Spatio-Temporal Variability in Remotely Sensed Vegetation Greenness Across Yellowstone National Park


Abstract
1. Introduction
2. Materials and Methods
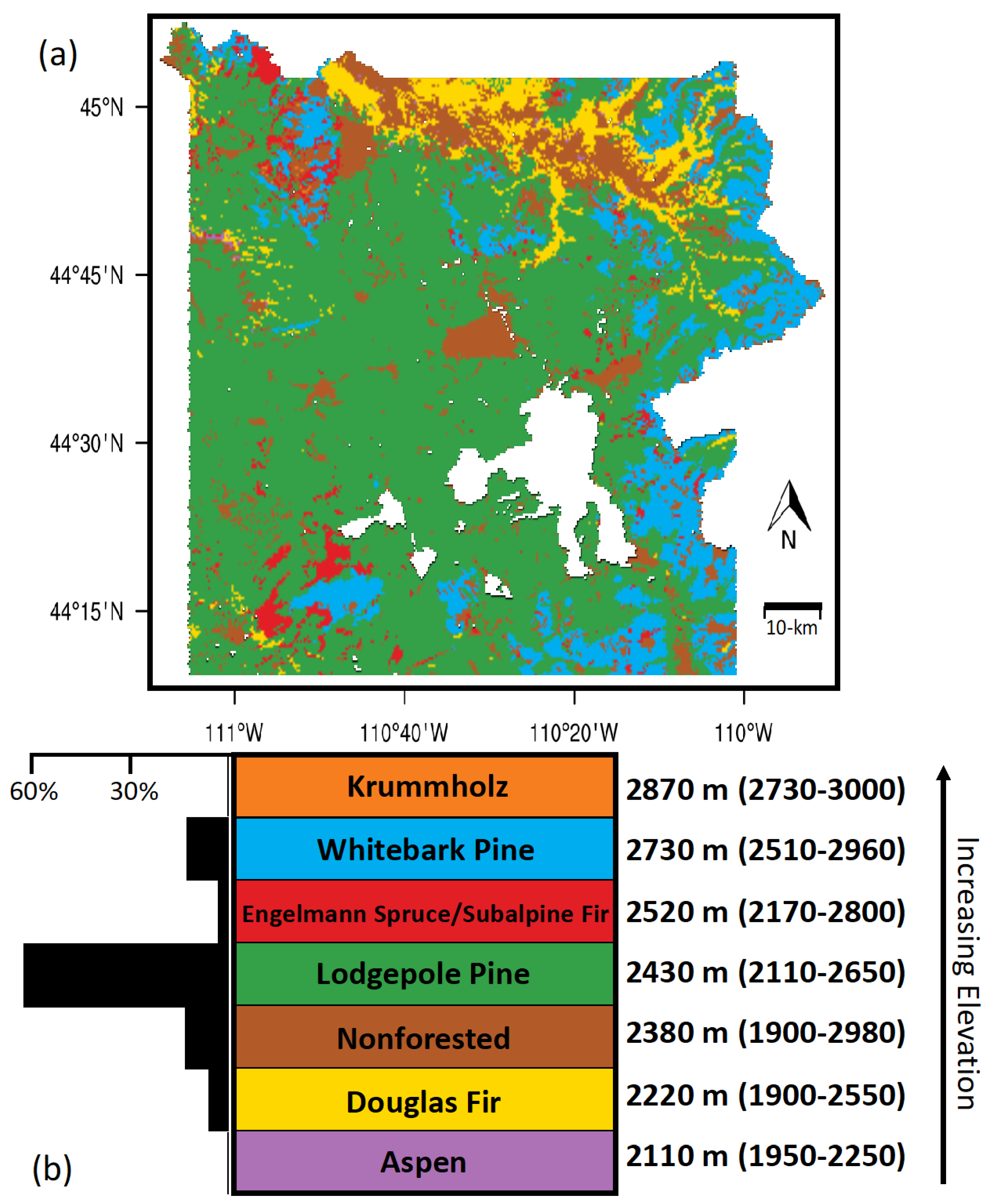
2.1. Study Area
2.2. Applied Topographic, Cover Type, Soil, Disturbance, and Climatic Datasets
2.3. Remotely Sensed Vegetation Indices
2.4. Summary of Methodology
3. Results
3.1. Focal Question 1: What are the Dominant Meteorological Drivers of Variability in Vegetation Greenness across Yellowstone National Park on the Seasonal to Interannual Time Scales?
