Discrimination of Canopy Structural Types in the Sierra Nevada Mountains in Central California



Abstract
1. Introduction
2. Study Site, Materials and Methods
2.1. Study Site
2.2. Data and Methods
2.2.1. Advanced Visible Infrared Imaging Spectrometer (AVIRIS) and Light Detection and Ranging (LiDAR) Preprocessing
2.2.2. Optical and LiDAR Metrics
- (1).
- Sub-pixel cover fractions of Green Vegetation (representing different forest cover types), Soil, Non-Photosynthetic vegetation (NPV), and shade were calculated using MESMA implemented in the Visualization and Image Processing for Environmental Research (VIPER) Tools package using ENVI image analysis software [78]. MESMA was run with the following constraints: 1. the maximum allowable RMSE = 2.5% and 2. the minimum and maximum allowable endmember fractions must fall between −0.05 and 1.05. Spectra of NPV and soil endmembers were independently collected during the overflight using an Analytical Spectral Devices (ASD) full-range spectrometer sensor, the FieldSpec3 Spectroradiometer (Analytical Spectral Devices Inc., Boulder, CO, USA), while green vegetation endmembers were selected from the AVIRIS-classic images. Several endmember spectra were selected for each cover type. The final endmembers chosen had high COB (count-based endmember selection (COB: [79])) and low MASA (minimum average spectral angle (MASA: [80])) and EAR (endmember average RMSE (EAR: [81])) values.
- (2).
- Narrow-band indices representing three types of spectral information were used: 1. those sensitive to the presence of photosynthetic pigments: NDVI [82], Red Edge Normalized Difference Vegetation Index (NDVI705) [83,84], Modified Red Edge Normalized Difference Vegetation Index (mNDVI705) [84,85], and Enhanced Vegetation Index (EVI) [86]; 2. those sensitive to water content: Normalized Difference Water Index (NDWI) [87], Normalized Difference Infrared Index (NDII) [88], and 3. an index sensitive to dry plant matter