1. Introduction
Recent interest in climate change and global warming has affected the distribution and function of vegetation by changing the environmental conditions of ecosystems [
1]. Since the Antarctic ecosystem is particularly sensitive to environmental change, studying its responses can enhance our understanding of such changes [
2,
3,
4,
5,
6]. The monitoring of terrestrial biodiversity in the Antarctic Peninsula and the South Shetland Islands, which are some of the most rapidly warming areas in the world [
6,
7], is vital as a proxy for climate change.
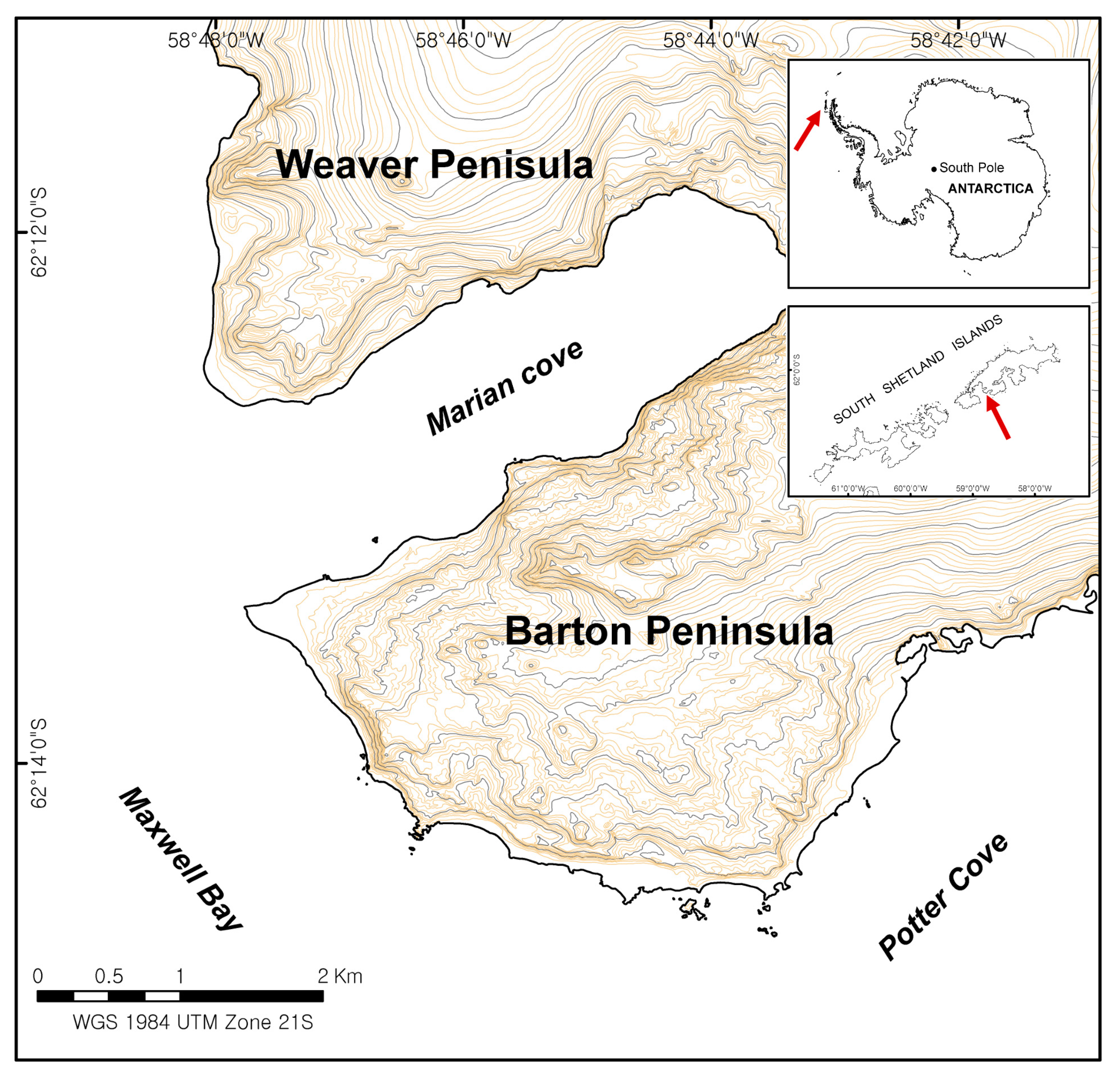
Detailed information on the maritime biodiversity in Antarctica has been collected via field surveys, but this information is limited to the areas most frequently visited by researchers, primarily near Antarctic research stations. Therefore, on King George Island, the largest of the South Shetland Islands and the most concentrated region of Antarctic research stations, various biodiversity monitoring studies via field surveys have been conducted. In the Maxwell Bay area of King George Island, vegetation cover was surveyed [
8], and the species density of bryophytes and lichens was calculated for the Barton Peninsula around King Sejong Station [
9]. Due to the extreme environmental conditions, only two species of flowering plants,
Deschampsia antarctica and
Colobanthus quitensis, grow in small and sparse patches; however, cryptogamic species, including 33 bryophytes (mosses) and 62 lichens, are dominant in most snow-free regions [
8,
9]. Changes in the lichen community diversity and composition of a deglaciated gradient were analyzed using five 50 × 50 cm grids at 24 sites in the Potter Peninsula [
10]. The present study reports that the patterns of these Antarctic lichen communities are dynamic and highly heterogeneous due to a strong influence of microsite factors and macroclimate variables. In Admiralty Bay, biomass and net annual production patterns were analyzed for four moss species in Antarctica [
11], and the driving mechanisms of vegetation, including soil characteristics and landscape elements (altitude and geomorphological variations), were investigated [
12] in the surrounding area of Arctowski Station. A distribution map was generated for 35 vegetation communities of the Keller Peninsula [
13], and the lichen biota dynamics of Lions Rump for the past 20 years were characterized, showing that the most significant changes had taken place in the forefield of a glacier and on the young moraines [
14]. In addition, the distributions of the representative flowering plant
Deschampsia antarctica [
15] and of mosses [
16] were surveyed and compared between the South Shetland Islands and the Antarctic Peninsula.
