
Use of NIRS in Wild Rodents’ Research: A Review of Timid Beginnings


Abstract
1. Brief Introduction to Near-Infrared Spectroscopy (NIRS)
2. The Importance of Rodent Food Studies
3. Why It Is Appropriate to Use the NIRS Method in Wild Rodents
4. Specifics of Using the NIRS Method in Wild Rodents
4.1. Analysis of Faeces
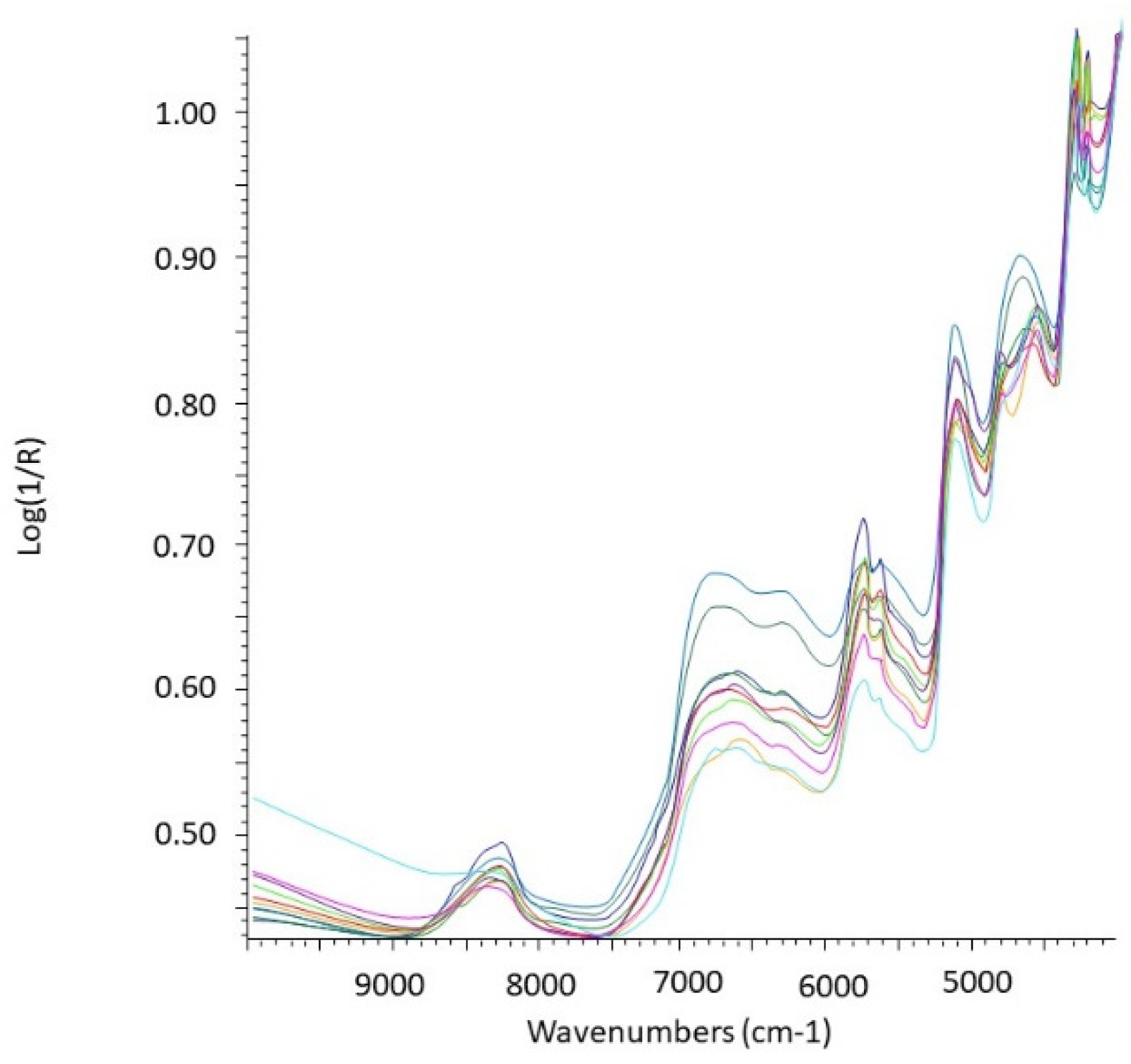
4.2. Analysis of Stomach Contents
4.3. Calibration
5. What Has Already Been Found?
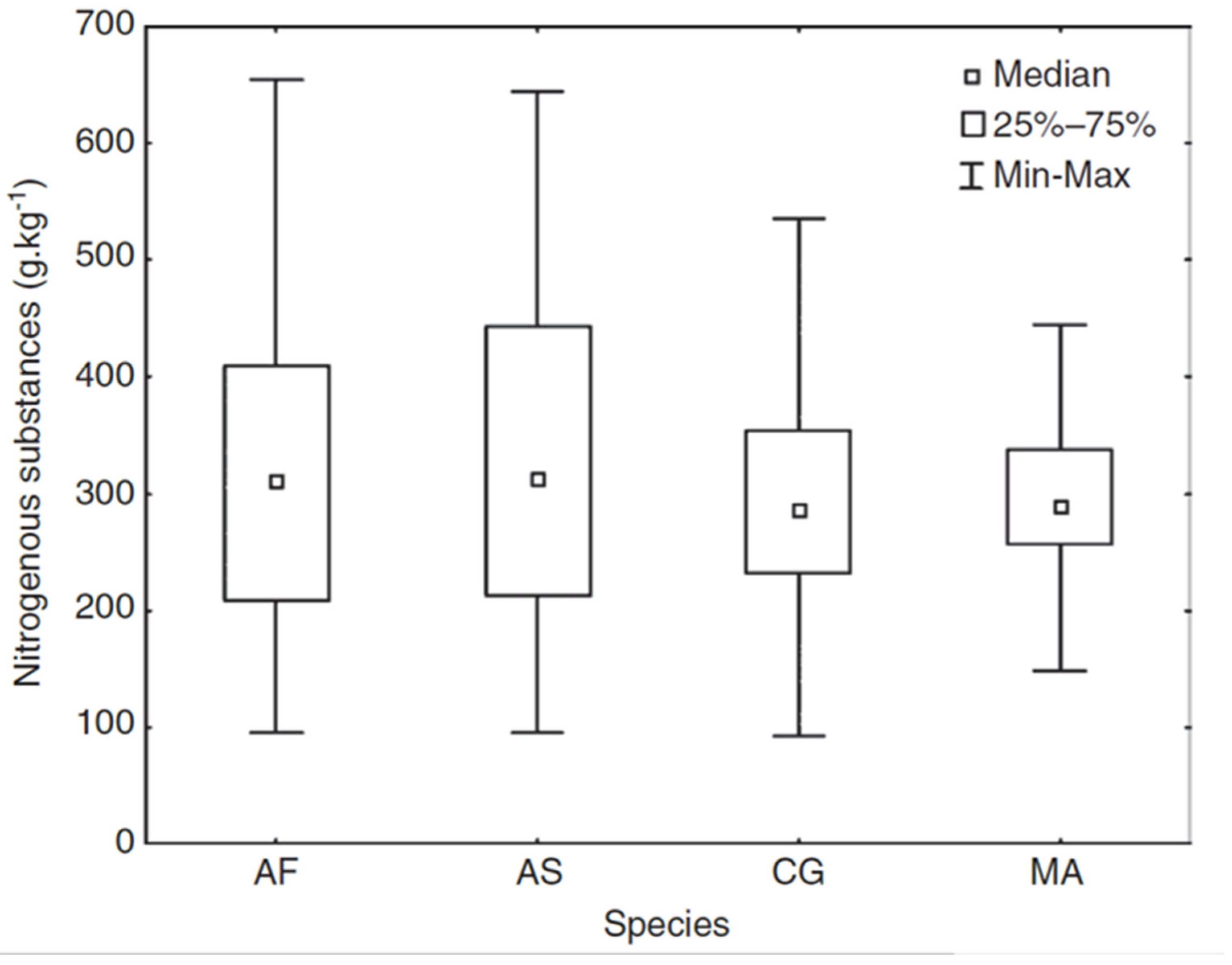
5.1. NIRS Analysis of Rodent Dietary Preferences
5.2. Rodent Stomach Sample Preparation for the NIRS Analysis
5.3. NIRS Analysis of Rodents Food Quality in Forest Habitats
5.4. NIRS Analysis of Rodents Food Quality in Arable Landscape
6. Suggestions for Further Research
- -
- Diseases. Small rodents are hosts and inter-hosts of a huge number of animal diseases and zoonoses, such as Francisella tularensis [61,62], Borrelia sp. [63], Yersinia sp. [64], Brucella sp. [65], Toxoplasma gondii [66], viruses and many others [67,68,69,70,71]. A reliable methodology for detecting pathogens (similar to [70,72]) from rodent faecal samples could help to estimate occurrence of these pathogens and make preventive screenings.
- -
- -
- Food quality changes and rodent population oscillations (especially population declines). For ecologists, the study of rodent seasonal oscillation is one of the unresolved topics. The role of diet has been widely discussed but we have not obtained clear conclusions yet [31,75,76]. NIRS makes it possible to detect the diet of many individuals in different populations during the oscillating cycle and it can help to clarify the role of food quality and quantity on population fluctuations.
- -
- Hormonal changes during population oscillation. According to recently published knowledge, gastrointestinal microbiota produce hormones, which affect hosts health and behaviour, such as cortisol and catecholamines (e.g., adrenaline) [77,78]. The levels of these stress hormones change according to population oscillation [79]. These hormones or their precursors might be detectable in faeces. The estimation of stress hormones levels would help to predict an increase/decrease in population and excessive use of poisons could be prevented. The study of hormone levels from faeces may be a promising method for many other types of studies, such as physiological, environmental, or agrotechnical studies.