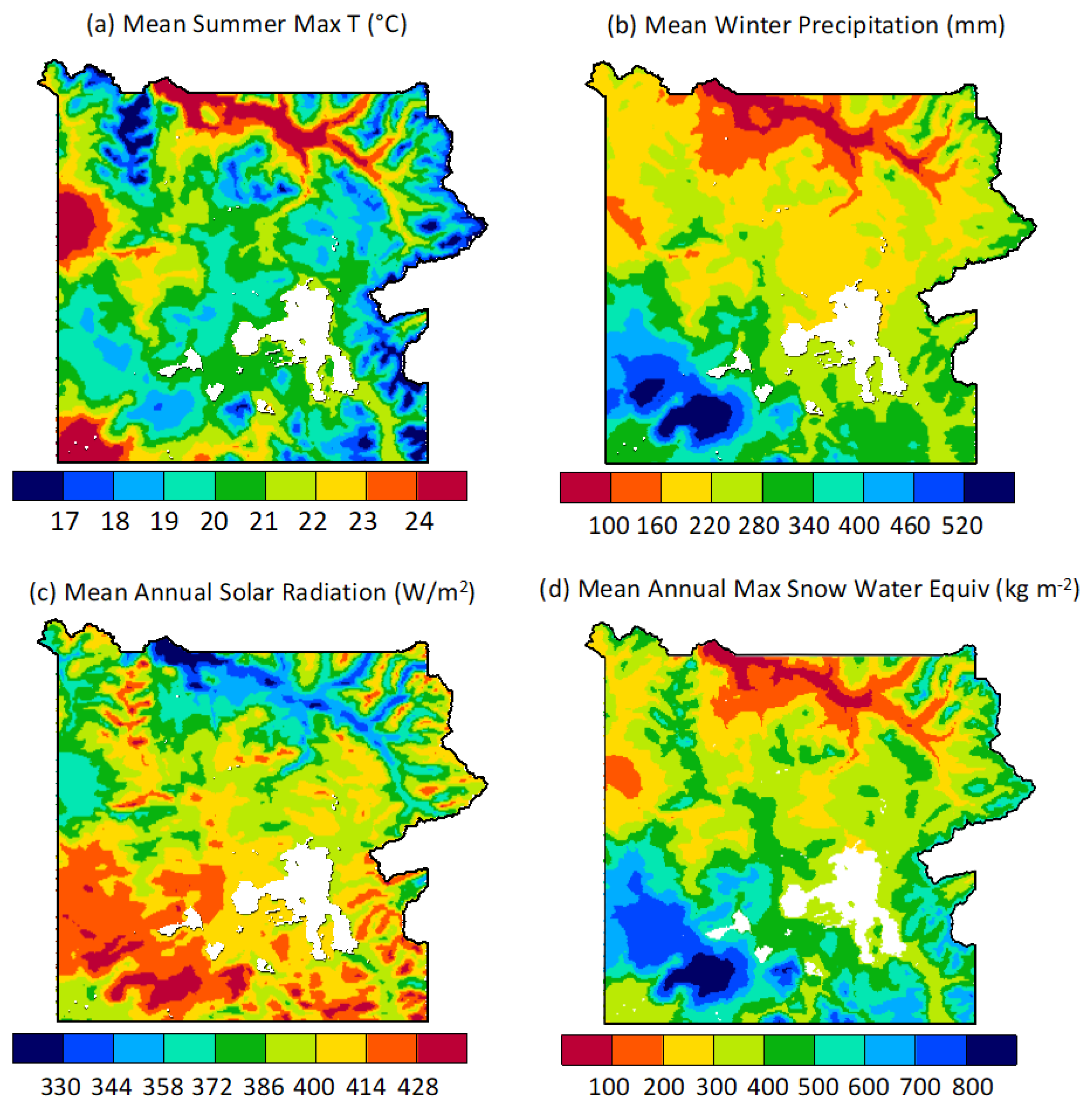
3.1.1. Climatic Heterogeneity across the Park
3.1.2. Park Summary Statistics on Vegetation Greenness Patterns
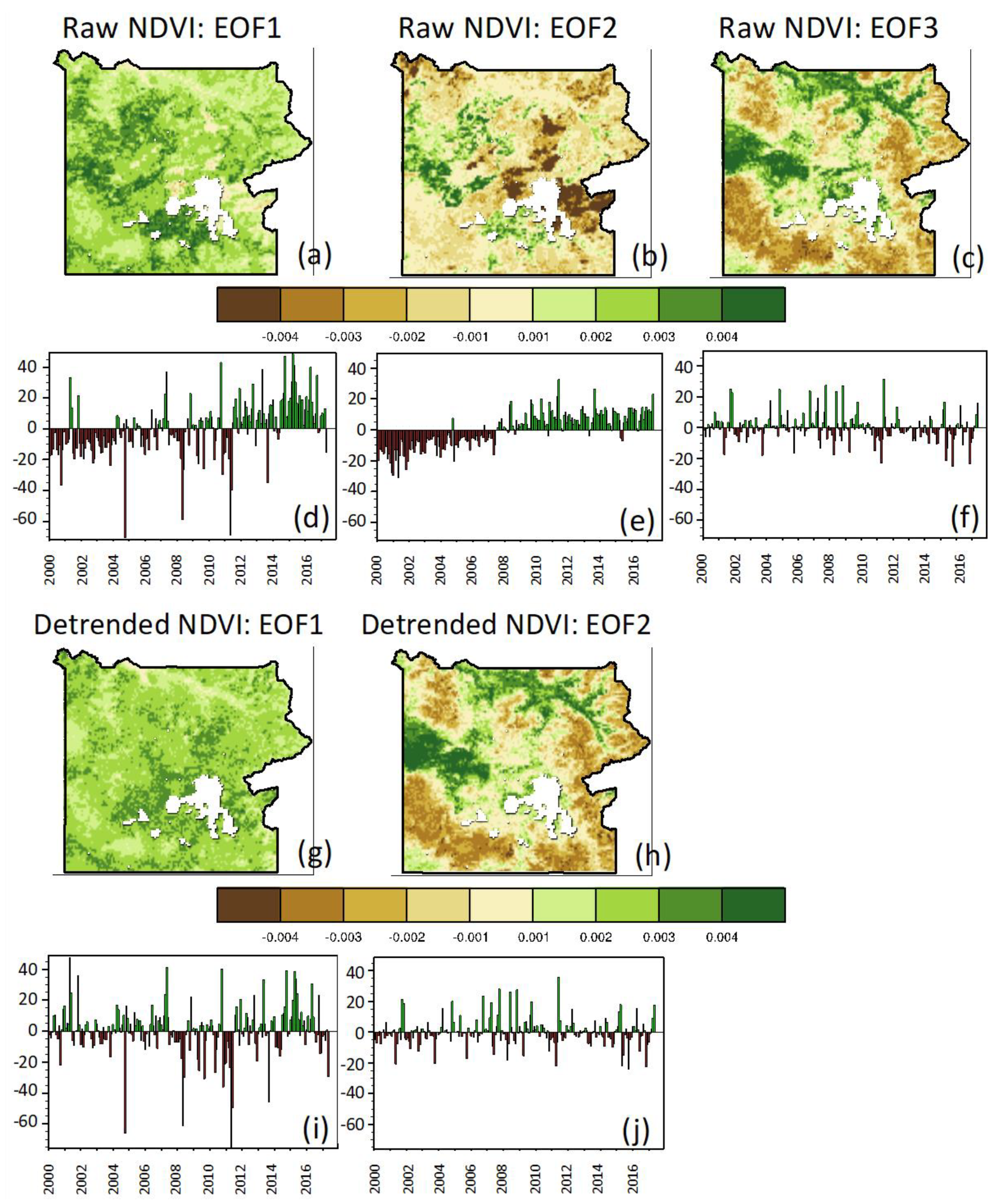
3.1.3. Spatio-Temporal Variability in Vegetation Greenness
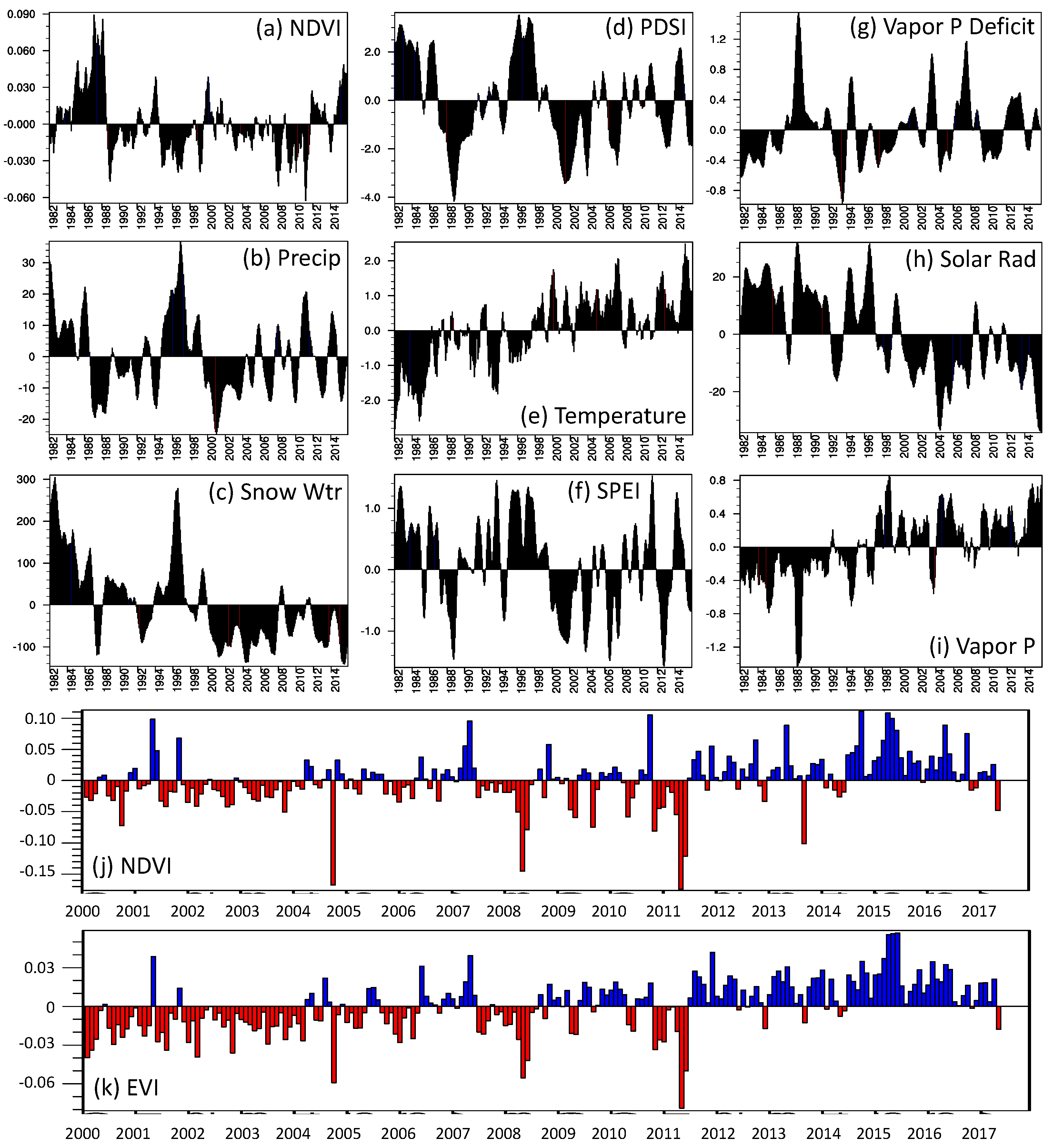
3.1.4. Climate–Phenology Correlations
3.2. Focal Question 2: How Have Patterns of Productivity Changed across the Park in Recent Decades in Response to Climate Change and Disturbances?