content: the Cellulose Absorption Index (CAI) [89]. Despite the similarity of these indexes, they provide different projections through the data space related to these processes and each provides useful information, as described later in the Results section. The formulas of the narrow-band indices used in this research are shown in Supplementary Material Table S1.
- (3).
- Spectral canopy water absorption derivatives: Derivative analysis was used to measure the wavelength position and magnitude of the NIR water absorption edges [90], abbreviated here as Wtr1EdgeWvl and Wtr1EdgeMag, the canopy water absorption features between 958–1073 nm and 1105–1168 nm, abbreviated as Wtr1AbAr and Wtr2AbAr, respectively, and the physically-derived equivalent water thickness (EWT; the depth of water/per pixel area) [90,91,92].
- (4).
- Principal component analysis (PCA) [93] was performed both using the full spectral range, as well as the independent regions: visible, near infrared, and shortwave infrared which was done to summarize significant information in all three regions of the spectrum. The PCA components that provided strong relationships with canopy structure [32] were included in this study.
2.2.3. LiDAR-Derived Structural Variables
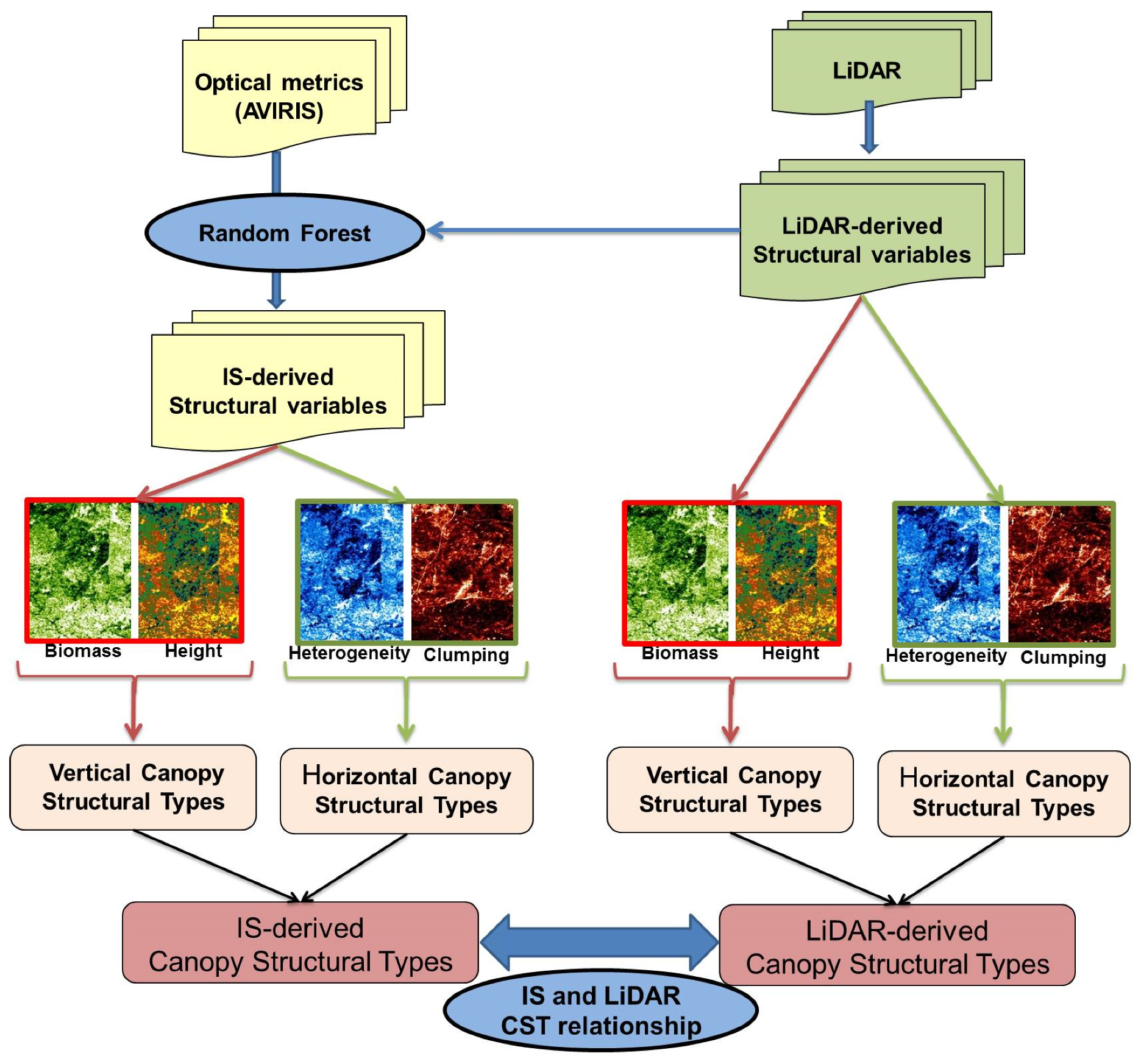
2.2.4. Modeling Structural Variables with Optical Metrics
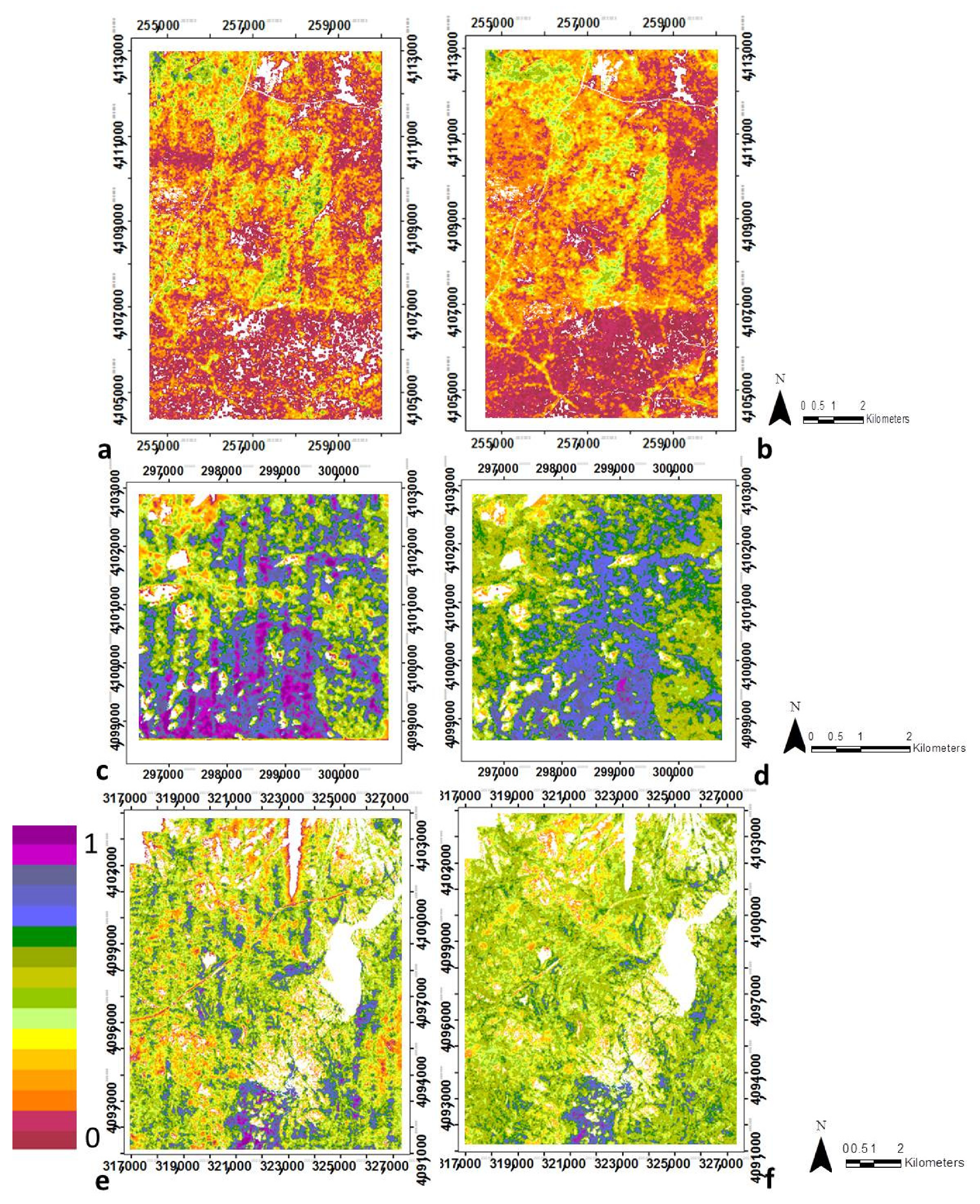
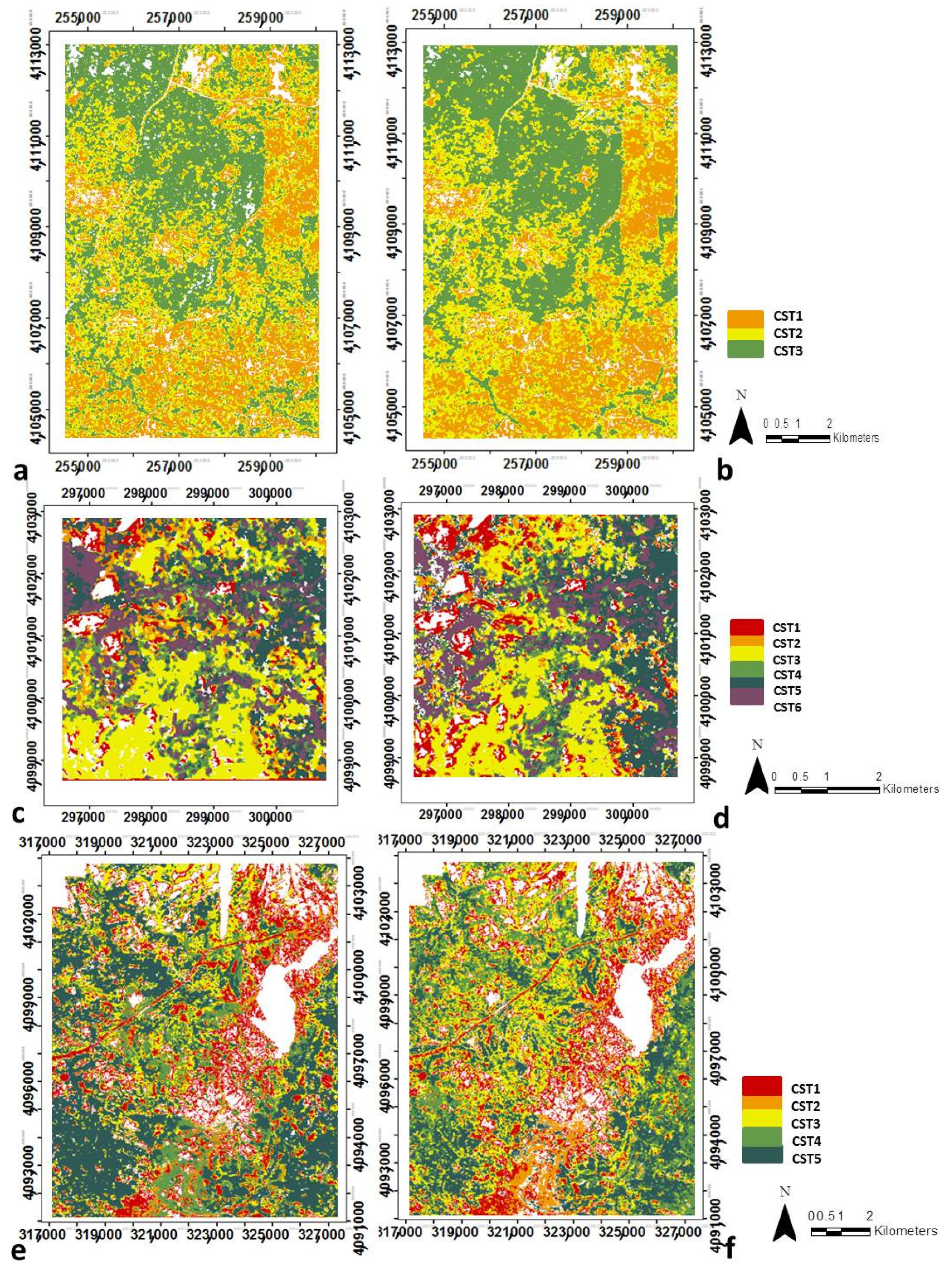
2.2.5. Canopy Structural Types Definition
3. Results
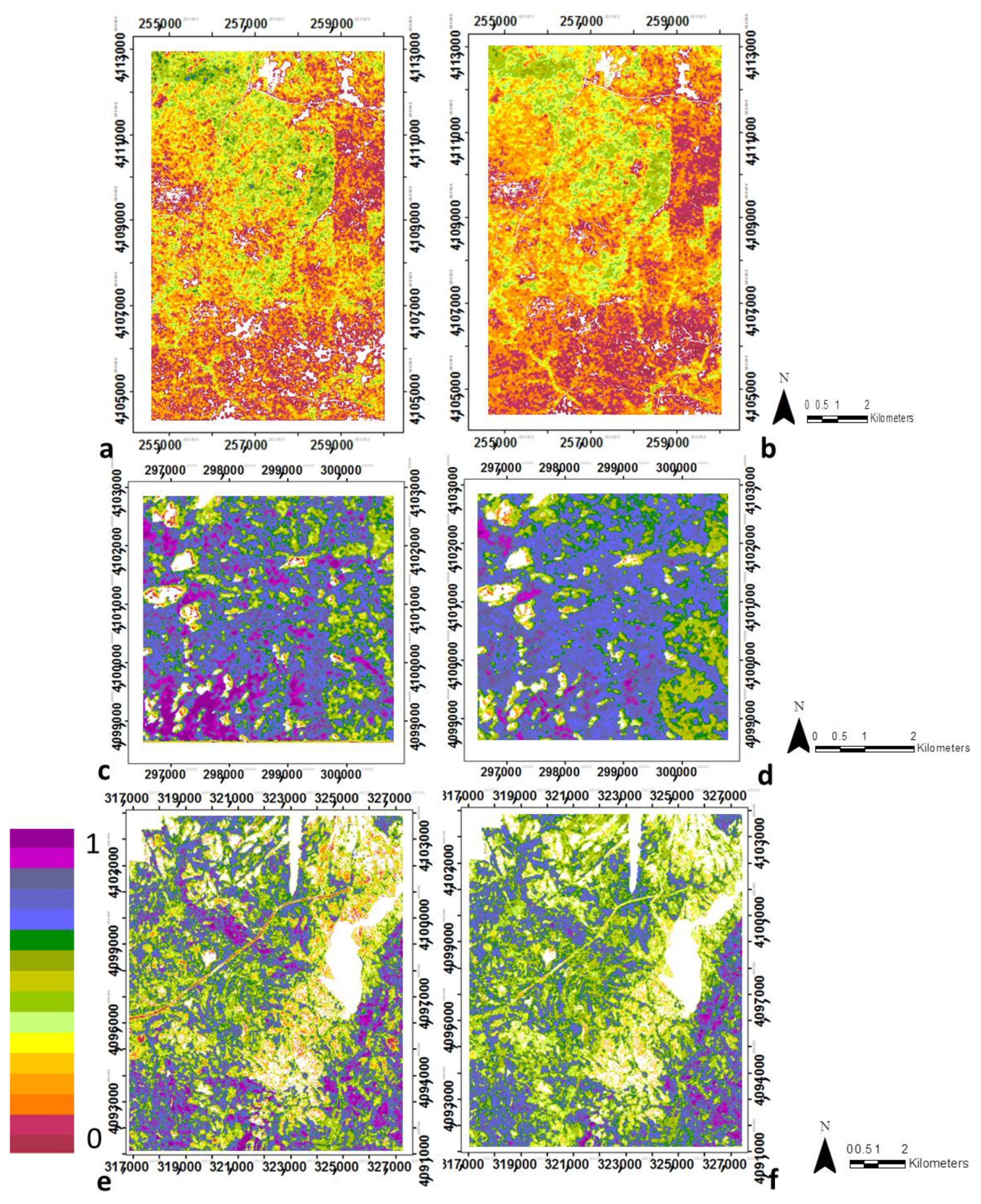
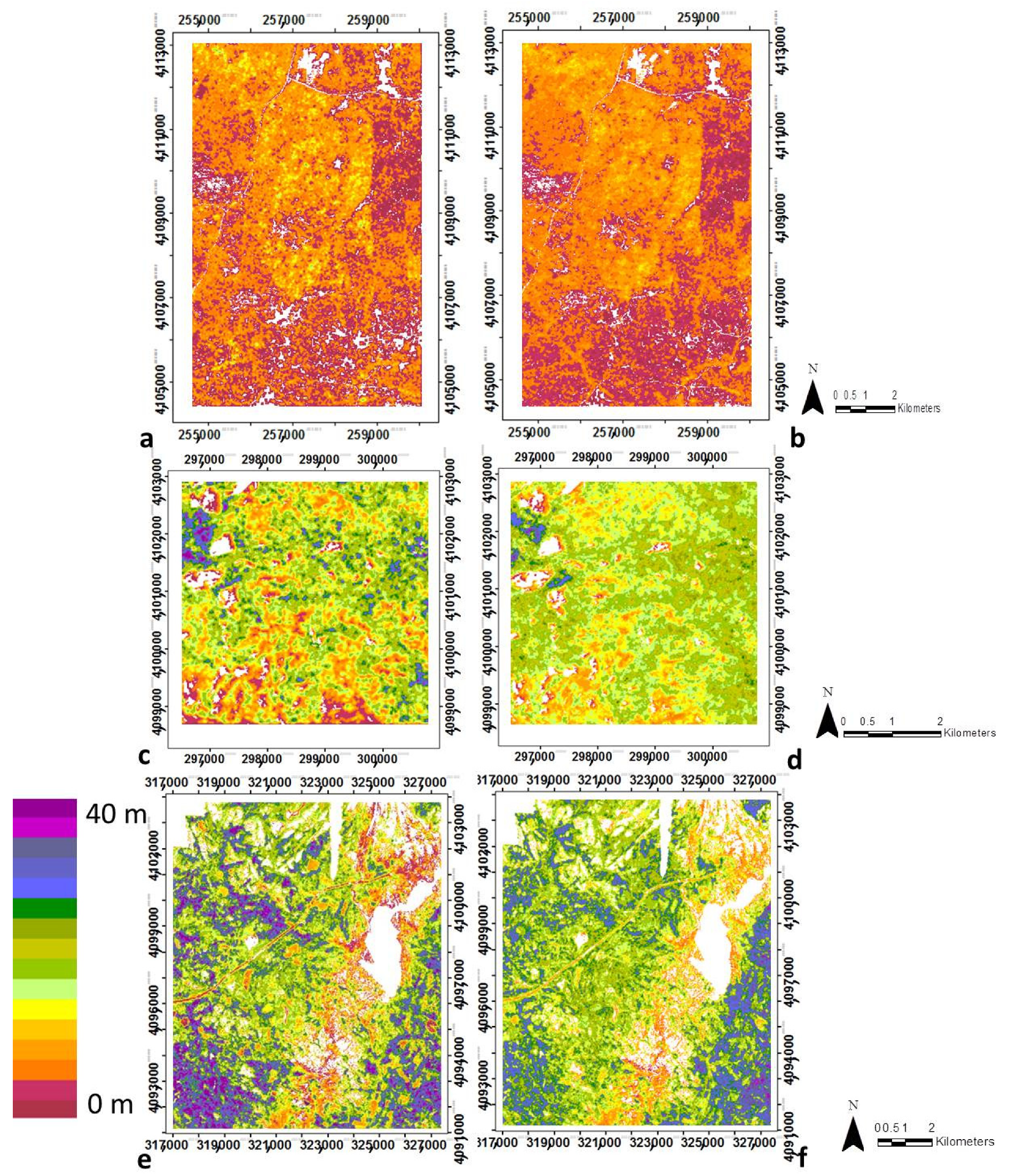
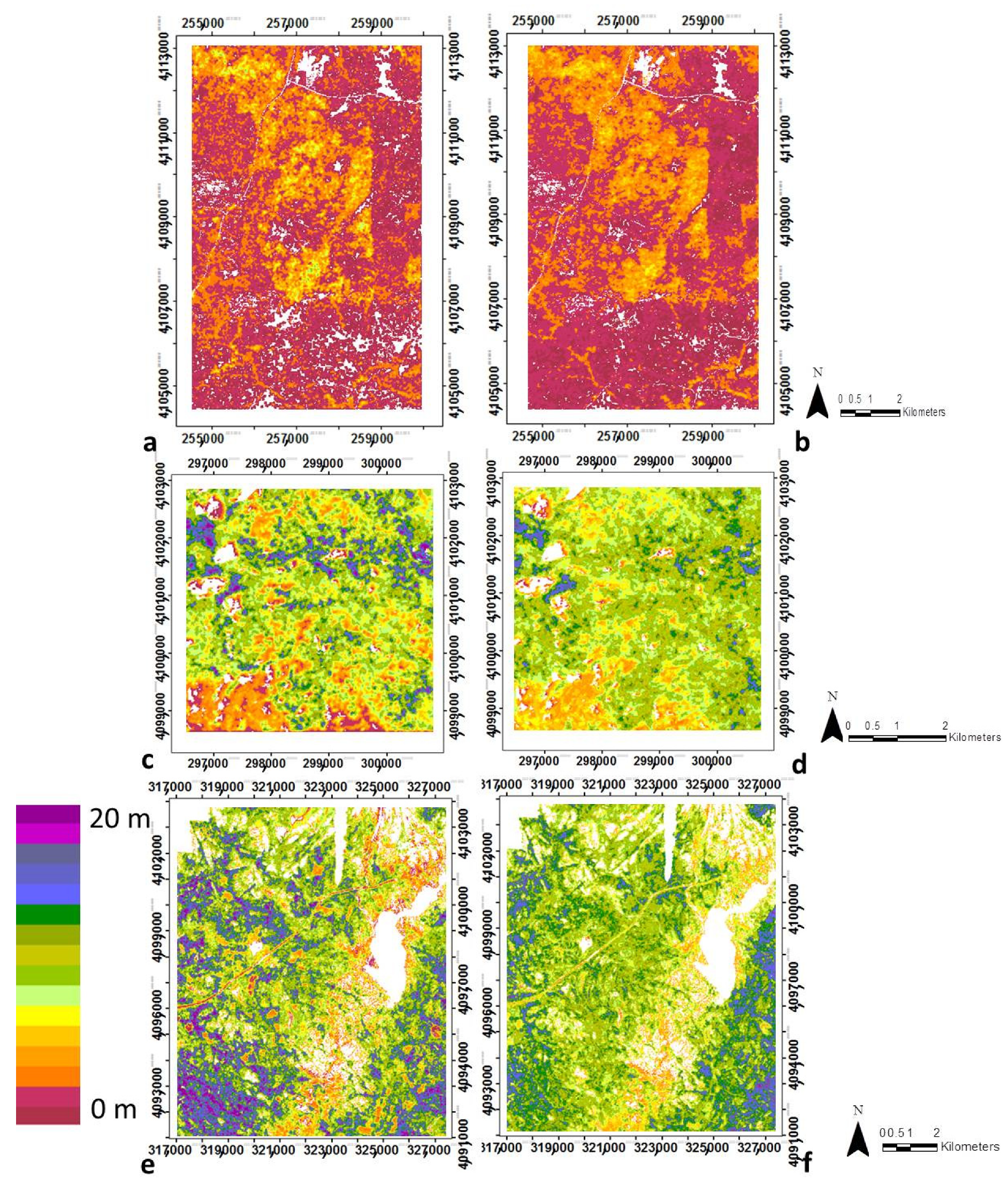
3.1. Modeling Structural Variables with Optical Metrics
3.2. Canopy Structural Types (CSTs)
3.2.1. Structural Types Defined in the San Joaquin Experimental Forest
3.2.2. Canopy Structural Types Defined in Soaproot Saddle
3.2.3. Structural Types Defined in Teakettle Experimental Forest
