
Climatic Regulation of Vegetation Phenology in Protected Areas along Western South America

 ,
,  , , , ,
, , , ,  , ,
, ,  and
and
Abstract
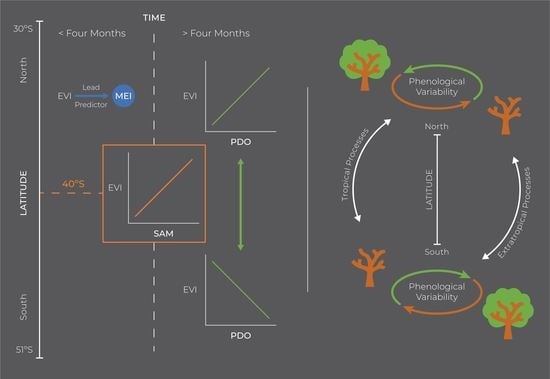
1. Introduction
2. Data and Methodological Analysis
2.1. Study Area
2.2. Phenology and Climatic Data
2.3. Data Analysis
3. Results
4. Discussion
5. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


References
- White, M.A.; Hoffman, F.; Hargrove, W.W.; Nemani, R.R. A global framework for monitoring phenological responses to climate change. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Donnelly, A.; Caffarra, A.; O’Neill, B.F. A review of climate-driven mismatches between interdependent phenophases in terrestrial and aquatic ecosystems. Int. J. Biometeorol. 2011, 55, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Renner, S.S.; Zohner, C.M. Climate change and phenological mismatch in trophic interactions among plants, insects, and vertebrates. Annu. Rev. Ecol. Syst. 2018, 49, 165–182. [Google Scholar] [CrossRef]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Inouye, D.W. Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology 2008, 89, 353–362. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Beard, K.H.; Kelsey, K.C.; Leffler, A.J.; Welker, J.M. The missing angle: Ecosystem consequences of phenological mismatch. Trends Ecol. Evol. 2019, 34, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Klos, R.J.; Wang, G.G.; Bauerle, W.L.; Rieck, J.R. Drought impact on forest growth and mortality in the southeast USA: An analysis using Forest Health and Monitoring data. Ecol. Appl. 2009, 19, 699–708. [Google Scholar] [CrossRef]
- Keenan, T.F.; Gray, J.; Friedl, M.A.; Toomey, M.; Bohrer, G.; Hollinger, D.Y.; Munger, J.W.; O’Keefe, J.; Schmid, H.P.; Wing, I.S.; et al. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat. Clim. Chang. 2014, 4, 598–604. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Ciais, P.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob. Chang. Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- Ma, Q.; Huang, J.G.; Hänninen, H.; Berninger, F. Reduced geographical variability in spring phenology of temperate trees with recent warming. Agric. For. Meteorol. 2018, 256–257, 526–533. [Google Scholar] [CrossRef]
- Yuan, H.; Wu, C.; Gu, C.; Wang, X. Evidence for satellite observed changes in the relative influence of climate indicators on autumn phenology over the Northern Hemisphere. Glob. Planet Chang. 2020, 187, 103131. [Google Scholar] [CrossRef]
- Maignan, F.; Bréon, F.M.; Bacour, C.; Demarty, J.; Poirson, A. Interannual vegetation phenology estimates from global AVHRR measurements: Comparison with in situ data and applications. Remote Sens. Environ. 2008, 112, 496–505. [Google Scholar] [CrossRef]
- Jamshidi, S.; Zand-Parsa, S.; Niyogi, D. Assessing Crop Water Stress Index of Citrus Using In-Situ Measurements, Landsat, and Sentinel-2 Data. Int. J. Remote Sens. 2021, 42, 1893–1916. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, S.; Feng, L.; Zhang, J.; Deng, F. Mapping Maize Cultivated Area Combining MODIS EVI Time Series and the Spatial Variations of Phenology over Huanghuaihai Plain. Appl. Sci. 2020, 10, 2667. [Google Scholar] [CrossRef]
- Ye, W.; van Dijk, A.I.; Huete, A.; Yebra, M. Global trends in vegetation seasonality in the GIMMS NDVI3g and their robustness. Int. J. Appl. Earth Obs. Geoinf. 2021, 94, 102238. [Google Scholar] [CrossRef]
- Kowalski, K.; Senf, C.; Hostert, P.; Pflugmacher, D. Characterizing spring phenology of temperate broadleaf forests using Landsat and Sentinel-2 time series. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102172. [Google Scholar] [CrossRef]