- -
- Field, orchard, and forest management. In arable landscapes, NIRS can ascertain the real impact of small rodents on crops. Although these mammals sometimes cause great damage in agriculture, it is not clear when and what they eat in the fields; sometimes there are many rodents, even in crops that are not suitable food for them (for example, corn, wheat, or barley for common voles) [80,81]. Weeds growing in fields with these crops may be a more suitable food for rodents [82]. NIRS can help answer the question: what is the proportion of crops and weeds in the diet of rodents? Of course, this proportion can change during the growing season or the ripening of the crop [83]. The NIRS method could help determine how much and when (at what stage of the population cycle, crop ripening, or season) rodents consume the target crops. With this knowledge, interventions against rodent damage could be more precise. Similarly in orchards [84,85] and forests [86,87]: what proportion of rodents’ diet is made up of the bark and sap of young trees? When do rodents consume them? Answers to such questions would allow more accurate crop protection and lower the use of rodenticides.
- -
- Studies of Norway rats in cities. Although rats are the most common mammalian health threat in cities, exhaustive information about their ecology is lacking [88]. Their faeces are easy to find and continuous monitoring of their health status (zoonoses occurrence, poison levels, and stress hormones) and population parameters (breeding etc.) could be helpful to planning their reduction.
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Givens, D.I.; De Boever, J.L.; Deaville, E.R. The principles, practices and some future applications of near infrared spectroscopy for predicting the nutritive value of foods for animals and humans. Nutr. Res. Rev. 1997, 10, 83–84. [Google Scholar] [CrossRef]
- Foley, W.J.; McIlwee, A.; Lawler, I.R.; Aragones, L.; Woolnough, A.P.; Berding, N. Ecological applications of near infrared reflectance spectroscopy—A tool for rapid, cost-effective prediction of composition of plant and animal tissues and aspects of animal performance. Oecologia 1998, 116, 293–305. [Google Scholar] [CrossRef]
- Shenk, J.S.; Westerhaus, M.O. The Application of near Infrared Reflectance Spectroscopy (NIRS) to Forage Analysis. In Forage Quality, Evaluation and Utilization; American Society of Agronomy: Madison, WI, USA, 1994; pp. 406–449. [Google Scholar]
- Tolleson, D.R.; Angerer, J.P. The application of near infrared spectroscopy to predict faecal nitrogen and phosphorus in multiple ruminant herbivore species. Rangel. J. 2021, 42, 415–423. [Google Scholar] [CrossRef]
- Kaneko, H.; Lawler, I.R. Can Near Infrared Spectroscopy be used to improve assessment of marine mammal diets via fecal analysis? Mar. Mammal Sci. 2006, 22, 261–275. [Google Scholar] [CrossRef]
- Moore, B.D.; Lawler, I.R.; Wallis, I.R.; Beale, C.M.; Foley, W.J. Palatability mapping: A koala’s eye view of spatial variation in habitat quality. Ecology 2010, 91, 3165–3176. [Google Scholar] [CrossRef] [PubMed]
- Wiedower, E.E.; Kouba, A.J.; Vance, C.K.; Hansen, R.L.; Stuth, J.W.; Tolleson, D.R. Fecal Near Infrared Spectroscopy to Discriminate Physiological Status in Giant Pandas. PLoS ONE 2012, 7, e38908. [Google Scholar] [CrossRef] [PubMed]
- Counsell, K.R.; Vance, C.K. Recent Advances of near Infrared Spectroscopy in Wildlife and Ecology Studies. NIR News 2016, 27, 29–32. [Google Scholar] [CrossRef]
- Kinoshita, K.; Miyazaki, M.; Morita, H.; Vassileva, M.; Tang, C.; Li, D.; Ishikawa, O.; Kusunoki, H.; Tsenkova, R. Spectral pattern of urinary water as a biomarker of estrus in the giant panda. Sci. Rep. 2012, 2, 7. [Google Scholar] [CrossRef]