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Asner, G.P.; Morton, D.C.; Anderson, L.O.; Saatchi, S.S.; Espírito-Santo, F.D.B.; Palace, M.; Souza, C., Jr. Regional ecosystem structure and function: Ecological insights from remote sensing of tropical forests. Trends Ecol. Evol. 2007, 22, 414–423. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Lachenbruch, B.; Dawson, T.E. (Eds.) Size and Age-Related Changes in Tree Structure and Function; Springer: New York, NY, USA, 2011; 509p. [Google Scholar]
- Thompson, R.M.; Brose, U.; Dunne, J.A.; Hall, R.O., Jr.; Hladyz, S.; Kitching, R.L.; Martinez, N.D.; Rantala, H.; Romanuk, T.N.; Stouffer, D.B. Food webs: Reconciling the structure and function of biodiversity. Trends Ecol. Evol. 2012, 27, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Jetz, W.; Cavender-Bares, J.; Pavlick, R.; Schimel, D.; Davis, F.W.; Asner, G.P.; Guralnick, R.; Kattge, J.; Latimer, A.M.; Moorcroft, P.; et al. Monitoring plant functional diversity from space. Nat. Plants 2016, 2, 16024. [Google Scholar] [CrossRef] [PubMed]
- Arora, V.K.; Boer, G.J.; Friedlingstein, P.; Eby, M.; Jones, C.D.; Christian, J.R.; Bonan, G.; Bopp, L.; Brovkin, V.; Cadule, P.; et al. Carbon-concentration and carbon-climate feedbacks in CMIP5 earth system models. J. Clim. 2013, 26, 5289–5314. [Google Scholar] [CrossRef]
- Cox, P.M.; Pearson, D.B.; Booth, B.B.; Friedlingstein, P.; Huntingford, C.; Jones, C.D.; Luke, C.M. Sensitivity of tropical carbon to climate change constrained by carbon dioxide variability. Nature 2013, 494, 341–344. [Google Scholar] [CrossRef]
- Schimel, D.; Stephens, B.B.; Fisher, J.B. Effect of increasing CO2 on the terrestrial carbon cycle. Proc. Natl. Acad. Sci. USA 2015, 112, 436–441. [Google Scholar] [CrossRef]
- Schimel, D.S.; Asner, G.P.; Moorcroft, P.R. Observing changing ecological diversity in the Anthropocene. Front. Ecol. Environ. 2013, 11, 129–137. [Google Scholar] [CrossRef]