- Sarvia, F.; De Petris, S.; Borgogno-Mondino, E. Exploring Climate Change Effects on Vegetation Phenology by MOD13Q1 Data: The Piemonte Region Case Study in the Period 2001–2019. Agronomy 2021, 11, 555. [Google Scholar] [CrossRef]
- Gore, M.; Abiodun, B.J.; Kucharski, F. Understanding the influence of ENSO patterns on drought over southern Africa using SPEEDY. Clim. Dyn. 2020, 54, 307–327. [Google Scholar] [CrossRef]
- Sarricolea, P.; Serrano-Notivoli, R.; Fuentealba, M.; Hernández-Mora, M.; de la Barrera, F.; Smith, P.; Meseguer-Ruiz, Ó. Recent wildfires in Central Chile: Detecting links between burned areas and population exposure in the wildland urban interface. Sci. Total Environ. 2020, 706, 135894. [Google Scholar] [CrossRef]
- Woodward, F.; Lomas, M.; Quaife, T. Global responses of terrestrial productivity to contemporary climatic oscillations. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 2779–2785. [Google Scholar] [CrossRef]
- Ito, A. Decadal variability in the terrestrial carbon budget caused by the Pacific Decadal Oscillation and Atlantic Multidecadal Oscillation. J. Meteorol. Soc. Ser. II 2011, 89, 441–454. [Google Scholar] [CrossRef]
- Dätwyler, C.; Grosjean, M.; Steiger, N.J.; Neukom, R. Teleconnections and relationship between the El Niño–Southern Oscillation (ENSO) and the Southern Annular Mode (SAM) in reconstructions and models over the past millennium. Clim. Past 2020, 16, 743–756. [Google Scholar] [CrossRef]
- Zhao, L.; Dai, A.; Dong, B. Changes in global vegetation activity and its driving factors during 1982–2013. Agric. For. Meteorol. 2018, 249, 198–209. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Maiorano, L. Contrasting effects of temperature and precipitation change on amphibian phenology, abundance and performance. Oecologia 2016, 181, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fu, Y.H.; Zeng, Z.; Huang, M.; Li, X.; Piao, S. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China. Glob. Chang. Biol. 2016, 22, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Jamshidi, S.; Zand-Parsa, S.; Niyogi, D. Physiological responses of orange trees subject to regulated deficit irrigation and partial root drying. Irrig. Sci. 2021, 39, 441–455. [Google Scholar] [CrossRef]
- Gordo, O.; Sanz, J.J. Impact of climate change on plant phenology in Mediterranean ecosystems. Glob. Chang. Biol. 2010, 16, 1082–1106. [Google Scholar] [CrossRef]
- Zhang, X.; Hodges, J.C.; Schaaf, C.B.; Friedl, M.A.; Strahler, A.H.; Gao, F. Global vegetation phenology from AVHRR and MODIS data. In Proceedings of the IGARSS 2001, Scanning the Present and Resolving the Future, IEEE 2001 International Geoscience and Remote Sensing Symposium (Cat. No. 01CH37217), Sydney, Australia, 9–13 July 2001; Volume 5, pp. 2262–2264. [Google Scholar]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Bhattacharjee, A.; Anadón, J.D.; Lohman, D.J.; Doleck, T.; Lakhankar, T.; Shrestha, B.B.; Thapa, P.; Devkota, D.; Tiwari, S.; Jha, A.; et al. The impact of climate change on biodiversity in Nepal: Current knowledge, lacunae, and opportunities. Climate 2017, 5, 80. [Google Scholar] [CrossRef]
- Pliscoff, P.; Fuentes-Castillo, T. Representativeness of terrestrial ecosystems in Chile’s protected area system. Environ. Conserv. 2011, 38, 303–311. [Google Scholar] [CrossRef]
- Jaksic, F.M. Ecological effects of El Niño in terrestrial ecosystems of western South America. Ecography 2001, 24, 241–250. [Google Scholar] [CrossRef]
- Squeo, F.A.; Loayza, A.P.; López, R.P.; Gutiérrez, J.R. Vegetation of Bosque Fray Jorge National Park and its surrounding matrix in the Coastal Desert of north-central Chile. J. Arid Environ. 2016, 126, 12–22. [Google Scholar] [CrossRef]
