
Phytoplankton Bloom Dynamics in the Baltic Sea Using a Consistently Reprocessed Time Series of Multi-Sensor Reflectance and Novel Chlorophyll-a Retrievals

 , ,
, ,
Abstract
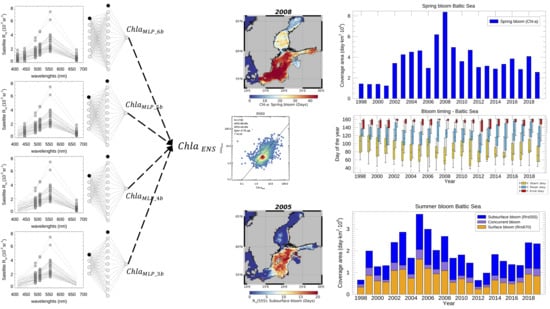
:
1. Introduction
2. Materials and Methods
2.1. Space-Borne
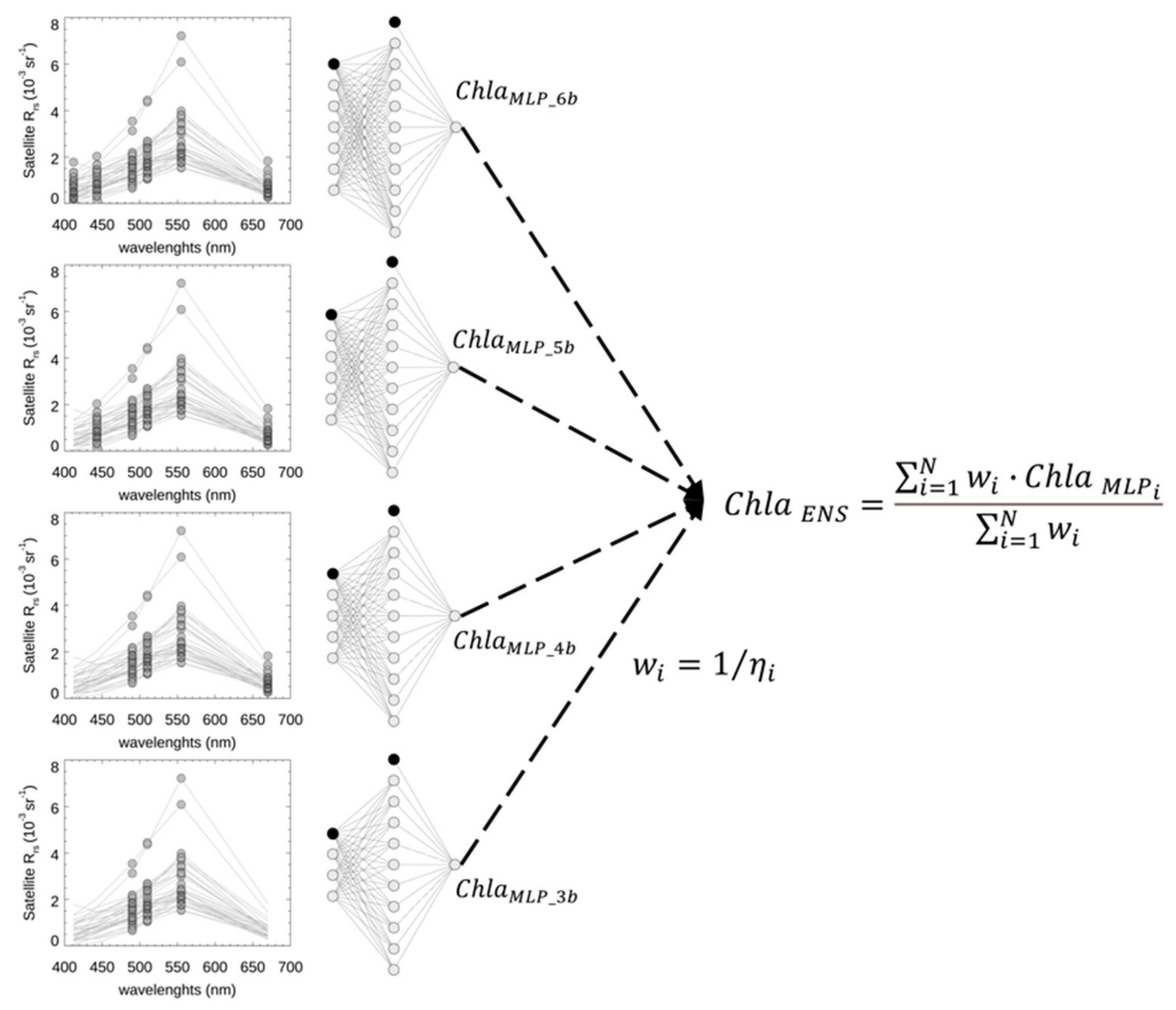
2.2. The Ensemble of MLP Bio-Optical Algorithms
- : 6 bands with values at 412, 443, 490, 510, 555, and 670 nm.
- : 5 bands with values at 443, 490, 510, 555, and 670 nm.
- : 4 bands with values at 490, 510, 555, and 670 nm.
- : 3 bands with values at 490, 510, and 555 nm.
2.2.1. MLP Scheme
2.2.2. Novelty Index and MLP Applicability Range
2.2.3. Ensemble Scheme
2.3. The Band-Ratio Bio-Optical Algorithms
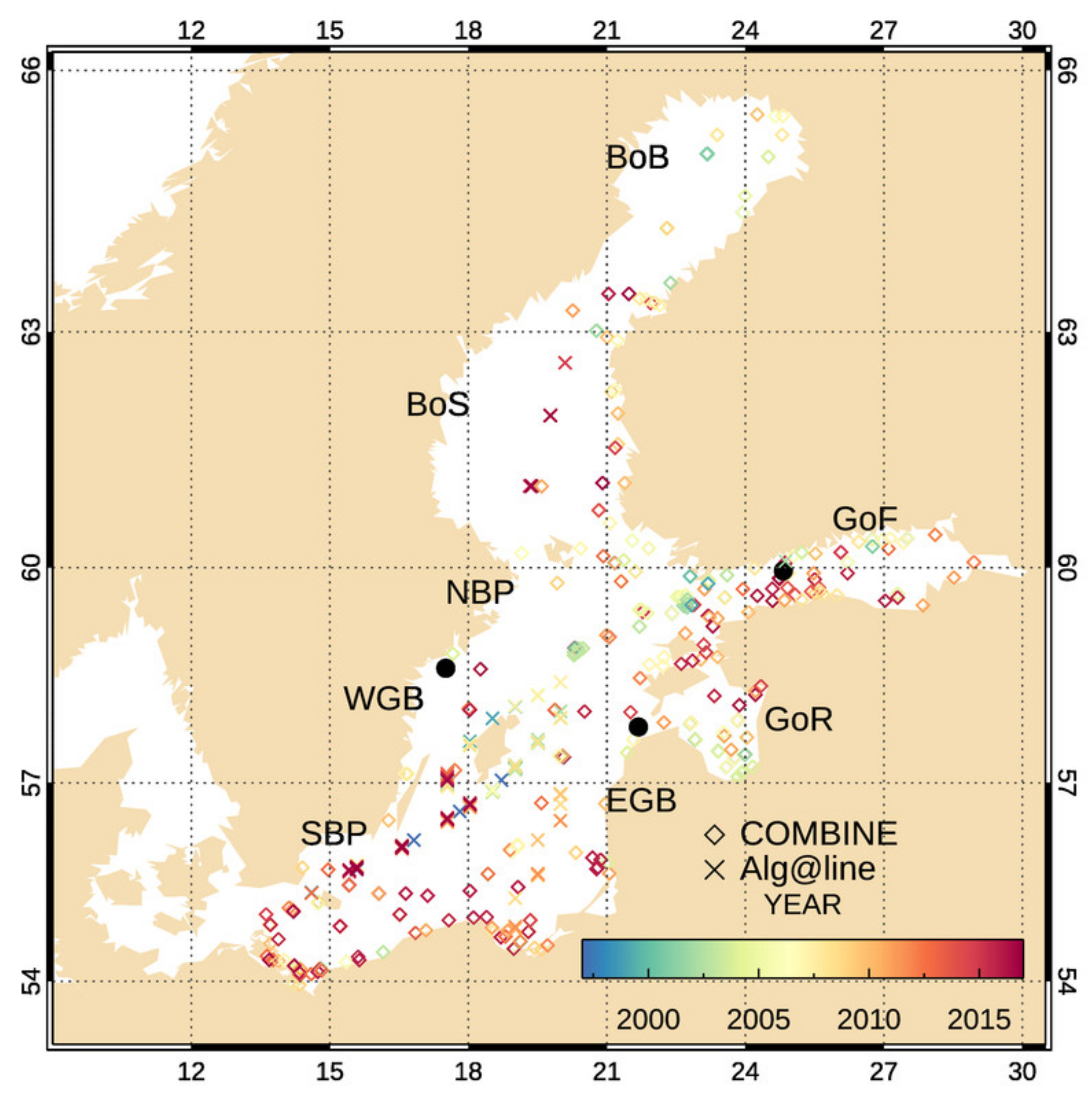
2.4. In Situ Data for Validation
2.4.1. In Situ Automated Radiometry
2.4.2. In Situ Data for the Assessment of the Retrieval
2.5. Statistical Figures
2.6. Indicators of the Spring and Summer Phytoplankton Bloom Dynamics
3. Results
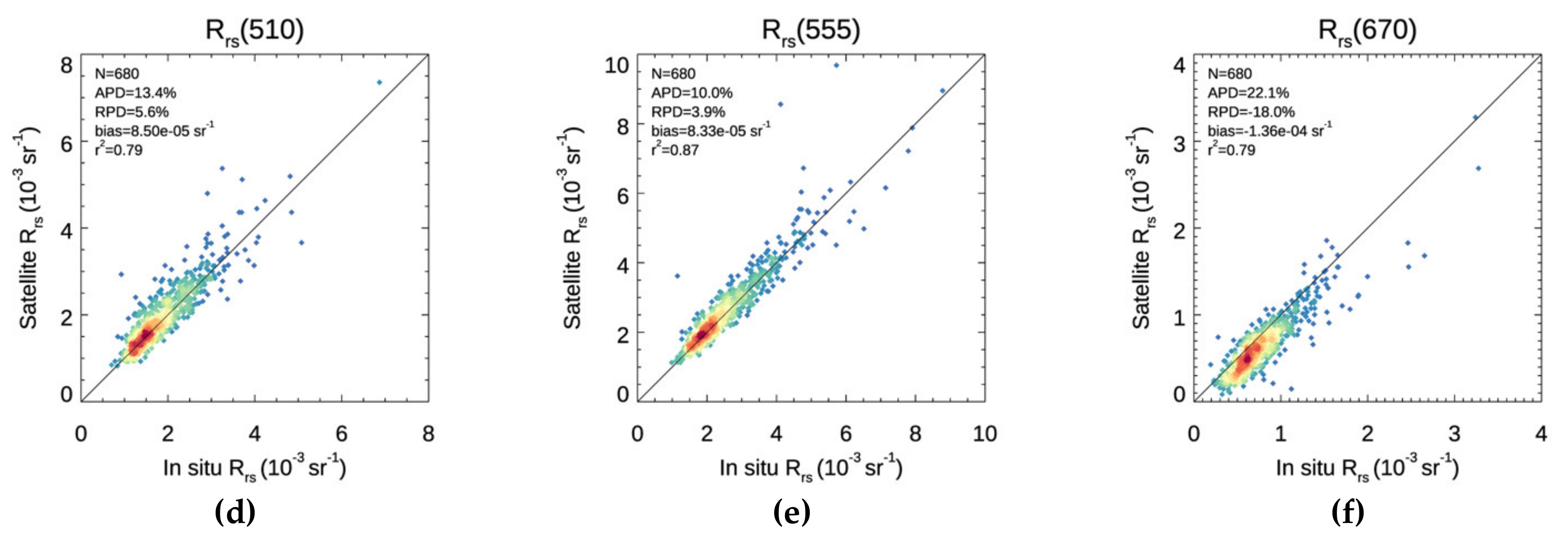
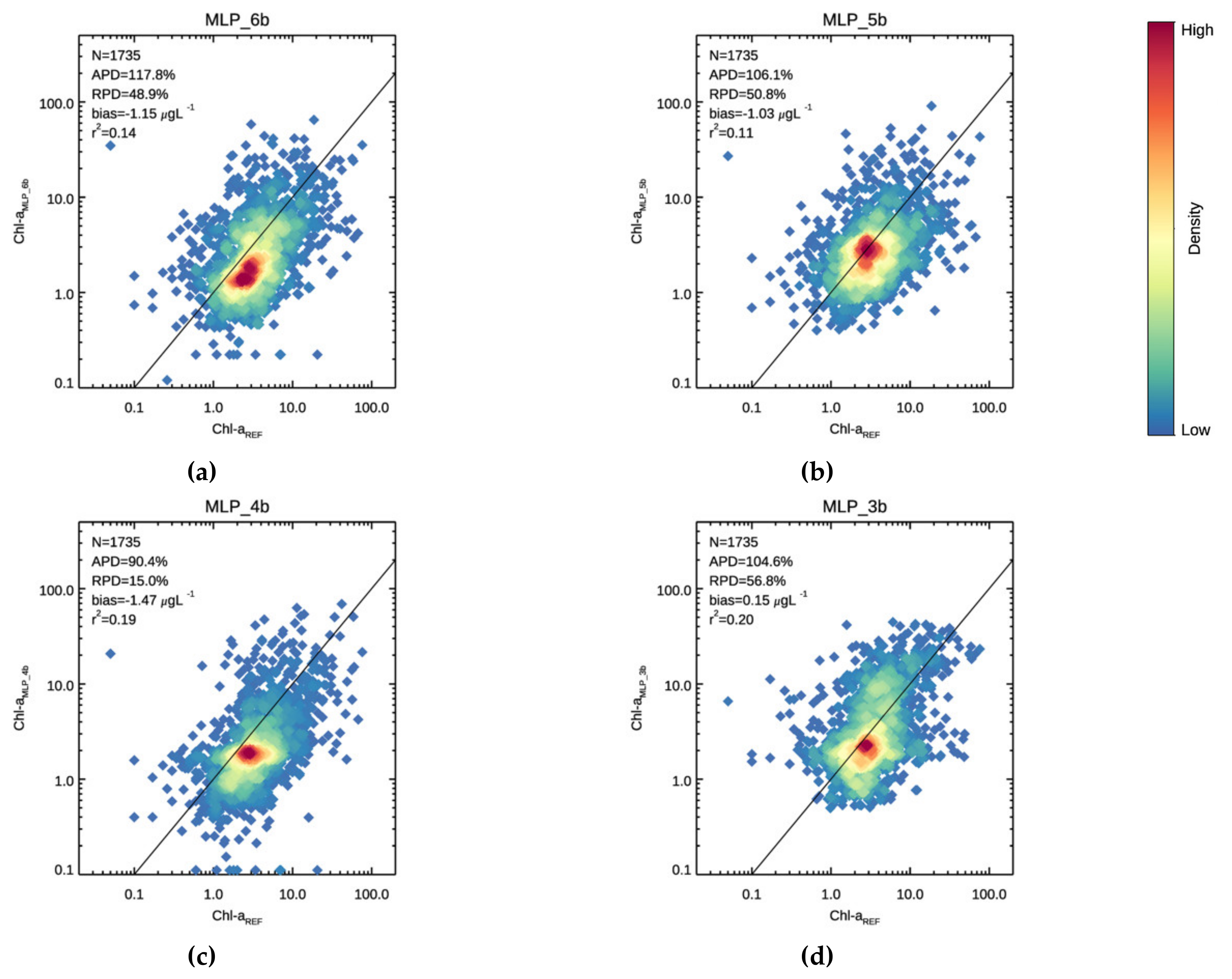
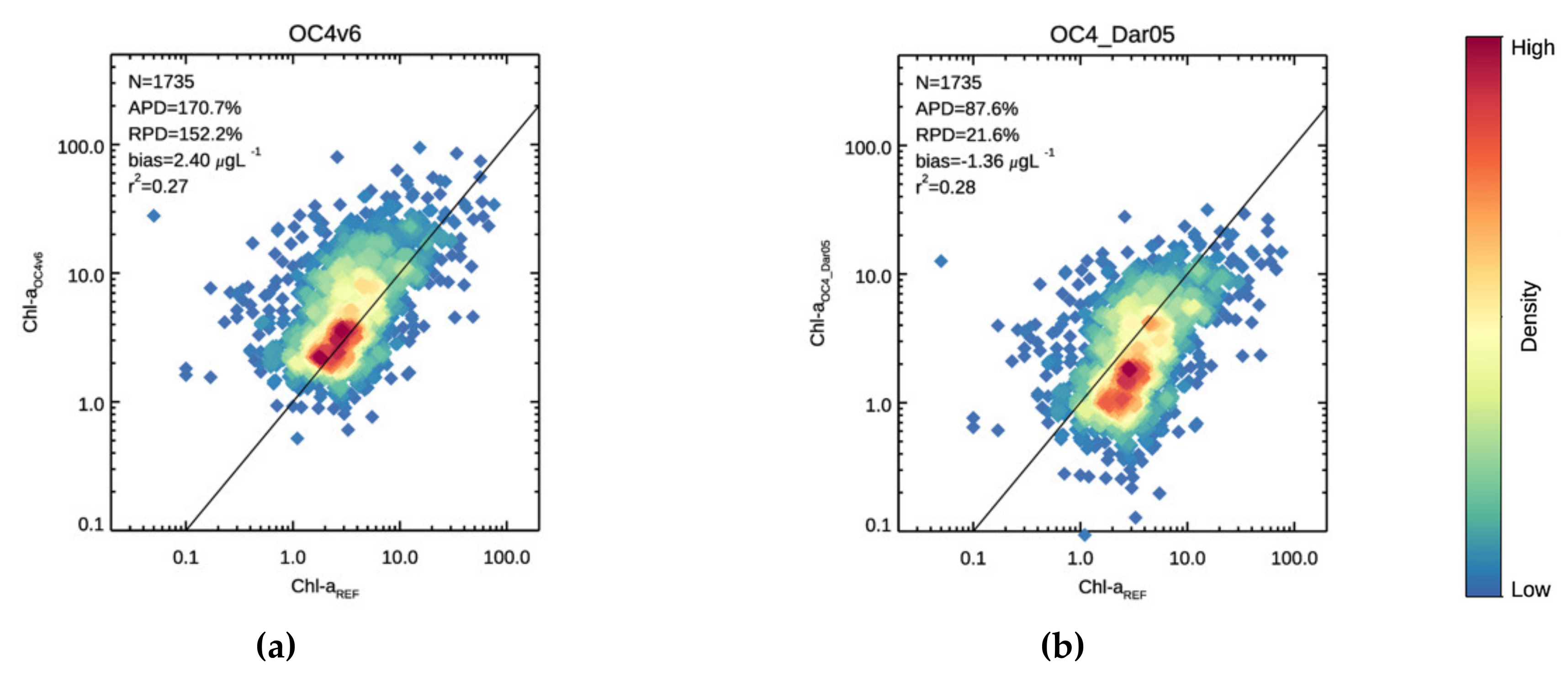
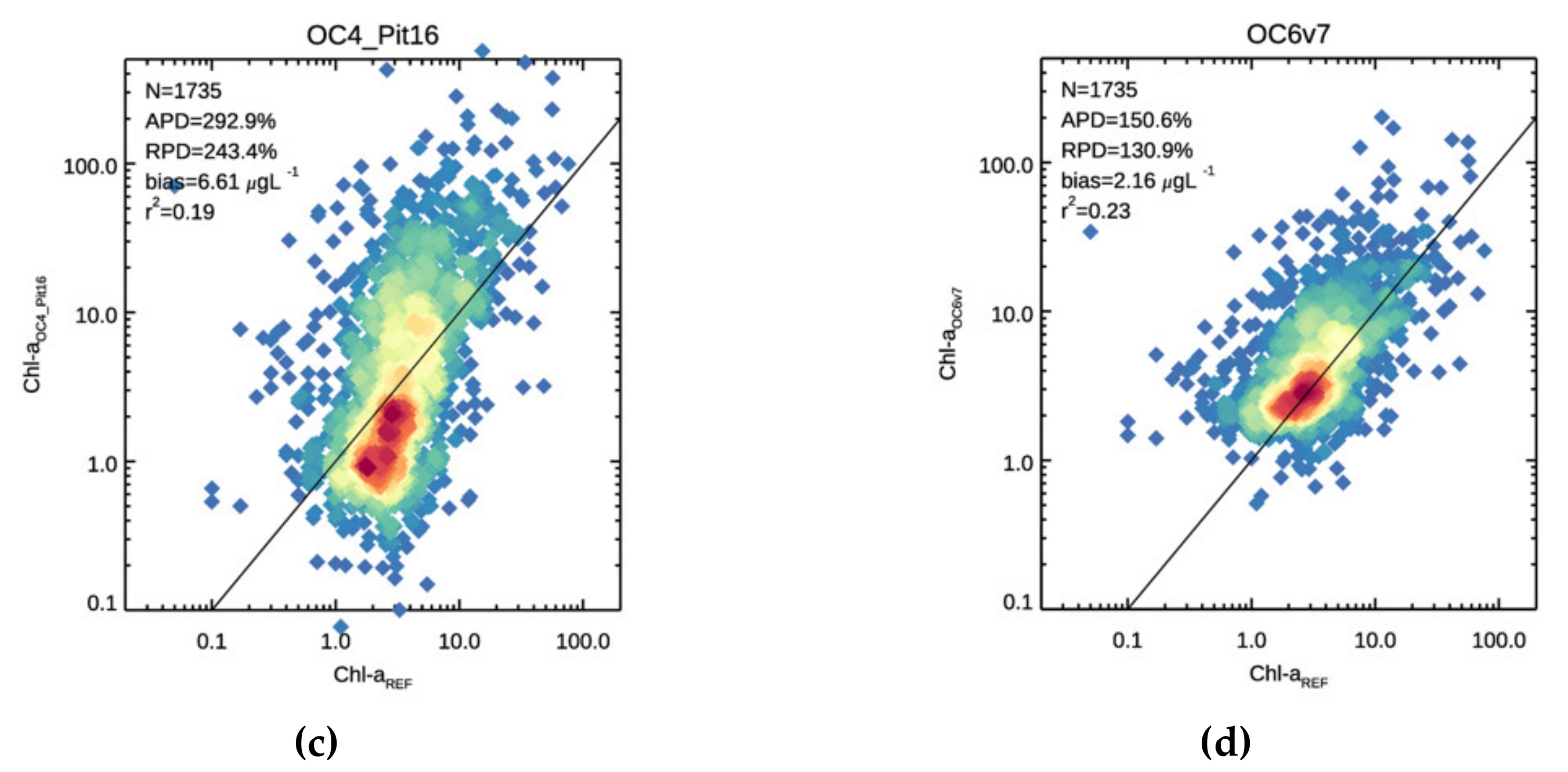
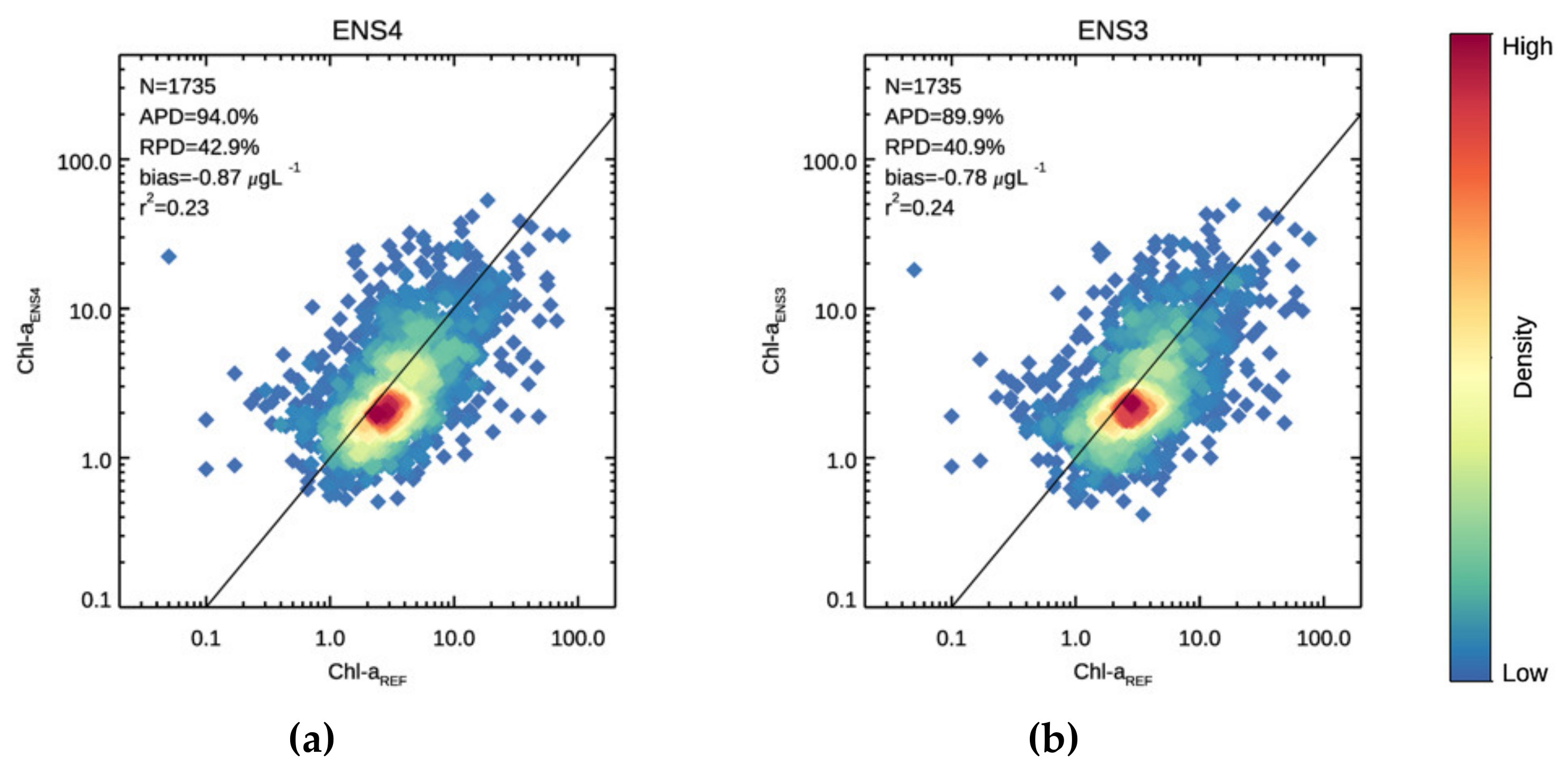
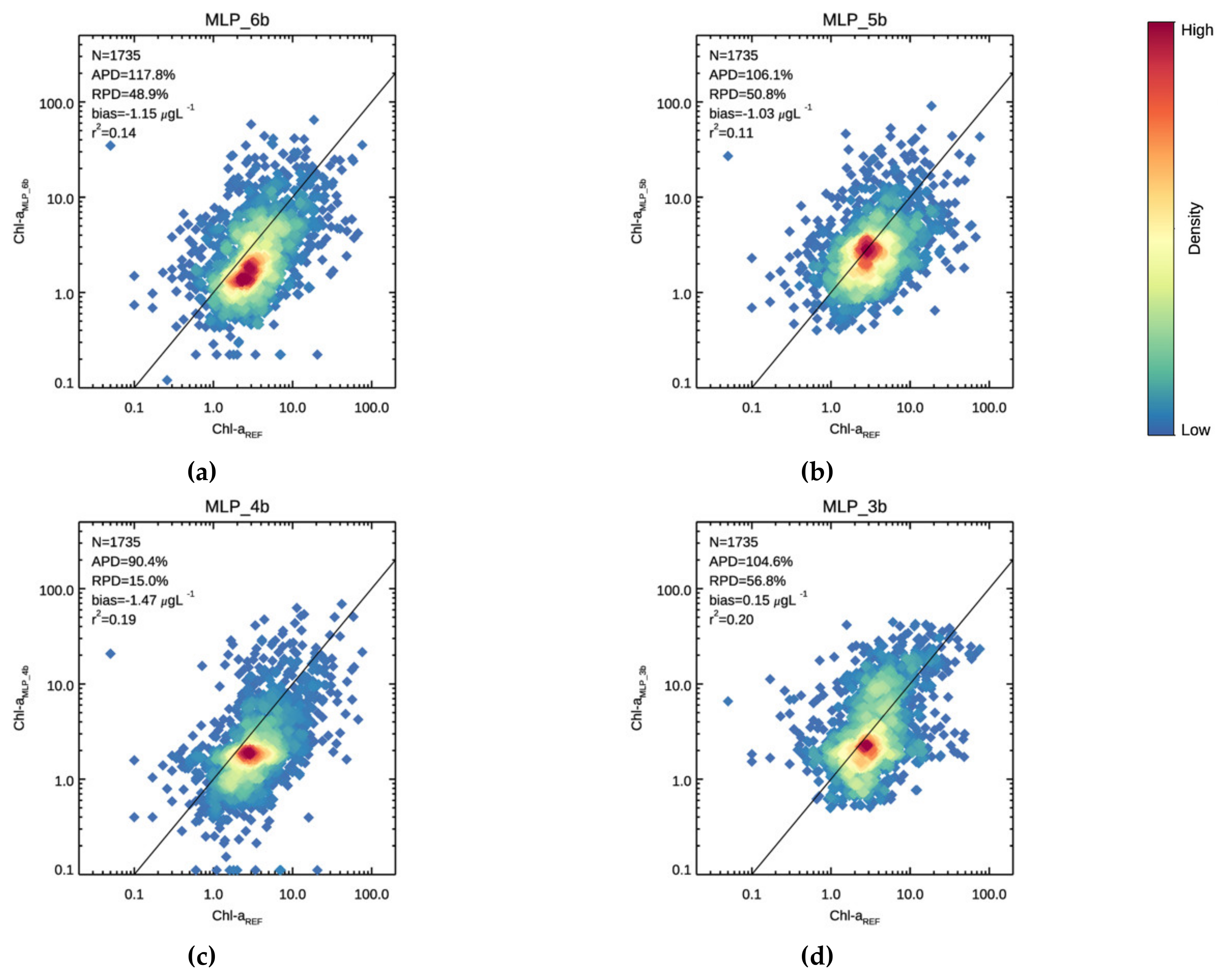
3.1. Validation
3.2. Chl-a Match-Ups with Alg@line Transects and COMBINE Dataset
3.3. Application to Environmental Reporting
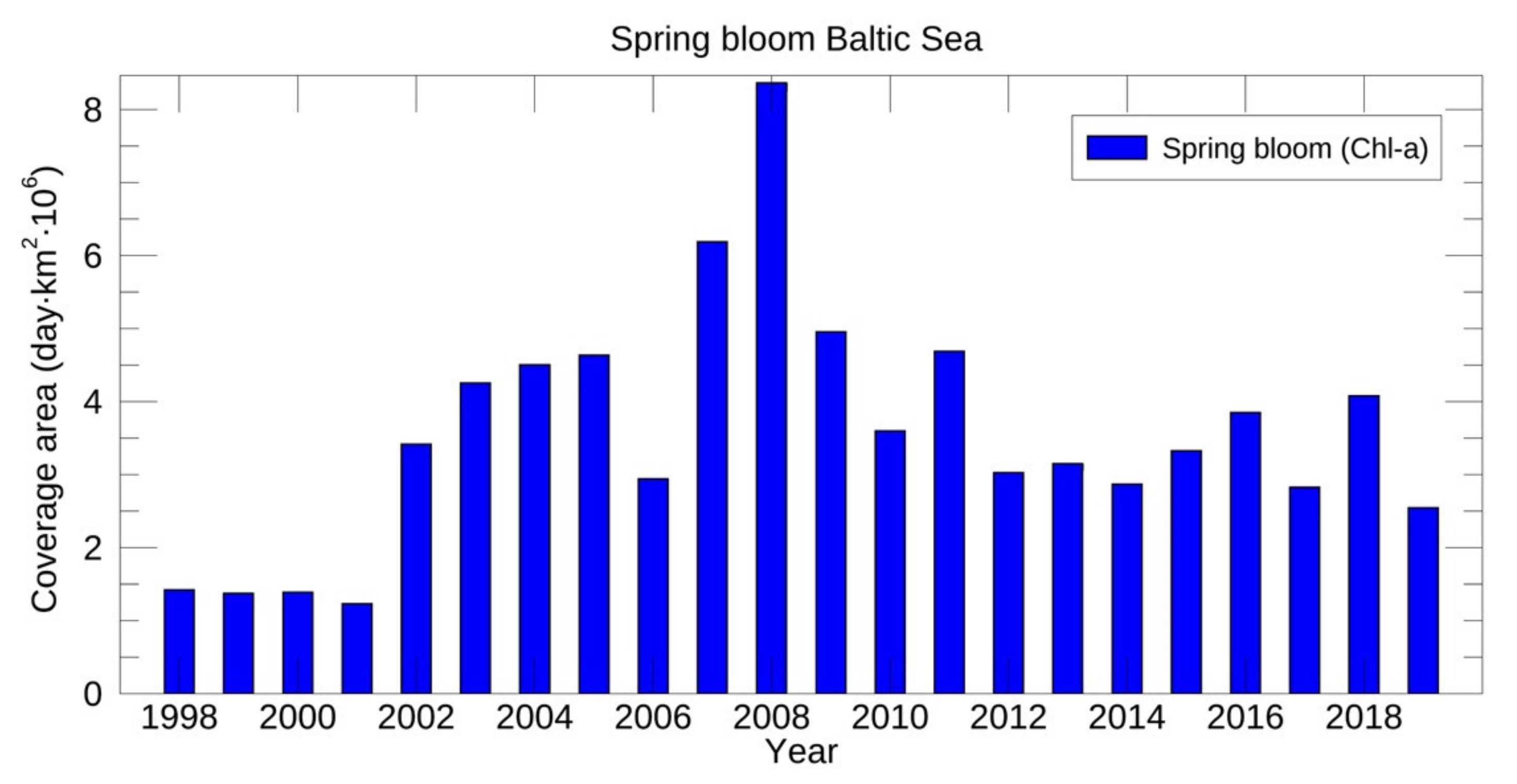
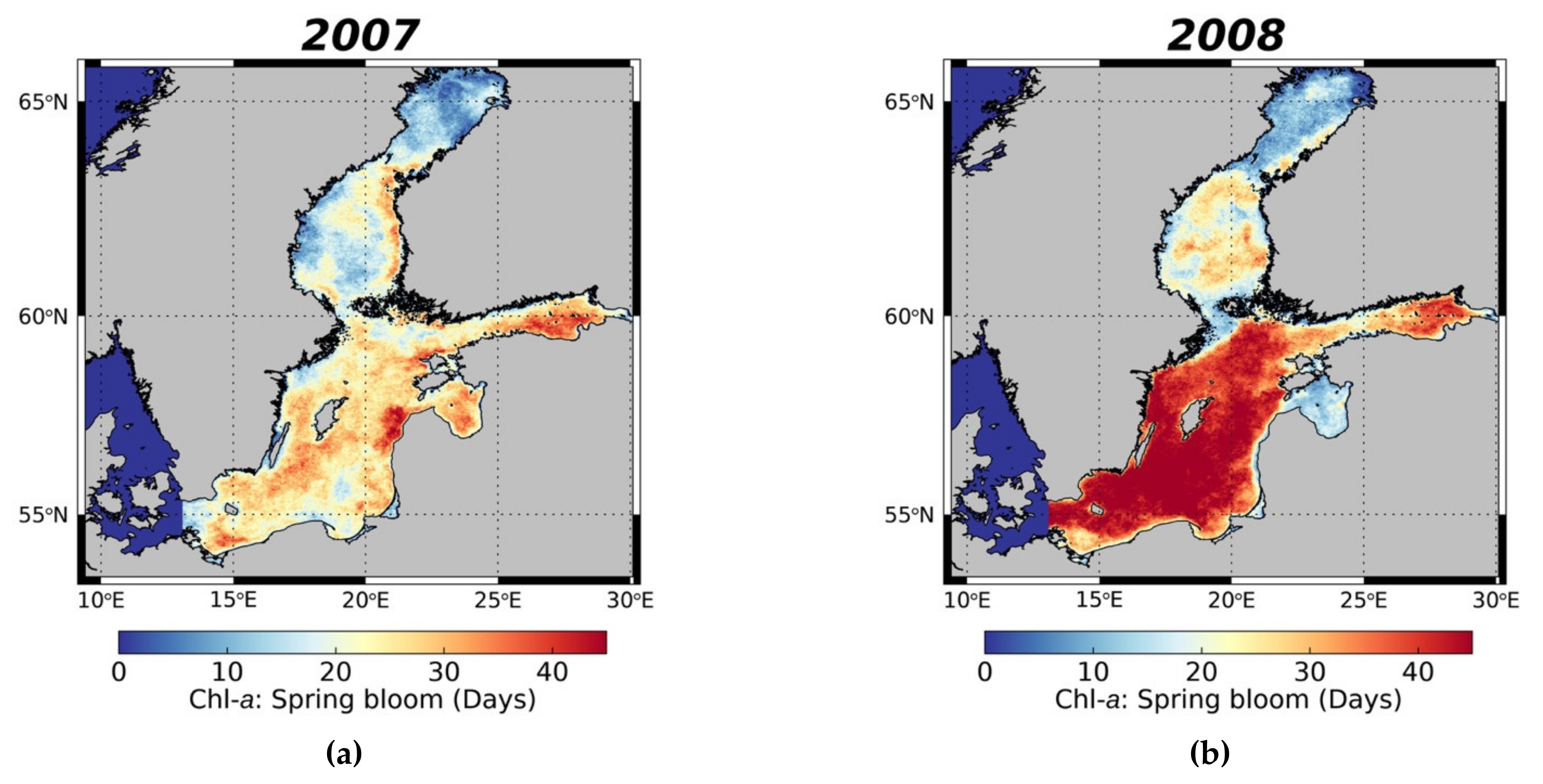
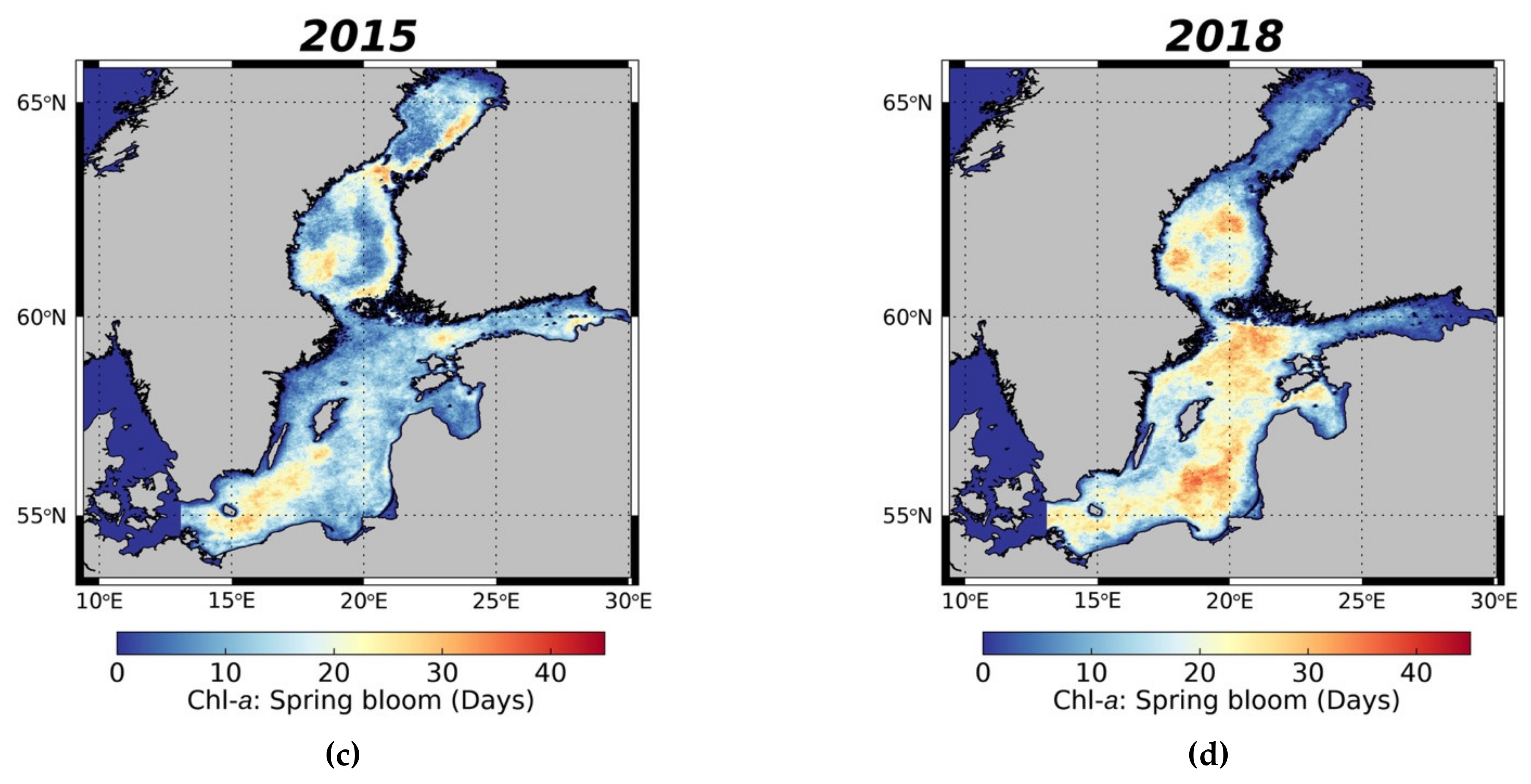
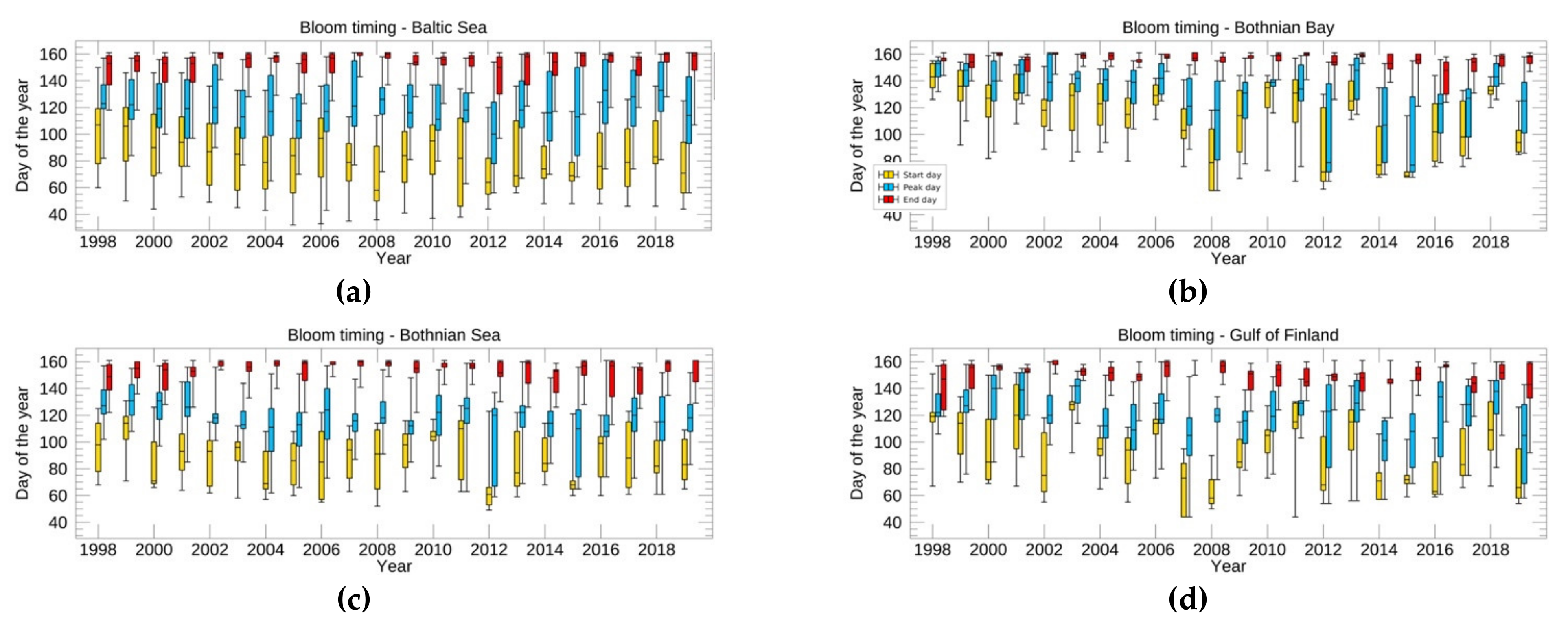
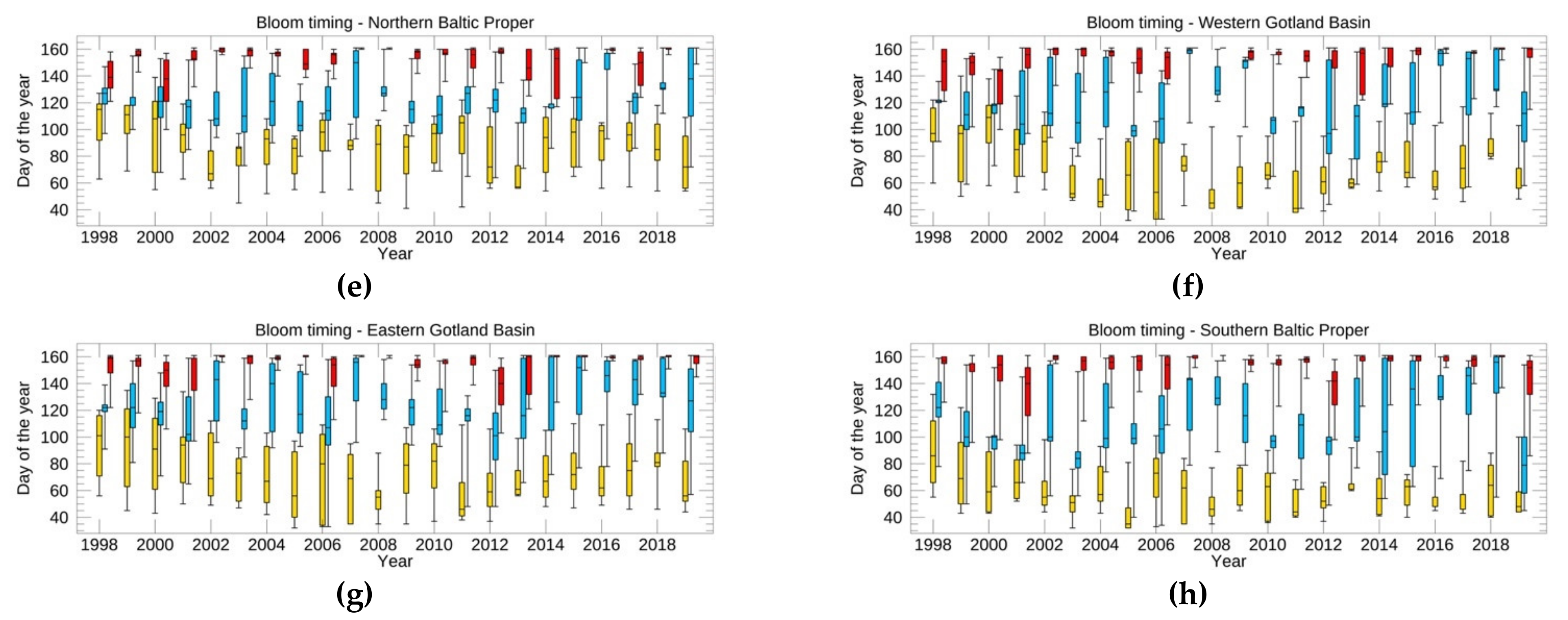
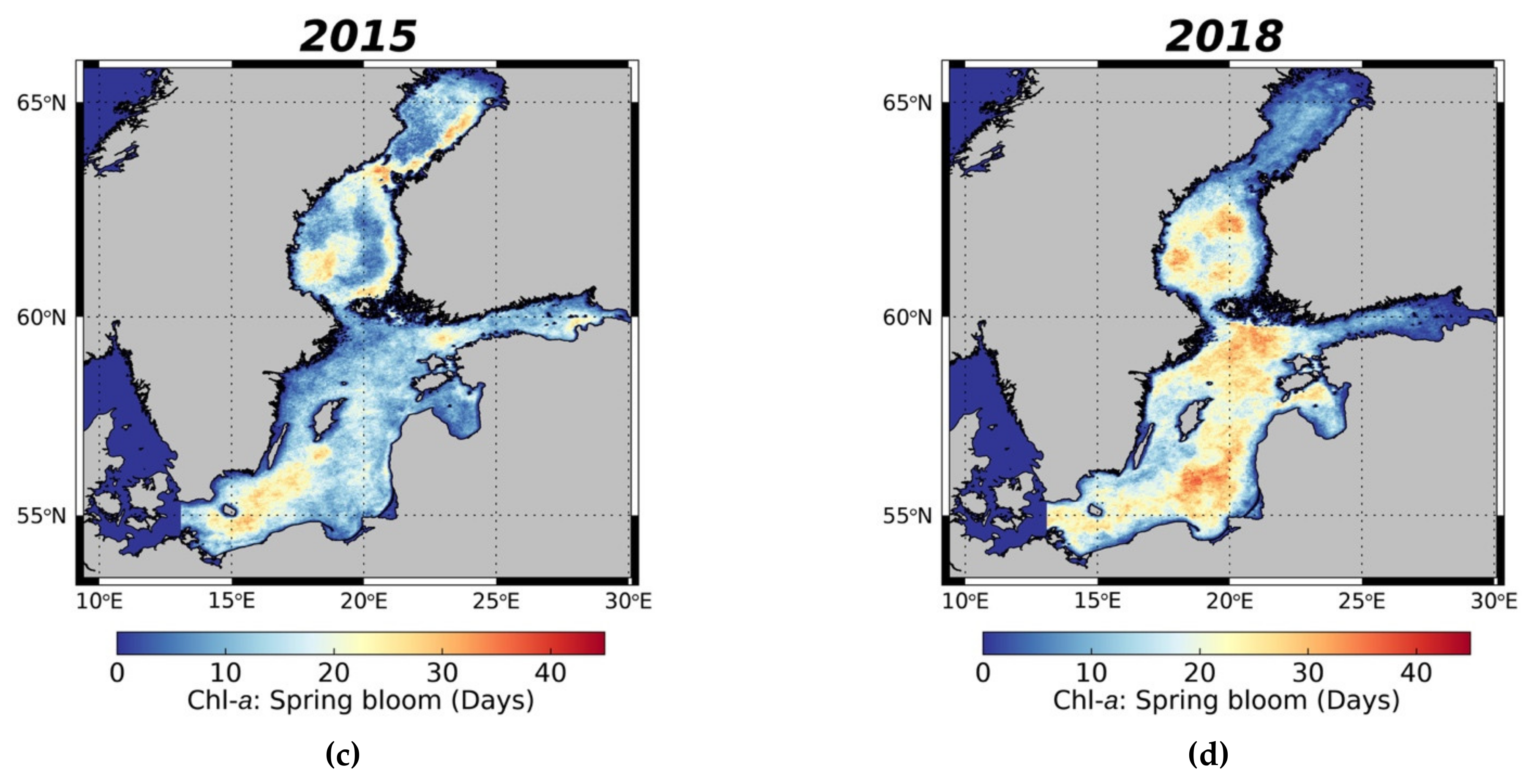
3.3.1. Spring Bloom Dynamics
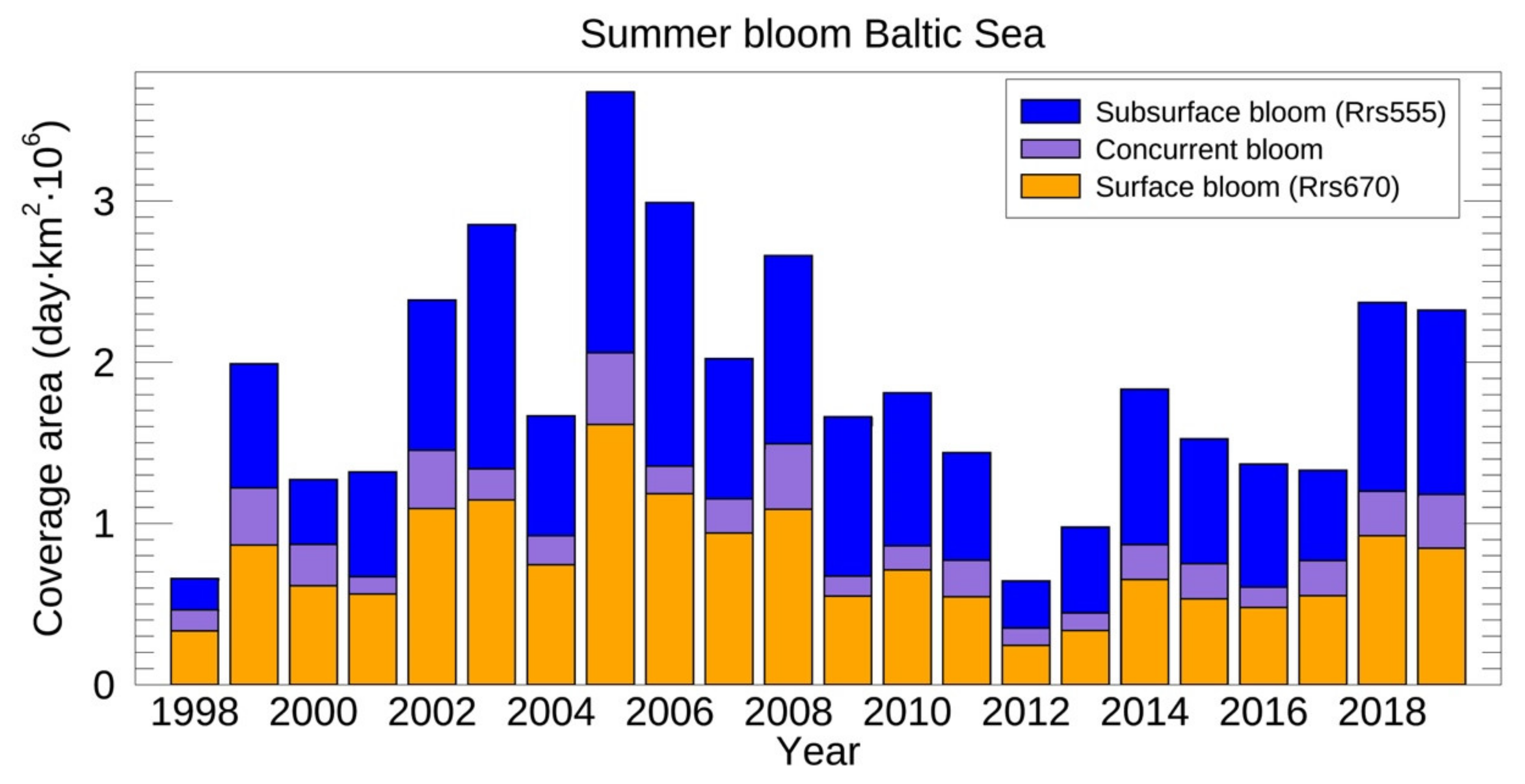
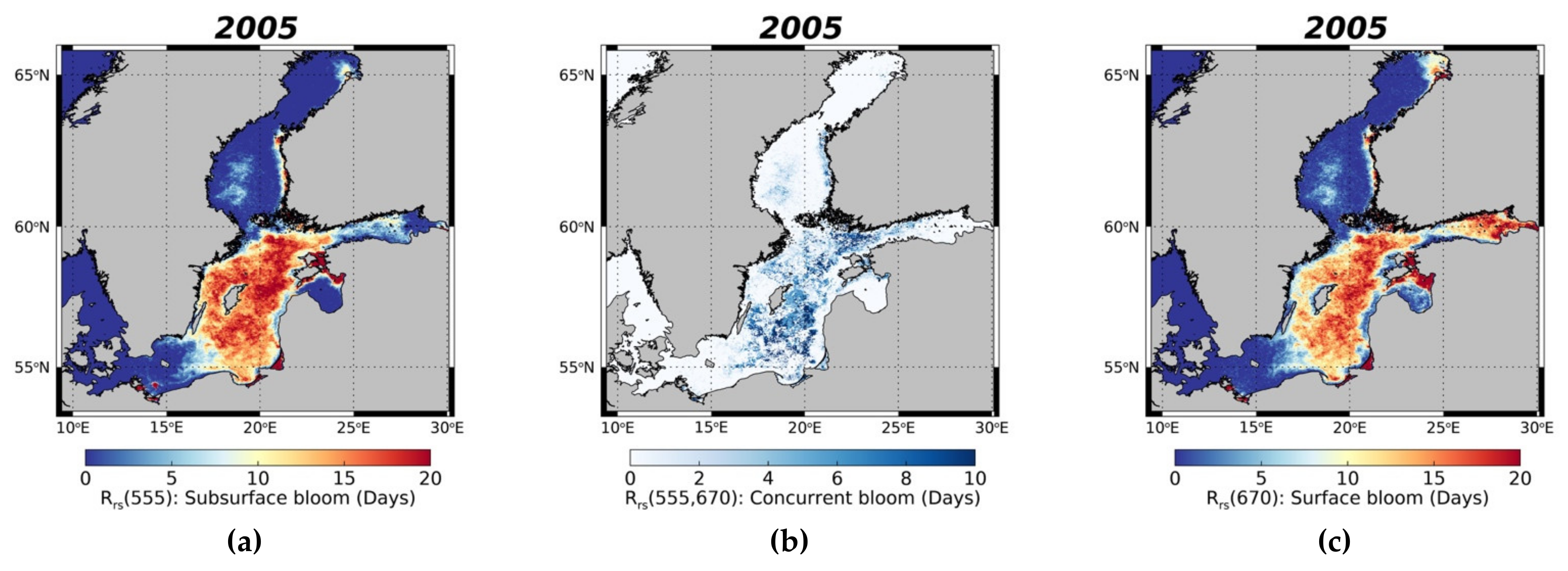
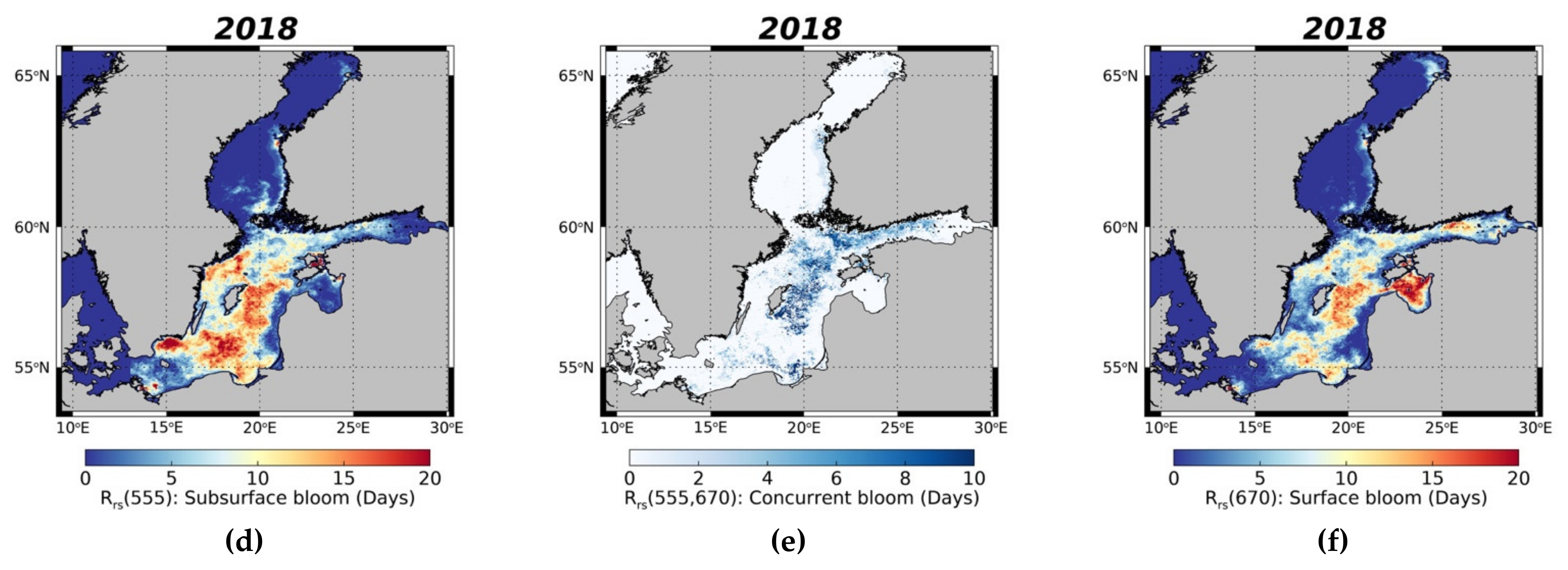
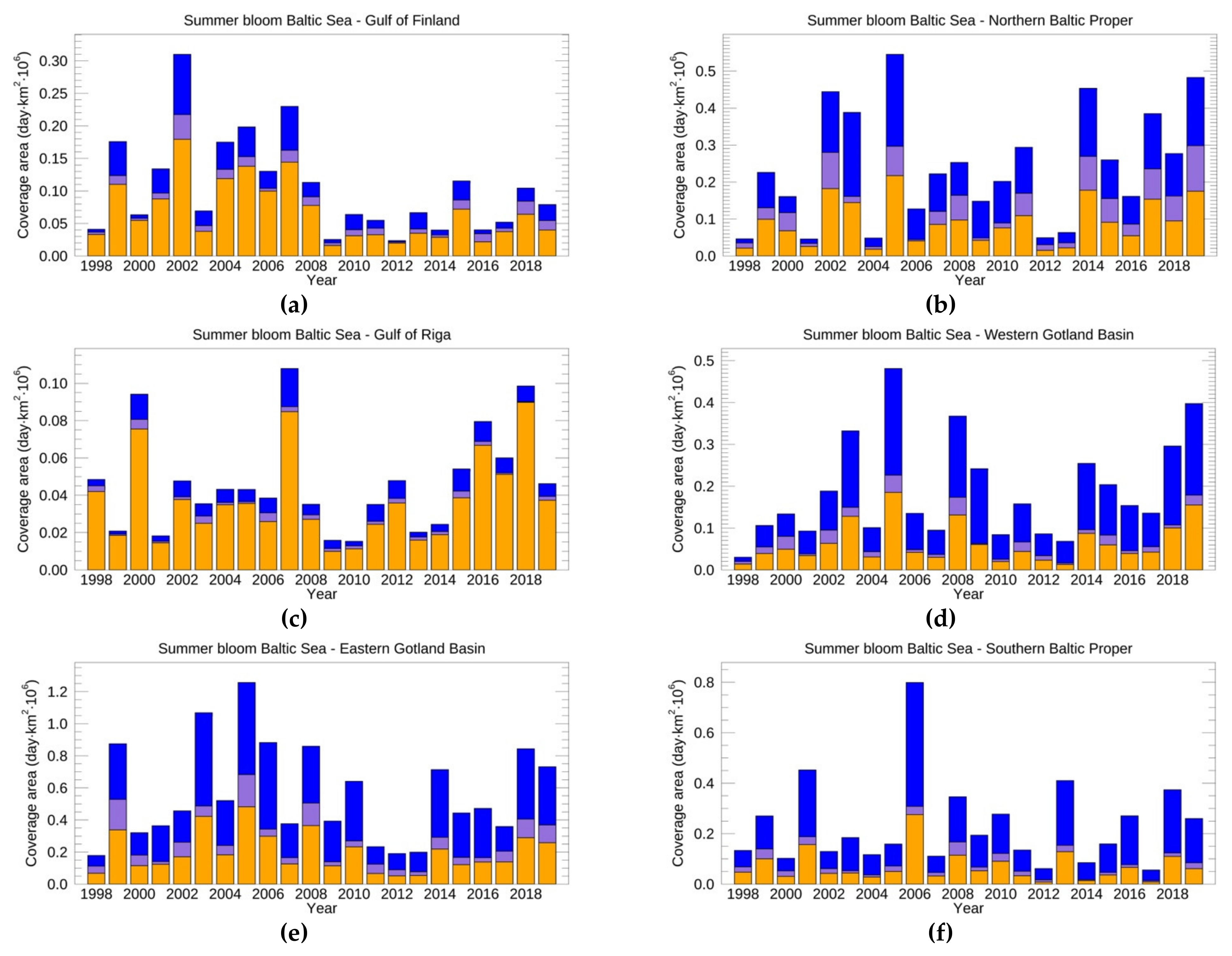
3.3.2. Surface and Subsurface Summer Blooms
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersen, J.H.; Axe, P.; Backer, H.; Carstensen, J.; Claussen, U.; Fleming-Lehtinen, V.; Järvinen, M.; Kaartokallio, H.; Knuuttila, S.; Korpinen, S.; et al. Getting the Measure of Eutrophication in the Baltic Sea: Towards Improved Assessment Principles and Methods. Biogeochemistry 2011, 106, 137–156. [Google Scholar] [CrossRef] [Green Version]
