Detection of Two Different Grapevine Yellows in Vitis vinifera Using Hyperspectral Imaging

 , , , , ,
, , , , ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Material
2.1.1. Greenhouse Plants
2.1.2. Field Samples
2.2. Molecular Analysis
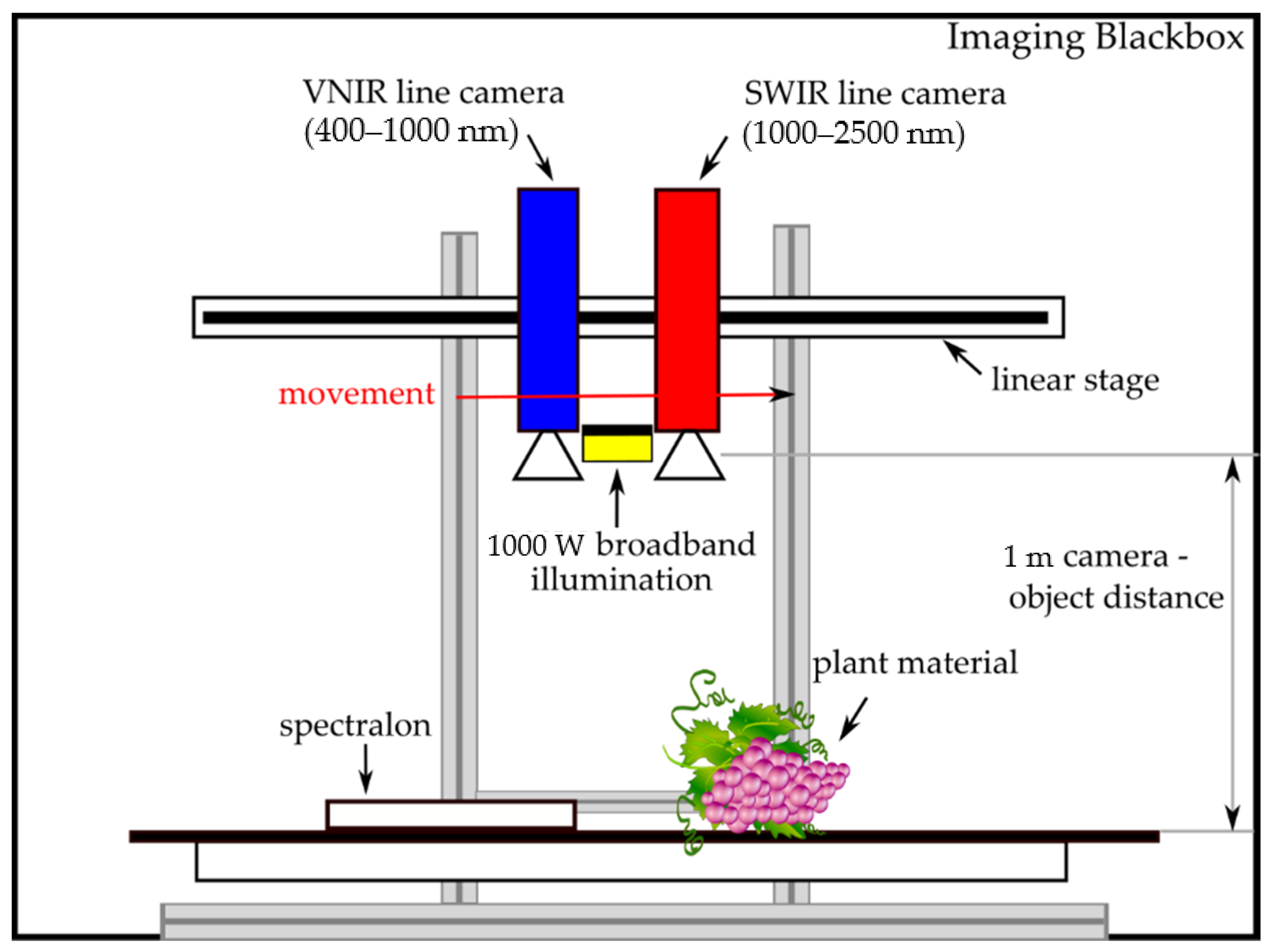
2.3. Hyperspectral Sensors and Data Acquisition
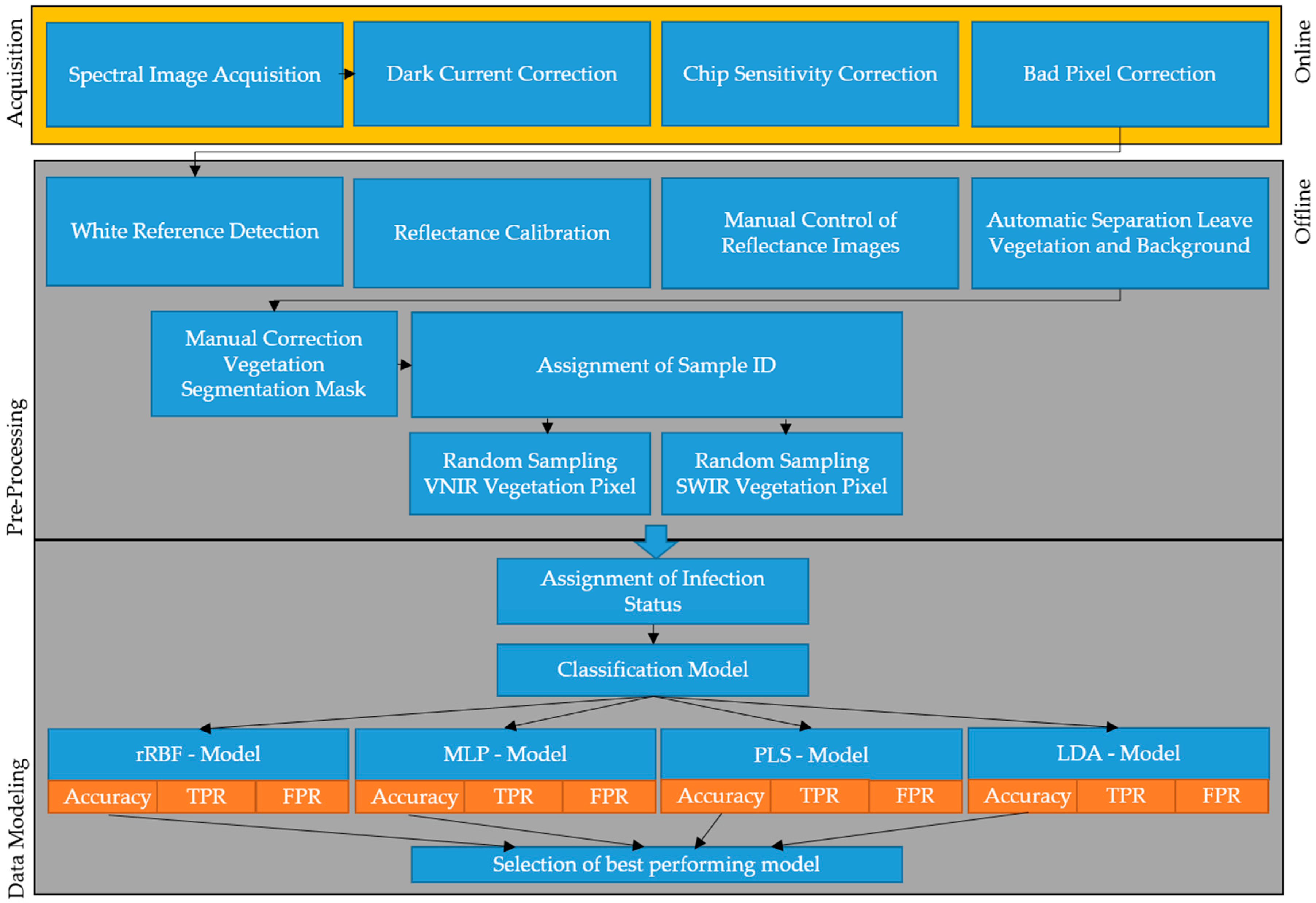
2.4. Data Calibration and Labeling
2.5. Model Development and Application
- Classification Accuracy (CA): Ratio calculated from the number of samples correctly classified among all possible samples.
