
5.3. The Impact of Grazing Cessation on Three Native Vegetation Communities
5.4. The Impact of Grazing Reintroduction on Three Native Vegetation Communities
5.5. The Influences of Climate Change on Three Native Vegetation Communities
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Alemu, A.W.; Kröbel, R.; McConkey, B.G.; Iwaasa, A.D. Effect of Increasing Species Diversity and Grazing Management on Pasture Productivity, Animal Performance, and Soil Carbon Sequestration of Re-Established Pasture in Canadian Prairie. Animals 2019, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Milton, S.J.; Dean, W.J.; Ellis, R.P. Rangeland health assessment: A practical guide for ranchers in arid Karoo shrublands. J. Arid. Environ. 1998, 39, 253–265. [Google Scholar] [CrossRef]
- Follett, R.; Stewart, C.; Bradford, J.; Pruessner, E.; Sims, P.L.; Vigil, M. Long-term pasture management impacts on eolian sand soils in the southern mixed-grass prairie. Quat. Int. 2020, 565, 84–93. [Google Scholar] [CrossRef]
- Bajgain, R.; Xiao, X.; Basara, J.; Doughty, R.; Wu, X.; Wagle, P.; Zhou, Y.; Gowda, P.; Steiner, J. Differential responses of native and managed prairie pastures to environmental variability and management practices. Agric. For. Meteorol. 2020, 294, 108137. [Google Scholar] [CrossRef]
- Ruggiero, N.; Kral, G.L. Prescribed cattle grazing as a tool for native prairie management: Lessons from the Tualatin River basin, Oregon. J. Soil Water Conserv. 2018, 73, 74A–78A. [Google Scholar] [CrossRef] [Green Version]
- Dunwiddie, P.W.; Bakker, J.D. The Future of Restoration and Management of Prairie-Oak Ecosystems in the Pacific Northwest. Northwest Sci. 2011, 85, 83–92. [Google Scholar] [CrossRef]
- Dennehy, C.; Alverson, E.R.; Anderson, H.E.; Clements, D.R.; Gilbert, R.; Kaye, T.N. Management Strategies for Invasive Plants in Pacific Northwest Prairies, Savannas, and Oak Woodlands. Northwest Sci. 2011, 85, 329–351. [Google Scholar] [CrossRef]
- Ahlering, M.; Carlson, D.; Vacek, S.; Jacobi, S.; Hunt, V.; Stanton, J.C.; Knutson, M.G.; Lonsdorf, E. Cooperatively improving tallgrass prairie with adaptive management. Ecosphere 2020, 11, 11. [Google Scholar] [CrossRef]
- Giglio, R.M.; Rocke, T.E.; Osorio, J.E.; Latch, E.K. Characterizing patterns of genomic variation in the threatened Utah prairie dog: Implications for conservation and management. Evol. Appl. 2021, 14, 1036–1051. [Google Scholar] [CrossRef] [PubMed]
- Gibon, A.; Ladet, S.; Balent, G. A socioecological assessment of the relationships between grassland management practices and landscape-level ecosystem services in Pyrenees National Park, France. Fourrages 2015, 224, 305–319. [Google Scholar]
- Vold, S.T.; Berkeley, L.I.; McNew, L.B. Effects of Livestock Grazing Management on Grassland Birds in a Northern Mixed-Grass Prairie Ecosystem. Rangel. Ecol. Manag. 2019, 72, 933–945. [Google Scholar] [CrossRef]
- Hunt, V.M.; Jacobi, S.K.; Gannon, J.J.; Zorn, J.E.; Moore, C.T.; Lonsdorf, E.V. A Decision Support Tool for Adaptive Management of Native Prairie Ecosystems. Interfaces 2016, 46, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Liu, S.; Tan, Z.; Sohl, T.L.; Wu, Y. Simulating the effects of management practices on cropland soil organic carbon changes in the Temperate Prairies Ecoregion of the United States from 1980 to 2012. Ecol. Model. 2017, 365, 68–79. [Google Scholar] [CrossRef]