4. Discussion
4.1. Focal Question 1: What are the Dominant Meteorological Drivers of Variability in Vegetation Greenness across Yellowstone National Park on the Seasonal to Interannual Time Scales?
4.2. Focal Question 2: How have Patterns of Productivity Changed across the Park in Recent Decades in Response to Climate Change and Disturbances?
4.3. Comparison of Results with Past Studies
4.4. Next Research Steps
5. Conclusions
- (1)
- The general dearth of continuous, long-term ecological measurements across Yellowstone National Park serves as a call to action, particularly given its long history of 147 years as an official national park, vast spatial extent, and priceless environmental and societal value. There is a clear need for new and expanded, long-term initiatives across Yellowstone National Park, and likely many other national parks, for citizen science phenology data collection, installation of pheno-cams and soil moisture probes, and ground measurements of LAI. Such observations are valuable for identifying species’ sensitivities to climate variability and change, elucidating remotely sensed phenological changes across diverse landscapes, and assessing the relative reliability of individual satellite-derived VIs. Such observational data needs are further motivated by concerns regarding the reliability of MODIS and Landsat NDVI and EVI across Yellowstone National Park from localized studies by Franks et al. [33], Garroutte [24], and Garroutte et al. [38]. Further studies of Yellowstone National Park should consider multiple available gridded meteorological datasets, noting that Daymet is characterized by small biases in temperature, but large biases in precipitation across the conterminous United States according to Behnke et al. [144]. It is also noted that all gridded solar radiation datasets, including Daymet, exhibit substantial seasonal biases that may affect regional-scale findings [145].
- (2)
- Remote sensing has been widely underutilized across the national park system, including Yellowstone, especially given the aforementioned deficiency in continuous ground-based phenological observations. The importance of monitoring protected landscapes, including national parks, through remote sensing was stressed by Pettorelli et al. [146] and Gillespie et al. [27] and is further emphasized by the present study. Specifically, there is a clear need for a systematic, remote-sensing assessment of species’ and landscape vulnerabilities to climate variability and change, including ecological droughts operating on a spectrum of time scales, across the Western United States’ national parks and the identification of potential drought refugia. Such insights would be highly valuable to park resource managers, aiming to adapt to a highly variable and rapidly changing climate. Further remote-sensing analyses should also expand to consider higher resolution Landsat and Visible Infrared Imaging Radiometer Suite (VIRRS) data and the potential value of solar-induced chlorophyll fluorescence [147,148,149,150,151] and MODIS-based chlorophyll/carotenoid indices of foliar pigment levels for tracking evergreen photosynthesis, which is a challenge with the standard NDVI [152].
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gutzler, D.S.; Robbins, T.O. Climate variability and projected change in the western United States: Regional downscaling and drought statistics. Clim. Dyn. 2010, 37, 835–849. [Google Scholar] [CrossRef]
- Westerling, A.L. Increasing western US forest wildfire activity: Sensitivity to changes in the timing of spring. Philos. Trans. R. Soc. 2016, 371, 20150178. [Google Scholar] [CrossRef] [PubMed]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef]
- Williams, A.P.; Allen, C.D.; Millar, C.I.; Swetnam, T.W.; Michaelsen, J.; Still, C.J.; Leavitt, S.W. Forest response to increasing aridity and warmth in the southwestern United States. Proc. Natl. Acad. Sci. USA 2010, 107, 21289–21294. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat.-Clim. Chang. 2012, 3, 292–297. [Google Scholar] [CrossRef]
- Notaro, M.; Mauss, A.; Williams, J.W. Projected vegetation changes for the American Southwest: Combined dynamical modeling and bioclimatic envelope approach. Ecol. Appl. 2012, 22, 1365–1388. [Google Scholar] [CrossRef] [PubMed]
- Bassman, J.H.; Johnson, J.D.; Fins, L.; Dobrowolski, J.P. Rocky Mountain ecosystems: Diversity, complexity and interactions. Tree Physiol. 2003, 23, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary productivity from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef]
- Jarvis, J. Quote from U.S. Senate Hearing of the Energy and Natural Resources Committee. Available online: https://www.energy.senate.gov/public/ (accessed on 28 October 2009).