- True Positive Rate (TPR): Ratio calculated from the number of samples detected correctly as infected among all possible infected samples.
- False Positive Rate (FPR): Ratio calculated from the number of samples detected incorrectly as infected among all possible control samples.
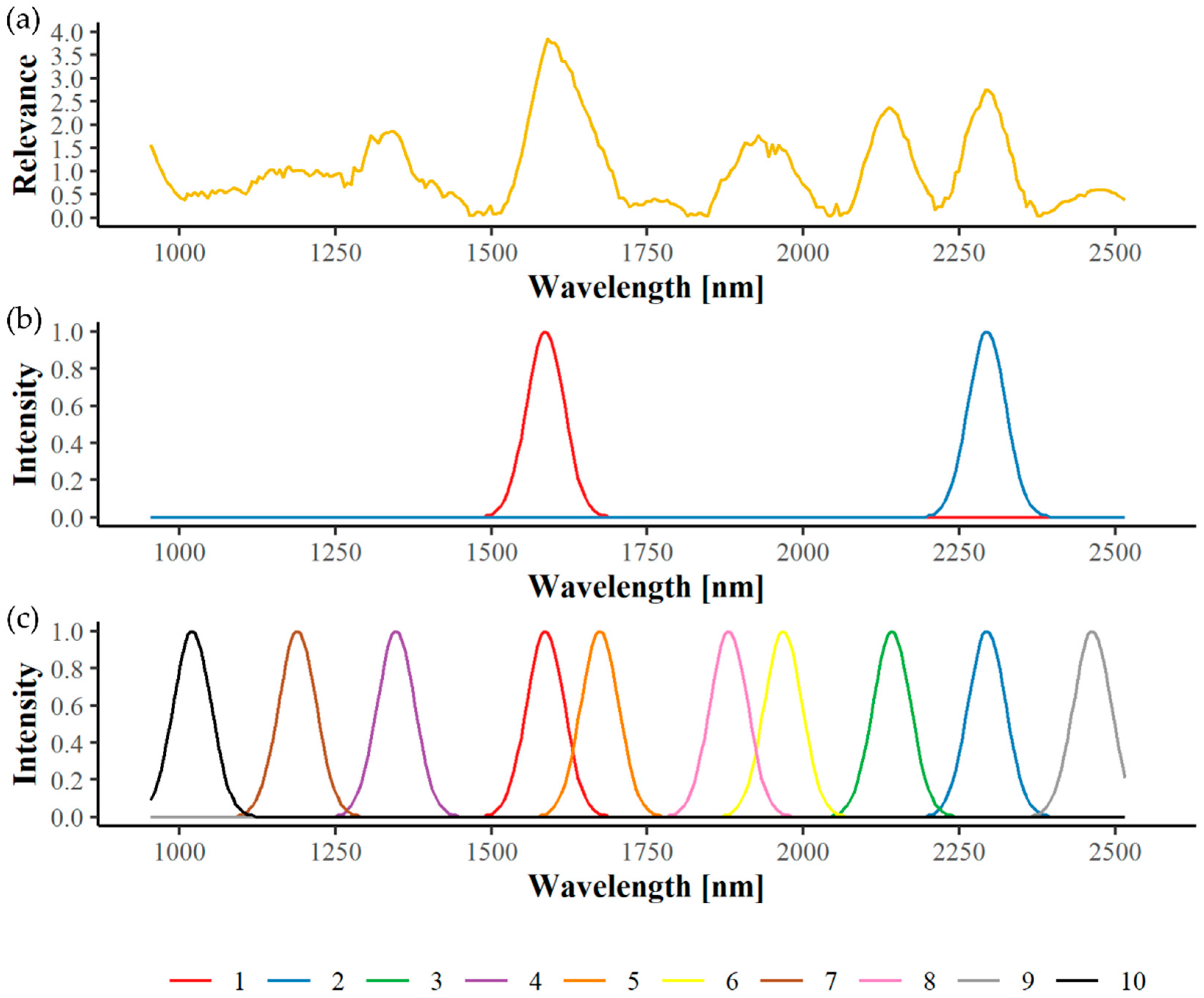
2.6. Spectral Relevance and Important Wavelengths
3. Results
3.1. Model Evaluation
3.1.1. Greenhouse Plants
3.1.2. Field Samples
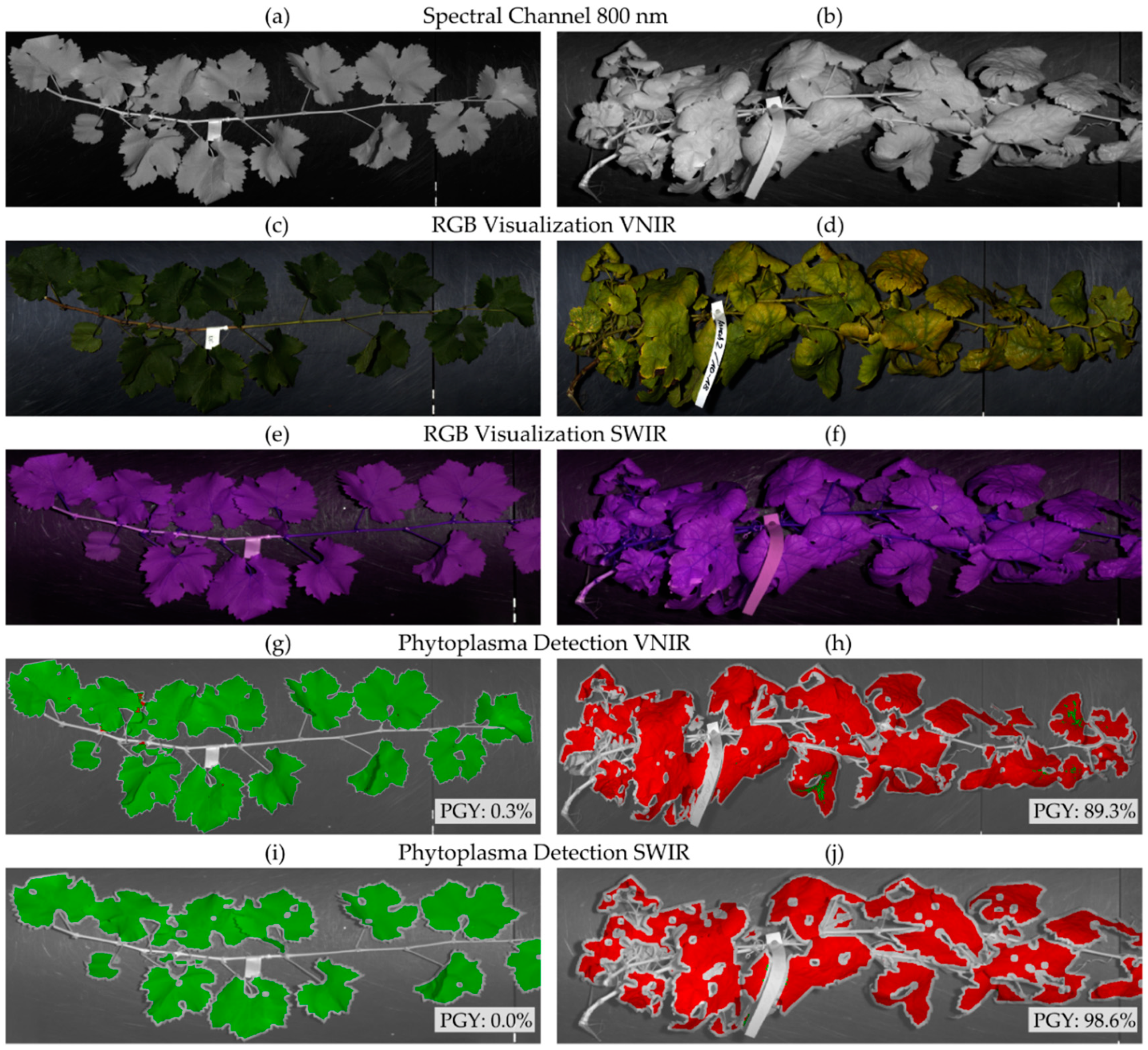
3.2. Model Application
3.2.1. Symptomatic Greenhouse Plants
3.2.2. Nonsymptomatic Greenhouse Plants
3.2.3. Symptomatic Field Material
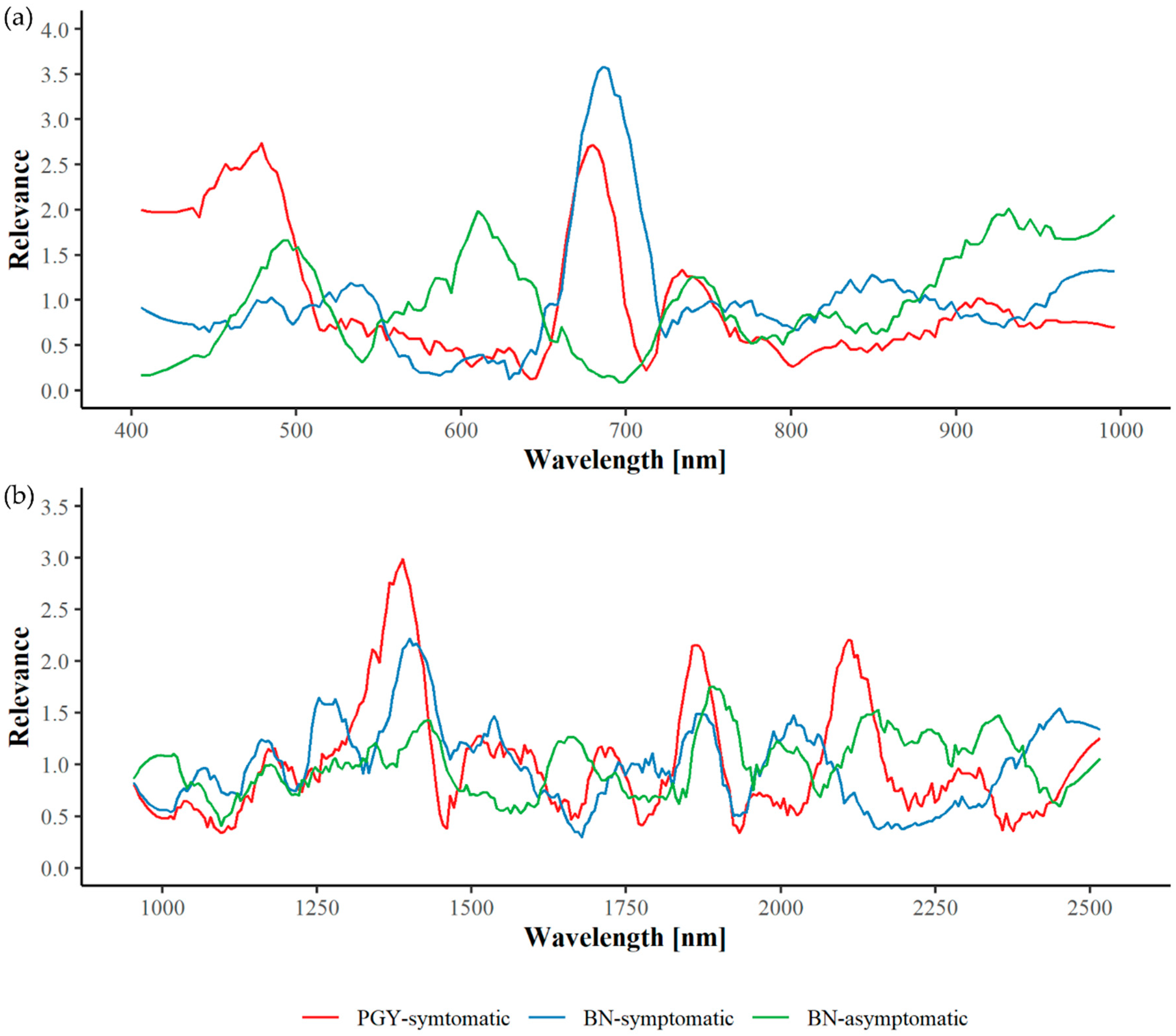
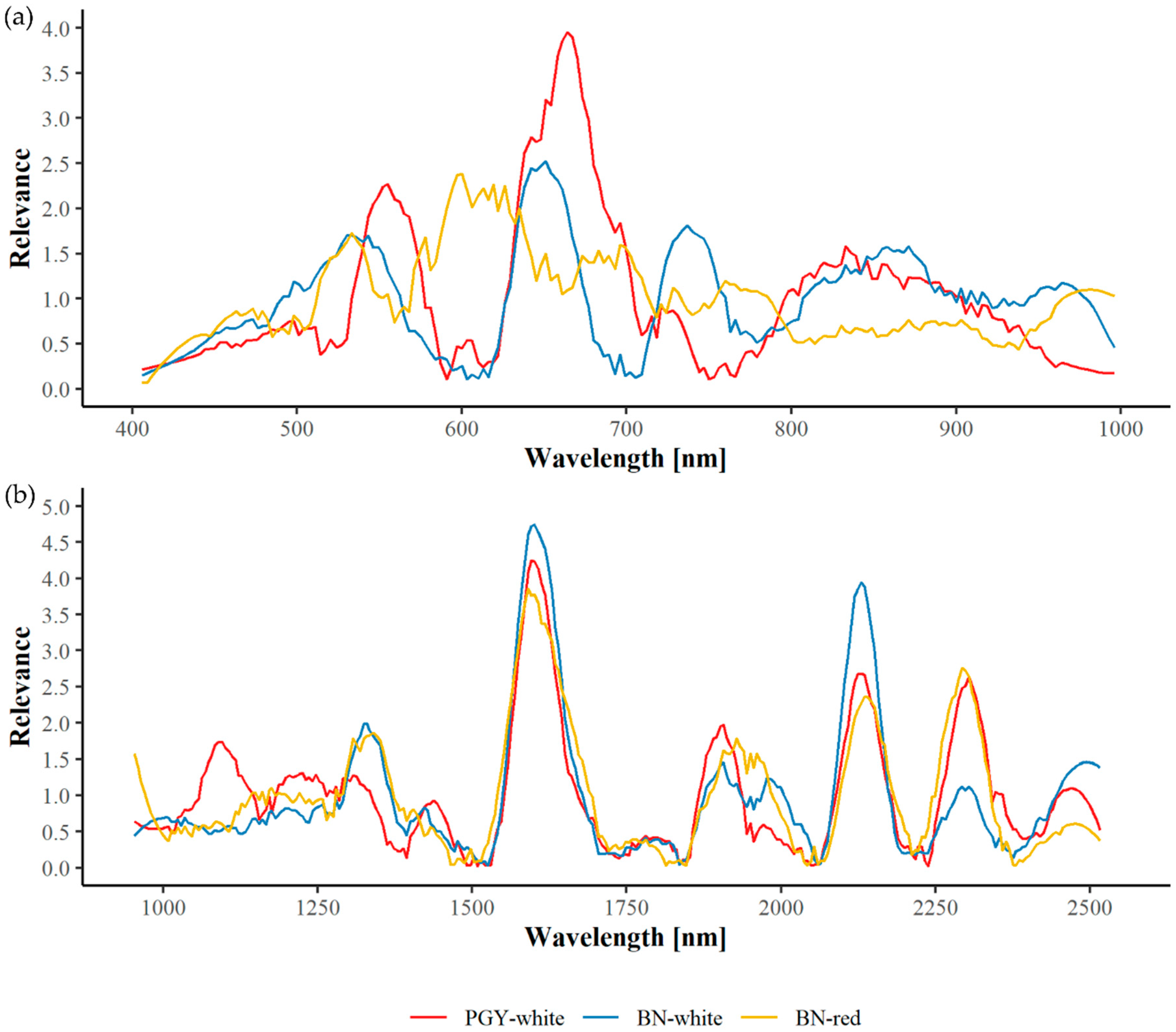
3.3. Spectral Relevance and Important Wavelengths
3.3.1. Greenhouse Plants
3.3.2. Field Material
4. Discussion
5. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Angelini, E.; Constable, F.; Duduk, B.; Fiore, N.; Quaglino, F.; Bertaccini, A. Grapevine phytoplasmas. In Phytoplasmas: Plant Pathogenic Bacteria-I; Rao, G., Bertaccini, A., Fiore, N., Liefting, L., Eds.; Springer: Singapore, 2018; pp. 123–151. [Google Scholar]
- Maixner, M.; Rüdel, M.; Daire, X.; Boudon-Padieu, E. Diversity of grapevine yellows in Germany. Vitis 1995, 34, 235–236. [Google Scholar]
- Quaglino, F.; Zhao, Y.; Casati, P.; Bulgari, D.; Bianco, P.A.; Wei, W.; Davis, R.E. ‘Candidatus Phytoplasma solani’, a novel taxon associated with stolbur- and bois noir-related diseases of plants. Int. J. Syst. Evol. Microbiol. 2013, 63, 2879–2894. [Google Scholar] [CrossRef] [PubMed]
- Sforza, R.; Clair, D.; Daire, X.; Larrue, J.; Boudon-Padieu, E. The role of Hyalesthes obsoletus (Hemiptera: Cixiidae) in the occurrence of bois noir of grapevines in France. J. Phytopathol. 1998, 146, 549–556. [Google Scholar] [CrossRef]
- Arnaud, G.; Malembic-Maher, S.; Salar, P.; Bonnet, P.; Maixner, M.; Marcone, C.; Boudon-Padieu, E.; Foissac, X. Multilocus sequence typing confirms the close genetic interrelatedness of three distinct Flavescence dorée phytoplasma strain clusters and group 16SrV phytoplasmas infecting grapevine and alder in Europe. Appl. Environ. Microbiol. 2007, 73, 4001–4010. [Google Scholar] [CrossRef] [PubMed]
- Maixner, M.; Reinert, W.; Darimont, H. Transmission of grapevine yellows by Oncopsis alni (Schrank)(Auchenorrhyncha: Macropsinae). Vitis 2000, 39, 83–84. [Google Scholar]
- Maixner, M.; Albert, A.; Johannesen, J. Survival relative to new and ancestral host plants, phytoplasma infection, and genetic constitution in host races of a polyphagous insect disease vector. Ecol. Evol. 2014, 4, 3082–3092. [Google Scholar] [CrossRef]
- Maixner, M.; Reinert, W. Oncopsis alni (Schrank)(Auchenorrhyncha: Cicadellidae) as a vector of the alder yellows phytoplasma of Alnus glutinosa (L.) Gaertn. Eur. J. Plant Pathol. 1999, 105, 87–94. [Google Scholar] [CrossRef]
- Caglayan, K.; Gazel, M.; Škorić, D. Transmission of phytoplasmas by agronomic practices. In Phytoplasmas: Plant Pathogenic Bacteria-II; Bertaccini, A., Weintraub, P., Rao, G., Mori, N., Eds.; Springer: Singapore, 2019; pp. 149–163. [Google Scholar]
- Bertaccini, A.; Duduk, B.; Paltrinieri, S.; Contaldo, N. Phytoplasmas and phytoplasma diseases: A severe threat to agriculture. Am. J. Plant Sci. 2014, 5, 1763–1788. [Google Scholar] [CrossRef]