- Fleming-Lehtinen, V.; Andersen, J.H.; Carstensen, J.; Łysiak-Pastuszak, E.; Murray, C.; Pyhälä, M.; Laamanen, M. Recent developments in assessment methodology reveal that the baltic sea eutrophication problem is expanding. Ecol. Indic. 2015, 48, 380–388. [Google Scholar] [CrossRef] [Green Version]
- HELCOM. HELCOM Baltic Sea Action Plan; HELCOM: Helsinki, Finland, 2007. [Google Scholar]
- HELCOM. State of the Baltic Sea—Second HELCOM Holistic Assessment 2011–2016; HELCOM: Helsinki, Finland, 2018. [Google Scholar]
- Malone, T.C.; Newton, A. The globalization of cultural eutrophication in the coastal ocean: Causes and consequences. Front. Mar. Sci. 2020, 7, 670. [Google Scholar] [CrossRef]
- HELCOM. Manual for Marine Monitoring in the COMBINE Programme of HELCOM; HELCOM: Helsinki, Finland, 2017. [Google Scholar]
- HELCOM. HELCOM Guidelines for Monitoring of Chlorophyll a; HELCOM: Helsinki, Finland, 2019. [Google Scholar]
- Ahlman, M.; Alenius, P.; Attila, J.; Arnkil, A.; Arponen, H.; Below, A.; Blankett, P.; Bäck, A.; Cederberg, T.; Forsman, L.; et al. Seurantakäsikirja Suomen Merenhoitosuunnitelman Seurantaohjelmaan Vuosille 2020–2026 (Manual for Marine Monitoring in Finland 2020–2026).; Suomen Ympäristökeskus: Helsinki, Finland, 2020; ISBN 978-952-11-5340-2. [Google Scholar]
- Blondeau-Patissier, D.; Gower, J.F.R.; Dekker, A.G.; Phinn, S.R.; Brando, V.E. A review of ocean color remote sensing methods and statistical techniques for the detection, mapping and analysis of phytoplankton blooms in coastal and open oceans. Prog. Oceanogr. 2014, 123, 123–144. [Google Scholar] [CrossRef] [Green Version]
- Groom, S.; Sathyendranath, S.; Ban, Y.; Bernard, S.; Brewin, R.; Brotas, V.; Brockmann, C.; Chauhan, P.; Choi, J.; Chuprin, A.; et al. Satellite Ocean Colour: Current Status and Future Perspective. Front. Mar. Sci. 2019, 6, 485. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, J.E.; Werdell, P.J. Chlorophyll Algorithms for Ocean Color Sensors—OC4, OC5 & OC6. Remote. Sens. Environ. 2019, 229, 32–47. [Google Scholar] [CrossRef]