- Gonzalez, P. Climate change impacts and carbon in U.S. national parks. Park Sci. 2011, 28, 11–15. [Google Scholar]
- Hansen, W.D.; Turner, M.G. Origins of abrupt change? Postfire subalpine conifer regeneration declines nonlinearly with warming and drying. Ecol. Monogr. 2019, 89, 1–19. [Google Scholar] [CrossRef]
- Post, E.; Pedersen, C.; Wilmers, C.C.; Forchhammer, M.C. Warming, plant phenology and the spatial dimension of trophic mismatch for large herbivores. Proc. R. Soc. Lond. B Biol. Sci. 2008, 275, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Forrest, J.; Miller-Rushing, A.J. Toward a synthetic understanding of the role of phenology in ecology and evolution. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3101–3112. [Google Scholar] [CrossRef]
- Miller-Rushing, A.; Evenden, A.; Gross, J.; Mitchell, B.; Sachs, S. Parks use phenology to improve management and communicate climate change. Park Sci. 2011, 28, 65–71. [Google Scholar]
- Stenseth, N.C.; Mysterud, A. Climate, changing phenology, and other life history traits: non-linearity and match-mismatch to the environment. Proc. Natl. Acad. Sci. USA 2002, 99, 13379–13381. [Google Scholar] [CrossRef]
- Visser, M.E.; Both, C.; Lambrechts, M.M. Global climate change leads to mistimed avian reproduction. Adv. Ecol. Res. 2004, 35, 89–110. [Google Scholar] [CrossRef]
- Visser, M.E.; Both, C. Shifts in phenology due to global climate change: The need for a yardstick. Proc. R. Soc. Biol. Sci. 2005, 272, 2561–2569. [Google Scholar] [CrossRef]
- Aikens, E.O.; Kauffman, M.J.; Merkle, J.A.; Dwinnell, S.P.H.; Fralick, G.L.; Monteith, K.L. The greenscape shapes surfing of resource waves in a large migratory herbivore. Ecol. Lett. 2017, 20, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Rosemartin, A.H.; Crimmis, T.M.; Enquist, C.A.F.; Gerst, K.L.; Kellermann, J.L.; Posthumus, E.E.; Denny, E.G.; Guertin, P.; Marsh, L.; Weltzin, J.F. Organizing phenological data resources to inform natural resource conservation. Biol. Conserv. 2014, 173, 90–97. [Google Scholar] [CrossRef]
- Monahan, W.B.; Rosemartin, A.; Gerst, K.L.; Fisichelli, N.A.; Ault, T.; Schwartz, M.D.; Gross, J.E.; Weltzin, J.F. Climate change is advancing spring onset across the U.S. national park system. Ecosphere 2016, 7, 1–17. [Google Scholar] [CrossRef]
- Primack, R.B.; Higuchi, H.; Miller-Rushing, A.J. The impact of climate change on cherry trees and other species in Japan. Biol. Conserv. 2009, 142, 1943–1949. [Google Scholar] [CrossRef]
- Myneni, R.B.; Hall, F.G.; Sellers, P.J.; Marshak, A.L. The interpretation of spectral vegetation indices. IEEE Trans. Geosci. Remote Sens. 1995, 33, 481–486. [Google Scholar] [CrossRef]
- Glenn, E.P.; Huete, A.R.; Nagler, P.L.; Nelson, S.G. Relationship between remotely-sensed vegetation indices, canopy attributes and plant physiological processes: What vegetation indices can and cannot tell us about the landscape. Sensors 2008, 8, 2136–2160. [Google Scholar] [CrossRef] [PubMed]
- Garroutte, E.L. Spatiotemporal Variation in Grassland Biomass and Quality Across the Upper Yellowstone River Basin: Variation Across Phenology and Land Use Gradients and Validation of Remote Sensing Vegetation Indices. Master’s Thesis, Montana State University, Bozeman, MT, USA, 2012; 122p. [Google Scholar]
- Nagendra, H.; Lucas, R.; Pradinho Honrado, J.; Jongman, R.H.G.; Tarantino, C.; Adamo, M.; Mairota, P. Remote sensing for conservation monitoring: Assessing protected areas, habitat extent, habitat condition, species diversity, and threats. Ecol. Indic. 2013, 33, 45–59. [Google Scholar] [CrossRef]
- Pereira, H.M.; Ferrier, S.; Walters, M.; Geller, G.N.; Jongman, R.H.G.; Scholes, R.J.; Bruford, M.W.; Brummitt, N.; Butchart, S.H.M.; Cardoso, A.C.; et al. Essential biodiversity variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef]
- Gillespie, T.W.; Ostermann-Kelm, S.; Dong, C.; Willis, K.S.; Okin, G.S.; MacDonald, G.M. Monitoring changes of NDVI in protected areas of southern California. Ecol. Indic. 2018, 88, 485–494. [Google Scholar] [CrossRef]
- Raul Romo Leon, J.; van Leeuwen, W.J.D.; Casady, G.M. Using MODIS-NDVI for the modeling of post-wildfire vegetation response as a function of environmental conditions and pre-fire restoration treatments. Remote Sens. 2012, 4, 598–621. [Google Scholar] [CrossRef]
- White, J.D.; Swint, P. Fire effects in the northern Chihuahuan Desert derived from Landsat-5 Thematic Mapper spectral indices. J. Appl. Remote Sens. 2014, 8, 083667. [Google Scholar] [CrossRef]
- O’Leary, D.S., III; Kellermann, J.L.; Wayne, C. Snowmelt timing, phenology, and growing season length in conifer forests of Crater Lake National Park, USA. Int. J. Biometeorol. 2018, 62, 273–285. [Google Scholar] [CrossRef]
- Brown, B.G. Comparison of vegetation-topography relationships at the alpine treeline ecotone. Phys. Geogr. 1994, 15, 125–145. [Google Scholar] [CrossRef]
- Wallace, C.S.A.; Walker, J.J.; Skirvin, S.M.; Patrick-Birdwell, C.; Weltzin, J.F.; Raichle, H. Mapping presence and predicting phenological status of invasive buffelgrass in southern Arizona using MODIS, climate and citizen science observation data. Remote Sens. 2016, 8, 524. [Google Scholar] [CrossRef]
- Franks, S.; Masek, J.G.; Turner, M.G. Monitoring forest regrowth following large scale fire using satellite data—A case study of Yellowstone National Park, USA. Eur. J. Remote Sens. 2013, 46, 1–19. [Google Scholar] [CrossRef]
- Jakubauskas, M.E.; Price, K.P. Empirical relationships between structural and spectral factors of Yellowstone Lodgepole pine forests. Photogram. Eng. Remote Sens. 1997, 63, 1375–1381. [Google Scholar]
- Zhao, R.F.; Meng, R.; Huang, C.; Zhao, M.; Zhao, F.A.; Gong, P.; Yu, L.; Zhu, Z. Long-term post-disturbance forest recovery in the Greater Yellowstone ecosystem analyzed using Landsat time series stack. Remote Sens. 2016, 8, 898. [Google Scholar] [CrossRef]
- Emmett, K.D.; Renwick, K.M.; Poulter, B. Disentangling climate and disturbance effects of regional vegetation greening trends. Ecosystems 2018. [Google Scholar] [CrossRef]
- Potter, C. Vegetation cover change in Yellowstone National Park detected using Landsat satellite image analysis. J. Biodivers. Manag. For. 2015, 4. [Google Scholar] [CrossRef]
- Garroutte, L.E.; Hansen, J.A.; Lawrence, L.R. Using NDVI and EVI to map spatiotemporal variation in the biomass and quality of forage for migratory elk in the Greater Yellowstone Ecosystem. Remote Sens. 2016, 8, 404. [Google Scholar] [CrossRef]
- Van Wagtendonk, J.W.; Root, R.R. The use of multi-temporal Landsat Normalized Difference Vegetation Index (NDVI) data for mapping fuel models in Yosemite National Park, USA. Int. J. Remote Sens. 2003, 24, 1639–1651. [Google Scholar] [CrossRef]
- Soulard, C.E.; Albano, C.M.; Villarreal, M.L.; Walker, J.J. Continuous 1985–2012 Landsat monitoring to assess fire effects on meadows in Yosemite National Park, California. Remote Sens. 2016, 8, 371. [Google Scholar] [CrossRef]
- National Park Service. National Park Service Climate Change Response Strategy; National Park Service Climate Change Response Program: Fort Collins, CO, USA, 2010.