- Kinoshita, K.; Kuze, N.; Kobayashi, T.; Miyakawa, E.; Narita, H.; Inoue-Murayama, M.; Idani, G.; Tsenkova, R. Detection of urinary estrogen conjugates and creatinine using near infrared spectroscopy in Bornean orangutans (Pongo Pygmaeus). Primates 2016, 57, 51–59. [Google Scholar] [CrossRef]
- Iverson, S.J.; Arnould, J.P.Y.; Boyd, I.L. Milk fatty acid signatures indicate both major and minor shifts in the diet of lactating Antarctic fur seals. Can. J. Zool. 1997, 75, 188–197. [Google Scholar] [CrossRef]
- Beć, K.B.; Grabska, J.; Huck, C.W. Near-Infrared Spectroscopy in Bio-Applications. Molecules 2020, 25, 2948. [Google Scholar] [CrossRef]
- Vance, C.K.; Tolleson, D.R.; Kinoshita, K.; Rodriguez, J.; Foley, W.J. Near Infrared Spectroscopy in Wildlife and Biodiversity. J. Near Infrared Spectrosc. 2016, 24, 1–25. [Google Scholar] [CrossRef]
- Beć, K.B.; Grabska, J.; Huck, C.W. Principles and Applications of Miniaturized Near-Infrared (NIR) Spectrometers. Chem. A Eur. J. 2021, 27, 1514–1532. [Google Scholar] [CrossRef]
- Pereira, J.F.Q.; Silva, C.S.; Vieira, M.J.L.; Pimentel, M.F.; Braz, A.; Honorato, R.S. Evaluation and identification of blood stains with handheld NIR spectrometer. Microchem. J. 2017, 133, 561–566. [Google Scholar] [CrossRef]
- Dickman, C.R. Rodent-Ecosystem Relationships: A Review. In Ecologically-Based Management of Rodent Pests; Singleton, G.R., Hinds, L.A., Leirs, H., Zhang, Z., Eds.; Australian Centre for International Agricultural Research: Canberra, Australia, 1999; Volume 59, pp. 113–133. [Google Scholar]
- Fischer, C.; Gayer, C.; Kurucz, K.; Riesch, F.; Tscharntke, T.; Batáry, P. Ecosystem services and disservices provided by small rodents in arable fields: Effects of local and landscape management. J. Appl. Ecol. 2018, 55, 548–558. [Google Scholar] [CrossRef]
- Verde Arregoitia, L.D.; D’Elía, G. Classifying rodent diets for comparative research. Mamm. Rev. 2021, 51, 51–65. [Google Scholar] [CrossRef]
- Hansson, L. Intraspecific Variation in Dynamics: Small Rodents between Food and Predation in Changing Landscapes. Oikos 1999, 86, 159–169. [Google Scholar] [CrossRef]
- Ieradi, L.A.; Moreno, S.; Bolívar, J.P.; Cappai, A.; Di Benedetto, A.; Cristaldi, M. Free-living rodents as bioindicators of genetic risk in natural protected areas. Environ. Pollut. 1998, 102, 265–268. [Google Scholar] [CrossRef]
- Singleton, G.R.; Jacob, J.; Krebs, C.J.; Monadjem, A. A meeting of mice and men: Rodent impacts on food security, human diseases and wildlife conservation; Ecosystem benefits; Fascinating biological models. Wildl. Res. 2015, 42, 83–85. [Google Scholar] [CrossRef][Green Version]
- Jacob, J.; Imholt, C.; Caminero-Saldaña, C.; Couval, G.; Giraudoux, P.; Herrero-Cófreces, S.; Horváth, G.; Luque-Larena, J.J.; Tkadlec, E.; Wymenga, E. Europe-wide outbreaks of common voles in 2019. J. Pest Sci. 2020, 93, 703–709. [Google Scholar] [CrossRef]
- Wilson, D.E.; Lacher, T.E.; Mittermeier, R.A. Handbook of the Mammals of the World, Volume 6 and 7: Lagomorphs and Rodents I and II.; Wilson, D.E., Lacher, T.E., Mittermeier, R.A., Eds.; Lynx: Barcelona, Spain, 2017. [Google Scholar]
- Singleton, G.R.; Belmain, S.R.; Brown, P.R.; Hardy, B. Rodent Outbreaks: Ecology and Impacts; Singleton, G.R., Belmain, S., Brown, P.R., Hardy, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2010; ISBN 9789712202575. [Google Scholar]
- Meerburg, B.G.; Singleton, G.R.; Leirs, H. The year of the rat ends—Time to fight hunger! Pest Manag. Sci. 2009, 65, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.; Tkadlec, E. Rodent outbreaks in Europe: Dynamics and damage. In Rodent Outbreaks—Ecology and Impacts; Singleton, G.R., Belmain, S., Brown, P.R., Hardy, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2010; pp. 207–223. ISBN 9789712202575. [Google Scholar]