- Asner, G.P.; Knapp, D.E.; Boardman, J.; Green, R.O.; Kennedy-Bowdoin, T.; Eastwood, M.; Martin, R.E.; Anderson, C.; Field, C.B. Carnegie Airborne Observatory-2: Increasing science dimensionality via high-fidelity multi-sensor fusion. Remote Sens. Environ. 2017, 124, 454–466. [Google Scholar] [CrossRef]
- Schneider, F.D.; Morsdorf, F.; Schmid, B.; Petchey, O.L.; Hueni, A.; Schimel, D.S.; Schaepman, M.E. Mapping functional diversity from remotely sensed morphological and physiological forest traits. Nat. Commun. 2017, 8, 1441. [Google Scholar] [CrossRef]
- Antonarakis, A.S.; Saatchi, S.S.; Chazdon, R.L.; Moorcroft, P.R. Using lidar and radar measurements to constrain predictions of forest ecosystem structure and function. Ecol. Appl. 2011, 21, 1120–1137. [Google Scholar] [CrossRef]
- Hansen, A.J.; Rotella, J.J.; Kraska, M.P.V.; Brown, D. Spatial patterns of primary productivity in the Greater Yellowstone Ecosystem. Landsc. Ecol. 2000, 15, 505–522. [Google Scholar] [CrossRef]
- Culbert, P.D.; Radeloff, V.C.; Flather, C.H.; Kellndorfer, J.M.; Rittenhouse, C.D.; Pidgeon, A.M. The Influence of Vertical and Horizontal Habitat Structure on Nationwide Patterns of Avian Biodiversity. Auk 2013, 130, 656–665. [Google Scholar] [CrossRef]
- Melin, M.; Matala, J.; Mehtätalo, L.; Pusenius, J.; Packalen, P. Ecological dimensions of airborne laser scanning—Analyzing the role of forest structure in moose habitat use within a year. Remote Sens. Environ. 2016, 173, 238–247. [Google Scholar] [CrossRef]
- North, M.P.; Kane, J.T.; Kane, V.R.; Asner, G.P. Cover of tall trees best predicts California spotted owl habitat. For. Ecol. Manag. 2017, 405, 166–178. [Google Scholar] [CrossRef]
- Goetz, S.; Hall, F.G. Terrestrial Ecology, Carbon Cycle, Land Use /Land Cover Change, and Biodiversity (TECLUB): Priority Science Questions and Measurements. White Paper Submitted to NRC Earth Science Decadal Survey RFI. 2015 Document Available from the Authors. Available online: https://cce.nasa.gov/cce/pdfs/TECLUB_Final_Report.pdf (accessed on 1 January 2018).