- Abella, S.R.; Menard, K.S.; Schetter, T.A.; Sprow, L.A.; Jaeger, J.F. Rapid and transient changes during 20 years of restoration management in savanna-woodland-prairie habitats threatened by woody plant encroachment. Plant Ecol. 2020, 221, 1201–1217. [Google Scholar] [CrossRef]
- Teague, W.; Dowhower, S.; Baker, S.; Haile, N.; DeLaune, P.; Conover, D. Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agric. Ecosyst. Environ. 2011, 141, 310–322. [Google Scholar] [CrossRef]
- Larson, D.; Ahlering, M.; Drobney, P.; Esser, R.; Larson, J.; Viste-Sparkman, K. Developing a Framework for Evaluating Tallgrass Prairie Reconstruction Methods and Management. Ecol. Restor. 2018, 36, 6–18. [Google Scholar] [CrossRef]
- Matthews, J.W.; Molano-Flores, B.; Ellis, J.; Marcum, P.B.; Handel, W.; Zylka, J.; Phillippe, L.R. Impacts of management and antecedent site condition on restoration outcomes in a sand prairie. Restor. Ecol. 2017, 25, 972–981. [Google Scholar] [CrossRef]
- Stanley, A.G.; Dunwiddie, P.; Kaye, T.N. Restoring Invaded Pacific Northwest Prairies: Management Recommendations from a Region-Wide Experiment. Northwest Sci. 2011, 85, 233–246. [Google Scholar] [CrossRef]
- Katsalirou, E.; Deng, S.; Gerakis, A.; Nofziger, D.L. Long-term management effects on soil P, microbial biomass P, and phosphatase activities in prairie soils. Eur. J. Soil Biol. 2016, 76, 61–69. [Google Scholar] [CrossRef]
- Morton, L.W.; Regen, E.; Engle, D.M.; Miller, J.R.; Harr, R.N. Perceptions of Landowners Concerning Conservation, Grazing, Fire, and Eastern Redcedar Management in Tallgrass Prairie. Rangel. Ecol. Manag. 2010, 63, 645–654. [Google Scholar] [CrossRef]
- Esparrago, J.; Kricsfalusy, V. Traditional grassland management and surrounding land use drive the abundance of a prairie plant species in urban areas. Landsc. Urban Plan. 2015, 142, 1–6. [Google Scholar] [CrossRef]
- Larson, D.L.; Hernández, D.L.; Larson, J.L.; Leone, J.B.; Pennarola, N. Management of remnant tallgrass prairie by grazing or fire: Effects on plant communities and soil properties. Ecosphere 2020, 11, 11. [Google Scholar] [CrossRef]
- Grman, E.; Bassett, T.; Brudvig, L.A. Confronting contingency in restoration: Management and site history determine outcomes of assembling prairies, but site characteristics and landscape context have little effect. J. Appl. Ecol. 2013, 50, 1234–1243. [Google Scholar] [CrossRef]
- Wagle, P.; Gowda, P.H.; Northup, B.K.; Starks, P.J.; Neel, J.P.S. Response of Tallgrass Prairie to Management in the U.S. Southern Great Plains: Site Descriptions, Management Practices, and Eddy Covariance Instrumentation for a Long-Term Experiment. Remote Sens. 2019, 11, 1988. [Google Scholar] [CrossRef] [Green Version]
- Guiden, P.W.; Barber, N.A.; Blackburn, R.; Farrell, A.; Fliginger, J.; Hosler, S.C.; King, R.B.; Nelson, M.; Rowland, E.G.; Savage, K.; et al. Effects of management outweigh effects of plant diversity on restored animal communities in tallgrass prairies. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Dong, J.; Xiao, X.; Wagle, P.; Zhang, G.; Zhou, Y.; Jin, C.; Torn, M.S.; Meyers, T.P.; Suyker, A.E.; Wang, J.; et al. Comparison of four EVI-based models for estimating gross primary production of maize and soybean croplands and tallgrass prairie under severe drought. Remote Sens. Environ. 2015, 162, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Wagle, P.; Xiao, X.; Torn, M.; Cook, D.R.; Matamala, R.; Fischer, M.L.; Jin, C.; Dong, J.; Biradar, C. Sensitivity of vegetation indices and gross primary production of tallgrass prairie to severe drought. Remote Sens. Environ. 2014, 152, 1–14. [Google Scholar] [CrossRef]