- Sathyendranath, S.; Brewin, R.; Brockmann, C.; Brotas, V.; Calton, B.; Chuprin, A.; Cipollini, P.; Couto, A.; Dingle, J.; Doerffer, R.; et al. An ocean-colour time series for use in climate studies: The experience of the ocean-colour climate change initiative (OC-CCI). Sensors 2019, 19, 4285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathyendranath, S.; Brewin, R.J.W.; Jackson, T.; Mélin, F.; Platt, T. Ocean-colour products for climate-change studies: What are their ideal characteristics? Remote. Sens. Environ. 2017, 203, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Szeto, M.; Werdell, P.J.; Moore, T.S.; Campbell, J.W. Are the world’s oceans optically different? J. Geophys. Res. 2011, 116, C00H04. [Google Scholar] [CrossRef] [Green Version]
- Volpe, G.; Colella, S.; Brando, V.E.; Forneris, V.; La Padula, F.; Di Cicco, A.; Sammartino, M.; Bracaglia, M.; Artuso, F.; Santoleri, R. Mediterranean Ocean Colour Level 3 Operational Multi-Sensor Processing. Ocean Sci. 2019, 15, 127–146. [Google Scholar] [CrossRef] [Green Version]
- Heiskanen, A.-S.; Bonsdorff, E.; Joas, M. Baltic Sea: A Recovering Future From Decades of Eutrophication. In Coasts and Estuaries; Elsevier: Amsterdam, The Netherlands, 2019; pp. 343–362. ISBN 978-0-12-814003-1. [Google Scholar]
- Kahru, M.; Horstmann, U.; Rud, O. Satellite detection of increased cyanobacteria blooms in the Baltic Sea: Natural fluctuation or ecosystem change? Ambio 1994, 23, 469–472. [Google Scholar]
- Leppäranta, M.; Myrberg, K. Physical Oceanography of the Baltic Sea; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-540-79702-9. [Google Scholar]
- Hjerne, O.; Hajdu, S.; Larsson, U.; Downing, A.S.; Winder, M. Climate driven changes in timing, composition and magnitude of the Baltic Sea Phytoplankton Spring Bloom. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Simis, S.G.H.; Ylöstalo, P.; Kallio, K.Y.; Spilling, K.; Kutser, T. Contrasting seasonality in optical-biogeochemical properties of the Baltic Sea. PLoS ONE 2017, 12, e0173357. [Google Scholar] [CrossRef]
- Zhang, D.; Lavender, S.; Muller, J.-P.; Walton, D.; Zou, X.; Shi, F. MERIS observations of phytoplankton phenology in the Baltic Sea. Sci. Total. Environ. 2018, 642, 447–462. [Google Scholar] [CrossRef]
- Finni, T.; Kononen, K.; Olsonen, R.; Wallström, K. The history of cyanobacterial blooms in the Baltic Sea. Ambio 2001, 30, 172–178. [Google Scholar] [CrossRef]