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.J. Lidar Remote Sensing for Ecosystem Studies. BioScience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Drake, J.B.; Knox, R.G.; Dubayah, R.O.; Clark, D.B.; Condit, R.; Blair, J.B.; Hofton, M. Above-ground biomass estimation in closed canopy Neotropical forests using lidar remote sensing: Factors affecting the generality of relationships. Glob. Ecol. Biogeogr. 2003, 12, 147. [Google Scholar] [CrossRef]
- Hilker, T.; Wulder, M.A.; Coops, N.C. Update of forest inventory data with lidar and high spatial resolution satellite imagery. Can. J. Remote Sens. 2008, 34, 5–12. [Google Scholar] [CrossRef]
- Hopkinson, C.; Chasmer, L. Testing LiDAR models of fractional cover across multiple forest ecozones. Remote Sens. Environ. 2009, 113, 275–288. [Google Scholar] [CrossRef]
- Tang, H.; Brolly, M.; Zhao, F.; Strahler, A.H.; Schaaf, C.L.; Ganguly, S.; Zhang, G.; Dubayah, R. Deriving and validating Leaf Area Index (LAI) at multiple spatial scales through lidar remote sensing: A case study in Sierra National Forest, CA. Remote Sens. Environ. 2014, 143, 131–141. [Google Scholar] [CrossRef]
- García, M.; Gajardo, J.; Riaño, D.; Zhao, K.; Martín, P.; Ustin, S. Canopy clumping appraisal using terrestrial and airborne laser scanning. Remote Sens. Environ. 2015, 161, 78–88. [Google Scholar] [CrossRef]
- Bouvier, M.; Durrieu, S.; Fournier, R.A.; Renaud, J. Generalizing predictive models of forest inventory attributes using an area-based approach with airborne LiDAR data. Remote Sens. Environ. 2015, 156, 322–334. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Fisk, J.; Thomas, R.Q.; Dubayah, R.; Moorcroft, P.R.; Shugart, H.H. Linking models and data on vegetation structure. J. Geophys. Res. 2010, 115, G00E10. [Google Scholar] [CrossRef]
- Lu, D. Aboveground biomass estimation using Landsat TM data in the Brazilian Amazon Basin. Int. J. Remote Sens. 2005, 26, 2509–2525. [Google Scholar] [CrossRef]
- Zheng, D.; Rademacher, J.; Chen, J.; Crow, T.; Breese, M.; Le Moine, J.; Ryu, S. Estimating aboveground biomass using Landsat 7 ETM + data across a Biomass estimation managed landscape in northern Wisconsin, USA. Remote Sens. Environ. 2004, 93, 402–411. [Google Scholar] [CrossRef]
- Lu, F. The potential and challenge of remote-sensing based biomass estimation. Int. J. Remote Sens. 2006, 27, 1297–1328. [Google Scholar] [CrossRef]
- St-Onge, B.; Hu, Y.; Vega, C. Mapping the height and above-ground biomass of a mixed forest using lidar and stereo Ikonos images. Int. J. Remote Sens. 2008, 29, 3343–3364. [Google Scholar] [CrossRef]
- Hyde, P.; Dubayah, R.; Walker, W.; Blair, J.B.; Hofton, M.; Hunsaker, C. Mapping forest structure for wildlife habitat analysis using multi-sensor (LiDAR, SAR/InSAR, ETM+, Quickbird) synergy. Remote Sens. Environ. 2006, 102, 63–73. [Google Scholar] [CrossRef]
- Swatantran, A.; Dubayah, R.; Roberts, D.; Hofton, M.; Blair, J.B. Mapping biomass and stress in the Sierra Nevada using lidar and hyperspectral data fusion. Remote Sens. Environ. 2011, 115, 2917–2930. [Google Scholar] [CrossRef]
- Huesca, M.; García, M.; Roth, K.L.; Casas, A.; Ustin, S.L. Canopy structural attributes derived from AVIRIS imaging spectroscopy data in a mixed broadleaf/conifer forest. Remote Sens. Environ. 2016, 182, 208–226. [Google Scholar] [CrossRef]
- Ollinger, S.V. Sources of variability in canopy reflectance and the convergent properties of plants. New Phytol. 2011, 189, 375–394. [Google Scholar] [CrossRef]
- Blackburn, G.A. Quantifying Chlorophylls and Carotenoids at Leaf and Canopy Scales: An Evaluation of Some Hyperspectral Approaches. Remote Sens. Environ. 1998, 66, 273–285. [Google Scholar] [CrossRef]
- Kokaly, R.F. Investigating a physical basis for spectroscopic estimates of leaf nitrogen concentration. Remote Sens. Environ. 2001, 75, 153–161. [Google Scholar] [CrossRef]
- Kokaly, R.F.; Asner, G.P.; Ollinger, S.V.; Martin, M.E.; Wessman, C.A. Characterizing canopy biochemistry from imaging spectroscopy and its application to ecosystem studies. Remote Sens. Environ. 2009, 113 (Suppl. 1), S78–S91. [Google Scholar] [CrossRef]
- Clevers, J.G.P.W. Beyond NDVI: Extraction of biophysical variables from remote sensing imagery. In Land Use and Land Cover Mapping in Europe: Practices and Trends; Manakos, I., Braun, M., Eds.; Remote Sensing and Digital Image Processing; Springer: Dordrecht, The Netherlands, 2014; Volume 18, pp. 363–381. [Google Scholar]
- Asner, G.P.; Martin, R.E.; Carranza-Jiménez, L.; Sinca, F.; Tupayachi, R.; Anderson, C.B.; Martinez, P. Functional and biological diversity of foliar spectra in tree canopies throughout the Andes to Amazon region. New Phytol. 2014, 204, 127–139. [Google Scholar] [CrossRef]
- Cheng, T.; Féret, J.B.; Jacquemoud, S.; Ustin, S.L. Deriving leaf mass per area (LMA) from foliar reflectance across a variety of plant species using continuous wavelet analysis. ISPRS J. Photogramm. 2014, 87, 28–38. [Google Scholar] [CrossRef]
- Féret, J.B.; Asner, G.P. Mapping tropical forest canopy diversity using high-fidelity imaging spectroscopy. Ecol. Appl. 2014, 24, 1289–1296. [Google Scholar] [CrossRef]
- Féret, J.B.; Asner, G.P. Microtopographic controls on lowland Amazonian canopy diversity from imaging spectroscopy. Ecol. Appl. 2014, 24, 1297–1310. [Google Scholar] [CrossRef]
- Serbin, S.P.; Singh, A.; McNeil, B.E.; Kingdon, C.C.; Townsend, P.A. Spectroscopic determination of leaf morphological and biochemical traits for northern temperate and boreal tree species. Ecol. Appl. 2014, 24, 1651–1669. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Spectranomics: Emerging science and conservation opportunities at the interface of biodiversity and remote sensing. Glob. Ecol. Conserv. 2016, 8, 212–219. [Google Scholar] [CrossRef]
- Meerdink, S.K.; Roberts, D.A.; King, J.Y.; Roth, K.L.; Dennison, P.E.; Amaral, C.H.; Hook, S.J. Linking seasonal foliar traits to SWIR-TIR spectroscopy across California ecosystems. Remote Sens. Environ. 2016, 186, 322–338. [Google Scholar] [CrossRef]
- Treitz, P.M.; Howarth, P.J. Hyperspectral remote sensing for estimating biophysical parameters of forest ecosystems. Prog. Phys. Geogr. 1999, 23, 359–390. [Google Scholar] [CrossRef]
- Goodenough, D.; Li, J.; Asner, G.; Schaepman, M.; Ustin, S.; Dyk, A. Combining Hyperspectral Remote Sensing and Physical Modeling for Applications in Land Ecosystems. In Proceedings of the IEEE International Symposium on Geoscience and Remote Sensing, Denver, CO, USA, 31 July–4 August 2006; pp. 2000–2004. [Google Scholar]
- Hall, F.G.; Shimabukuro, Y.E.; Huemmrich, K.F. Remote Sensing of Forest Biophysical Structure Using Mixture Decomposition and Geometric Reflectance Models. Ecol. Appl. 1995, 5, 993–1013. [Google Scholar] [CrossRef]
- Ogunjemiyo, S.; Parker, G.; Roberts, D. Reflections in bumpy terrain: Implications of canopy surface variations for the radiation balance of vegetation. IEEE Geosci. Remote Sens. 2005, 2, 90–93. [Google Scholar] [CrossRef]
- Ahmed, O.S.; Franklin, S.E.; Wulder, M.A.; White, J.C. Characterizing stand level forest canopy cover and height using Landsat time series, samples of airborne LiDAR, and the Random Forest algorithm. ISPRS J. Photogramm. 2015, 101, 89–101. [Google Scholar] [CrossRef]
- Chan, J.C.; Paelinckx, D. Evaluation of Random Forest and Adaboost treebased ensemble classification and spectral band selection for ecotope mapping using airborne hyperspectral imagery. Remote Sens. Environ. 2008, 112, 2999–3011. [Google Scholar] [CrossRef]
- Naidoo, L.; Cho, M.A.; Mathieu, R.; Asner, G. Classification of savanna tree species, in the Greater Kruger National Park region, by integrating hyperspectral and LiDAR data in a Random Forest data mining environment. ISPRS J. Photogramm. 2012, 69, 167–179. [Google Scholar] [CrossRef]
- Pierce, A.D.; Farris, C.A.; Taylor, A.H. Use of random forests for modeling and mapping forest canopy fuels for fire behavior analysis in Lassen Volcanic National Park, California, USA. For. Ecol. Manag. 2012, 279, 77–89. [Google Scholar] [CrossRef]
- Belgiu, M.; Dragut, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- USFS 2016a. US Forest Service. Available online: http://www.fs.fed.us/psw/ef/san_joaquin/ (accessed on 1 November 2017).