- Wood, B.J.; Singleton, G.R. Rodents in agriculture and forestry. In Rodent Pests and Their Control, 2nd ed.; University of Reading: Reading, UK, 2015; pp. 33–80. ISBN 9781845938178. [Google Scholar]
- Hansen, S.C.; Stolter, C.; Jacob, J. The smell to repel: The effect of odors on the feeding behavior of female rodents. Crop Prot. 2015, 78, 270–276. [Google Scholar] [CrossRef]
- Heroldová, M. Diet of four rodent species from Robinia pseudo-acacia stands in South Moravia. Acta Theriol. 1994, 39, 333–337. [Google Scholar] [CrossRef]
- Holišová, V. The food of Clethrionomys glareolus at different population densities. Acta Sci. Nat. Brno 1971, 5, 1–43. [Google Scholar]
- Butet, A. Does food quality Drive Cycle in Microtus arvalis? Study on a French Atlantic Marsh Population. Proc. I Eur. Congr. Mammal. 1996, 177, 177–188. [Google Scholar]
- Windley, H.R.; Wallis, I.R.; DeGabriel, J.L.; Moore, B.D.; Johnson, C.N.; Foley, W.J. A faecal index of diet quality that predicts reproductive success in a marsupial folivore. Oecologia 2013, 173, 203–212. [Google Scholar] [CrossRef]
- Wolff, J.O.; Sherman, P.W. Rodent Societies: An Ecological and Evolutionary Perspective; The University Chicago Press: Chicago, IL, USA; London, UK, 2007; ISBN 9780226905372. [Google Scholar]
- Heroldová, M.; Čižmár, D.; Tkadlec, E. Predicting rodent impact in crop fields by near-infrared reflectance spectroscopy analysis of their diet preferences. Crop Prot. 2010, 29, 773–776. [Google Scholar] [CrossRef]
- Lean, I.J.; Golder, H.M.; Black, J.L.; King, R.; Rabiee, A.R. In vivo indices for predicting acidosis risk of grains in cattle: Comparison with in vitro methods. J. Anim. Sci. 2013, 91, 2823–2835. [Google Scholar] [CrossRef]
- André, J.; Lawler, I.R. Near infrared spectroscopy as a rapid and inexpensive means of dietary analysis for a marine herbivore, dugong Dugong dugon. Mar. Ecol. Prog. Ser. 2003, 257, 259–266. [Google Scholar] [CrossRef][Green Version]
- Chivers, D.J.; Langer, P. The Digestive System in Mammals: Food, Form and Function; Cambridge University Press: Cambridge, UK, 1994; ISBN 0-521-44016-5. [Google Scholar]
- Langer, P. The digestive tract and life history of small mammals. Mamm. Rev. 2002, 32, 107–131. [Google Scholar] [CrossRef]
- Butet, A.; Delettre, Y.R. Diet differentiation between European arvicoline and murine rodents. Acta Theriol. 2011, 56, 297–304. [Google Scholar] [CrossRef]
- Givens, D.I.; Deaville, E.R. The current and future role of near infrared reflectance spectroscopy in animal nutrition: A review. Aust. J. Agric. Res. 1999, 50, 1131–1145. [Google Scholar] [CrossRef]
- Steyaert, S.M.; Hütter, F.J.; Elfström, M.; Zedrosser, A.; Hackländer, K.; Lê, M.H.; Windisch, W.M.; Swenson, J.E.; Isaksson, T. Faecal spectroscopy: A practical tool to assess diet quality in an opportunistic omnivore. Wildl. Biol. 2012, 18, 431–438. [Google Scholar] [CrossRef]
- Zijlstra, R.T.; Swift, M.L.; Wang, L.F.; Scott, T.A.; Edney, M.J. Short communication: Near infrared reflectance spectroscopy accurately predicts the digestible energy content of barley for pigs. Can. J. Anim. Sci. 2011, 91, 301–304. [Google Scholar] [CrossRef]
- Rivero-Marcotegui, A.; Olivera-Olmedo, J.E.; Valverde-Visus, F.S.; Palacios-Sarrasqueta, M.; Grijalba-Uche, A.; García-Merlo, S. Water, fat, nitrogen, and sugar content in feces: Reference intervals in children. Clin. Chem. 1998, 44, 1540–1544. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Bjørnstad, O.N.; Stenseth, N.C. Density Dependence in Voles and Mice: A Comparative Study. Ecology 1999, 80, 638–650. [Google Scholar] [CrossRef]