- Despain, D. Yellowstone Vegetation: Consequences of Environment and History in a Natural Setting; Boulder, Roberts Rinehart Publishers: Lanham, MD, USA, 1990. [Google Scholar]
- Whitlock, C. Postglacial vegetation and climate of Grand Teton and southern Yellowstone National Parks. Ecol. Monogr. 1993, 63, 173–198. [Google Scholar] [CrossRef]
- Bartlein, P.J.; Whitlock, C.; Shafer, S.L. Future climate in the Yellowstone National Park region and its potential impact on vegetation. Conserv. Biol. 1997, 11, 782–792. [Google Scholar] [CrossRef]
- Kokaly, R.F.; Despain, D.G.; Clark, R.N.; Livo, K.E. Mapping vegetation in Yellowstone National Park using spectral feature analysis of AVIRIS data. Remote Sens. Environ. 2003, 84, 437–456. [Google Scholar] [CrossRef]
- Schoennagel, T.; Turner, M.G.; Kashian, D.M.; Fall, A. Influence of fire regimes on lodgepole pine stand age and density across the Yellowstone National Park (USA) landscape. Landsc. Ecol. 2006, 21, 1281–1296. [Google Scholar] [CrossRef]
- Forester, J.D.; Ives, A.R.; Turner, M.G.; Anderson, D.P.; Fortin, D.; Beyer, H.L.; Smith, D.W.; Boyce, M.S. State-space models link elk movement patterns to landscape characteristics in Yellowstone National Park. Ecol. Monogr. 2007, 77, 285–299. [Google Scholar] [CrossRef]
- Wilmers, C.C.; Ram, K.; Watson, F.G.R.; White, P.J.; Smith, D.W.; Levi, T. Climate and vegetation phenology: Predicting the effects of warming temperatures. In Yellowstone’s Wildlife in Transition; White, P.J., Garrott, R.A., Plumb, G.E., Eds.; Harvard University Press: Cambridge, MA, USA, 2013; pp. 147–163. [Google Scholar]
- Chang, T.; Hansen, A.J. Historic and projected climate change in the greater Yellowstone ecosystem. Yellowstone Sci. 2015, 23, 14–19. [Google Scholar]
- Romme, W.H.; Turner, M.G. Ecological implications of climate change in Yellowstone: Moving into uncharted territory? Yellowstone Sci. 2015, 23, 6–13. [Google Scholar]
- Ault, T.R.; Macalady, A.K.; Pederson, G.T.; Betancourt, J.L.; Schwartz, M.D. Northern Hemisphere modes of variability and the timing of spring in western North America. J. Clim 2011, 24, 4003–4014. [Google Scholar] [CrossRef]
- Gonzalez, P. Climate Change and Ecological Impacts at Yellowstone National Park, USA; National Park Service, U.S. Department of the Interior: Washington, DC, USA, 2012; 22p.
- Tercek, M.; Rodman, A.; Thoma, D. Trends in Yellowstone snowpack. Yellowstone Sci. 2015, 23, 20–27. [Google Scholar]
- Schook, D.M. Climatic and Hydrologic Processes Leading to Recent Wetland Losses in Yellowstone National Park, USA. Master’s Thesis, Colorado State University, Fort Collins, CO, USA, 2012. [Google Scholar]
- Ray, A.; Sepulveda, A.; Hossack, B.; Patla, D.; Thoma, D.; Al-Chokhachy, R.; Litt, A. Monitoring Greater Yellowstone ecosystem wetlands: Can long-term monitoring help us understand their future? Yellowstone Sci. 2015, 23, 44–53. [Google Scholar]
- McMenamin, S.K.; Hadly, E.A.; Wright, C.K. Climatic change and wetland desiccation cause amphibian decline in Yellowstone National Park. Proc. Natl. Acad. Sci. USA 2008, 105, 16988–16993. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Jewett, J.T.; Lawrence, R.L.; Marshall, L.A.; Gessler, P.A.; Powell, S.L.; Savage, S.L. Spatiotemporal relationships between climate and white bark mortality in the Greater Yellowstone ecosystem. For. Sci. 2011, 57, 320–335. [Google Scholar]
- Donato, D.C.; Simard, M.; Romme, W.H.; Harvey, B.J.; Turner, M.G. Evaluating post-outbreak management effects on future fuel profiles and stand structure in bark beetle-impacted forests of Greater Yellowstone. For. Ecol. Manag. 2013, 303, 160–174. [Google Scholar] [CrossRef]
- Macfarlane, W.W.; Logan, J.A.; Kern, W.R. An innovative aerial assessment of Greater Yellowstone Ecosystem mountain pine beetle-caused whitebark pine mortality. Ecol. Appl. 2013, 23, 421–437. [Google Scholar] [CrossRef]
- Hansen, E.M.; Johnson, M.C.; Bentz, B.J.; Vandygriff, J.C.; Munson, A.S. Fuel loads and simulated fire behavior in “old stage” beetle-infested ponderosa pine of the Colorado Plateau. For. Sci. 2015, 61, 644–664. [Google Scholar] [CrossRef]
- Jakubos, B.; Romme, W.H. Invasion of subalpine meadows by lodgepole pine in Yellowstone National Park, Wyoming, U.S.A. Arctic Alpine Res. 1993, 25, 382–390. [Google Scholar] [CrossRef]
- Turner, M.G.; Romme, W.H.; Reed, R.A.; Tuskan, G.A. Postfire aspen seedling recruitment across the Yellowstone (USA) landscape. Landsc. Ecol. 2003, 18, 127–140. [Google Scholar] [CrossRef]
- Hansen, W.D.; Romme, W.H.; Ba, A.; Turner, M.G. Shifting ecological filters mediate postfire expansion of seedling aspen (Populus tremuloides) in Yellowstone. For. Ecol. Manag. 2016, 362, 218–230. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Branstator, G.W.; Arkin, P.A. Origins of the 1988 North American drought. Science 1988, 242, 1640–1645. [Google Scholar] [CrossRef]
- Renkin, R.A.; Despain, D.G. Fuel moisture, forest type, and lightning-caused fire in Yellowstone National Park. Can. J. For. Res. 1992, 22, 37–45. [Google Scholar] [CrossRef]
- Turner, M.G.; Hargrove, W.; Gardner, R.; Romme, W. Effects of fire on landscape heterogeneity in Yellowstone National Park, Wyoming. J. Veg. Sci. 1994, 5, 731–742. [Google Scholar] [CrossRef]
- Turner, M.G.; Romme, W.H.; Tinker, D.B. Surprises and lessons from the 1988 Yellowstone fires. Front. Ecol. Environ. 2003, 1, 351–358. [Google Scholar] [CrossRef]
- Bessie, W.C.; Johnson, E.A. The relative importance of fuels and weather on fire behavior in sub-alpine forests. Ecology 1995, 76, 747–762. [Google Scholar] [CrossRef]
- Schullery, P. The fires and fire policy. Bioscience 1989, 39, 686–694. [Google Scholar] [CrossRef]
- Rothermel, R.C.; Hartford, R.A.; Chase, C.H. Fire Growth Maps for the 1988 Greater Yellowstone Area Fire; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1994; p. 64.
- Daly, C.; Halbleib, M.; Smith, J.I.; Wayne, P.G.; Doggett, M.K.; Taylor, G.H.; Curtis, J.; Pasteris, P.P. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Int. J. Climatol. 2008. [Google Scholar] [CrossRef]
- Palmer, W.C. Meteorological Drought; U.S. Weather Bureau Research Paper 45; U.S. Government Printing Office: Washington, DC, USA, 1965; 65p.