However, periodic field surveys of extended areas are financially and physically costly, and the human activity can potentially destroy or disturb the ecosystem. Additionally, survey-based approaches maintain inherent uncertainty when quantifying vegetation cover and variations over relatively long time periods. As an alternative, remote sensing techniques are widely used to periodically monitor quantitative and qualitative states of extensive and/or inaccessible regions. Several studies have been conducted to assess polar vegetational biodiversity using remote sensing approaches over the past decade. Théau et al. [
17] used a classification method and spectral mixture analysis for mapping several types of lichen over Northern Quebec using Landsat imagery. They acquired field data using a helicopter and determined the types and percent of coverage by visual inspection. Large-scale mapping analyses using the normalized difference vegetation index (NDVI) from medium-resolution Landsat [
18] and SPOT [
19] images have been conducted; however, detailed information from these studies is not available due to the limited spatial and spectral resolution of the images and ground truthing efforts. Nonetheless, such research has suggested that a combination of remote sensing data and techniques could potentially help obtain information. Shin et al. [
20] used a spectral mixture analysis technique to quantify vegetation abundance at the sub-pixel level from high-resolution KOMPSAT-2 (Korea Multi-Purpose Satellite-2) and QuickBird images of the Barton Peninsula; however, they employed image-driven endmembers instead of field spectra. Liu and Treitz [
21] used ground-based near-infrared (NIR) images to develop a vegetation cover model for the Canadian High Arctic, and then applied this model to high-resolution satellite images to examine the spatio-temporal patterns of the vegetation cover over the extended areas. Sun et al. [
22] investigated vegetation abundance and health mapping from WorldView-2 data using spectral mixture analysis in the Fildes Peninsula and Ardley Island, King George Island. They categorized mosses and lichens according to their associated health conditions and acquired reference field spectra over approximately 300–1000 nm for large sampling plots (2 m × 2 m), which are suitable for obtaining high resolution satellite data. Accordingly, past vegetation mapping studies of remote sensing data have focused on the analysis of vegetation/spectral indices and on the classification/unmixing of a few vegetation species or conditions due to the relative lack of spectral information and images of the Arctic/Antarctic vegetation.