- Belli, G.; Bianco, P.; Conti, M. Grapevine yellows in Italy: Past, present and future. J. Plant Pathol. 2010, 92, 303–326. [Google Scholar] [CrossRef]
- Eveillard, S.; Jollard, C.; Labroussaa, F.; Khalil, D.; Perrin, M.; Desqué, D.; Salar, P.; Razan, F.; Hévin, C.; Bordenave, L.; et al. Contrasting susceptibilities to Flavescence dorée in Vitis vinifera, rootstocks and wild Vitis species. Front. Plant Sci. 2016, 7, 1762. [Google Scholar] [CrossRef]
- Maixner, M. Grapevine yellows—Current developments and unsolved questions. In Proceedings of the 15th Meeting of ICVG, Stellenbosch, South Africa, 3–7 April 2006; pp. 86–88. [Google Scholar]
- Bianco, P.A.; Romanazzi, G.; Mori, N.; Myrie, W.; Bertaccini, A. Integrated management of phytoplasma diseases. In Phytoplasmas: Plant Pathogenic Bacteria-II; Bertaccini, A., Weintraub, P., Rao, G., Mori, N., Eds.; Springer: Singapore, 2019; pp. 237–258. [Google Scholar]
- Mahlein, A.K. Plant disease detection by imaging sensors—Parallels and specific demands for precision agriculture and plant phenotyping. Plant Dis. 2016, 100, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Bock, C.H.; Barbedo, J.G.; del Ponte, E.M.; Bohnenkamp, D.; Mahlein, A.-K. From visual estimates to fully automated sensor-based measurements of plant disease severity: Status and challenges for improving accuracy. Phytopathol. Res. 2020, 2, 9. [Google Scholar] [CrossRef]
- Arens, N.; Backhaus, A.; Doll, S.; Fischer, S.; Seiffert, U.; Mock, H.P. Non-invasive presymptomatic detection of Cercospora beticola infection and identification of early metabolic responses in sugar beet. Front. Plant Sci. 2016, 7, 1377. [Google Scholar] [CrossRef] [PubMed]
- Polder, G.; van der Heijden, G.W.A.M.; van Doorn, J.; Baltissen, T.A.H.M.C. Automatic detection of tulip breaking virus (TBV) in tulip fields using machine vision. Biosyst. Eng. 2014, 117, 35–42. [Google Scholar] [CrossRef]
- Behmann, J.; Bohnenkamp, D.; Paulus, S.; Mahlein, A.-K. Spatial referencing of hyperspectral images for tracing of plant disease symptoms. J. Imaging 2018, 4, 143. [Google Scholar] [CrossRef]
- Delalieux, S.; Somers, B.; Verstraeten, W.W.; van Aardt, J.A.N.; Keulemans, W.; Coppin, P. Hyperspectral indices to diagnose leaf biotic stress of apple plants, considering leaf phenology. Int. J. Remote Sens. 2009, 30, 1887–1912. [Google Scholar] [CrossRef]
- Kuska, M.; Wahabzada, M.; Leucker, M.; Dehne, H.W.; Kersting, K.; Oerke, E.C.; Steiner, U.; Mahlein, A.K. Hyperspectral phenotyping on the microscopic scale: Towards automated characterization of plant-pathogen interactions. Plant Methods 2015, 11, 28. [Google Scholar] [CrossRef]
- Barthel, D.; Fischnaller, S.; Eisenstecken, D.; Kerschbamer, C.; Messner, M.; Dordevic, N.; Robatscher, P.; Janik, K. Near-infrared spectroscopy analysis—A useful tool to detect apple proliferation diseased trees? Phytopathog. Mollicutes 2019, 9, 79–80. [Google Scholar] [CrossRef]
- Albetis, J.; Duthoit, S.; Guttler, F.; Jacquin, A.; Goulard, M.; Poilvé, H.; Féret, J.-B.; Dedieu, G. Detection of Flavescence dorée grapevine disease using unmanned aerial vehicle (UAV) multispectral imagery. Remote Sens. 2017, 9, 308. [Google Scholar] [CrossRef]
- Albetis, J.; Jacquin, A.; Goulard, M.; Poilvé, H.; Rousseau, J.; Clenet, H.; Dedieu, G.; Duthoit, S. On the potentiality of UAV multispectral imagery to detect Flavescence dorée and grapevine trunk diseases. Remote Sens. 2018, 11, 23. [Google Scholar] [CrossRef]