- Alley, W.M. The Palmer drought severity index: Limitations and assumptions. J. Clim. Appl. Meteorol. 1984, 23, 1100–1109. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Mcevoy, D.J.; Redmond, K.T. The west wide drought tracker: Drought monitoring at fine spatial scales. Bull. Am. Meteor. Soc. 2017, 98, 1815–1820. [Google Scholar] [CrossRef]
- Chen, T.; van der Werf, G.R.; de Jeu, R.A.M.; Wang, G.; Dolman, A.J. A global analysis of the impact of drought on net primary productivity. Hydrol. Earth Syst. Sci. 2013, 17, 3885–3894. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Begueria, S.; Trigo, R.; Lopez-Moreno, J.I.; Azorin-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef]
- Daly, C. Guidelines for assessing the suitability of spatial climate data sets. Int. J. Climatol. 2006, 26, 707–721. [Google Scholar] [CrossRef]
- Thornton, P.E.; Thornton, M.M.; Mayer, B.W.; Wei, Y.; Devarakonda, R.; Vose, R.S.; Cook, R.B. Daymet: Daily Surface Weather Data on a 1-km Grid for North America, Version 3; ORNL DAAC: Oak Ridge, TN, USA, 2013. [Google Scholar] [CrossRef]
- Menne, M.J.; Durre, I.; Vose, R.S.; Gleason, B.E.; Houston, T.G. An overview of the global historical climate network-daily database. J. Atmos. Ocean. Technol. 2012, 29, 897–910. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Reed, B.C.; Schwartz, M.D.; Xiao, X. Remote sensing phenology: Status and the way forward. In Phenology of Ecosystem Processes: Applications in Global Change Research; Noormets, A., Ed.; Springer: New York, NY, USA, 2009; pp. 231–246. [Google Scholar]
- Tucker, C.J.; Vanpraet, C.L.; Sharman, M.J.; Ittersum, G.V. Satellite remote sensing of total herbaceous biomass production in the Senegalese Sahel: 1980–1984. Remote Sens. Environ. 1985, 17, 233–249. [Google Scholar] [CrossRef]
- Cabrera-Bosquet, L.; Molero, G.; Stellacci, A.M.; Bort, J.; Nogués, S.; Araus, J.L. NDVI as a potential tool for predicting biomass, plant nitrogen content and growth in wheat genotypes subjected to different water and nitrogen conditions. Cereal Res. Commun. 2011, 39, 147–159. [Google Scholar] [CrossRef]
- Tucker, C.J.; Sellers, P.J. Satellite remote sensing of primary production. Int. J. Remote Sens. 1986, 7, 1395–1416. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Epstein, H.E.; Lauenroth, W.K.; Burke, I.C. ANPP estimates from NDVI for the central grassland region of the United States. Ecology 1997, 78, 953–958. [Google Scholar] [CrossRef]
- Seaquist, J.W.; Olsson, L.; Ardö, J. A remote sensing-based primary production model for grassland biomes. Ecol. Model. 2003, 169, 131–155. [Google Scholar] [CrossRef]
- Hamel, S.; Garel, M.; Festa-Bianchet, M.; Gaillard, J.M.; Côté, S.D. Spring Normalized Difference Vegetation Index (NDVI) predicts annual variation in timing of peak faecal crude protein in mountain ungulates. J. Appl. Ecol. 2009, 46, 582–589. [Google Scholar] [CrossRef]
- Doiron, M.; Legagneux, P.; Gauthier, G.; Lévesque, E. Broad-scale satellite Normalized Difference Vegetation Index data predict plant biomass and peak date of nitrogen concentration in Arctic tundra vegetation. Appl. Veg. Sci. 2013, 16, 343–351. [Google Scholar] [CrossRef]
- Jones, C.; Maness, N.; Stone, M.; Jayasekara, R. Chlorophyll estimation using multispectral reflectance and height sensing. Trans. ASAE 2007, 50, 1867–1872. [Google Scholar] [CrossRef]
- Christianson, D.; Creel, S. Fecal chlorophyll describes the link between primary production and consumption in a terrestrial herbivore. Ecol. Appl. 2009, 19, 1323–1335. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant remote sensing vegetation indices: A review of developments and applications. J. Sens. 2017, 2017. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Kawamura, K.; Akiyama, T.; Yokota, H.; Tsutsumi, M.; Yasuda, T.; Watanabe, O.; Wang, S. Comparing MODIS vegetation indices with AVHRR NDVI for monitoring the forage quantity and quality in Inner Mongolia grassland, China. Grassl. Sci. 2005, 51, 33–40. [Google Scholar] [CrossRef]
- Sims, D.; Rahman, A.; Cordova, V.; El-Masria, B.; Baldocchi, D.; Flanagan, L.; Goldstein, A.; Hollinger, D.; Misson, L.; Schmid, H.; et al. On the use of MODIS EVI to assess gross primary productivity of North American ecosystems. J. Geophys. Res.—Biogeosci. 2006, 111. [Google Scholar] [CrossRef]
- Matsushita, B.; Yang, W.; Chen, J.; Onda, Y.; Qiu, G. Sensitivity of the Enhanced Vegetation Index (EVI) and Normalized Difference Vegetation Index (NDVI) to topographic effects: A case study in high-density cypress forest. Sensors 2007, 7, 2636–2651. [Google Scholar] [CrossRef] [PubMed]
- Sesnie, S.E.; Dickson, B.G.; Rosenstock, S.S.; Rundall, J.M. A comparison of Landsat TM and MODIS vegetation indices for estimating forage phenology in desert bighorn sheep (Ovis canadensis nelsoni) habitat in the Sonoran Desert, USA. Int. J. Remote Sens. 2011, 33, 276–286. [Google Scholar] [CrossRef]
- Chen, X.Q.; Pan, W.F. Relationships among phenological growing season, time-integrated normalized difference vegetation index and climate forcing in the temperate region of eastern China. Int. J. Climatol. 2002, 22, 1781–1792. [Google Scholar] [CrossRef]
- Pinzon, J.E.; Tucker, C.J. A non-stationary 1981–2012 AVHRR NDVI3g time series. Remote Sens. 2014, 6, 6929–6960. [Google Scholar] [CrossRef]
- Zhu, Z.; Bi, J.; Pan, Y.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.; Nemani, R.R.; Myneni, R.B. Global data sets of vegetation leaf area index (LAI)3g and fraction of photosynthetically active radiation (FPAR)3g derived from Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI3g) for the period 1981 to 2011. Remote Sens. 2013, 5, 927–948. [Google Scholar] [CrossRef]
- Didan, K.; Munoz, A.B.; Solano, R.; Huete, A. MODIS Vegetation Index User’s Guide (MOD13 Series); Version 3.0; University of Arizona: Tucson, AZ, USA, 2015. [Google Scholar]
- Miura, T.; Huete, A.R.; Yoshioka, H. Evaluation of sensor calibration uncertainties on vegetation indices for MODIS. IEEE Trans. Geosci. Remote Sens. 2000, 38, 1399–1409. [Google Scholar] [CrossRef]
- Pebesma, E.J. Multivariate geostatistics in S: the gstat package. Comput. Geosci. 2004, 30, 683–691. [Google Scholar] [CrossRef]
- Fukuoka, A. A Study of 10-Day Forecast. (A Synthetic Report); The Geophysical Magazine: Tokyo, Japan, 1951; Volume XXII, pp. 177–218. [Google Scholar]
- Lorenz, E.N. Empirical Orthogonal Functions and Statistical Weather Prediction; Technical Report, Statistical Forecast Project Report 1; Department of Meteorology, MIT: Cambridge, MA, USA, 1956; p. 49. [Google Scholar]
- Kutzbach, J.E. Empirical eigenvectors of sea-level pressure, surface temperature and precipitation complexes over North America. J. Appl. Meteorol. 1967, 6, 791–802. [Google Scholar] [CrossRef]