- Wehlage, D.C.; Gamon, J.A.; Thayer, D.; Hildebrand, D.V. Interannual Variability in Dry Mixed-Grass Prairie Yield: A Comparison of MODIS, SPOT, and Field Measurements. Remote Sens. 2016, 8, 872. [Google Scholar] [CrossRef] [Green Version]
- Dong, T.; Liu, J.; Shang, J.; Qian, B.; Huffman, T.; Zhang, Y.; Champagne, C.; Daneshfar, B. Assessing the Impact of Climate Variability on Cropland Productivity in the Canadian Prairies Using Time Series MODIS FAPAR. Remote Sens. 2016, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Brennan, J.R.; Johnson, P.S.; Hanan, N.P. Comparing stability in random forest models to map Northern Great Plains plant communities in pastures occupied by prairie dogs using Pleiades imagery. Biogeosciences 2020, 17, 1281–1292. [Google Scholar] [CrossRef] [Green Version]
- Cui, T.; Martz, L.; Zhao, L.; Guo, X. Investigating the impact of the temporal resolution of MODIS data on measured phenology in the prairie grasslands. GISci. Remote Sens. 2020, 57, 395–410. [Google Scholar] [CrossRef]
- Wagle, P.; Gowda, P.H.; Neel, J.P.; Northup, B.K.; Zhou, Y. Integrating eddy fluxes and remote sensing products in a rotational grazing native tallgrass prairie pasture. Sci. Total Environ. 2020, 712, 136407. [Google Scholar] [CrossRef]
- Rigge, M.; Smart, A.; Wylie, B.; Gilmanov, T.; Johnson, P. Linking Phenology and Biomass Productivity in South Dakota Mixed-Grass Prairie. Rangel. Ecol. Manag. 2013, 66, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.M.; Hill, M.J.; Zhang, Y. Estimating Ground Cover in the Mixed Prairie Grassland of Southern Alberta Using Vegetation Indices Related to Physiological Function. Can. J. Remote Sens. 2015, 41, 51–66. [Google Scholar] [CrossRef]
- Yang, X.; Guo, X. Investigating vegetation biophysical and spectral parameters for detecting light to moderate grazing effects: A case study in mixed grass prairie. Open Geosci. 2011, 3, 336–348. [Google Scholar] [CrossRef]
- Przeszlowska, A.; Trlica, M.J.; Weltz, M.A. Near-Ground Remote Sensing of Green Area Index on the Shortgrass Prairie. Rangel. Ecol. Manag. 2006, 59, 422–430. [Google Scholar] [CrossRef]
- Yang, X.; Kovach, E.; Guo, X. Biophysical and spectral responses to various burn treatments in the northern mixed-grass prairie. Can. J. Remote Sens. 2013, 39, 175–184. [Google Scholar] [CrossRef]
- Phillips, R.L.; Ngugi, M.K.; Hendrickson, J.; Smith, A.; West, M. Mixed-Grass Prairie Canopy Structure and Spectral Reflectance Vary with Topographic Position. Environ. Manag. 2012, 50, 914–928. [Google Scholar] [CrossRef] [Green Version]
- Kaskie, K.D.; Wimberly, M.C.; Bauman, P.J. Rapid assessment of juniper distribution in prairie landscapes of the northern Great Plains. Int. J. Appl. Earth Obs. Geoinf. 2019, 83, 83. [Google Scholar] [CrossRef]