- Kahru, M.; Elmgren, R.; Di Lorenzo, E.; Savchuk, O. Unexplained interannual oscillations of cyanobacterial blooms in the Baltic Sea. Sci. Rep. 2018, 8, 6365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attila, J.; Koponen, S.; Kallio, K.; Lindfors, A.; Kaitala, S.; Ylöstalo, P. MERIS Case II Water Processor Comparison on Coastal Sites of the Northern Baltic Sea. Remote. Sens. Environ. 2013, 128, 138–149. [Google Scholar] [CrossRef]
- Darecki, M.; Stramski, D. An Evaluation of MODIS and SeaWiFS Bio-Optical Algorithms in the Baltic Sea. Remote. Sens. Environ. 2004, 89, 326–350. [Google Scholar] [CrossRef]
- Ligi, M.; Kutser, T.; Kallio, K.; Attila, J.; Koponen, S.; Paavel, B.; Soomets, T.; Reinart, A. Testing the performance of empirical remote sensing algorithms in the Baltic Sea waters with modelled and in situ reflectance data. Oceanologia 2017, 59, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Kratzer, S.; Moore, G. Inherent optical properties of the Baltic Sea in comparison to other seas and oceans. Remote. Sens. 2018, 10, 418. [Google Scholar] [CrossRef] [Green Version]
- Berthon, J.-F.; Zibordi, G. Optically black waters in the Northern Baltic Sea. Geophys. Res. Lett. 2010, 37. [Google Scholar] [CrossRef]
- Zibordi, G.; Berthon, J.-F.; Mélin, F.; D’Alimonte, D. Cross-Site Consistent in Situ Measurements for Satellite Ocean Color Applications: The BiOMaP Radiometric Dataset. Remote. Sens. Environ. 2011, 115, 2104–2115. [Google Scholar] [CrossRef]
- Aavaste, A.; Sipelgas, L.; Uiboupin, R.; Uudeberg, K. Impact of thermohaline conditions on vertical variability of optical properties in the Gulf of Finland (Baltic Sea): Implications for water quality remote sensing. Front. Mar. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Scheinin, M.; Asmala, E. Ubiquitous Patchiness in Chlorophyll a Concentration in Coastal Archipelago of Baltic Sea. Front. Mar. Sci. 2020, 7, 563. [Google Scholar] [CrossRef]