- CZO. 2016: Southern Sierra Critical Zone Observatory. Available online: http://criticalzone.org/sierra/infrastructure/field-area/flux-tower-at-soaproot-saddle/ (accessed on 1 November 2017).
- USFS 2016b. US. Forest Service. Available online: http://www.fs.fed.us/psw/ef/teakettle/ (accessed on 1 November 2017).
- Wang, Z.; Schaaf, C.B.; Lewis, P.; Knyazikhin, Y.; Schull, M.A.; Strahler, A.H.; Yao, T.; Myneni, R.B.; Chopping, M.J.; Blair, B.J. Retrieval of canopy height using moderate-resolution imaging spectroradiometer (MODIS) data. Remote Sens. Environ. 2011, 115, 1595–1601. [Google Scholar] [CrossRef]
- Alexander, C.; Bøcher, P.K.; Arge, L.; Svenning, J.C. Regional-scale mapping of tree cover, height and main phenological tree types using airborne laser scanning data. Remote Sens. Environ. 2014, 147, 156–172. [Google Scholar] [CrossRef]
- Mutwiri1, F.K.; Odera, P.A.; Kinyanjui, M.J. Estimation of Tree Height and Forest Biomass Using Airborne LiDAR Data: A Case Study of Londiani Forest Block in the Mau Complex, Kenya. J. For. 2017, 7, 255–269. [Google Scholar] [CrossRef]
- Sumnall, M.J.; Hill, R.A.; Hinsley, S.A. Comparison of small-footprint discrete return and full waveform airborne lidar data for estimating multiple forest variables. Remote Sens. Environ. 2016, 173, 214–223. [Google Scholar] [CrossRef]
- Coops, N.C.; Hilker, T.; Wulder, M.A.; St-Onge, B.; Newnham, G.; Siggins, A.; Trofymow, J.A. Estimating canopy structure of Douglas-fir forest stands from discrete-return LiDAR. Trees 2007, 21, 295–310. [Google Scholar] [CrossRef]
- Hirata, Y.; Furuya, N.; Suzuki, M.; Yamamoto, H. Airborne laser scanning in forest management: Individual tree identification and laser pulse penetration in a stand with different levels of thinning. For. Ecol. Manag. 2009, 258, 752–760. [Google Scholar] [CrossRef]
- Kane, V.R.; McGaughey, R.J.; Bakker, J.D.; Gersonde, R.F.; Lutz, J.A.; Franklin, J.F. Comparisons between field- and LiDAR-based measures of stand structural complexity. Can. J. For. Res. 2010, 40, 761–773. [Google Scholar] [CrossRef]
- Magnussen, S.; Boudewyn, P. Derivations of stand heights from airborne laser scanner data with canopy-based quantile estimators. Can. J. For. Res. 1998, 28, 1016–1031. [Google Scholar] [CrossRef]
- Rooker-Jensen, J.L.; Humes, K.S.; Conner, T.; Williams, C.J.; DeGroot, J. Estimation of biophysical characteristics for highly variable mixed-conifer stands using small-footprint lidar. Can. J. For. Res. 2006, 36, 1129–1138. [Google Scholar] [CrossRef]
- Sibona, E.; Vitali, A.; Meloni, F.; Caffo, L.; Dotta, A.; Lingua, E.; Motta, R.; Garbarino, M. Direct Measurement of Tree Height Provides Different Results on the Assessment of LiDAR Accuracy. Forests 2017, 8, 7. [Google Scholar] [CrossRef]
- Su, Y.; Ma, Q.; Guo, Q. Fine-resolution forest tree height estimation across the Sierra Nevada through the integration of spaceborne LiDAR, airborne LiDAR, and optical imagery. Int. J. Digit. Earth 2017, 10, 307–323. [Google Scholar] [CrossRef]
- Andersen, H.E.; Reutebuch, S.E.; McGaughey, R.J. A rigorous assessment of tree height measurements obtained using airborne lidar and conventional field methods. Can. J. Remote Sens. 2006, 32, 355–366. [Google Scholar] [CrossRef]
- Nilsson, M. Estimation of Tree Heights and Stand Volume Using an Airborne Lidar System. Remote Sens. Environ. 1996, 56, 1–7. [Google Scholar] [CrossRef]
- Stereńczak, K.; Zasada, M. Accuracy of tree height estimation based on LIDAR data analysis. Folia For. Pol. 2011, 53, 123–129. [Google Scholar]
- Montagnoli, A.; Fusco, S.; Terzaghi, M.; Kirschbaum, A.; Pflugmacher, D.; Cohen, W.D.; Scippa, G.S.; Chiatante, D. Estimating forest aboveground biomass by low density lidar data in mixed broadleaved forests in the Italian Pre-Alps. For. Ecosyst. 2015, 2, 10. [Google Scholar] [CrossRef]
- Ritchie, J.C.; Humes, K.S.; Weltz, M.A. Laser altimeter measurements at WalnutGulch watershed, Arizona. J. Soil Water Conserv. 1995, 50, 440–442. [Google Scholar]
- Green, R.O.; Eastwood, M.L.; Sarture, C.M.; Chrien, T.G.; Aronsson, M.; Chippendale, B.J.; Faust, J.A.; Pavri, B.E.; Chovit, C.J.; Solis, M.S.; et al. Imaging spectroscopy and the Airborne Visible Infrared Imaging Spectrometer (AVIRIS). Remote Sens. Environ. 1998, 65, 227–248. [Google Scholar] [CrossRef]
- Soenen, S.A.; Peddle, D.R.; Coburn, C.A. A Modified Sun-Canopy-Sensor Topographic Correction in Forested Terrain. IEEE Trans. Geosci. Remote Sens. 2005, 43, 2148–2159. [Google Scholar] [CrossRef]
- Gu, D.; Gillespie, A. Topographic normalization of Landsat TM images of forests based on subpixel Sun-Canopy-Sensor geometry. Remote Sens. Environ. 1998, 64, 166–175. [Google Scholar] [CrossRef]
- NEON. National Ecological Observatory Network; Battelle: Boulder, CO, USA, 2016; Available online: http://data.neonscience.org/ (accessed on 1 November 2017).