- Lindsay, E.J.; King, D.; Davidson, A.M.; Daneshfar, B. Canadian Prairie Rangeland and Seeded Forage Classification Using Multiseason Landsat 8 and Summer RADARSAT-2. Rangel. Ecol. Manag. 2019, 72, 92–102. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, X.; Qin, Y.; Doughty, R.B.; Dong, J.; Zou, Z. Characterizing the encroachment of juniper forests into sub-humid and semi-arid prairies from 1984 to 2010 using PALSAR and Landsat data. Remote Sens. Environ. 2018, 205, 166–179. [Google Scholar] [CrossRef]
- Finger, D.J.; McPherson, M.L.; Houskeeper, H.F.; Kudela, R.M. Mapping bull kelp canopy in northern California using Landsat to enable long-term monitoring. Remote Sens. Environ. 2021, 254, 112243. [Google Scholar] [CrossRef]
- Fraser, R.H.; Olthof, I.; Carrière, M.; Deschamps, A.; Pouliot, D. Detecting long-term changes to vegetation in northern Canada using the Landsat satellite image archive. Environ. Res. Lett. 2011, 6, 045502. [Google Scholar] [CrossRef] [Green Version]
- Mandanici, E.; Bitelli, G. Multi-Image and Multi-Sensor Change Detection for Long-Term Monitoring of Arid Environments with Landsat Series. Remote Sens. 2015, 7, 14019–14038. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Feng, D.; Yu, L.; Cheng, Y.; Zhang, M.; Liu, X.; Xu, Y.; Fang, L.; Zhu, Z.; Gong, P. Long-Term Land Cover Dynamics (1986–2016) of Northeast China Derived from a Multi-Temporal Landsat Archive. Remote Sens. 2019, 11, 599. [Google Scholar] [CrossRef] [Green Version]
- Sulla-Menashe, D.; Friedl, M.A.; Woodcock, C.E. Sources of bias and variability in long-term Landsat time series over Canadian boreal forests. Remote Sens. Environ. 2016, 177, 206–219. [Google Scholar] [CrossRef]
- Lopes, C.L.; Mendes, R.; Caçador, I.; Dias, J.M. Evaluation of long-term estuarine vegetation changes through Landsat imagery. Sci. Total Environ. 2019, 653, 512–522. [Google Scholar] [CrossRef]
- Graf, W.; Kleinn, C.; Schall, P.; Nauss, T.; Detsch, F.; Magdon, P. Analyzing the relationship between historic canopy dynamics and current plant species diversity in the herb layer of temperate forests using long-term Landsat time series. Remote Sens. Environ. 2019, 232. [Google Scholar] [CrossRef]
- Guo, X.; Wilmshurst, J.; McCanny, S.; Fargey, P.; Richard, P. Measuring Spatial and Vertical Heterogeneity of Grasslands Using Remote Sensing Techniques. J. Environ. Inform. 2004, 3, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Guo, X. Evaluating the impacts of nearly 30 years of conservation on grassland ecosystem using Landsat TM images. Grassl. Sci. 2015, 61, 227–242. [Google Scholar] [CrossRef]
- Xu, D.; Guo, X.; Li, Z.; Yang, X.; Yin, H. Measuring the dead component of mixed grassland with Landsat imagery. Remote Sens. Environ. 2014, 142, 33–43. [Google Scholar] [CrossRef]
- Gazette, M. Where Buffalo Roam. Available online: http://www.canada.com/topics/travel/story.html?id=bc04003c-e581-4fab-be85-0e847b66afe7 (accessed on 15 April 2015).
- Xu, D.; Pu, Y.; Guo, X. A Semi-Automated Method to Extract Green and Non-Photosynthetic Vegetation Cover from RGB Images in Mixed Grasslands. Sensors 2020, 20, 6870. [Google Scholar] [CrossRef]