- Mélin, F.; Zibordi, G.; Berthon, J.-F.; Bailey, S.; Franz, B.; Voss, K.; Flora, S.; Grant, M. Assessment of MERIS Reflectance Data as Processed with SeaDAS over the European Seas. Opt. Express 2011, 19, 25657–25671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, P.; Simis, S.G.H.; Tilstone, G.H. Radiometric Validation of Atmospheric Correction for MERIS in the Baltic Sea Based on Continuous Observations from Ships and AERONET-OC. Remote. Sens. Environ. 2017, 200, 263–280. [Google Scholar] [CrossRef] [Green Version]
- Zibordi, G.; Melin, F.; Berthon, J.-F. A Regional Assessment of OLCI Data Products. IEEE Geosci. Remote Sens. Lett. 2018, 15, 1490–1494. [Google Scholar] [CrossRef]
- D’Alimonte, D.; Kajiyama, T.; Saptawijaya, A. Ocean color remote sensing of atypical marine optical cases. IEEE Trans. Geosci. Remote Sens. 2016, 54, 6574–6586. [Google Scholar] [CrossRef]
- Pitarch, J.; Volpe, G.; Colella, S.; Krasemann, H.; Santoleri, R. Remote Sensing of Chlorophyll in the Baltic Sea at Basin Scale from 1997 to 2012 Using Merged Multi-Sensor Data. Ocean Sci. 2016, 12, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Odermatt, D.; Gitelson, A.; Brando, V.E.; Schaepman, M. Review of constituent retrieval in optically deep and complex waters from satellite imagery. Remote Sens. Environ. 2012, 118, 116–126. [Google Scholar] [CrossRef] [Green Version]
- D’Alimonte, D.; Zibordi, G.; Berthon, J.-F.; Canuti, E.; Kajiyama, T. Performance and Applicability of Bio-Optical Algorithms in Different European Seas. Remote Sens. Environ. 2012, 124, 402–412. [Google Scholar] [CrossRef]
- Kratzer, S.; Vinterhav, C. Improvement of MERIS Level 2 Products in Baltic Sea Coastal Areas by Applying the Improved Contrast between Ocean and Land Processor (ICOL)—Data Analysis and Validation. Oceanologia 2010, 52, 211–236. [Google Scholar] [CrossRef]
- Kyryliuk, Dmytro; Kratzer, Susanne Evaluation of Sentinel-3A OLCI Products Derived Using the Case-2 Regional CoastColour Processor over the Baltic Sea. Sensors 2019, 19, 3609. [CrossRef] [Green Version]
- Hieronymi, M.; Müller, D.; Doerffer, R. The OLCI Neural Network Swarm (ONNS): A Bio-Geo-Optical Algorithm for Open Ocean and Coastal Waters. Front. Mar. Sci. 2017, 4, 140. [Google Scholar] [CrossRef] [Green Version]
- Toming, K.; Kutser, T.; Uiboupin, R.; Arikas, A.; Vahter, K.; Paavel, B. Mapping Water Quality Parameters with Sentinel-3 Ocean and Land Colour Instrument Imagery in the Baltic Sea. Remote Sens. 2017, 9, 1070. [Google Scholar] [CrossRef] [Green Version]