- Holmgren, M.; Scheffer, M.; Ezcurra, E.; Gutiérrez, J.R.; Mohren, G.M. El Niño effects on the dynamics of terrestrial ecosystems. Trends Ecol. Evol. 2001, 16, 89–94. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Alvarez-Garreton, C.; Barichivich, J.; Boisier, J.P.; Christie, D.; Galleguillos, M.; LeQuesne, C.; McPhee, J.; Zambrano-Bigiarini, M. The 2010–2015 megadrought in central Chile: Impacts on regional hydroclimate and vegetation. Hydrol. Earth Syst. Sci. 2017, 21, 6307–6327. [Google Scholar] [CrossRef]
- Úbeda, X.; Sarricolea, P. Wildfires in Chile: A review. Glob. Planet Change 2016, 146, 152–161. [Google Scholar] [CrossRef]
- Urrutia-Jalabert, R.; González, M.E.; González-Reyes, Á.; Lara, A.; Garreaud, R. Climate variability and forest fires in central and south-central Chile. Ecosphere 2018, 9, e02171. [Google Scholar] [CrossRef]
- Holz, A.; Paritsis, J.; Mundo, I.A.; Veblen, T.T.; Kitzberger, T.; Williamson, G.J.; Aráoz, E.; Bustos-Schindler, C.; González, M.E.; Grau, H.R.; et al. Southern Annular Mode drives multicentury wildfire activity in southern South America. Proc. Natl. Acad. Sci. USA 2017, 114, 9552–9557. [Google Scholar] [CrossRef]
- Viale, M.; Garreaud, R. Orographic effects of the subtropical and extratropical Andes on upwind precipitating clouds. J. Geophys. Res. Atmos. 2015, 120, 4962–4974. [Google Scholar] [CrossRef]
- Boisier, J.P.; Rondanelli, R.; Garreaud, R.D.; Muñoz, F. Anthropogenic and natural contributions to the Southeast Pacific precipitation decline and recent megadrought in central Chile. Geophys. Res. Lett. 2016, 43, 413–421. [Google Scholar] [CrossRef]
- Quintana, J.; Aceituno, P. Changes in the rainfall regime along the extratropical west coast of South America (Chile): 30–43 ºS. Atmósfera 2012, 25, 1–22. [Google Scholar]
- Alaniz, A.J. Chilean forest: Recent trends and perspectives. Persp. Chall. 2019, 1–308. [Google Scholar]
- Madariaga, G.C.; Navarro, R.C.; Baeza, Á.P.; Verrugio, G.C. Structural and biometric characterization of Nothofagus betuloides production forests in the Magellan Region, Chile. J. Sustain. For. 2007, 24, 123–140. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853. [Google Scholar] [CrossRef]
- Díaz, I.A.; Armesto, J.J.; Reid, S.; Sieving, K.E.; Willson, M.F. Linking forest structure and composition: Avian diversity in successional forests of Chiloé Island, Chile. Biol. Conserv. 2005, 123, 91–101. [Google Scholar] [CrossRef]
- Echeverría, C.; Newton, A.C.; Lara, A.; Benayas, J.M.R.; Coomes, D.A. Impacts of forest fragmentation on species composition and forest structure in the temperate landscape of Southern Chile. Glob. Ecol. Biogeogr. 2007, 16, 426–439. [Google Scholar] [CrossRef]
- Petit, I.J.; Campoy, A.N.; Hevia, M.J.; Gaymer, C.F.; Squeo, F.A. Protected areas in Chile: Are we managing them? Rev. Chil. Hist. Nat. 2018, 91, 1. [Google Scholar] [CrossRef]
- Jusys, T. Quantifying avoided deforestation in Pará: Protected areas, buffer zones and edge effects. J. Nat. Conserv. 2016, 33, 10–17. [Google Scholar] [CrossRef]
- Núñez, J.; Rivera, D.; Oyarzún, R.; Arumí, J. Influence of Pacific Ocean multidecadal variability on the distributional properties of hydrological variables in north-central Chile. J. Hydrol. 2013, 501, 227–240. [Google Scholar] [CrossRef]
- Saldías, G.S.; Largier, J.L.; Mendes, R.; Pérez-Santos, I.; Vargas, C.A.; Sobarzo, M. Satellite-measured interannual variability of turbid river plumes off central-southern Chile: Spatial patterns and the influence of climate variability. Prog. Oceanogr. 2016, 146, 212–222. [Google Scholar] [CrossRef]
- Cazelles, B. Symbolic dynamics for identifying similarity between rhythms of ecological time series. Ecol. Lett. 2004, 7, 755–763. [Google Scholar] [CrossRef]
- Cazelles, B.; Chavez, M.; Magny, G.C.d.; Guégan, J.F.; Hales, S. Time-dependent spectral analysis of epidemiological time-series with wavelets. J. R. Soc. Interface 2007, 4, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, C.; Xing, H. The Analysis of Time Series: An Introduction with R; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2019. [Google Scholar]