- Munier, S.; Carrer, D.; Planque, C.; Camacho, F.; Albergel, C.; Calvet, J.-C. Satellite Leaf Area Index: Global Scale Analysis of the Tendencies Per Vegetation Type over the Last 17 Years. Remote Sens. 2018, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- Newnham, G.J.; Verbesselt, J.; Grant, I.F.; Anderson, S.A. Relative Greenness Index for assessing curing of grassland fuel. Remote Sens. Environ. 2011, 115, 1456–1463. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, D.; Fang, J. Satellite-based Studies on Large-Scale Vegetation Changes in China. J. Integr. Plant Biol. 2012, 54, 713–728. [Google Scholar] [CrossRef]
- Daughtry, C.; Hunt, E.; McMurtrey, J. Assessing crop residue cover using shortwave infrared reflectance. Remote Sens. Environ. 2004, 90, 126–134. [Google Scholar] [CrossRef]
- Nagler, P.; Daughtry, C.; Goward, S. Plant Litter and Soil Reflectance. Remote Sens. Environ. 2000, 71, 207–215. [Google Scholar] [CrossRef]
- Pacheco, A.; McNairn, H. Evaluating multispectral remote sensing and spectral unmixing analysis for crop residue mapping. Remote Sens. Environ. 2010, 114, 2219–2228. [Google Scholar] [CrossRef]
- Mohammadi, A.; Costelloe, J.F.; Ryu, D. Application of time series of remotely sensed normalized difference water, vegetation and moisture indices in characterizing flood dynamics of large-scale arid zone floodplains. Remote Sens. Environ. 2017, 190, 70–82. [Google Scholar] [CrossRef]
- Gao, B.-C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Xiao, J.; Moody, A. A comparison of methods for estimating fractional green vegetation cover within a desert-to-upland transition zone in central New Mexico, USA. Remote Sens. Environ. 2005, 98, 237–250. [Google Scholar] [CrossRef]
- Gutman, G.; Ignatov, A. The derivation of the green vegetation fraction from NOAA/AVHRR data for use in numerical weather prediction models. Int. J. Remote Sens. 1998, 19, 1533–1543. [Google Scholar] [CrossRef]
- Curran, P.; Steven, M. Multispectral remote sensing for the estimation of green leaf area index [and discussion]. Philos. Trans. R. Soc. London. Ser. A Math. Phys. Sci. 1983, 309, 257–270. [Google Scholar]
- Tucker, C.; Vanpraet, C.; Sharman, M.; Van Ittersum, G. Satellite remote sensing of total herbaceous biomass production in the Senegalese Sahel: 1980–1984. Remote Sens. Environ. 1985, 17, 233–249. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Jianlong, L.; Tiangang, L.; Quangong, C. Estimating grassland yields using remote sensing and GIS technologies in China. N. Z. J. Agric. Res. 1998, 41, 31–38. [Google Scholar] [CrossRef]
- Xu, D.; Guo, X. Some Insights on Grassland Health Assessment Based on Remote Sensing. Sensors 2015, 15, 3070–3089. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Guo, X. A Study of Soil Line Simulation from Landsat Images in Mixed Grassland. Remote Sens. 2013, 5, 4533–4550. [Google Scholar] [CrossRef] [Green Version]
- Bhatti, A.; Mulla, D.; Frazier, B. Estimation of soil properties and wheat yields on complex eroded hills using geostatistics and thematic mapper images. Remote Sens. Environ. 1991, 37, 181–191. [Google Scholar] [CrossRef]
- Ben-Dor, E.; Inbar, Y.; Chen, Y. The reflectance spectra of organic matter in the visible near-infrared and short wave infrared region (400–2500 nm) during a controlled decomposition process. Remote Sens Environ. 1997, 61, 1–15. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Acquisition Dates | Sensor | Landsat |
|---|---|---|---|
| 1986 | 04-05, 06-24, 08-27, 09-28 | TM | Landsat 5 |
| 1987 | 04-24, 05-10, 06-11, 08-30, 10-01 | TM | Landsat 5 |