- Le Traon, P.Y.; Reppucci, A.; Alvarez Fanjul, E.; Aouf, L.; Behrens, A.; Belmonte, M.; Bentamy, A.; Bertino, L.; Brando, V.E.; Kreiner, M.B.; et al. From Observation to Information and Users: The Copernicus Marine Service Perspective. Front. Mar. Sci. 2019, 6, 234. [Google Scholar] [CrossRef] [Green Version]
- von Schuckmann, K.; Le Traon, P.-Y.; Smith, N.; Pascual, A.; Djavidnia, S.; Gattuso, J.-P.; Grégoire, M.; Nolan, G.; Aaboe, S.; Aguiar, E.; et al. Copernicus Marine Service Ocean State Report, Issue 3. J. Oper. Oceanogr. 2019, 12, S1–S123. [Google Scholar] [CrossRef]
- D’Alimonte, D.; Zibordi, G.; Berthon, J.-F.; Canuti, E.; Kajiyama, T. Bio-Optical Algorithms for European Seas: Performance and Applicability of Neural-Net Inversion Schemes; Publications Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Darecki, M.; Kaczmarek, S.; Olszewski, J. SeaWiFS Ocean Colour Chlorophyll Algorithms for the Southern Baltic Sea. Int. J. Remote Sens. 2005, 26, 247–260. [Google Scholar] [CrossRef]
- NASA Ocean Biology Processing Group. Chlorophyll a (Chlor_a) Product Summary; NASA: Washington, DC, USA, 2021.
- NASA Ocean Biology Processing Group. Visible and Infrared Imager/Radiometer Suite (VIIRS) Ocean Color Data; NASA: Washington, DC, USA, 2018.
- NASA Ocean Biology Processing Group. SEAWIFS-ORBVIEW-2 Level 2 Ocean Color Data Version R2018.0; NASA: Washington, DC, USA, 2018.
- NASA Ocean Biology Processing Group. Moderate-Resolution Imaging Spectroradiometer (MODIS) Aqua Ocean Color Data; NASA: Washington, DC, USA, 2018.
- NASA Ocean Biology Processing Group. VIIRS-SNPP Level 2 Ocean Color Data Version R2018.0; NASA: Washington, DC, USA, 2017.
- Steinmetz, F.; Deschamps, P.-Y.; Ramon, D. Atmospheric Correction in Presence of Sun Glint: Application to MERIS. Opt. Express 2011, 19, 9783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregg, W.W. Ocean-Colour Data Merging; IOCCG: Dartmouth, Canada, 2007; pp. 1–68. [Google Scholar]
- Lee, Z.; Carder, K.L.; Arnone, R.A. Deriving Inherent Optical Properties from Water Color: A Multiband Quasi-Analytical Algorithm for Optically Deep Waters. Appl. Opt. 2002, 41, 5755. [Google Scholar] [CrossRef] [PubMed]