- Cazelles, B.; Chavez, M.; Berteaux, D.; Ménard, F.; Vik, J.O.; Jenouvrier, S.; Stenseth, N.C. Wavelet analysis of ecological time series. Oecologia 2008, 156, 287–304. [Google Scholar] [CrossRef]
- Cazelles, B.; Cazelles, K.; Chavez, M. Wavelet analysis in ecology and epidemiology: Impact of statistical tests. J. R. Soc. Interface 2014, 11, 20130585. [Google Scholar] [CrossRef]
- Morisette, J.T.; Richardson, A.D.; Knapp, A.K.; Fisher, J.I.; Graham, E.A.; Abatzoglou, J.; Wilson, B.E.; Breshears, D.D.; Henebry, G.M.; Hanes, J.M.; et al. Tracking the rhythm of the seasons in the face of global change: Phenological research in the 21st century. Front. Ecol. Environ. 2009, 7, 253–260. [Google Scholar] [CrossRef]
- Taig-Johnston, M.; Strom, M.K.; Calhoun, K.; Nowak, K.; Ebensperger, L.A.; Hayes, L. The ecological value of long-term studies of birds and mammals in Central America, South America and Antarctica. Rev. Chil. Hist. Nat. 2017, 90, 1–13. [Google Scholar] [CrossRef]
- Buermann, W.; Anderson, B.; Tucker, C.J.; Dickinson, R.E.; Lucht, W.; Potter, C.S.; Myneni, R.B. Interannual covariability in Northern Hemisphere air temperatures and greenness associated with El Niño-Southern Oscillation and the Arctic Oscillation. J. Geophys. Res. Atmos. 2003, 108. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Dai, A.; Van Der Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Chen, W.; Feng, J.; Wu, R. Roles of ENSO and PDO in the link of the East Asian winter monsoon to the following summer monsoon. J. Clim. 2013, 26, 622–635. [Google Scholar] [CrossRef]
- Schneider, W.; Donoso, D.; Garcés-Vargas, J.; Escribano, R. Water-column cooling and sea surface salinity increase in the upwelling region off central-south Chile driven by a poleward displacement of the South Pacific High. Prog. Oceanogr. 2017, 151, 38–48. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The central Chile mega drought (2010–2018): A climate dynamics perspective. Int. J. Climatol. 2020, 40, 421–439. [Google Scholar] [CrossRef]
- McWethy, D.B.; Pauchard, A.; García, R.A.; Holz, A.; González, M.E.; Veblen, T.T.; Stahl, J.; Currey, B. Landscape drivers of recent fire activity (2001–2017) in south-central Chile. PLoS ONE 2018, 13, e0201195. [Google Scholar] [CrossRef]
- Lara, C.; Cazelles, B.; Saldías, G.S.; Flores, R.P.; Paredes, Á.L.; Broitman, B.R. Coupled Biospheric Synchrony of the Coastal Temperate Ecosystem in Northern Patagonia: A Remote Sensing Analysis. Remote Sens. 2019, 11, 2092. [Google Scholar] [CrossRef]
- Hessl, A.; Allen, K.J.; Vance, T.; Abram, N.J.; Saunders, K.M. Reconstructions of the southern annular mode (SAM) during the last millennium. Prog. Phys. Geogr. 2017, 41, 834–849. [Google Scholar] [CrossRef]
- Marshall, G.J. Trends in the Southern Annular Mode from observations and reanalyses. J. Clim. 2003, 16, 4134–4143. [Google Scholar] [CrossRef]
- Gong, D.; Wang, S. Definition of Antarctic oscillation index. Geophys. Res. Lett. 1999, 26, 459–462. [Google Scholar] [CrossRef]
- Garreaud, R. Record-breaking climate anomalies lead to severe drought and environmental disruption in western Patagonia in 2016. Clim. Res. 2018, 74, 217–229. [Google Scholar] [CrossRef]
- Wu, X.; Mao, J. Interdecadal modulation of ENSO-related spring rainfall over South China by the Pacific Decadal Oscillation. Clim. Dyn. 2016, 47, 3203–3220. [Google Scholar] [CrossRef]
- Schoennagel, T.; Veblen, T.T.; Romme, W.H.; Sibold, J.S.; Cook, E.R. ENSO and PDO variability affect drought-induced fire occurrence in Rocky Mountain subalpine forests. Ecol. Appl. 2005, 15, 2000–2014. [Google Scholar] [CrossRef]