- Hannachi, A.; Jolliffe, I.T.; Stephenson, D.B. Empirical orthogonal functions and related techniques in atmospheric science: A review. Int. J. Climatol. 2007, 27, 1119–1152. [Google Scholar] [CrossRef]
- Wang, H.; Liu, D.; Montenegro, A.; Zhu, X. NDVI and vegetation phenology dynamics under the influence of sunshine duration on the Tibetan plateau. Int. J. Climatol. 2015, 35, 687–698. [Google Scholar] [CrossRef]
- Mann, H.B. Non-parametric tests against trend. Econometrica 1945, 13, 163–171. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods, 4th ed.; Charles Griffin: London, UK, 1975. [Google Scholar]
- Gilbert, R.O. Statistical Methods for Environmental Pollution Monitoring; Wiley, Van Nostrand Reinhold Company: New York, NY, USA, 1987; 336p. [Google Scholar]
- Whitlock, C.; Bartlein, P.J. Spatial variations of Holocene climatic change in the Yellowstone region. Quat. Res. 1993, 39, 231–238. [Google Scholar] [CrossRef]
- Tercek, M.; Rodman, A. Forecasts of 21st century snowpack and implications for snowmobile and snowcoach use in Yellowstone National Park. PLoS ONE 2016, 11, e0159218. [Google Scholar] [CrossRef] [PubMed]
- Vitasse, Y.; Bresson, C.C.; Kremer, A.; Michalet, R.; Delzon, S. Quantifying phenological plasticity to temperature in two temperate tree species. Funct. Ecol. 2010, 24, 1211–1218. [Google Scholar] [CrossRef]
- Hwang, T.; Song, C.; Vose, J.M.; Band, L.E. Topography-mediated controls on local vegetation phenology estimated from MODIS vegetation index. Landsc. Ecol. 2011, 26, 541–556. [Google Scholar] [CrossRef]
- Hwang, T.; Band, L.E.; Miniat, C.F.; Song, C.; Bolstad, P.V.; Vose, J.M.; Love, J.P. Divergent phenological response to hydroclimate variability in forested mountain watersheds. Glob. Chang. Biol. 2014, 20, 2580–2595. [Google Scholar] [CrossRef] [PubMed]
- Elmore, A.J.; Guinn, S.M.; Minsley, B.J.; Richardson, A.D. Landscape controls on the timing of spring, autumn, and growing season length in mid-Atlantic forests. Glob. Chang. Biol. 2012, 18, 656–674. [Google Scholar] [CrossRef]
- Wehner, M.; Easterling, D.R.; Lawrimore, J.H.; Heim, R.R., Jr.; Vose, R.S.; Santer, B.D. Projections of future drought in the continental United States and Mexico. J. Hydrometeorol. 2011, 12, 1359–1377. [Google Scholar] [CrossRef]
- Cook, B.I.; Ault, T.R.; Smerdon, J.E. Unprecedented 21st century drought risk in the American Southwest and Central Plains. Sci. Adv. 2015, 1. [Google Scholar] [CrossRef] [PubMed]
- Harpold, A.A.; Molotch, N.P.; Musselman, K.N.; Bales, R.C.; Kitchner, P.B.; Litvak, M.; Brooks, P.D. Soil moisture response to snowmelt timing in mixed-conifer subalpine forests. Hydrol. Process. 2015, 29, 2782–2798. [Google Scholar] [CrossRef]
- Andrews, S.F.; Flanagan, L.B.; Sharp, E.J.; Cai, T. Variation in water potential, hydraulic characteristics and water source use in montane Douglas-fir and lodgepole pine trees in southwestern Alberta and consequences for seasonal changes in photosynthetic capacity. Tree Physiol. 2012, 32, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Odum, P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Kröel-Dulay, G.; Ransijn, J.; Schmidt, I.K.; Beier, C.; De Angelis, P.; de Dato, G.; Dukes, J.S.; Emmett, B.; Estiarte, M.; Garadnai, J.; et al. Increased sensitivity to climate change in disturbed ecosystems. Nat. Commun. 2015, 6, 6682. [Google Scholar] [CrossRef]
- Sepulveda, A.J.; Tercek, M.T.; Al-Chokachy, R.; Ray, A.M.; Thoma, D.P.; Hossack, B.R.; Pederson, G.T.; Rodman, A.W.; Olliff, T. The shifting climate portfolio of the Greater Yellowstone area. PLoS ONE 2015, 10, e0145060. [Google Scholar] [CrossRef]
- Gray, S.T.; Graumlich, L.J.; Betancourt, J.L. Annual precipitation in the Yellowstone National Park region since AD 1173. Quart. Res. 2007, 68, 18–27. [Google Scholar] [CrossRef]
- Newman, W.B.; Watson, F.G.R. The central Yellowstone landscape: Terrain, geology, climate, vegetation. In The Ecology of Large Mammals in Central Yellowstone: Sixteen Years of Integrated Field Studies; Garrott, R.A., White, P.J., Watson, F.G.R., Eds.; Elsevier: San Diego, CA, USA, 2009; pp. 17–35. [Google Scholar]
- Wilmers, C.C.; Getz, W.M. Gray wolves as climate change buffers in Yellowstone. PLoS Biol. 2005, 3, e92. [Google Scholar] [CrossRef] [PubMed]
- Shuman, B. Recent Wyoming temperature trends, their drivers, and impacts in a 14,000-year context. Clim. Chang. 2012, 112, 429–447. [Google Scholar] [CrossRef]
- Changnon, S.A. The 1988 drought, barges, and diversion. Bull. Am. Meteorol. Soc. 1989, 70, 1092–1104. [Google Scholar] [CrossRef]
- Nyland, R. Patterns of lodgepole pine regeneration following the 1988 Yellowstone fires. For. Ecol. Manag. 1998, 111, 23–33. [Google Scholar] [CrossRef]
- Turner, M.G.; Whitby, T.G.; Tinker, D.B.; Romme, W.H. Twenty-four years after the Yellowstone fires: Are postfire lodgepole pine stands converging in structure and function? Ecology 2016, 97, 1260–1273. [Google Scholar] [CrossRef]
- Kashian, D.M.; Tinker, D.B.; Turner, M.G.; Scarpace, F.L. Spatial heterogeneity of lodgepole pine sapling densities following the 1988 fires in Yellowstone National Park, Wyoming, USA. Can. J. For. Res. 2004, 34, 2263–2276. [Google Scholar] [CrossRef]
- Turner, M.G.; Tinker, D.B.; Romme, W.H.; Kashian, D.M.; Litton, C.M. Landscape patterns of sapling density, leaf area, and aboveground net primary production in postfire lodgepole pine forests, Yellowstone National Park (USA). Ecosystems 2004, 7, 751–775. [Google Scholar] [CrossRef]
- Turner, M.G.; Romme, W.H.; Gardner, R.H. Prefire heterogeneity, fire severity, and early postfire plant reestablishment in subalpine forests of Yellowstone National Park, Wyoming. Int. J. Wildland Fire 1999, 9, 21–36. [Google Scholar] [CrossRef]
- Nash, M.S.; Wickham, J.; Christensen, J.; Wade, T. Changes in landscape greenness and climatic factors over 25 years (1989–2013) in the USA. Remote Sens. 2017, 9, 295. [Google Scholar] [CrossRef]
- Schrag, A.M.; Bunn, A.G.; Graumlich, L.J. Influence of bioclimatic variables on treeline conifer distribution in the Greater Yellowstone Ecosystem: Implications for species of conservation concern. J. Biogeogr. 2008, 35, 698–710. [Google Scholar] [CrossRef]
- Warwell, M.V.; Rehfeldt, G.E.; Crookston, N.L. Modeling contemporary climate profiles of whitebark pine (Pinus albicaulis) and predicting responses to global warming. In Proceedings of the Conference Whitebark Pine: A Pacific Coast Perspective, Ashland, OR, USA, 27–31 August 2007; U.S. Department of Agriculture, Forest Service, Pacific Northwest Region: Portland, OR, USA, 2007. Available online: https://www.fs.fed.us/rm/pubs_other/rmrs_2006_warwell_m001.pdf (accessed on 1 January 2017).