- Al-Saddik, H.; Simon, J.C.; Cointault, F. Development of spectral disease indices for ‘Flavescence dorée’ grapevine disease identification. Sensors 2017, 17, 2772. [Google Scholar] [CrossRef]
- Al-Saddik, H.; Laybros, A.; Billiot, B.; Cointault, F. Using image texture and spectral reflectance analysis to detect yellowness and Esca in grapevines at leaf-level. Remote Sens. 2018, 10, 618. [Google Scholar] [CrossRef]
- Al-Saddik, H.; Simon, J.-C.; Cointault, F. Assessment of the optimal spectral bands for designing a sensor for vineyard disease detection: The case of “Flavescence dorée”. Precis. Agric. 2019, 20, 398–422. [Google Scholar] [CrossRef]
- Maixner, M.; Ahrens, U.; Seemüller, E. Detection of the German grapevine yellows (Vergilbungskrankheit) MLO in grapevine, alternative hosts and a vector by a specific PCR procedure. Eur. J. Plant Pathol. 1995, 101, 241–250. [Google Scholar] [CrossRef]
- Lorenz, K.; Schneider, B.; Ahrens, U.; Seemüller, E. Detection of the apple proliferation and pear decline phytoplasmas by PCR amplification of ribosomal and nonribosomal DNA. Phytopathology 1995, 85, 771–776. [Google Scholar] [CrossRef]
- Schneider, B.; Seemüller, E.; Smart, C.; Kirkpatrick, B. Phylogenetic classification of plant pathogenic mycoplasma-like organisms or phytoplasmas. In Molecular and Diagnostic Procedures in Mycoplasmology; Razin, S., Tully, J., Eds.; Academic Press: San Diego, CA, USA, 1995; Volume 1, pp. 369–380. [Google Scholar]
- Marcone, C.; Ragozzino, A.; Seemüller, E. Detection of an elm yellows-related phytoplasma in eucalyptus trees affected by little-leaf disease in Italy. Plant Dis. 1996, 80, 669–673. [Google Scholar] [CrossRef]
- Bendel, N.; Kicherer, A.; Backhaus, A.; Köckerling, J.; Maixner, M.; Bleser, E.; Klück, H.-C.; Seiffert, U.; Voegele, R.T.; Töpfer, R. Detection of grapevine leafroll-associated virus 1 and 3 in white and red grapevine cultivars using hyperspectral imaging. Remote Sens. 2020, 12, 1693. [Google Scholar] [CrossRef]
- Bendel, N.; Kicherer, A.; Backhaus, A.; Klück, H.-C.; Seiffert, U.; Fischer, M.; Voegele, R.T.; Töpfer, R. Evaluating the suitability of hyper- and multispectral imaging to detect foliar symptoms of the grapevine trunk disease Esca in vineyards. Plant Methods 2020, 16, 142. [Google Scholar] [CrossRef]
- Cybenko, G. Approximation by superpositions of a sigmoidal function. Math Control Sign. Syst. 1989, 2, 303–314. [Google Scholar] [CrossRef]
- Asaari, M.S.M.; Mishra, P.; Mertens, S.; Dhondt, S.; Inzé, D.; Wuyts, N.; Scheunders, P. Close-range hyperspectral image analysis for the early detection of stress responses in individual plants in a high-throughput phenotyping platform. ISPRS J. Photogram. Remote Sens. 2018, 138, 121–138. [Google Scholar] [CrossRef]
- Fortuna, L.; Graziani, S.; Rizzo, A.; Xibilia, M.G. Soft Sensors for Monitoring and Control of Industrial Processes; Springer Science & Business Media: London, UK, 2007. [Google Scholar]
- Krzanowski, W. Principles of Multivariate Analysis: A User’s Perspective; Clarendon Press: Oxford, UK, 1988; pp. 291–301. [Google Scholar]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Møller, M.F. A scaled conjugate gradient algorithm for fast supervised learning. Neural Netw. 1993, 6, 525–533. [Google Scholar] [CrossRef]