- Suchomel, J.; Heroldová, M.; Mlček, J.; Šustová, K.; Růžičková, J.; Remeš, M. Winter Diet Preferences of Apodemus flavicollis Under Influence of Supplementary Food in Pheasantry. In Proceedings of the 9th International Mammalogical Congress, The Scientific Council of Japan, The Mammalogical Society of Japan, Sapporo, Japan, 31 July 2005; p. 301. [Google Scholar]
- Cox, P.G.; Rayfield, E.J.; Fagan, M.J.; Herrel, A.; Pataky, T.C.; Jeffery, N. Functional evolution of the feeding system in rodents. PLoS ONE 2012, 7, e36299. [Google Scholar] [CrossRef]
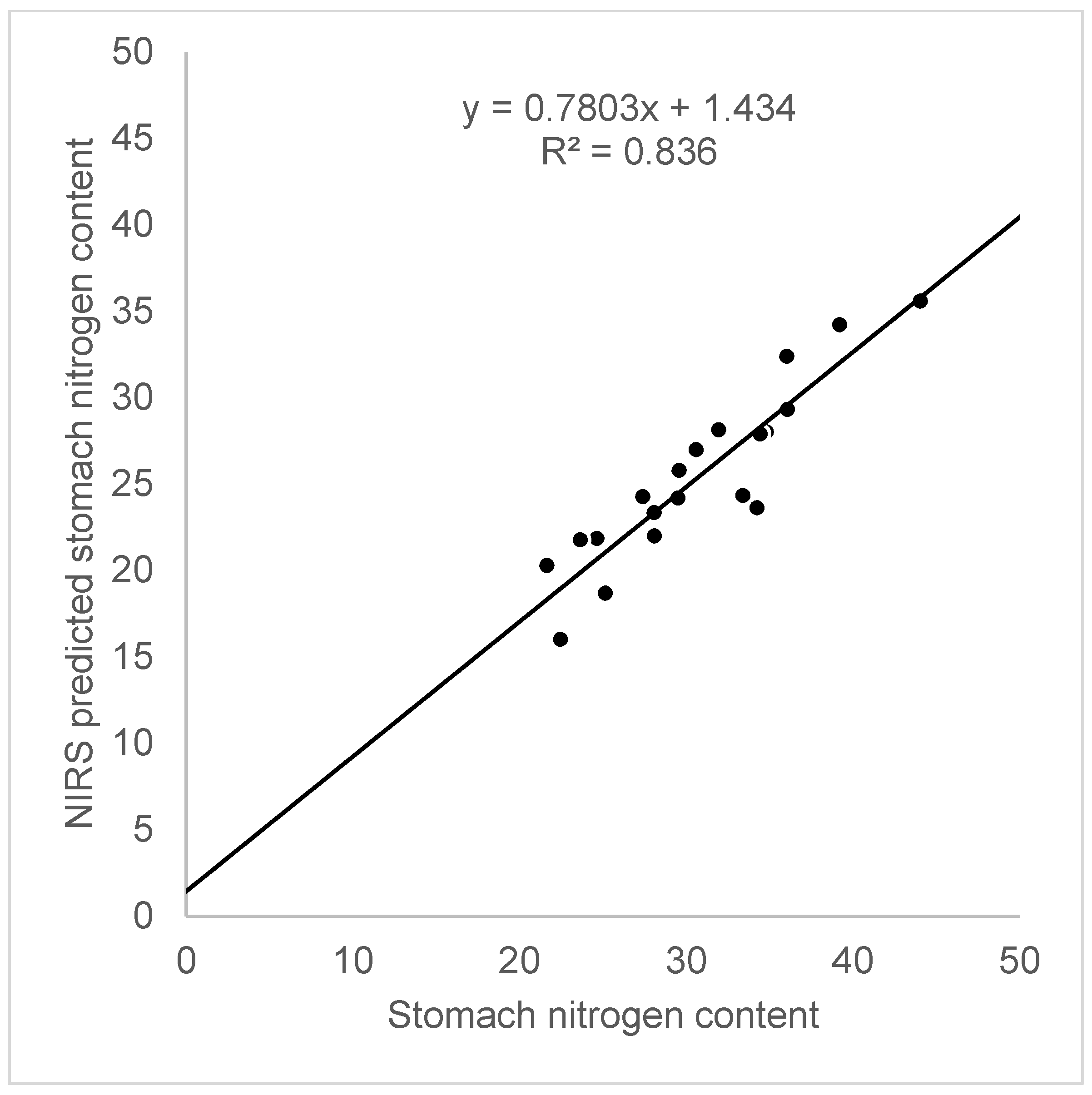
- Čepelka, L.; Heroldová, M.; Jánová, E.; Suchomel, J.; Čižmár, D. Rodent stomach sample preparation for nitrogen NIRS analysis. Mamm. Biol. 2017, 87, 13–16. [Google Scholar] [CrossRef]
- Čepelka, L.; Heroldová, M.; Jánová, E.; Suchomel, J. The Dynamics of Nitrogenous Substances in Rodent Diet in a Forest Environment. Mammalia 2014, 78, 327–333. [Google Scholar] [CrossRef]
- Čepelka, L.; Heroldová, M.; Jánová, E.; Suchomel, J. Dynamics of nitrogenous substance content in the diet of the wood mouse (Apodemus sylvaticus). Acta Univ. Agric. Silvic. Mendel. Brun. 2013, 61, 1–7. [Google Scholar] [CrossRef]
- Jánová, E.; Bryja, J.; Čižmár, D.; Čepelka, L.; Heroldová, M. A new method for assessing food quality in common vole (Microtus arvalis) populations. Eur. J. Wildl. Res. 2015, 61, 57–62. [Google Scholar] [CrossRef]
- Jánová, E.; Heroldová, M.; Čepelka, L. Rodent food quality and its relation to crops and other environmental and population parameters in an agricultural landscape. Sci. Total Environ. 2016, 562, 164–169. [Google Scholar] [CrossRef]
- Heroldová, M.; Jánová, E. Feeding strategy of two rodent species in a set-Aside field and its influence on alimentary tract morphometry. Mammalia 2019, 83, 34–40. [Google Scholar] [CrossRef]
- Burgin, C.J.; Colella, J.P.; Kahn, P.L.; Upham, N.S. How many species of mammals are there? J. Mammal. 2018, 99, 1–14. [Google Scholar] [CrossRef]
- Vance, C.K.; Kouba, A.J.; Willard, S.T. Near Infrared Spectroscopy Applications in Amphibian Ecology and Conservation: Gender and Species Identification. NIR News 2014, 25, 10–15. [Google Scholar] [CrossRef]
- Tolleson, D.R.; Schafer, D.W. Evaluation of non-invasive bioforensic techniques for determining the age of hot-iron brand burn scars in cattle. Transl. Anim. Sci. 2021, 1–10. [Google Scholar] [CrossRef]
- Goeser, J.P.; Hoffman, P.C.; Combs, D.K. Modification of a rumen fluid priming technique for measuring in vitro neutral detergent fiber digestibility. J. Dairy Sci. 2009, 92, 3842–3848. [Google Scholar] [CrossRef]
- Gagnon, D.D.; Hancock, C.; Mccue, A.; Beckett-Brown, N.; Gagnon, J.; Williams, L.; Marsh, D.; Munten, S. Muscle cooling modulates tissue oxidative and biochemical responses but not energy metabolism during exercise. Eur. J. Appl. Physiol. 2020, 120, 1761–1775. [Google Scholar] [CrossRef]
- Drozdz, A. Digestibility and Assimilation of Natural Foods in Small Rodents. Acta Theriol. 1968, 13, 367–389. [Google Scholar] [CrossRef]
- Kurien, B.T.; Everds, N.E.; Scofield, R.H. Experimental animal urine collection: A review. Lab. Anim. 2004, 38, 333–361. [Google Scholar] [CrossRef]
- Rose, C.; Parker, A.; Jefferson, B.; Cartmell, E. The characterization of feces and urine: A review of the literature to inform advanced treatment technology. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1827–1879. [Google Scholar] [CrossRef] [PubMed]
- Rossow, H.; Sissonen, S.; Koskela, K.A.; Kinnunen, P.M.; Hemmilä, H.; Niemimaa, J.; Huitu, O.; Kuusi, M.; Vapalahti, O.; Henttonen, H.; et al. Detection of Francisella tularensis in Voles in Finland. Vector Borne Zoonotic Dis. 2014, 14, 193–198. [Google Scholar] [CrossRef]