- Lee, Z.; Carder, K.L.; Arnone, R.A. Update of the Quasi-Analytical Algorithm (QAA_V6); IOCCG: Dartmouth, Canada, 2014. [Google Scholar]
- Mélin, F.; Sclep, G. Band Shifting for Ocean Color Multi-Spectral Reflectance Data. Opt. Express 2015, 23, 2262. [Google Scholar] [CrossRef]
- D’Alimonte, D.; Zibordi, G.; Kajiyama, T.; Berthon, J.-F. Comparison between MERIS and Regional High-Level Products in European Seas. Remote Sens. Environ. 2014, 140, 378–395. [Google Scholar] [CrossRef]
- Kajiyama, T.; D’Alimonte, D.; Zibordi, G. Algorithms Merging for the Determination of Chlorophyll-a Concentration in the Black Sea. IEEE Geosci. Remote Sens. Lett. 2019, 16, 677–681. [Google Scholar] [CrossRef]
- Jacobs, R.A.; Jordan, M.I.; Nowlan, S.J.; Hinton, G.E. Adaptive mixtures of local experts. Neural Comput. 1991, 3, 79–87. [Google Scholar] [CrossRef]
- Yuksel, S.E.; Wilson, J.N.; Gader, P.D. Twenty Years of Mixture of Experts. IEEE Trans. Neural Netw. Learn. Syst. 2012, 23, 1177–1193. [Google Scholar] [CrossRef]
- Nabney, I. NETLAB: Algorithms for Pattern Recognitions; Springer: London, UK, 2002; ISBN 978-1-85233-440-6. [Google Scholar]
- Bishop, C.M. Neural Networks for Pattern Recognition; Oscar Publications: New Delhi, India, 2005; ISBN 978-0-19-566799-8. [Google Scholar]
- O’Reilly, J.E.; Maritorena, S.; Mitchell, B.G.; Siegel, D.A.; Carder, K.L.; Garver, S.A.; Kahru, M.; McClain, C. Ocean Color Chlorophyll Algorithms for SeaWiFS. J. Geophys. Res. 1998, 103, 24937–24953. [Google Scholar] [CrossRef] [Green Version]
- Zibordi, G.; Mélin, F.; Berthon, J.-F.; Holben, B.; Slutsker, I.; Giles, D.; D’Alimonte, D.; Vandemark, D.; Feng, H.; Schuster, G.; et al. AERONET-OC: A Network for the Validation of Ocean Color Primary Products. J. Atmos. Oceanic Technol. 2009, 26, 1634–1651. [Google Scholar] [CrossRef]
- Zibordi, G.; Holben, B.N.; Talone, M.; D’Alimonte, D.; Slutsker, I.; Giles, D.M.; Sorokin, M.G. Advances in the Ocean Color Component of the Aerosol Robotic Network (AERONET-OC). J. Atmos. Ocean. Technol. 2020, 1. [Google Scholar] [CrossRef]
- NASA AERONET Ocean Color. Available online: https://aeronet.gsfc.nasa.gov/new_web/ocean_color.html (accessed on 10 April 2020).