- Moody, J.; Darken, C.J. Fast learning in networks of locally-tuned processing units. Neural Comput. 1989, 1, 281–294. [Google Scholar] [CrossRef]
- Backhaus, A.; Bollenbeck, F.; Seiffert, U. Robust classification of the nutrition state in crop plants by hyperspectral imaging and artificial neural networks. In Proceedings of the 3rd Workshop on Hyperspectral Image and Signal Processing: Evolution in Remote Sensing (Whispers), Lisbon, Portugal, 6–9 June 2011; pp. 1–4. [Google Scholar]
- Dehghani, R.; Mahdavi-Amiri, N. Scaled nonlinear conjugate gradient methods for nonlinear least squares problems. Numer. Algorithms 2019, 82, 1–20. [Google Scholar] [CrossRef]
- Becker, F.; Backhaus, A.; Johrden, F.; Flitter, M. Optimal multispectral sensor configurations through machine learning for cognitive agriculture. Automatisierungstechnik Spec. Issue Cognetive Agric. 2020. Accepted for Publication. [Google Scholar]
- Martinetz, T.M.; Berkovich, S.G.; Schulten, K.J. ‘Neural-gas’ network for vector quantization and its application to time-series prediction. IEEE Transact. Neural Netw. 1993, 4, 558–569. [Google Scholar] [CrossRef]
- Siedliska, A.; Baranowski, P.; Zubik, M.; Mazurek, W.; Sosnowska, B. Detection of fungal infections in strawberry fruit by VNIR/SWIR hyperspectral imaging. Postharvest Biol. Technol. 2018, 139, 115–126. [Google Scholar] [CrossRef]
- Wiegmann, M.; Backhaus, A.; Seiffert, U.; Thomas, W.T.; Flavell, A.J.; Pillen, K.; Maurer, A. Optimizing the procedure of grain nutrient predictions in barley via hyperspectral imaging. PLoS ONE 2019, 14, e0224491. [Google Scholar] [CrossRef]
- Abdulridha, J.; Ehsani, R.; de Castro, A. Detection and differentiation between laurel wilt disease, phytophtora disease, and salinity damage using hyperspectral sensing technique. Agriculture 2016, 6, 56. [Google Scholar] [CrossRef]
- Terlizzi, F.; Credi, R. Uneven distribution of stolbur phytoplasma in Italian grapevines as revealed by nested-PCR. Bull. Insect. 2007, 60, 365–366. [Google Scholar]
- Afonso, A.M.; Guerra, R.; Cavaco, A.M.; Pinto, P.; Andrade, A.; Duarte, A.; Power, D.M.; Marques, N.T. Identification of asymptomatic plants infected with Citrus tristeza virus from a time series of leaf spectral characteristics. Comput. Electron. Agric. 2017, 141, 340–350. [Google Scholar] [CrossRef]
- Mannini, F. Hot water treatment and field coverage of mother plant vineyards to prevent propagation material from phytoplasma infections. Bull. Insect. 2007, 60, 311–312. [Google Scholar]
- Wang, D.; Vinson, R.; Holmes, M.; Seibel, G.; Bechar, A.; Nof, S.; Tao, Y. Early detection of tomato spotted wilt virus by hyperspectral imaging and outlier removal auxiliary classifier generative adversarial nets (OR-AC-GAN). Sci. Rep. 2019, 9, 4377. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.-H.; Chung, W.-C.; Liao, J.-Y.; Chung, C.-L.; Kuo, Y.-F.; Lin, T.-T. Strawberry foliar anthracnose assessment by hyperspectral imaging. Comput. Electron. Agric. 2016, 122, 1–9. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Rumpf, T.; Welke, P.; Dehne, H.W.; Plümer, L.; Steiner, U.; Oerke, E.C. Development of spectral indices for detecting and identifying plant diseases. Remote Sens. Environ. 2013, 128, 21–30. [Google Scholar] [CrossRef]