Incorporating the field spectra of the target areas into the remote sensing analyses can greatly assist with the assessments of inaccessible areas. A spectral library is a set of digital reflectance spectra measured in the laboratory or the field, or remotely by air or spacecraft. They are employed to support imaging spectroscopy studies of the Earth and other planets and include informative wavelength data of both natural and artificial surfaces, such as rocks, minerals, soils, vegetation, snow, and ice [
23]. A spectral library can play an important role in identifying the components of unknown targets and quantifying their abundance in mixed pixels. Widely used spectral libraries are contributed to by the Advanced Spaceborne Thermal Emission Reflection Radiometer (ASTER), the NASA Jet Propulsion Laboratory (JPL), the Johns Hopkins University, and the United States Geological Survey (USGS). However, these spectral libraries have been primarily developed for use in mid-latitudinal regions. As the landscapes of the polar regions are quite different, the current libraries may not be applicable for interpreting the ecosystems and landcover types in the Antarctic region. Limited research has been aimed at developing a spectral library of polar vegetation. Goswami and Matharasi [
24] developed a vegetation spectral library for the Arctic, Antarctic, and Chihuahuan Desert regions using a hand-held portable spectrometer in the visible and NIR (VNIR) wavelength range. Calvino-Cancela and Martin-Herreroin [
25] developed a spectral library for 13 representative moss and lichen species in the Barton Peninsula in the VNIR wavelength range (380–1000 nm). Their classified field-measured spectra showed high overall accuracy in principal component analysis–linear discriminant analysis. However, the analysis also concluded that some species, such as