| 1988 | 04-10, 05-28, 07-15, 07-31, 10-03 | TM | Landsat 5 |
| 1989 | 07-26, 09-28 | TM | Landsat 4 |
| 1989 | 07-02, 07-18, 09-04 | TM | Landsat 5 |
| 1990 | 05-02, 08-22, 09-07, 09-23, 10-09 | TM | Landsat 5 |
| 1991 | 04-03, 05-21, 09-26 | TM | Landsat 5 |
| 1992 | 07-26, 09-28 | TM | Landsat 5 |
| 1993 | 05-10, 08-14 | TM | Landsat 5 |
| 1994 | 04-11, 06-30, 08-17, 09-18 | TM | Landsat 5 |
| 1995 | 05-16, 06-01, 06-17, 08-04, 09-21, 10-23 | TM | Landsat 5 |
| 1996 | 08-22, 10-09 | TM | Landsat 5 |
| 1997 | 05-05, 07-24, 08-25, 09-10 | TM | Landsat 5 |
| 1998 | 05-08, 07-27, 08-12, 08-28, 09-13 | TM | Landsat 5 |
| 1999 | 04-25, 07-14, 09-16 | TM | Landsat 5 |
| 1999 | 08-23 | ETM+ | Landsat 7 |
| 2000 | 04-27, 06-30, 08-01 | TM | Landsat 5 |
| 2000 | 04-19, 07-08, 08-09, 09-26 | ETM+ | Landsat 7 |
| 2001 | 08-12, 10-15 | ETM+ | Landsat 7 |
| 2002 | 05-19, 06-20, 07-06, 08-23 | TM | Landsat 5 |
| 2002 | 04-25, 07-30, 10-02 | ETM+ | Landsat 7 |
| 2003 | 08-10, 08-26 | TM | Landsat 5 |
| 2003 | 04-12, 05-14 | ETM+ | Landsat 7 |
| 2004 | 09-29 | TM | Landsat 5 |
| 2005 | 05-11, 07-14, 07-30, | TM | Landsat 5 |
| 2006 | 04-12, 07-17, 09-03, 10-05 | TM | Landsat 5 |
| 2007 | 07-04, 08-05, 08-21 | TM | Landsat 5 |
| 2008 | 04-17, 08-07, 08-23 | TM | Landsat 5 |
| 2009 | 04-20, 05-22, 08-10, 09-11, 09-27 | TM | Landsat 5 |
| 2010 | 06-26, 09-30 | TM | Landsat 5 |
| 2011 | 06-13, 07-15, 07-31, 10-03 | TM | Landsat 5 |
| 2013 | 05-01, 06-18, 07-04, 08-05, 08-21, 10-08 | OLI | Landsat 8 |
| 2014 | 08-08, 09-25 | OLI | Landsat 8 |
| 2015 | 04-21, 06-08, 07-10, 08-27, 09-12, 09-28, 10-14 | OLI | Landsat 8 |
| 2016 | 06-10, 09-14 | OLI | Landsat 8 |
| 2017 | 07-13 | OLI | Landsat 8 |
| 2018 | 05-15, 10-22 | OLI | Landsat 8 |
| 2019 | 06-03, 08-06, 09-23 | OLI | Landsat 8 |
| 2020 | 08-24, 09-25 | OLI | Landsat 8 |
| Response Variable (y) | Explanatory Variable (y) | Equation of Linear Regression | R Square | p Value |
|---|---|---|---|---|
| Difference in fresh biomass | Difference in NIR reflectance | y = 1543x − 43 | 0.546 | <0.05 |
| Difference in soil organic matter | Difference in SWIR1 reflectance | y = −182x + 2.3 | 0.672 | <0.05 |
| Difference in green cover | Difference in SWIR2 reflectance | y = −348x + 5.7 | 0.579 | <0.05 |
| Difference in litter cover | Difference in LST | y = −479x + 0.12 | 0.605 | <0.05 |
| Indicators | Types | Grazing Cessation | Grazing Reintroduction | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| First Linear Segment | Second Linear Segment | First Linear Segment | Second Linear Segment | ||||||||||
| Time Period | Slope | p Value | Time Period | Slope | p Value | Time Period | Slope | p Value | Time Period | Slope | p Value | ||
| Difference in NIR reflectance | Upland grassland | 1985–1999 | −0.0014 | <0.001 | 1999–2006 | NA | >0.05 | 2006–2014 | 0.0013 | <0.001 | 2014–2020 | NA | >0.05 |
| Sloped grassland | 1985–1997 | −0.0021 | <0.001 | 1997–2006 | NA | >0.05 | 2006–2010 | NA | >0.05 | 2010–2020 | 0.0020 | <0.001 | |
| Valley grassland | 1985–1996 | −0.0029 | <0.001 | 1996–2006 | NA | >0.05 | 2006–2012 | NA | >0.05 | 2012–2020 | 0.0019 | <0.001 | |
| Difference in SWIR1 reflectance | Upland grassland | 1985–1999 | −0.0016 | <0.001 | 1999–2006 | 0.0004 | <0.05 | 2006–2013 | 0.0014 | <0.001 | 2013–2020 | NA | >0.05 |