- Jeske, K.; Tomaso, H.; Imholt, C.; Schulz, J.; Beerli, O.; Suchomel, J.; Heroldová, M.; Jacob, J.; Staubach, C.; Ulrich, R.G. Detection of Francisella tularensis in three vole species in Central Europe. Transbound. Emerg. Dis. 2019, 66, 1029–1032. [Google Scholar] [CrossRef] [PubMed]
- Hanincova, K.; Schäfer, S.M.; Etti, S.; Sewell, H.S.; Taragelová, V.; Ziak, D.; Labuda, M.; Kurtenbach, K. Association of Borrelia afzelii with rodents in Europe. Parasitology 2003, 126, 11–20. [Google Scholar] [CrossRef]
- Backhans, A.; Fellström, C.; Lambertz, S.T. Occurrence of pathogenic Yersinia enterocolitica and Yersinia pseudotuberculosis in small wild rodents. Epidemiol. Infect. 2011, 139, 1230–1238. [Google Scholar] [CrossRef]
- Zheludkov, M.M.; Tsirelson, L.E. Reservoirs of Brucella infection in nature. Biol. Bull. 2010, 37, 709–715. [Google Scholar] [CrossRef]
- Gotteland, C.; Chaval, Y.; Villena, I.; Galan, M.; Geers, R.; Aubert, D.; Poulle, M.L.; Charbonnel, N.; Gilot-Fromont, E. Species or local environment, what determines the infection of rodents by Toxoplasma gondii? Parasitology 2014, 141, 259–268. [Google Scholar] [CrossRef]
- Obiegala, A.; Heuser, E.; Ryll, R.; Imholt, C.; Fürst, J.; Prautsch, L.M.; Plenge-Bönig, A.; Ulrich, R.G.; Pfeffer, M. Norway and black rats in Europe: Potential reservoirs for zoonotic arthropod-borne pathogens? Pest Manag. Sci. 2019, 75, 1556–1563. [Google Scholar] [CrossRef]
- Mrochen, D.M.; Schulz, D.; Fischer, S.; Jeske, K.; El Gohary, H.; Reil, D.; Imholt, C.; Trübe, P.; Suchomel, J.; Tricaud, E.; et al. Wild rodents and shrews are natural hosts of Staphylococcus aureus. Int. J. Med. Microbiol. 2018, 308, 590–597. [Google Scholar] [CrossRef]
- Heroldová, M.; Pejčoch, M.; Bryja, J.; Jánová, E.; Suchomel, J.; Tkadlec, E. Tula Virus in Populations of Small Terrestrial Mammals in a Rural Landscape. Vector Borne Zoonotic Dis. 2010, 10, 599–603. [Google Scholar] [CrossRef]
- Sakudo, A.; Suganuma, Y.; Kobayashi, T.; Onodera, T.; Ikuta, K. Near-infrared spectroscopy: Promising diagnostic tool for viral infections. Biochem. Biophys. Res. Commun. 2006, 341, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Hardestam, J.; Karlsson, M.; Falk, K.I.; Olsson, G.; Klingström, J.; Lundkvist, Å. Puumala hantavirus excretion kinetics in bank voles (Myodes glareolus). Emerg. Infect. Dis. 2008, 14, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.M.B.; Mutsaers, M.; Garcia, G.A.; David, M.R.; Pavan, M.G.; Petersen, M.T.; Corrêa-Antônio, J.; Couto-Lima, D.; Maes, L.; Dowell, F.; et al. High throughput estimates of Wolbachia, Zika and chikungunya infection in Aedes aegypti by near-infrared spectroscopy to improve arbovirus surveillance. Commun. Biol. 2021, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vávrová, M.; Gargošová, Z.H.; Šucman, E.; Večerek, V.; Kořinek, P.; Zukal, J.; Zejda, J.; Sebestiánová, N.; Kubištová, I. Game animals and small terrestrial mammals—Suitable bioindicators for the pollution assessment in agrarian ecosystems. Fresenius Environ. Bull. 2003, 12, 165–172. [Google Scholar]
- Casale, M.; Bagnasco, L.; Giordani, P.; Mariotti, M.G.; Malaspina, P. NIR spectroscopy as a tool for discriminating between lichens exposed to air pollution. Chemosphere 2015, 134, 355–360. [Google Scholar] [CrossRef]
- Yoccoz, N.G.; Stenseth, N.C.; Henttonen, H.; Prévot-Julliard, A.-C. Effects of food addition on the seasonal density dependent structure of bank vole Clethrionomys glareolus populations. J. Anim. Ecol. 2001, 70, 713–720. [Google Scholar] [CrossRef]
- White, T.C.R. The role of food, weather and climate in limiting the abundance of animals. Biol. Rev. 2008, 83, 227–248. [Google Scholar] [CrossRef]
- Asano, Y.; Hiramoto, T.; Nishino, R.; Aiba, Y.; Kimura, T.; Yoshihara, K.; Koga, Y.; Sudo, N. Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, 1288–1295. [Google Scholar] [CrossRef]