Deschampsia and
Prasiola, were challenging to discriminate using their library spectra.
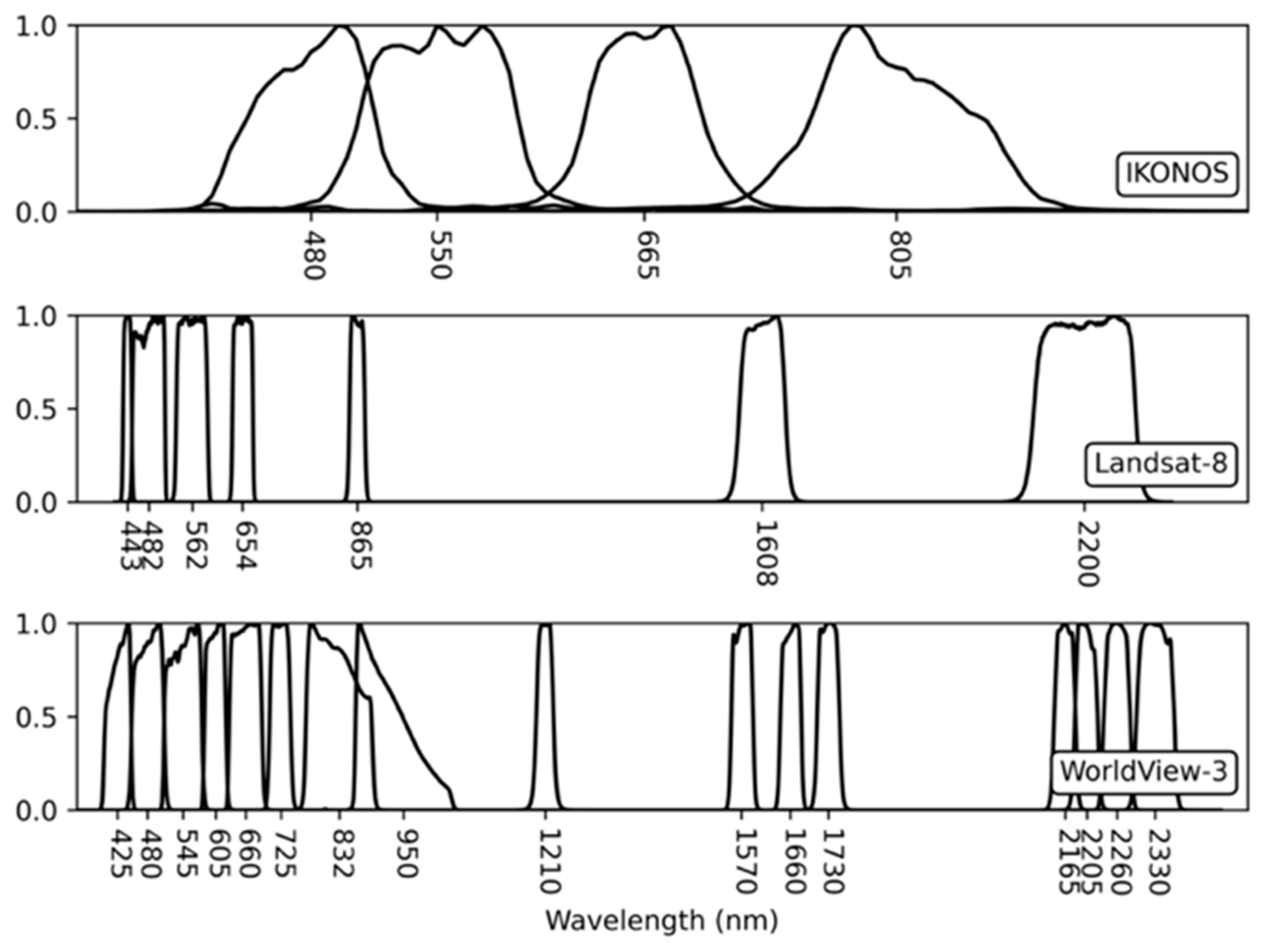
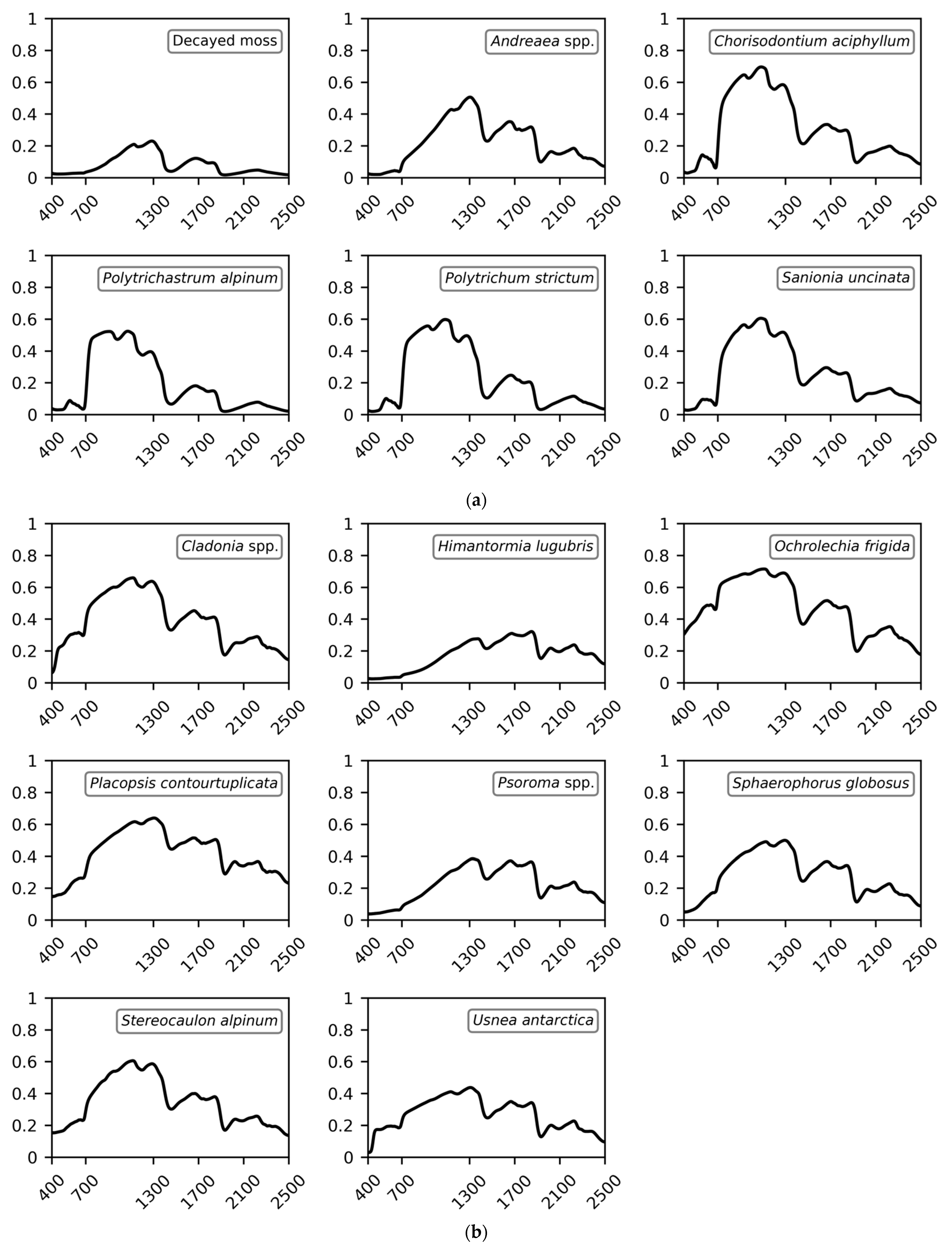
Although the spectral library is crucial for quantitative remote sensing analysis, particularly for inaccessible or difficult-to-access locations such as the polar regions, limited spectral information for ground objects in the polar regions is available. This study had two research goals: (1) the development of a spectral library for Antarctic vegetation; and (2) the investigation of the spectral characteristics of Antarctic vegetation to determine which spectral ranges are the most effective for identifying Antarctic vegetation in remote sensing data. To this end, we first developed a spectral library for 16 representative vegetation species and decayed moss, including mosses, lichens, vascular plants, and one alga often discovered in the ice-free areas of Antarctica. Iterative field spectral data across the visible–shortwave-infrared (SWIR) range (400–2500 nm) were collected for homogeneous vegetation patches and then compiled to generate representative spectra. Both spectrometers and hyperspectral imaging sensors typically use the VNIR (400–1000 nm), SWIR (1000–2500 nm), or full-spectrum (i.e., VNIR–SWIR; 400–2500 nm) ranges. As VNIR sensors are lighter, cheaper, and have a better signal-to-noise ratio than SWIR or full-spectrum sensors, using a VNIR spectrometer or integrating a VNIR imaging sensor to a small unmanned aerial vehicle (UAV) has fewer technical challenges. Therefore, along with developing a spectral library of the Antarctic vegetation species over a full-spectral range, analyses were carried out using spectral discrimination measures to observe the spectral characteristics and potential separability of the Antarctic vegetation by wavelength range. Since sufficient remote sensing images with corresponding surveyed data over the study area are unavailable, artificial images were created and modeled using the developed spectral library, field information, and spectral response functions of widely used remote imaging sensors. The potential performance of the vegetation discrimination was then evaluated for practical classification and unmixing tasks. In particular, we addressed the importance of SWIR wavelengths in discriminating or quantifying Antarctic vegetation species effectively.
4. Discussion
Research on vegetation species classification using remote sensing data is currently popular. However, as we discussed in the introduction, this research has been conducted based on limited field-survey information or image-driven spectral information due to the unavailability of spectral libraries for Antarctic vegetation species. We believe that the spectral library developed in this study can be used as ground truth and can assist in understanding remote sensing data in various disciplines. Along with the development of the spectral library, two research questions were raised in this study: (1) which spectral ranges are required to effectively analyze Antarctic vegetation in remote sensing? (2) which sensor is appropriate for classifying or quantifying vegetation species? To address these questions, we conducted experiments to compare interspecies separability and performed image classification/unmixing using simulated remote sensing imagery according to spectral resolution.