- Wu, D.; Zhao, X.; Liang, S.; Zhou, T.; Huang, K.; Tang, B.; Zhao, W. Time-lag effects of global vegetation responses to climate change. Glob. Chang. Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef]
- Gutierrez, J.R.; Meserve, P.L.; Kelt, D.A.; Engilis Jr, A.; Previtali, M.A.; Milstead, W.B.; Jaksic, F.M. Long-term research in Bosque Fray Jorge National Park: Twenty years studying the role of biotic and abiotic factors in a Chilean semiarid scrubland. Rev. Chil. Hist. Nat. 2010, 83, 69–98. [Google Scholar] [CrossRef][Green Version]
- Meserve, P.L.; Kelt, D.A.; Previtali, M.A.; Milstead, W.B.; Gutiérrez, J.R. Global climate change and small mammal populations in north-central Chile. J. Mammal. 2011, 92, 1223–1235. [Google Scholar] [CrossRef]
- Meserve, P.L.; Kelt, D.A.; Gutiérrez, J.R.; Previtali, M.A.; Milstead, W.B. Biotic interactions and community dynamics in the semiarid thorn scrub of Bosque Fray Jorge National Park, north-central Chile: A paradigm revisited. J. Arid Environ. 2016, 126, 81–88. [Google Scholar] [CrossRef]
- Le Maire, G.; Marsden, C.; Nouvellon, Y.; Grinand, C.; Hakamada, R.; Stape, J.L.; Laclau, J.P. MODIS NDVI time-series allow the monitoring of Eucalyptus plantation biomass. Remote Sens. Environ. 2011, 115, 2613–2625. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Lara, C.; Saldías, G.S.; Paredes, A.L.; Cazelles, B.; Broitman, B.R. Temporal variability of MODIS phenological indices in the temperate rainforest of Northern Patagonia. Remote Sens. 2018, 10, 956. [Google Scholar] [CrossRef]
- Mutanga, O.; Skidmore, A.K. Narrow band vegetation indices overcome the saturation problem in biomass estimation. Int. J. Remote Sens. 2004, 25, 3999–4014. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| National Park | Area () | Year Created | Vegetation Formation |
|---|---|---|---|
| (1) Fray Jorge (S) | 86.06 | 1941 | Desertic scrub |
| (2) La Campana (S) | 78.18 | 1967 | Deciduous forest |
| Sclerophyllous forest | |||
| Alpine dwarf scrub | |||
| (3) Alerce Costero (S) | 236.14 | 2010 | Valdivian Laured- |
| Leaved Forest | |||
| Fitzroya Forest | |||
| (4) Tantauco (S) | 1016.4 | 2005 | Deciduous forest |
| (5) San Rafael (S) | 18,693.40 | 1959 | Deciduous forest |
| Sclerophyllous forest | |||
| Evergreen forest | |||
| Alpine dwarf scrub | |||
| Deciduous shrub | |||
| Evergreen shrub | |||
| Peatbog | |||
| (6) Torres del Paine (S) | 2216.47 | 1959 | Deciduous forest |
| Evergreen forest | |||
| Alpine dwarf scrub | |||
| Matorral arborescent | |||
| Deciduous shrub |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lara, C.; Saldías, G.S.; Cazelles, B.; Rivadeneira, M.M.; Muñoz, R.; Galán, A.; Paredes, Á.L.; Fierro, P.; Broitman, B.R. Climatic Regulation of Vegetation Phenology in Protected Areas along Western South America. Remote Sens. 2021, 13, 2590. https://doi.org/10.3390/rs13132590
Lara C, Saldías GS, Cazelles B, Rivadeneira MM, Muñoz R, Galán A, Paredes ÁL, Fierro P, Broitman BR. Climatic Regulation of Vegetation Phenology in Protected Areas along Western South America. Remote Sensing. 2021; 13(13):2590. https://doi.org/10.3390/rs13132590
Chicago/Turabian StyleLara, Carlos, Gonzalo S. Saldías, Bernard Cazelles, Marcelo M. Rivadeneira, Richard Muñoz, Alexander Galán, Álvaro L. Paredes, Pablo Fierro, and Bernardo R. Broitman. 2021. "Climatic Regulation of Vegetation Phenology in Protected Areas along Western South America" Remote Sensing 13, no. 13: 2590. https://doi.org/10.3390/rs13132590
APA StyleLara, C., Saldías, G. S., Cazelles, B., Rivadeneira, M. M., Muñoz, R., Galán, A., Paredes, Á. L., Fierro, P., & Broitman, B. R. (2021). Climatic Regulation of Vegetation Phenology in Protected Areas along Western South America. Remote Sensing, 13(13), 2590. https://doi.org/10.3390/rs13132590