- Roberts, D.A.; Gardner, M.; Church, R.; Ustin, S.; Scheer, G.; Green, R.O. Mapping chaparral in the Santa Monica Mountains using multiple endmember spectral mixture models. Remote Sens. Environ. 1998, 65, 267–279. [Google Scholar] [CrossRef]
- Roberts, D.; Halligan, K.; Dennison, P. 2007 ViperTools. Available online: http://www.vipertools.org/ (accessed on 1 June 2017).
- Roberts, D.A.; Dennison, P.E.; Gardner, M.E.; Hetzel, Y.; Ustin, S.L.; Lee, C.T. Evaluation of the potential of Hyperion for fire danger assessment by comparison to the Airborne Visible/Infrared Imaging Spectrometer. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1297–1310. [Google Scholar] [CrossRef]
- Dennison, P.E.; Halligan, K.Q.; Roberts, D.A. A comparison of error metrics and constraints for multiple endmember spectral mixture analysis and spectral angle mapper. Remote Sens. Environ. 2004, 93, 359–367. [Google Scholar] [CrossRef]
- Dennison, P.E.; Roberts, D.A. Endmember Selection for Multiple Endmember Spectral Mixture Analysis using Endmember Average RSME. Remote Sens. Environ. 2003, 87, 123–135. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. In Proceedings of the Third ERTS Symposium, NASA, Washington, DC, USA, 10–14 December 1973; Volume 1, pp. 309–317. [Google Scholar]
- Gitelson, A.; Merzlyak, M.N. Spectral Reflectance Changes Associated with Autumn Senescence of Aesculus hippocastanum L. and Acer platanoides L. Leaves. Spectral Features and Relation to Chlorophyll Estimation. J. Plant Physiol. 1994, 143, 286–292. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Datt, B. A New Reflectance Index for Remote Sensing of Chlorophyll Content in Higher Plants: Tests using Eucalyptus Leaves. J. Plant Physiol. 1999, 154, 30–36. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Gao, B. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Hunt, E.R., Jr.; Rock, B.N. Detection of changes in leaf water content using Near- and Middle-Infrared reflectances. Remote Sens. Environ. 1989, 30, 43–54. [Google Scholar]
- Daughtry, C.S.T. Discriminating crop residues from soil by shortwave infrared reflectance. Agron. J. 2001, 93, 125–131. [Google Scholar] [CrossRef]
- Clark, M.L.; Roberts, D.A. Species-Level Differences in Hyperspectral Metrics among Tropical Rainforest Trees as Determined by a Tree-Based Classifier. Remote Sens. 2012, 4, 1820–1855. [Google Scholar] [CrossRef]
- Roberts, D.A.; Green, R.O.; Adams, J.B. Temporal and spatial patterns in vegetation and atmospheric properties from AVIRIS. Remote Sens. Environ. 1997, 62, 223–240. [Google Scholar] [CrossRef]
- Ustin, S.L.; Riaño, D.; Hunt, E.R. Estimating canopy water content from spectroscopy. Isr. J. Plant Sci. 2012, 60, 9–23. [Google Scholar] [CrossRef]
- Richards, J.A. Remote Sensing Digital Image Analysis: An Introduction; Springer: Berlin, Germany, 1999; p. 240. [Google Scholar]
- Lovell, J.L.; Jupp, D.L.B.; Culvenor, D.S.; Coops, N.C. Using airborne and ground based ranging LiDAR to measure canopy structure in Australian forests. Can. J. Remote Sens. 2003, 29, 607–622. [Google Scholar] [CrossRef]
- Morsdorf, F.; Kotz, B.; Meier, E.; Itten, K.I.; Allgower, B. Estimation of LAI and fractional cover from small footprint airborne laser scanning data based on gap fraction. Remote Sens. Environ. 2006, 104, 50–61. [Google Scholar] [CrossRef]
- Palace, M.W.; Sullivan, F.B.; Ducey, M.J.; Treuhaft, R.N.; Herrick, C.; Shimbo, J.Z.; Mota-E-Silva, J. Estimating forest structure in a tropical forest using field measurements, a synthetic model and discrete return lidar data. Remote Sens. Environ. 2015, 161, 1–11. [Google Scholar] [CrossRef]
- Stark, S.C.; Leitold, V.; Wu, J.L.; Hunter, M.O.; de Castilho, C.V.; Costa, F.R.C.; McMahon, S.M.; Parker, G.G.; Shimabukuro, M.T.; Lefsky, M.A.; et al. Amazon forest carbon dynamics predicted by profiles of canopy leaf area and light environment. Ecol. Lett. 2012, 15, 1406–1414. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Ball, G.H.; Hall, D.J. Isodata: A Method of Data Analysis and Pattern Classification; Stanford Research Institute: Menlo Park, CA, USA, 1965. [Google Scholar]
- USFS 2016c. US. Department of Agriculture Forest Service. CALVEG (California Vegetation Map). Available online: http://www.fs.fed.us/r5/rsl/projects/mapping (accessed on 1 June 2017).
- Galidaki, G.; Zianis, D.; Gitas, I.; Radoglou, K.V.; Tsakiri-Strati, M.; Woodhouse, I.; Mallinis, G. Vegetation biomass estimation with remote sensing: Focus on forest and other wooded land over the Mediterranean ecosystem. Int. J. Remote Sens. 2017, 38, 1940–1966. [Google Scholar] [CrossRef]
- Kumar, L.; Sinha, P.; Taylor, S.; Alqurashi, A.F. Review of the use of remote sensing for biomass estimation to support renewable energy generation. J. Appl. Remote Sens. 2015, 9, 097696. [Google Scholar] [CrossRef]
- Morsdorf, F.; Frey, O.; Meier, E.; Itten, K.I.; Allgower, B. Assessment of the influence of flying altitude and scan angle on biophysical vegetation products derived from airborne laser scanning. Int. J. Remote Sens. 2008, 29, 1387–1406. [Google Scholar] [CrossRef]
- Næsset, E. Effects of different sensors, flying altitudes, and pulse repetition frequencies on forest canopy metrics and biophysical stand properties derived from small-footprint airborne laser data. Remote Sens. Environ. 2009, 113, 148–159. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LiDAR Metrics | IS Metrics |
|---|---|
| Maximum height (Hmax) | Green vegetation (GV) fraction, Non-Photosynthetic vegetation (NPV) fraction, soil fraction, shade fraction; [78] |