| Sloped grassland | 1985–1998 | −0.0014 | <0.001 | 1998–2008 | 0.0003 | <0.001 | 2008–2020 | 0.0017 | <0.001 | NA | NA | NA | |
| Valley grassland | 1985–2006 | −0.0015 | <0.001 | NA | NA | NA | 2006–2014 | 0.0009 | <0.001 | 2014–2020 | −0.0009 | <0.05 | |
| Difference in SWIR2 reflectance | Upland grassland | 1985–1998 | −0.0012 | <0.001 | 1998–2006 | NA | >0.05 | 2006–2013 | 0.0010 | <0.001 | 2013–2020 | NA | >0.05 |
| Sloped grassland | 1985–1998 | −0.0008 | <0.001 | 1998–2008 | NA | >0.05 | 2008–2020 | 0.0018 | <0.001 | NA | NA | NA | |
| Valley grassland | 1985–2006 | −0.0013 | <0.001 | NA | NA | NA | 2006–2015 | 0.0016 | <0.001 | 2015–2020 | −0.0009 | <0.05 | |
| Difference in LST | Upland grassland | 1985–1991 | NA | >0.05 | 1991–2000 | −0.04 | <0.001 | 2000–2014 | 0.10 | <0.001 | 2014–2020 | −0.17 | <0.001 |
| Sloped grassland | 1985–1994 | NA | >0.05 | 1994–2006 | −0.01 | <0.05 | 2006–2014 | 0.15 | <0.001 | 2014–2020 | NA | >0.05 | |
| Valley grassland | 1985–1994 | NA | >0.05 | 1994–2005 | −0.11 | <0.001 | 2005–2014 | 0.25 | <0.001 | 2014–2020 | 0.12 | >0.05 | |
| Indicators | Types | Grazing Cessation | Grazing Reintroduction | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| First Linear Segment | Second Linear Segment | First Linear Segment | Second Linear Segment | ||||||||||
| Time Period | Slope | p Value | Time Period | Slope | p Value | Time Period | Slope | p Value | Time Period | Slope | p Value | ||
| Difference in NDVI | Upland grassland | 1985–2000 | −0.0007 | <0.001 | 2000–2007 | NA | >0.05 | 2007–2020 | 0.0008 | <0.001 | NA | NA | NA |
| Sloped grassland | 1985–1997 | −0.0012 | <0.001 | 1997–2006 | NA | >0.05 | 2006–2014 | NA | >0.05 | 2014–2020 | 0.0014 | <0.001 | |
| Valley grassland | 1985–1997 | −0.0015 | <0.001 | 1997–2006 | 0.0009 | <0.001 | 2006–2013 | −0.0007 | <0.001 | 2013–2020 | 0.0020 | <0.001 | |
| Difference in NDWI | Upland grassland | 1985–2006 | −0.0004 | <0.001 | NA | NA | NA | 2006–2020 | 0.0005 | <0.001 | NA | NA | NA |
| Sloped grassland | 1985–1997 | −0.0013 | <0.001 | 1997–2006 | NA | >0.05 | 2006–2013 | NA | >0.05 | 2013–2020 | 0.0013 | <0.001 | |
| Valley grassland | 1985–1995 | −0.0021 | <0.001 | 1995–2006 | 0.0012 | <0.001 | 2006–2014 | 0.0039 | <0.001 | 2014–2020 | 0.0020 | <0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, D.; Harder, J.K.; Xu, W.; Guo, X. Evaluating the Impact of Grazing Cessation and Reintroduction in Mixed Prairie Using Raster Time Series Analysis of Landsat Data. Remote Sens. 2021, 13, 3397. https://doi.org/10.3390/rs13173397
Xu D, Harder JK, Xu W, Guo X. Evaluating the Impact of Grazing Cessation and Reintroduction in Mixed Prairie Using Raster Time Series Analysis of Landsat Data. Remote Sensing. 2021; 13(17):3397. https://doi.org/10.3390/rs13173397
Chicago/Turabian StyleXu, Dandan, Jeff K. Harder, Weixin Xu, and Xulin Guo. 2021. "Evaluating the Impact of Grazing Cessation and Reintroduction in Mixed Prairie Using Raster Time Series Analysis of Landsat Data" Remote Sensing 13, no. 17: 3397. https://doi.org/10.3390/rs13173397
APA StyleXu, D., Harder, J. K., Xu, W., & Guo, X. (2021). Evaluating the Impact of Grazing Cessation and Reintroduction in Mixed Prairie Using Raster Time Series Analysis of Landsat Data. Remote Sensing, 13(17), 3397. https://doi.org/10.3390/rs13173397