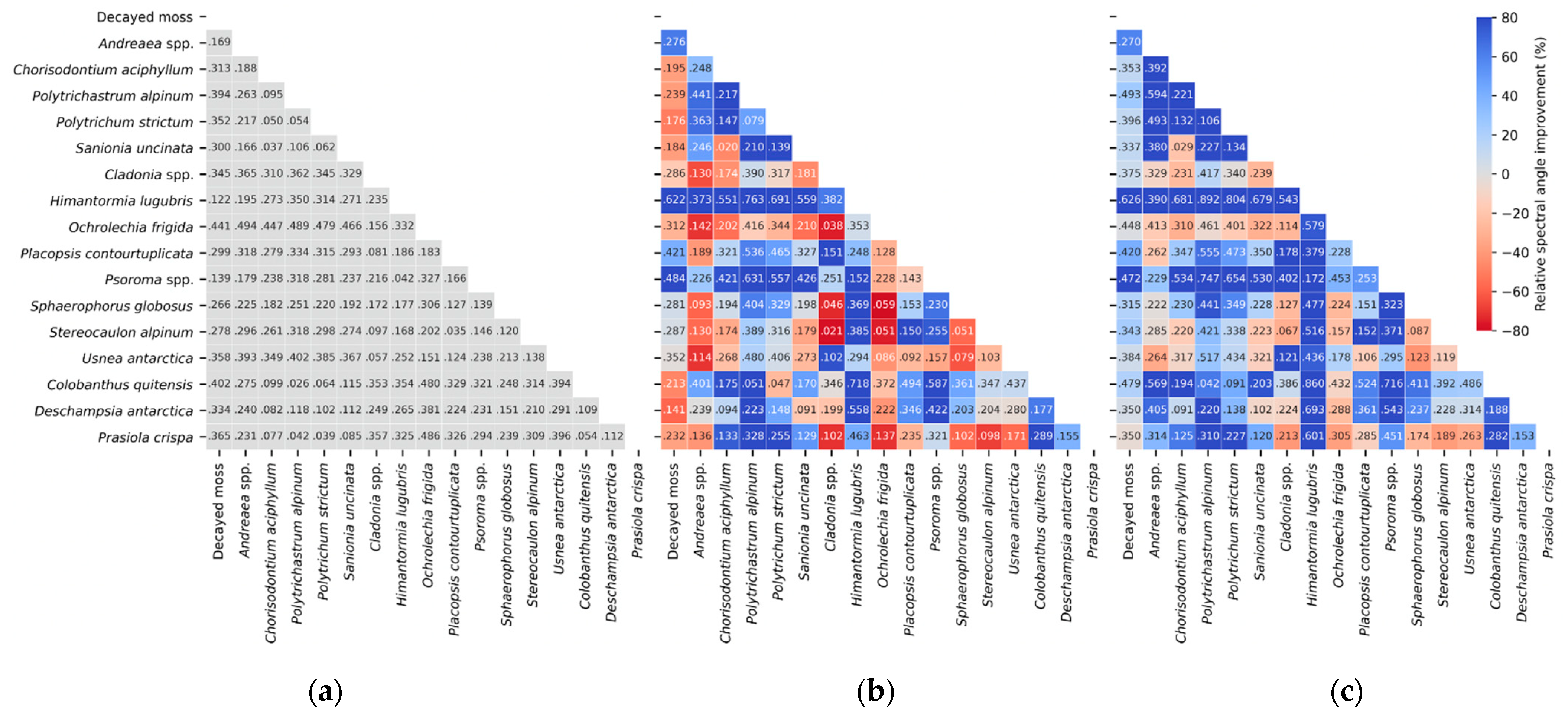
First, whereas only visible and NIR wavelengths have been investigated for the Antarctic vegetation in previous research [
25], this study explored a full spectral range and revealed the importance of using SWIR wavelengths for accurately identifying various vegetation classes. The VNIR range could generally explain cell structure and leaf pigments better than longer wavelengths; however, vegetation in cold regions may have relatively weaker spectral signals corresponding to this information and may present stronger water- or biochemical-related information in the SWIR region, compared with vegetation in temperate latitudes. This was tested using individual spectra and simulated image data discussed in
Section 3. Particularly, the identification of
Deschampsia and
Prasiola, which has previously been reported as a challenge when using data from across the VNIR range [
25], was improved at 1000–2500 nm.