- Knauer, U.; Matros, A.; Petrovic, T.; Zanker, T.; Scott, E.S.; Seiffert, U. Improved classification accuracy of powdery mildew infection levels of wine grapes by spatial-spectral analysis of hyperspectral images. Plant Methods 2017, 13, 47. [Google Scholar] [CrossRef]
- Musetti, R.; Buxa, S.V.; de Marco, F.; Loschi, A.; Polizzotto, R.; Kogel, K.-H.; van Bel, A.J. Phytoplasma-triggered Ca2+ influx is involved in sieve-tube blockage. Mol. Plant Microbe Interact. 2013, 26, 379–386. [Google Scholar] [CrossRef]
- Hren, M.; Nikolic, P.; Rotter, A.; Blejec, A.; Terrier, N.; Ravnikar, M.; Dermastia, M.; Gruden, K. ‘Bois noir’ phytoplasma induces significant reprogramming of the leaf transcriptome in the field grown grapevine. BMC Genom. 2009, 10, 460. [Google Scholar] [CrossRef]
- Bertamini, M.; Nedunchezhian, N.; Tomasi, F.; Grando, M. Phytoplasma [Stolbur-subgroup (Bois Noir-BN)] infection inhibits photosynthetic pigments, ribulose-1, 5-bisphosphate carboxylase and photosynthetic activities in field grown grapevine (Vitis vinifera L. cv. Chardonnay) leaves. Physiol. Mol. Plant Path. 2002, 61, 357–366. [Google Scholar] [CrossRef]
- Blackburn, G.A. Hyperspectral remote sensing of plant pigments. J. Exp. Bot. 2007, 58, 855–867. [Google Scholar] [CrossRef]
- Curran, P.J. Remote sensing of foliar chemistry. Remote Sens. Environ. 1989, 30, 271–278. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I. Visible and near-infrared reflectance techniques for diagnosing plant physiological status. Trends Plant Sci. 1998, 3, 151–156. [Google Scholar] [CrossRef]
- Margaria, P.; Ferrandino, A.; Caciagli, P.; Kedrina, O.; Schubert, A.; Palmano, S. Metabolic and transcript analysis of the flavonoid pathway in diseased and recovered Nebbiolo and Barbera grapevines (Vitis vinifera L.) following infection by Flavescence dorée phytoplasma. Plant Cell Environ. 2014, 37, 2183–2200. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Lee, E.; Bogs, J.; McDavid, D.A.; Thomas, M.R.; Robinson, S.P. White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J. 2007, 49, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Gitelson, A.A.; Mezlyak, M.N.; Chivkunova, O.B. Optical properties and nondestructive estimation of anthocyanin content in plant leaves. Photochem. Photobiol. 2001, 74, 38–45. [Google Scholar] [CrossRef]
- Blackburn, G.A. Spectral indices for estimating photosynthetic pigment concentrations: A test using senescent tree leaves. Int. J. Remote Sens. 1998, 19, 657–675. [Google Scholar] [CrossRef]
- Christensen, N.M.; Axelsen, K.B.; Nicolaisen, M.; Schulz, A. Phytoplasmas and their interactions with hosts. Trends Plant Sci. 2005, 10, 526–535. [Google Scholar] [CrossRef]
- Negro, C.; Sabella, E.; Nicolì, F.; Pierro, R.; Materazzi, A.; Panattoni, A.; Aprile, A.; Nutricati, E.; Vergine, M.; Miceli, A.; et al. Biochemical changes in leaves of Vitis vinifera cv. Sangiovese infected by Bois noir phytoplasma. Pathogens 2020, 9, 269. [Google Scholar] [CrossRef]
- Nagler, P.; Daughtry, C.; Goward, S. Plant litter and soil reflectance. Remote Sens. Environ. 2000, 71, 207–215. [Google Scholar] [CrossRef]
- Wang, Z.; Skidmore, A.K.; Wang, T.; Darvishzadeh, R.; Hearne, J. Applicability of the PROSPECT model for estimating protein and cellulose + lignin in fresh leaves. Remote Sens. Environ. 2015, 168, 205–218. [Google Scholar] [CrossRef]