- Kurz, W.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyk, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef]
- Bjornlie, D.D.; Van Manen, F.T.; Ebinger, M.R.; Haroldson, M.A.; Thompson, D.J.; Costello, C.M. Whitebark pine, population density, and home-range size of grizzly bears in the Greater Yellowstone ecosystem. PLoS ONE 2014, 9, e88160. [Google Scholar] [CrossRef]
- Lanner, R.M.; Gilbert, B.K. Nutritive value of whitebark pine seeds, and the questions of their variable dormancy. In Proceedings of the International Workshop on Subalpine Stone Pines and Their Environment: The Status of our Knowledge, St. Mortiz, Switzerland, 5–11 September 1994; Schmidt, W.C., Holtmeier, F.K., Eds.; U.S. Forest Service, General Technical Report INT-GTR-309. Intermountain Research Station: Ogden, UT, USA, 1994; pp. 206–211. [Google Scholar]
- Tomback, D.F.; Kendall, K.C. Biodiversity losses: the downward spiral. In Whitebark Pine Communities: Ecology and Restoration; Tomback, D.F., Arno, S.F., Keane, R.E., Eds.; Island Press: Washington, DC, USA, 2001; pp. 243–262. [Google Scholar]
- Landenburger, L.; Lawrence, R.L.; Podruzyn, S.; Schwartz, C.C. Mapping regional distribution of a single tree species: Whitebark pine in the Greater Yellowstone ecosystem. Sensors 2008, 8, 4983–4994. [Google Scholar] [CrossRef]
- Behnke, R.; Vavrus, S.; Allstadt, A.; Albright, T.; Thogmartin, W.E.; Radeloff, V.C. Evaluation of downscaled, gridded climate data for the conterminous United States. Ecol. Appl. 2016, 26, 1338–1351. [Google Scholar] [CrossRef]
- Slater, A.G. Surface solar radiation in North America: A comparison of observations, reanalyses, satellite, and derived products. J. Hydrometeorol. 2016, 17, 401–420. [Google Scholar] [CrossRef]
- Pettorelli, N. Climate change as a main driver of ecological research. J. Appl. Ecol. 2012, 49, 542–545. [Google Scholar] [CrossRef]
- Meroni, M.; Rossini, M.; Guanter, L.; Alonso, L.; Rascher, U.; Colombo, R.; Moreno, J. Remote sensing of solar-induced chlorophyll fluorescence: Review of methods and applications. Remote Sens. Environ. 2009, 113, 2037–2051. [Google Scholar] [CrossRef]
- Frankenberg, C.; Butz, A.; Toon, G.C. Disentangling chlorophyll fluorescence from atmospheric scattering effects in O2A-band spectra of reflected sun-light. Geophys. Res. Lett. 2011, 38, L03801. [Google Scholar] [CrossRef]
- Joiner, J.; Yoshida, Y.; Vasilkov, A.P.; Yoshida, Y.; Corp, L.A.; Middleton, E.M. First observations of global and seasonal terrestrial chlorophyll fluorescence from space. Biogeosciences 2011, 8, 637–651. [Google Scholar] [CrossRef]
- Joiner, J.; Yoshida, Y.; Vasilkov, A.P.; Schaefer, K.; Jung, M.; Guanter, L.; Zhang, Y.; Garrity, S.; Middleton, E.M.; Huemmrich, K.F.; et al. The seasonal cycle of satellite chlorophyll observations and its relationship to vegetation phenology and ecosystem-atmosphere carbon exchange. Remote Sens. Environ. 2014, 152, 375–391. [Google Scholar] [CrossRef]
- Jeong, S.-J.; Schimel, D.; Frankenberg, C.; Drewry, D.T.; Fisher, J.B.; Verma, M.; Berry, J.A.; Lee, J.-E.; Joiner, J. Application of satellite solar-induced chlorophyll fluorescence to understanding large-scale variations in vegetation phenology and function over northern high latitude forests. Remote Sens. Environ. 2017, 190, 178–187. [Google Scholar] [CrossRef]
- Gamon, J.A.; Huemmrich, K.F.; Wong, C.Y.S.; Ensminger, I.; Garrity, S.; Hollinger, D.Y.; Noormets, A.; Peñuelas, J. A remotely sensed pigment index reveals photosynthetic phenology in evergreen conifers. Proc. Natl. Acad. Sci. USA 2016, 113, 13087–13092. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Period | Data | Results | Comparison |
|---|---|---|---|---|
| Kokaly et al. [45] | 1996 | Airborne Visible/Infrared Imaging Spectrometer | Whitebark pine is abundant on the slopes of Mount Washburn. | Consistent with National Park Service (NPS) YELL dataset mapping. |
| Franks et al. [33] | 1986–2006 | Landsat NDVI and band 5; field data of LAI, annual net primary productivity (ANPP), sapling density | Satellite NDVI can describe most of the variability in observed forest regrowth as seen in leaf area index (LAI) and annual net primary productivity (ANPP). | Yields credibility to satellite data in present study. |
| Chang and Hansen [49] | 1948–2010 | Parameter–elevation Relationships on Independent Slopes Model (PRISM); Topography Weather (TopoWx) | Annual temperature in Yellowstone and Grand Teton National Parks have increased at +0.16 °C decade−1. | The current study identifies a much more rapid warming in recent decades (1982–2015) of +0.71 °C decade−1. |
| Potter [37] | 1987–2010 | Landsat NDVI | NDVI over unburned areas has been decreasing due to reduced snowpack. | Both studies lack a greenhouse-induced green-up. |
| Sepulveda et al. [125] | 1948–2012 | Snow Telemetry (SNOTEL); Cooperative Observer Network | Winter and summer temperatures have risen the most. | The current study finds the most rapid warming in autumn–winter in recent decades (1982–2015). |
| Tercek et al. [53] | 1961–2012 | SNOTEL | Snow–water equivalent declined at 70% of sites. | Declining snowpack is apparent in both studies. |
| Garroutte et al. [38] | 2007–2012 | MODIS NDVI, EVI | NDVI explains more variation in forage biomass/quality than EVI. | Useful insight as the current study does not compare the two VIs’ reliability. |
| Turner et al. [132] | 1988, 2012 | Plot data of LAI, biomass, ANPP | Stand functional properties (e.g., LAI, biomass, ANPP) may converge earlier than stand structural properties (e.g., stem density/diameter) after 1988 fires. | Phenological greening trends have been rapidly occurring. |
| Emmett et al. [36] | 1989–2014 | USGS EROS Center NDVI | Fire disturbance history is a main driver of NDVI trends. The areal extent of greening exceeds that of browning in the Greater Yellowstone Ecosystem. | Both studies agree that Yellowstone is greening, largely due to the post-1988 fire recovery; trend analysis back to 1982 actually shows declining NDVI due to large positive anomalies in the early 1980s. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Notaro, M.; Emmett, K.; O’Leary, D. Spatio-Temporal Variability in Remotely Sensed Vegetation Greenness Across Yellowstone National Park. Remote Sens. 2019, 11, 798. https://doi.org/10.3390/rs11070798
Notaro M, Emmett K, O’Leary D. Spatio-Temporal Variability in Remotely Sensed Vegetation Greenness Across Yellowstone National Park. Remote Sensing. 2019; 11(7):798. https://doi.org/10.3390/rs11070798
Chicago/Turabian StyleNotaro, Michael, Kristen Emmett, and Donal O’Leary. 2019. "Spatio-Temporal Variability in Remotely Sensed Vegetation Greenness Across Yellowstone National Park" Remote Sensing 11, no. 7: 798. https://doi.org/10.3390/rs11070798
APA StyleNotaro, M., Emmett, K., & O’Leary, D. (2019). Spatio-Temporal Variability in Remotely Sensed Vegetation Greenness Across Yellowstone National Park. Remote Sensing, 11(7), 798. https://doi.org/10.3390/rs11070798