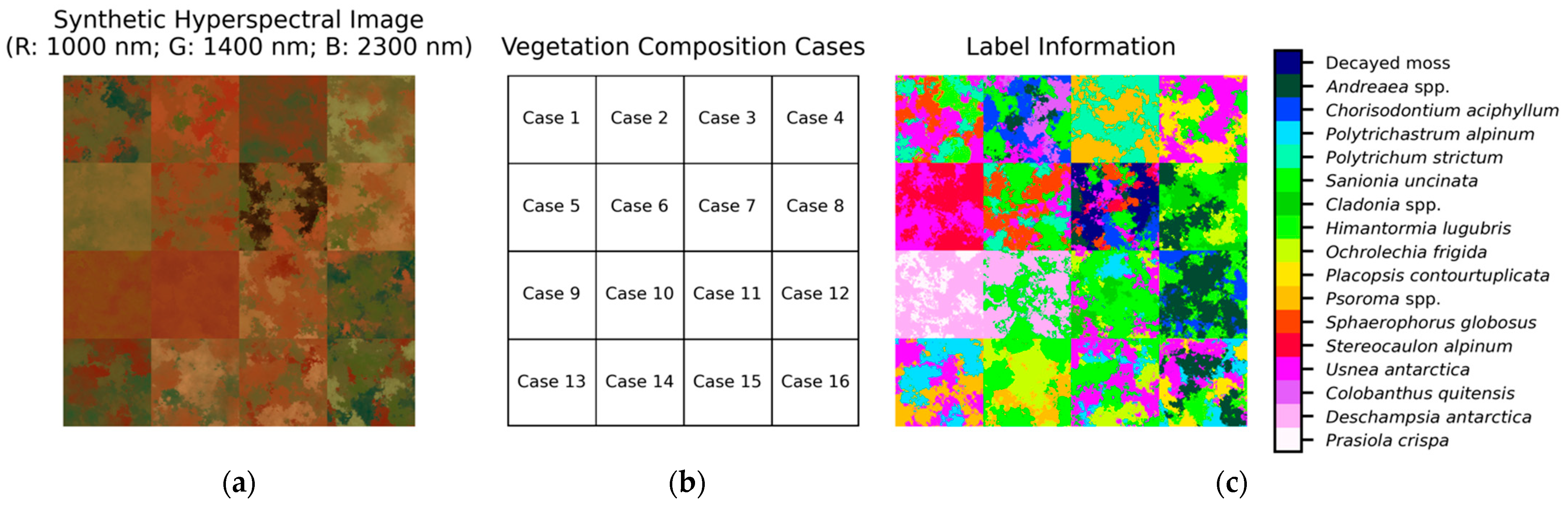
With respect to the second question: the synthetic images in
Section 3.3 were generated using library spectra to model popular remote sensing images, including multi- and hyperspectral sensors, owing to the unavailability of real image data for the study area. SAM-based classification and linear unmixing methods were applied to the generated images to investigate which sensor was optimal for analyzing vegetation distributions in practical applications. These experiments represented ideal scenarios since simulated images of known library spectra, using assumed linear mixture models and the ground truth, were used. With true remote sensing imagery, the resulting accuracy will be lower, as it will be difficult to associate the library spectra with the images owing to different data acquisition conditions. However, under the same experimental conditions, hyperspectral data significantly outperformed multispectral sensor data in identifying and quantifying Antarctic vegetation. In particular, these results also indicate that spectral information from the SWIR wavelengths was more effective than that from the VNIR. However, it is worth noting that multispectral sensors showed reasonable classification performance for several dominant species. Particularly, multispectral sensors with more SWIR range bands showed better results than those with fewer bands. Therefore, considering the difficulty in acquiring high-quality hyperspectral data, the purpose of the application, and the vegetation compositions of the study area, the selection of proper remote sensing data would be important.
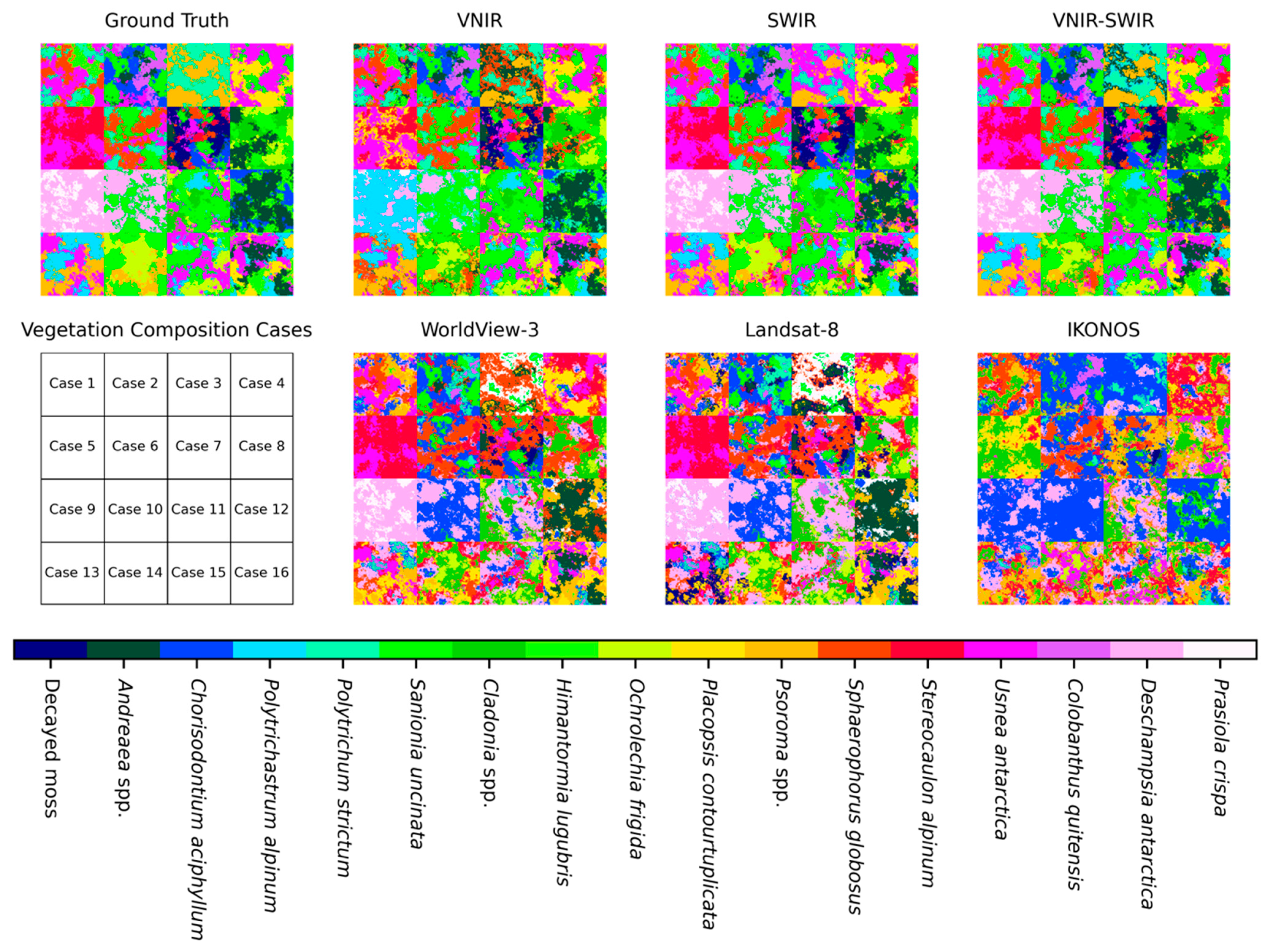
Although we demonstrated the potential performance of several sensors to analyze vegetation species according to their spectral resolutions, it is difficult to conclude which species are clearly separated by which sensor or wavelength region. For example, in Case 12, consisting of
Andreaea,
Chorisodontium and
Himantormia, as shown in
Figure 8, the VNIR region images showed better classification results than the SWIR and VNIR-SWIR region images. Some
Andreaea and