- Pagliari, L.; Musetti, R. Phytoplasmas: An introduction. In Phytoplasmas. Methods in Molecular Biology; Musetti, R., Pagliari, L., Eds.; Humana Press: New York, NY, USA, 2019; Volume 1875, pp. 1–6. [Google Scholar]
- Sinha, R.; Khot, L.R.; Rathnayake, A.P.; Gao, Z.; Naidu, R.A. Visible-near infrared spectroradiometry-based detection of grapevine leafroll-associated virus 3 in a red-fruited wine grape cultivar. Comput. Electron. Agric. 2019, 162, 165–173. [Google Scholar] [CrossRef]
- Al-Saddik, H.; Laybros, A.; Simon, J.-C.; Cointault, F. Protocol for the definition of a multi-spectral sensor for specific foliar disease detection: Case of “Flavescence dorée”. In Phytoplasmas. Methods in Molecular Biology; Musetti, R., Pagliari, L., Eds.; Humana Press: New York, NY, USA, 2019; Volume 1875, pp. 213–238. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Cultivar | Disease | PCR Result | ||
|---|---|---|---|---|---|
| Negative | Positive Symptomatic | Positive Nonsymptomatic | |||
| 2017 | ‘Riesling’ | BN | 132 | 16 | 29 |
| 2018 | ‘Scheurebe’ | PGY | 190 | 8 | – |
| PCR Result | Cultivar | Number of Shoots |
|---|---|---|
| Negative | White + Red | 15 |
| PGY | White | 12 |
| BN | White | 84 |
| BN | Red | 37 |
| BN + PGY | Red | 3 |
| Specification | HySpex VNIR 1800 | HySpex SWIR 384 |
|---|---|---|
| Wavelength range (nm) | 400–1000 | 1000–2500 |
| Spectral bands | 256 | 288 |
| Spectral pixels | 1800 | 384 |
| Spectral resolution (nm) | 3.26 | 5.45 |
| Spatial resolution (mm/pixel) | 0.17 | 0.65 |
| Field of view | 17° | 16° |
| Maximum framerate (Hz) | 100 | 400 |
| Dynamic range (bit) | 16 | 16 |
| Detector type | CMOS | MCT at 150 K |
| Method | Formula |
|---|---|
| Vector L2 normalization | |
| Vector SNV normalization [34] |
| Method | Hyper-Parameter | Reference |
|---|---|---|
| Linear Discriminance Model (LDA) | No hyperparameters | [37] |
| Partially Least Square (PLS) | Number of components: 20 | [38] |
| Multi-Layer Perceptron (MLP) | Number of hidden layers: 3 Optimization method: scaled conjugate gradient backpropagation Neurons per hidden layer: 50, 25, 10 | [34,39] |
| Radial-Basis Function Network with Relevance (rRBF) | Number of radial basis functions: 30 Optimization method: scaled nonlinear conjugate gradient | [40,41,42] |
| Disease | Symptoms | Model | Classification Accuracy (%) | True Positive Rate (%) | False Positive Rate (%) | |||
|---|---|---|---|---|---|---|---|---|
| VNIR | SWIR | VNIR | SWIR | VNIR | SWIR | |||
| PGY | Yes | LDA | 77 | 88 | 78 | 85 | 24 | 9 |
| PLS | 77 | 88 | 77 | 86 | 24 | 9 | ||
| MLP | 86 | 88 | 83 | 85 | 22 | 10 | ||
| rRBF | 89 | 92 | 89 | 90 | 11 | 5 | ||
| BN | Yes | LDA | 62 | 65 | 57 | 64 | 26 | 29 |
| PLS | 63 | 65 | 54 | 64 | 28 | 34 | ||
| MLP | 68 | 73 | 65 | 72 | 30 | 27 | ||
| rRBF | 70 | 74 | 68 | 79 | 30 | 15 | ||
| BN | No | LDA | 58 | 60 | 52 | 64 | 36 | 44 |
| PLS | 59 | 60 | 57 | 62 | 39 | 42 | ||
| MLP | 62 | 62 | 63 | 65 | 37 | 46 | ||
| rRBF | 63 | 64 | 68 | 79 | 33 | 36 | ||
| Disease | Symptom Coloration | Model | Classification Accuracy (%) | True Positive Rate (%) | False Positive Rate (%) | |||
|---|---|---|---|---|---|---|---|---|
| VNIR | SWIR | VNIR | SWIR | VNIR | SWIR | |||
| PGY | White | LDA | 96 | 75 | 95 | 75 | 4 | 26 |
| PLS | 96 | 99 | 95 | 97 | 4 | 0 | ||
| MLP | 97 | 99 | 97 | 98 | 3 | 1 | ||
| rRBF | 96 | 98 | 96 | 97 | 3 | 1 | ||
| BN | White | LDA | 88 | 89 | 84 | 82 | 8 | 3 |