| Mean height (Hmean) | Normalized Difference Vegetation Index (NDVI); [82] |
| Median height (Hmedian) | Red Edge Normalized Difference Vegetation Index (NDVI705); [83,84] |
| Standard deviation of height (Hstd) | Modified Red Edge Normalized Difference Vegetation Index (mNDVI705); [84,85] |
| Standard deviation of Canopy Height Model (CHMstd) | Enhanced Vegetation Index (EVI); [86] |
| Fractional Cover (FC) [18,91] | Normalized Difference Water Index (NDWI); [87] |
| Fractional Cover form the first returns (FC-1rtn); [18] | Normalized Difference Infrared Index (NDII); [88] |
| Leaf Area Index (LAI); [92] | Cellulose Absorption Index (CAI); [89] |
| Vegetation Vertical Profile integral (VVIint) [93,94] | Wavelength positon of the NIR water absorption edge (Wtr1EdgeWvl), [90] |
| Clumping Index [23] | Magnitude positon of the NIR water absorption edge (Wtr1EdgeMag), [90] |
| Canopy water absorption feature between 958-1073 nm (Wtr1AbAr), [90] | |
| Canopy water absorption feature between 1105-1168 nm (Wtr2AbAr), [90] | |
| Equivalent Water Thickness (EWT); [90,91,92] | |
| Principal Component: PC1, PC2, PC1_visible, PC2_visible, PC1_NIR, PC2_NIR, PC1_SWIR1 and PC1_SWIR2 [93]. |
| Cover Type | Description |
|---|---|
| CON | Conifer forest/woodland |
| HDW | Hardwood forest/woodland |
| MIX | Mixed conifer and hardwood forest/woodland |
| SHB | Shrub |
| HEB | Herbaceous |
| BAR | Barren [Rock/Soil/Sand/ Snow] |
| WAT | Water |
| AGR | Agriculture |
| URB | Urban |
| Biomass | Height | Vegetation Heterogeneity | Clumping | ||
|---|---|---|---|---|---|
| R2(TV) | 0.83/0.81 | 0.80/0.69 | 0.78/0.72 | 0.76/0.70 | |
| RMSE (T/V) | 0.01/0.09 | 1.08/3.52 | 2.82/1.68 | 0.01/0.09 | |
| MPSE (T/V) | 15.50/19.43 | 25.36/50.75 | 25.37/36.95 | 19.99/23.57 | |
| SJER | R2 (T/V) | 0.80/0.56 | 0.67/0.40 | 0.73/0.52 | 0.73/0.53 |
| RMSE(T/V) | 0.06/0.06 | 1.01/1.12 | 0.51/0.43 | 0.06/0.04 | |
| MPSE(T/V) | 16.63/18.32 | 21.37/30.61 | 16.93/16.75 | 18.62/12.26 | |
| SOAP | R2 (T/V) | 0.66/0.35 | 0.59/0.33 | 0.60/0.52 | 0.58/0.17 |
| RMSE(T/V) | 0.06/0.08 | 2.28/2.23 | 1.25/1.20 | 0.07/0.01 | |
| MPSE(T/V) | 11.39/16.20 | 23.35/75.99 | 20.79/59.99 | 20.35/25.32 | |
| TEAK | R2 (T/V) | 0.80/0.64 | 0.78/0.57 | 0.71/0.49 | 0.72/0.44 |
| RMSE(T/V) | 0.07/0.08 | 2.94/3.15 | 1.33/1.40 | 0.07/0.08 | |
| MPSE(T/V) | 15.62/2013 | 23.26/39.75 | 22.91/27.86 | 20.33/26.76 | |
| B | H | VH | C | |
|---|---|---|---|---|
| SJER (LiDAR/IS) | ||||
| CST1 | 0.06/0.07 | 1.86/2.14 | 0.62/0.75 | < 0.5 (low) |
| CST2 | 0.18/0.17 | 4.28/4.18 | 1.49/1.46 | < 0.5 (low) |
| CST3 | 0.34/0.34 | 6.54/6.26 | 2.70/2.58 | < 0.5 (low) |
| SOAP(LiDAR/IS) | ||||
| CST1 | 0.32/0.37 | 4.96/7.27 | 3.11/4.10 | 0.30/0.30 |
| CST2 | 0.59/0.59 | 10.91/12.79 | 6.11/6.45 | 0.41/0.45 |
| CST3 | 0.71/0.68 | 8.23/9.73 | 4.32/6.10 | 0.70/0.59 |
| CST4 | 0.68/0.70 | 12.10/14.04 | 7.61/8.28 | 0.65/0.62 |
| CST5 | 0.57/0.59 | 16.33/15.87 | 8.31/8.93 | 0.44/0.50 |
| CST6 | 0.77/0.75 | 18.13/17.06 | 9.72/9.68 | 0.63/0.64 |
| TEAK(LiDAR/IS) | ||||
| CST1 | 0.30/0.23 | 7.40/6.03 | 4.29/3.42 | 0.29/0.22 |
| CST2 | 0.48/0.56 | 5.78/8.84 | 3.93/5.12 | 0.58/0.59 |
| CST3 | 0.53/0.50 | 14.08/14.32 | 7.61/7.59 | 0.37/0.39 |
| CST4 | 0.65/0.63 | 13.33/19.99 | 7.22/9.71 | 0.62/0.43 |
| CST5 | 0.69/0.71 | 22.57/20.62 | 10.75/9.87 | 0.49/0.58 |
| VCT &NVCS | SJER | SOAP | TEAK | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CST1 | CST2 | CST3 | CST1 | CST2 | CST3 | CST4 | CST5 | CST6 | CST1 | CST2 | CST3 | CST4 | CST5 | ||
| LiDAR | AGRICULTURE | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| BARREN | 0 | 0 | 0 | 17 | 1 | 0 | 0 | 0 | 0 | 17 | 14 | 2 | 2 | 0 | |
| HARDWOOD | 24 | 43 | 55 | 16 | 12 | 23 | 3 | 3 | 1 | 1 | 4 | 0 | 5 | 0 | |
| GRASS | 74 | 53 | 18 | 2 | 1 | 0 | 0 | 0 | 0 | 3 | 0 | 1 | 0 | 0 | |
| MIX | 1 | 4 | 27 | 49 | 68 | 55 | 78 | 74 | 73 | 4 | 13 | 4 | 14 | 5 | |
| SHRUB | 0 | 1 | 0 | 9 | 6 | 12 | 2 | 1 | 0 | 10 | 13 | 2 | 2 | 0 | |
| CONIFER | 0 | 0 | 0 | 7 | 11 | 9 | 17 | 22 | 26 | 66 | 55 | 90 | 77 | 94 | |
| Closed tree canopy | 29 | 65 | 75 | 81 | 79 | 90 | 10 | 21 | 32 | 59 | 70 | ||||
| Sparse tree canopy | 17 | 1 | 0 | 0 | 0 | 0 | 28 | 16 | 16 | 6 | 4 | ||||
| Imaging Spectroscopy | AGRICULTURE | 0 | 0 | 0 | 0 | 0 | |||||||||
| BARREN | 18 | 9 | 2 | 0 | 0 | ||||||||||
| HARDWOOD | 1 | 9 | 0 | 0 | 0 | ||||||||||
| GRASS | 2 | 1 | 1 | 0 | 0 | ||||||||||
| MIX | 6 | 20 | 5 | 1 | 6 | ||||||||||
| SHRUB | 9 | 10 | 3 | 1 | 0 | ||||||||||
| CONIFER | 64 | 51 | 89 | 97 | 93 | ||||||||||
| Closed tree canopy | 9 | 30 | 31 | 58 | 84 | ||||||||||
| Sparse tree canopy | 32 | 13 | 15 | 6 | 2 | ||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huesca, M.; Roth, K.L.; García, M.; Ustin, S.L. Discrimination of Canopy Structural Types in the Sierra Nevada Mountains in Central California. Remote Sens. 2019, 11, 1100. https://doi.org/10.3390/rs11091100
Huesca M, Roth KL, García M, Ustin SL. Discrimination of Canopy Structural Types in the Sierra Nevada Mountains in Central California. Remote Sensing. 2019; 11(9):1100. https://doi.org/10.3390/rs11091100
Chicago/Turabian StyleHuesca, Margarita, Keely L. Roth, Mariano García, and Susan L. Ustin. 2019. "Discrimination of Canopy Structural Types in the Sierra Nevada Mountains in Central California" Remote Sensing 11, no. 9: 1100. https://doi.org/10.3390/rs11091100
APA StyleHuesca, M., Roth, K. L., García, M., & Ustin, S. L. (2019). Discrimination of Canopy Structural Types in the Sierra Nevada Mountains in Central California. Remote Sensing, 11(9), 1100. https://doi.org/10.3390/rs11091100