Himantormia pixels in the SWIR and VNIR-SWIR region images were misclassified to
Usnea and
Psoroma, respectively. However, the
Andreaea species in Cases 2, 7, and 16 were successfully identified in both the VNIR and SWIR range data, with even WorldView-3 and Landsat-8 classifying them relatively well, compared with the other species.
Himantormia in Cases 1 and 16 were also correctly classified using the VNIR and SWIR datasets. This should indicate that while the spectral resolution of the sensor is crucial, types of species that consist of image pixels are also important for determining the sensor’s classification and unmixing performance.
Conventional surveys assessing vegetation cover over quadrants in the Barton Peninsula have been conducted for several years across sparse locations. However, these surveys are costly, and their findings do not adequately quantify the overall vegetation cover. Recent advances in sensor technologies have facilitated the development of miniaturized hyperspectral sensors for drones. In future work, the spectral library created here, and hyperspectral UAV data should be incorporated to generate detailed vegetation maps of the entire Barton Peninsula, as an alternative to conducting physically intensive field surveys. Satellite remote sensing data, including a new hyperspectral sensor PRISMA (PRecursore IperSpettrale della Missione Applicativa) [
54] or high-resolution multispectral data, should also be investigated using the spectral library, based on research goals and sensor performance. In addition, intraspecies spectral variations exist according to color, shape, and environmental conditions. As the spectral library was created with multiple measurements for each species and was temporally distanced from rain or snow events, the spectra used should be considered the most representative signals of the species. However, developing a more detailed library that factors in environmental and physiological conditions is a worthwhile alternative to the potentially destructive contact that currently occurs during the characterization of vegetation.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}