- Boonstra, R.; Hik, D.; Singleton, G.R.; Tinnikov, A. The Impact of Predator-Induced Stress on the Snowshoe Hare Cycle. Ecol. Monogr. 1998, 68, 371–394. [Google Scholar] [CrossRef]
- Korpimäki, E.; Brown, P.R.; Jacob, J.; Pech, R.P. The Puzzles of Population Cycles and Outbreaks of Small Mammals Solved? Bioscience 2004, 54, 1071–1079. [Google Scholar] [CrossRef]
- Jánová, E.; Heroldová, M.; Konecny, A.; Bryja, J.; Konečný, A. Traditional and diversified crops in South Moravia (Czech Republic): Habitat preferences of common vole and mice species. Mamm. Biol. 2011, 76, 570–576. [Google Scholar] [CrossRef]
- Dell’Agnello, F.; Natali, C.; Bertolino, S.; Fattorini, L.; Fedele, E.; Foggi, B.; Martini, M.; Pisani, C.; Riga, F.; Sgarlata, A.; et al. Assessment of seasonal variation of diet composition in rodents using DNA barcoding and Real-Time PCR. Sci. Rep. 2019, 9, 14124. [Google Scholar] [CrossRef] [PubMed]
- Htwe, N.M.; Singleton, G.R.; Johnson, D.E. Interactions between rodents and weeds in a lowland rice agro-ecosystem: The need for an integrated approach to management. Integr. Zool. 2019, 14, 396–409. [Google Scholar] [CrossRef]
- Heroldová, M.; Zejda, J.; Zapletal, M.; Obdržálková, D.; Jánová, E.; Bryja, J.; Tkadlec, E. Foraging strategy of small rodents in winter rape. Plant Soil Environ. 2004, 50, 175–181. [Google Scholar] [CrossRef]
- Bertolino, S.; Asteggiano, L.; Saladini, M.A.; Giordani, L.; Vittone, G.; Alma, A. Environmental factors and agronomic practices associated with Savi’s pine vole abundance in Italian apple orchards. J. Pest Sci. 2015, 88, 135–142. [Google Scholar] [CrossRef]
- Suchomel, J.; Šipoš, J.; Čepelka, L.; Heroldová, M. The impact of Microtus arvalis and Lepus europaeus on apple trees by trunk bark gnawing. Plant Prot. Sci. 2019, 55, 142–147. [Google Scholar] [CrossRef]
- Krojerová-Prokešová, J.; Homolka, M.; Heroldová, M.; Barančeková, M.; Baňař, P.; Kamler, J.; Modlinger, R.; Purchart, L.; Zejda, J.; Suchomel, J. Patterns of vole gnawing on saplings in managed clearings in Central European forests. For. Ecol. Manag. 2018, 408, 137–147. [Google Scholar] [CrossRef]
- Leiva, M.J.; Díaz-Maqueda, A. Fast-growing seeds and delayed rodent predatory activity in the seeding season: A combined mechanism to escape and survive rodent predation in Quercus ilex subsp. ballota L. acorns and seedlings. For. Ecol. Manag. 2016, 380, 23–30. [Google Scholar] [CrossRef]
- Byers, K.A.; Lee, M.J.; Patrick, D.M.; Himsworth, C.G. Rats about town: A systematic review of rat movement in urban ecosystems. Front. Ecol. Evol. 2019, 7, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constituent | N | Mean | SD | SEC | R2 cal | SECV | R2cv | SD/SECV |
|---|---|---|---|---|---|---|---|---|
| Nitrogen content | 34 | 25.3 | 4.8 | 43.5 | 0.84 | 44.1 | 0.84 | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čepelka, L.; Jánová, E.; Suchomel, J.; Heroldová, M. Use of NIRS in Wild Rodents’ Research: A Review of Timid Beginnings. Remote Sens. 2021, 13, 3268. https://doi.org/10.3390/rs13163268
Čepelka L, Jánová E, Suchomel J, Heroldová M. Use of NIRS in Wild Rodents’ Research: A Review of Timid Beginnings. Remote Sensing. 2021; 13(16):3268. https://doi.org/10.3390/rs13163268
Chicago/Turabian StyleČepelka, Ladislav, Eva Jánová, Josef Suchomel, and Marta Heroldová. 2021. "Use of NIRS in Wild Rodents’ Research: A Review of Timid Beginnings" Remote Sensing 13, no. 16: 3268. https://doi.org/10.3390/rs13163268
APA StyleČepelka, L., Jánová, E., Suchomel, J., & Heroldová, M. (2021). Use of NIRS in Wild Rodents’ Research: A Review of Timid Beginnings. Remote Sensing, 13(16), 3268. https://doi.org/10.3390/rs13163268