- Thuillier, G.; Floyd, L.; Woods, T.N.; Cebula, R.; Hilsenrath, E.; Hersé, M.; Labs, D. Solar irradiance reference spectra. In Geophysical Monograph Series; Pap, J.M., Fox, P., Frohlich, C., Hudson, H.S., Kuhn, J., McCormack, J., North, G., Sprigg, W., Wu, S.T., Eds.; American Geophysical Union: Washington, DC, USA, 2004; Volume 141, pp. 171–194. ISBN 978-0-87590-406-1. [Google Scholar]
- Fleming, V.; Kaitala, S. Phytoplankton Spring Bloom Intensity Index for the Baltic Sea Estimated for the Years 1992 to 2004. Hydrobiologia 2006, 554, 57–65. [Google Scholar] [CrossRef]
- Kaitala, S.; Zibordi, G.; Melin, F.; Seppälä, J.; Ylöstalo, P. Coastal Water Monitoring and Remote Sensing Products Validation Using Ferrybox and Above-Water Radiometric Measurements. EARSeL eProceedings 2008, 7, 75–80. [Google Scholar]
- Kahru, M.; Nommann, S. The Phytoplankton Spring Bloom in the Baltic Sea in 1985, 1986: Multitude of Spatio-Temporal Scales. Cont. Shelf Res. 1990, 10, 329–354. [Google Scholar] [CrossRef]
- Raudsepp, U.; She, J.; Brando, V.E.; Santoleri, R.; Sammartino, M.; Koõuts, M.; Uiboupin, R.; Maljutenko, I. Phytoplankton blooms in the Baltic Sea. In Copernicus Marine Service Ocean State Report. Issue 3. J. Oper. Oceanogr. 2019, 12, s21–s26. [Google Scholar]
- Raudsepp, U.; She, J.; Brando, V.; Kõuts, M.; Lagemaa, P.; Sammartino, M.; Santoleri, R. Eutrophication and hypoxia in the Baltic Sea. In Copernicus Marine Service Ocean State Report. Issue 2. J. Oper. Oceanogr. 2018, 11, s110–s116. [Google Scholar]
- Groetsch, P.M.M.; Simis, S.G.H.; Eleveld, M.A.; Peters, S.W.M. Spring Blooms in the Baltic Sea Have Weakened but Lengthened from 2000 to 2014. Biogeosciences 2016, 13, 4959–4973. [Google Scholar] [CrossRef] [Green Version]
- Siegel, D.A.; Doney, S.C.; Yoder, J.A. The North Atlantic Spring Phytoplankton Bloom and Sverdrup’s Critical Depth Hypothesis. Science 2002, 296, 730–733. [Google Scholar] [CrossRef]
- HELCOM. HELCOM Subbasins with Coastal and Offshore Division 2018 (ID: 4). Available online: https://maps.helcom.fi/arcgis/rest/services/MADS/Sea_environmental_monitoring/MapServer/4 (accessed on 10 April 2020).
- Hansson, M.; Pamberton, P.; Håkansson, B.; Reinart, A.; Alikas, K. Operational Nowcasting of Algal Blooms in the Baltic Sea Using MERIS and MODIS. In Proceedings of the ESA Living Planet Symposium, Bergen, Norway, 28 June–2 July 2010. [Google Scholar]
- Öberg, J. Cyanobacterial Blooms in the Baltic Sea. HELCOM Baltic Sea Environment Fact Sheet 2017; HELCOM: Helsinki, Finland, 2018. [Google Scholar]
- Kahru, M.; Savchuk, O.; Elmgren, R. Satellite measurements of cyanobacterial bloom frequency in the Baltic Sea: Interannual and spatial variability. Mar. Ecol. Prog. Ser. 2007, 343, 15–23. [Google Scholar] [CrossRef]
- Kutser, T. Quantitative Detection of Chlorophyll in Cyanobacterial Blooms by Satellite Remote Sensing. Limnol. Oceanogr. 2004, 49, 2179–2189. [Google Scholar] [CrossRef]
- Kutser, T.; Metsamaa, L.; Strömbeck, N.; Vahtmäe, E. Monitoring Cyanobacterial Blooms by Satellite Remote Sensing. Estuar. Coast. Shelf Sci. 2006, 67, 303–312. [Google Scholar] [CrossRef]
- Reinart, A.; Kutser, T. Comparison of Different Satellite Sensors in Detecting Cyanobacterial Bloom Events in the Baltic Sea. Remote Sens. Environ. 2006, 102, 74–85. [Google Scholar] [CrossRef]
- Hansson, M.; Håkansson, B. The Baltic Algae Watch System—A remote sensing application for monitoring cyanobacterial blooms in the Baltic Sea. J. Appl. Remote Sens 2007, 1, 011507. [Google Scholar] [CrossRef]
- Valente, A.; Sathyendranath, S.; Brotas, V.; Groom, S.; Grant, M.; Taberner, M.; Antoine, D.; Arnone, R.; Balch, W.M.; Barker, K.; et al. A Compilation of Global Bio-Optical in Situ Data for Ocean-Colour Satellite Applications—Version Two. Earth Syst. Sci. Data 2019, 11, 1037–1068. [Google Scholar] [CrossRef] [Green Version]
- Pitarch, J.; Bellacicco, M.; Marullo, S.; van der Woerd, H.J. Global Maps of Forel–Ule Index, Hue Angle and Secchi Disk Depth Derived from 21 Years of Monthly ESA Ocean Colour Climate Change Initiative Data. Earth Syst. Sci. Data 2021, 13, 481–490. [Google Scholar] [CrossRef]
- Kahru, M.; Elmgren, R. Multidecadal Time Series of Satellite-Detected Accumulations of Cyanobacteria in the Baltic Sea. Biogeosciences 2014, 11, 3619–3633. [Google Scholar] [CrossRef] [Green Version]
- Kahru, M.; Elmgren, R.; Kaiser, J.; Wasmund, N.; Savchuk, O. Cyanobacterial Blooms in the Baltic Sea: Correlations with environmental factors. Harmful Algae 2020, 92, 101739. [Google Scholar] [CrossRef]
- Uotila, P.; Vihma, T.; Haapala, J. Atmospheric and Oceanic Conditions and the Extremely Low Bothnian Bay Sea Ice Extent in 2014/2015. Geophys. Res. Lett. 2015, 42, 7740–7749. [Google Scholar] [CrossRef]
- Anttila, S.; Fleming-Lehtinen, V.; Attila, J.; Junttila, S.; Alasalmi, H.; Hällfors, H.; Kervinen, M.; Koponen, S. A novel earth observation based ecological indicator for cyanobacterial blooms. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 145–155. [Google Scholar] [CrossRef]
- Hansson, M. HELCOM Baltic Sea Environment Fact Sheet 2005. Cyanobacterial Blooms in the Baltic Sea; HELCOM: Helsinki, Finland, 2005. [Google Scholar]
- Moore, T.S.; Dowell, M.D.; Bradt, S.; Ruiz Verdu, A. An Optical Water Type Framework for Selecting and Blending Retrievals from Bio-Optical Algorithms in Lakes and Coastal Waters. Remote Sens. Environ. 2014, 143, 97–111. [Google Scholar] [CrossRef] [Green Version]
- Moore, T.S.; Campbell, J.W.; Dowell, M.D. A Class-Based Approach to Characterizing and Mapping the Uncertainty of the MODIS Ocean Chlorophyll Product. Remote Sens. Environ. 2009, 113, 2424–2430. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Average | Standard Deviation | Min | 5th Percentile | 25th Percentile | Median | 75th Percentile | 95th Percentile | Max | |
|---|---|---|---|---|---|---|---|---|---|---|
| Alg@line | 658 | 3.44 | 3.05 | 0.10 | 1.11 | 1.95 | 2.80 | 3.99 | 7.33 | 48.24 |
| COMBINE | 1077 | 5.73 | 7.29 | 0.05 | 0.80 | 2.13 | 3.40 | 6.16 | 18.10 | 76.10 |
| ALL | 1735 | 4.86 | 6.15 | 0.05 | 1.00 | 2.08 | 3.13 | 5.16 | 14.30 | 76.10 |
| ALL (N = 1735) | Alg@line (N = 658) | COMBINE (N = 1077) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| APD | RPD | Bias | r2 | APD | RPD | Bias | r2 | APD | RPD | Bias | r2 | |
| 170.6 | 152.2 | 2.40 | 0.275 | 51.4 | 24.0 | −0.081 | 0.114 | 243.5 | 230.5 | 3.92 | 0.266 | |
| 87.6 | 21.6 | −1.36 | 0.283 | 50.8 | −41.3 | −1.81 | 0.118 | 110.0 | 60.1 | −1.09 | 0.276 | |
| 292.8 | 243.4 | 6.61 | 0.194 | 55.6 | −27.5 | −1.21 | 0.084 | 437.9 | 408.9 | 11.39 | 0.188 | |
| 150.6 | 130.9 | 2.16 | 0.230 | 42.5 | 13.9 | −0.38 | 0.133 | 216.7 | 202.4 | 3.72 | 0.222 | |
| 117.8 | 48.9 | −1.15 | 0.137 | 50.6 | −37.1 | −1.74 | 0.116 | 158.8 | 101.4 | −0.79 | 0.117 | |
| 106.1 | 50.8 | −1.02 | 0.109 | 42.4 | −18.6 | −1.29 | 0.040 | 145.0 | 93.3 | −0.86 | 0.095 | |
| 90.4 | 15.0 | −1.46 | 0.190 | 49.6 | −42.1 | −1.87 | 0.067 | 115.4 | 49.9 | −1.22 | 0.180 | |
| 104.6 | 56.8 | 0.15 | 0.204 | 50.6 | −11.6 | −1.01 | 0.049 | 137.5 | 98.6 | 0.86 | 0.193 | |
| 94.0 | 42.9 | −0.87 | 0.229 | 40.8 | −27.4 | −1.48 | 0.099 | 126.4 | 85.8 | −0.50 | 0.216 | |
| 89.9 | 40.9 | −0.78 | 0.238 | 39.8 | −24.1 | −1.39 | 0.076 | 120.6 | 80.6 | −0.41 | 0.228 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brando, V.E.; Sammartino, M.; Colella, S.; Bracaglia, M.; Di Cicco, A.; D’Alimonte, D.; Kajiyama, T.; Kaitala, S.; Attila, J. Phytoplankton Bloom Dynamics in the Baltic Sea Using a Consistently Reprocessed Time Series of Multi-Sensor Reflectance and Novel Chlorophyll-a Retrievals. Remote Sens. 2021, 13, 3071. https://doi.org/10.3390/rs13163071
Brando VE, Sammartino M, Colella S, Bracaglia M, Di Cicco A, D’Alimonte D, Kajiyama T, Kaitala S, Attila J. Phytoplankton Bloom Dynamics in the Baltic Sea Using a Consistently Reprocessed Time Series of Multi-Sensor Reflectance and Novel Chlorophyll-a Retrievals. Remote Sensing. 2021; 13(16):3071. https://doi.org/10.3390/rs13163071
Chicago/Turabian StyleBrando, Vittorio E., Michela Sammartino, Simone Colella, Marco Bracaglia, Annalisa Di Cicco, Davide D’Alimonte, Tamito Kajiyama, Seppo Kaitala, and Jenni Attila. 2021. "Phytoplankton Bloom Dynamics in the Baltic Sea Using a Consistently Reprocessed Time Series of Multi-Sensor Reflectance and Novel Chlorophyll-a Retrievals" Remote Sensing 13, no. 16: 3071. https://doi.org/10.3390/rs13163071
APA StyleBrando, V. E., Sammartino, M., Colella, S., Bracaglia, M., Di Cicco, A., D’Alimonte, D., Kajiyama, T., Kaitala, S., & Attila, J. (2021). Phytoplankton Bloom Dynamics in the Baltic Sea Using a Consistently Reprocessed Time Series of Multi-Sensor Reflectance and Novel Chlorophyll-a Retrievals. Remote Sensing, 13(16), 3071. https://doi.org/10.3390/rs13163071