| PLS | 88 | 90 | 84 | 82 | 8 | 3 | ||
| MLP | 89 | 90 | 86 | 85 | 7 | 5 | ||
| rRBF | 88 | 91 | 84 | 86 | 8 | 3 | ||
| BN | Red | LDA | 92 | 94 | 86 | 91 | 1 | 3 |
| PLS | 92 | 95 | 86 | 91 | 1 | 2 | ||
| MLP | 94 | 94 | 90 | 93 | 3 | 4 | ||
| rRBF | 93 | 96 | 89 | 94 | 2 | 2 | ||
| VNIR | SWIR | |||
|---|---|---|---|---|
| Application per Plant | CA (%) | 84 | 96 | |
| PGY | TPR (%) | 100 | 100 | |
| FPR (%) | 17 | 4 | ||
| CA (%) | 68 | 79 | ||
| BN | TPR (%) | 81 | 81 | |
| FPR (%) | 34 | 22 |
| VNIR | SWIR | ||
|---|---|---|---|
| Application per Plant | CA (%) | 68 | 64 |
| TPR (%) | 63 | 86 | |
| FPR (%) | 29 | 41 |
| Disease | Symptom Coloration | VNIR | SWIR | ||
|---|---|---|---|---|---|
| Application per Plant | CA (%) | 100 | 100 | ||
| PGY | White | TPR (%) | 100 | 100 | |
| FPR (%) | 0 | 0 | |||
| CA (%) | 96 | 96 | |||
| BN | White | TPR (%) | 96 | 95 | |
| FPR (%) | 7 | 0 | |||
| CA (%) | 98 | 98 | |||
| BN | Red | TPR (%) | 97 | 97 | |
| FPR (%) | 0 | 0 |
| Disease | Symptoms | VNIR | SWIR | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
| PGY | Yes | 679 | 459 | 492 | 423 | 748 | 905 | 965 | 541 | 832 | 606 | 1373 | 1861 | 2125 | 1545 | 1709 | 2288 | 1214 | 2459 | 2000 | 1031 |
| BN | Yes | 689 | 971 | 861 | 539 | 486 | 752 | 914 | 811 | 431 | 631 | 1400 | 2451 | 1865 | 2010 | 1549 | 1239 | 1160 | 1734 | 1055 | 2313 |
| BN | No | 932 | 975 | 503 | 616 | 890 | 734 | 579 | 835 | 455 | 784 | 1893 | 1433 | 2180 | 2362 | 1658 | 1343 | 2000 | 2268 | 2102 | 1170 |
| Disease | Cultivar | VNIR | SWIR | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
| PGY | White | 672 | 639 | 557 | 845 | 891 | 801 | 498 | 718 | 945 | 443 | 1587 | 2132 | 2305 | 1648 | 1918 | 1072 | 1239 | 2458 | 1784 | 1391 |
| BN | White | 637 | 741 | 667 | 862 | 553 | 966 | 509 | 812 | 913 | 452 | 1582 | 2131 | 1649 | 1353 | 2466 | 1981 | 2297 | 1204 | 1869 | 1034 |
| BN | Red | 626 | 528 | 586 | 673 | 773 | 969 | 718 | 458 | 899 | 839 | 1586 | 2294 | 2140 | 1347 | 1670 | 1965 | 1188 | 1881 | 2462 | 1019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bendel, N.; Backhaus, A.; Kicherer, A.; Köckerling, J.; Maixner, M.; Jarausch, B.; Biancu, S.; Klück, H.-C.; Seiffert, U.; Voegele, R.T.; et al. Detection of Two Different Grapevine Yellows in Vitis vinifera Using Hyperspectral Imaging. Remote Sens. 2020, 12, 4151. https://doi.org/10.3390/rs12244151
Bendel N, Backhaus A, Kicherer A, Köckerling J, Maixner M, Jarausch B, Biancu S, Klück H-C, Seiffert U, Voegele RT, et al. Detection of Two Different Grapevine Yellows in Vitis vinifera Using Hyperspectral Imaging. Remote Sensing. 2020; 12(24):4151. https://doi.org/10.3390/rs12244151
Chicago/Turabian StyleBendel, Nele, Andreas Backhaus, Anna Kicherer, Janine Köckerling, Michael Maixner, Barbara Jarausch, Sandra Biancu, Hans-Christian Klück, Udo Seiffert, Ralf T. Voegele, and et al. 2020. "Detection of Two Different Grapevine Yellows in Vitis vinifera Using Hyperspectral Imaging" Remote Sensing 12, no. 24: 4151. https://doi.org/10.3390/rs12244151
APA StyleBendel, N., Backhaus, A., Kicherer, A., Köckerling, J., Maixner, M., Jarausch, B., Biancu, S., Klück, H.-C., Seiffert, U., Voegele, R. T., & Töpfer, R. (2020). Detection of Two Different Grapevine Yellows in Vitis vinifera Using Hyperspectral Imaging. Remote Sensing, 12(24), 4151. https://doi.org/10.3390/rs12244151