Remote Sensing of Explosives-Induced Stress in Plants: Hyperspectral Imaging Analysis for Remote Detection of Unexploded Threats



Abstract
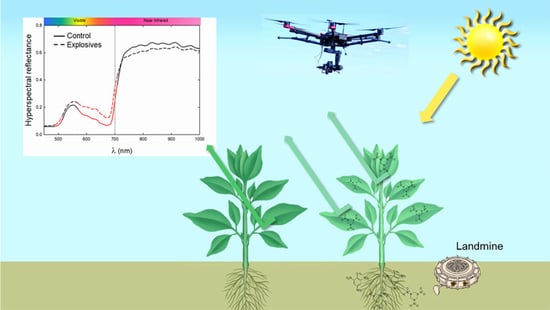
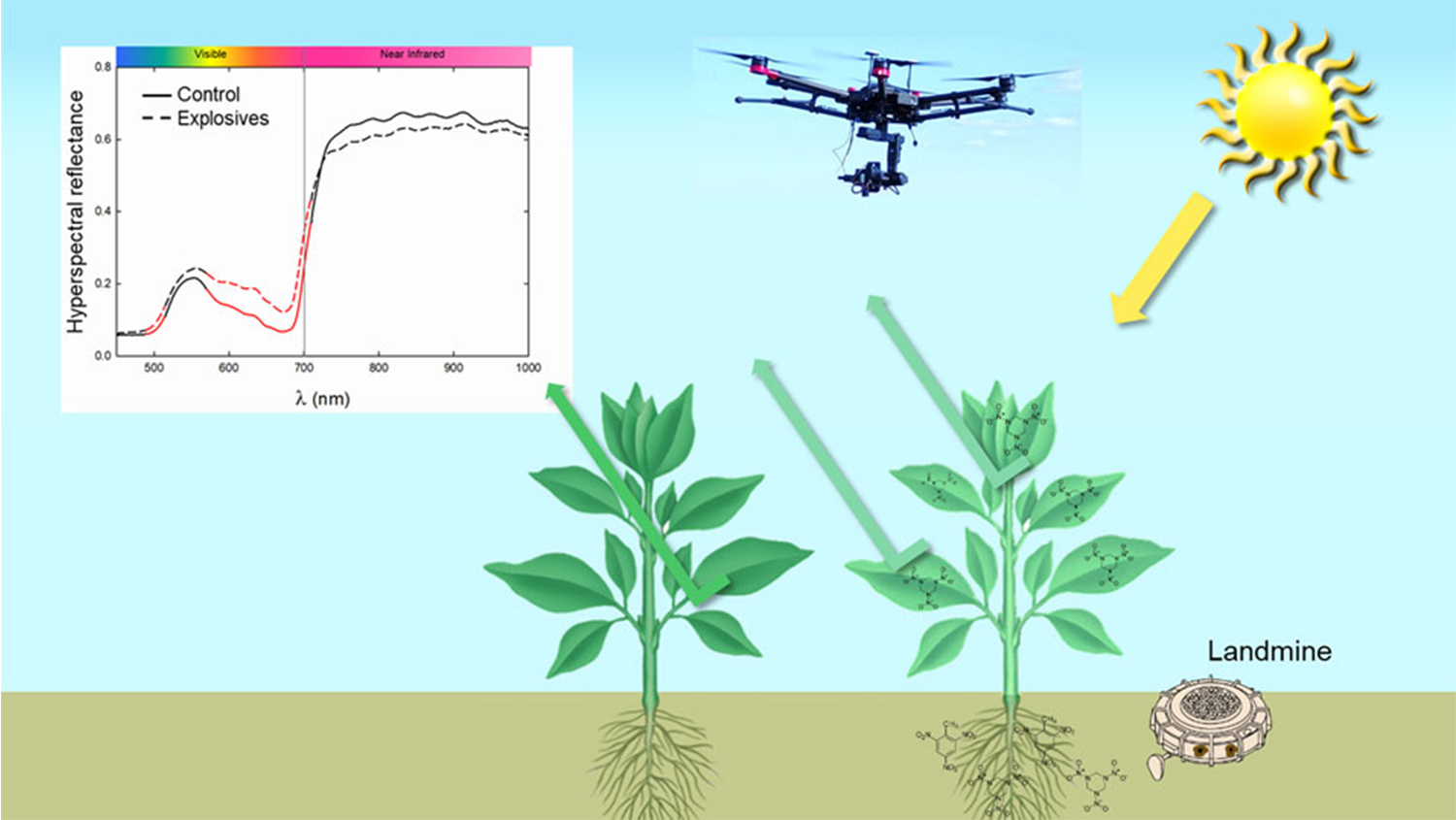
1. Introduction
2. Materials and Methods
2.1. Plants
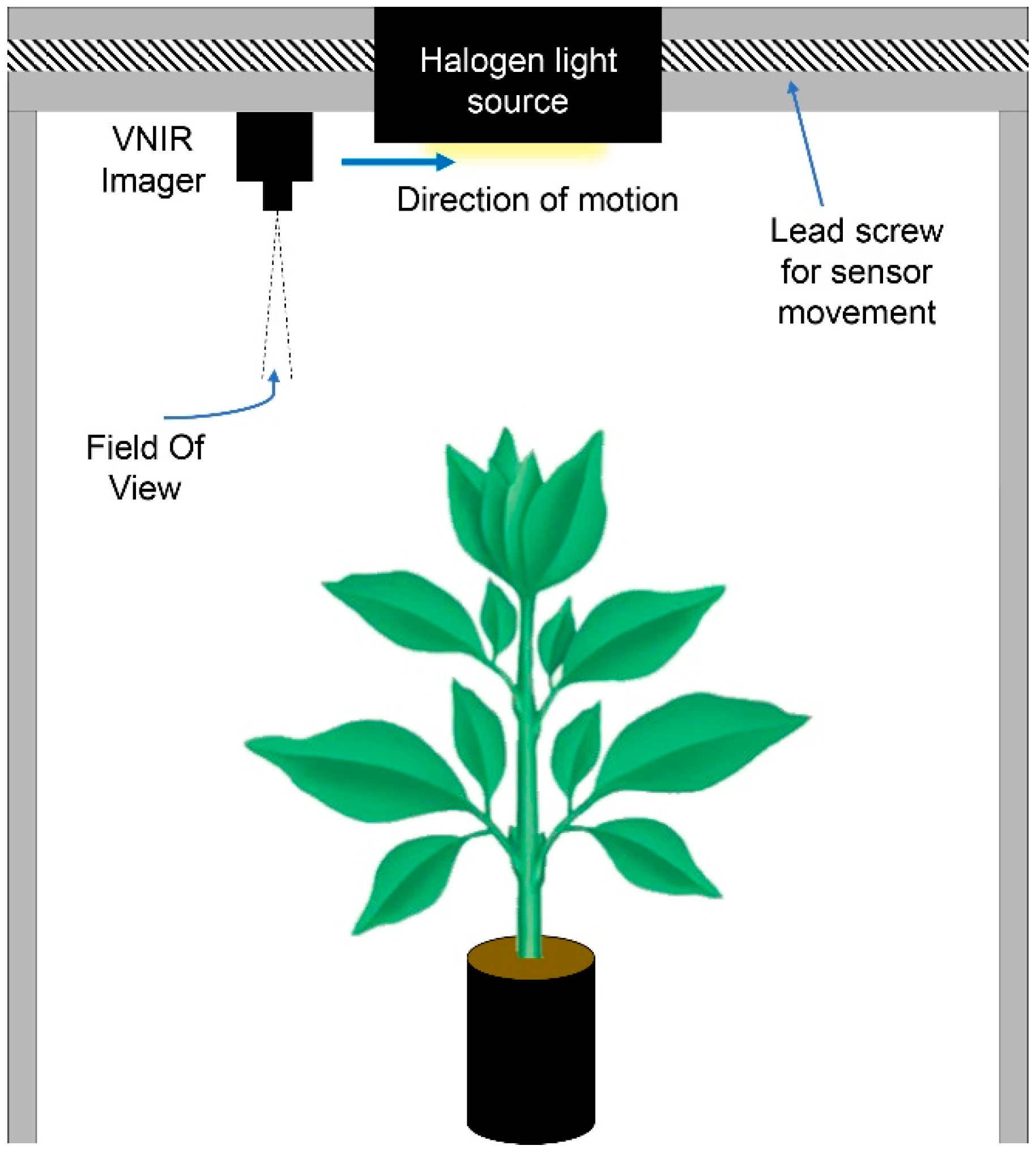
2.2. Hyperspectral Imaging
2.3. Statistics
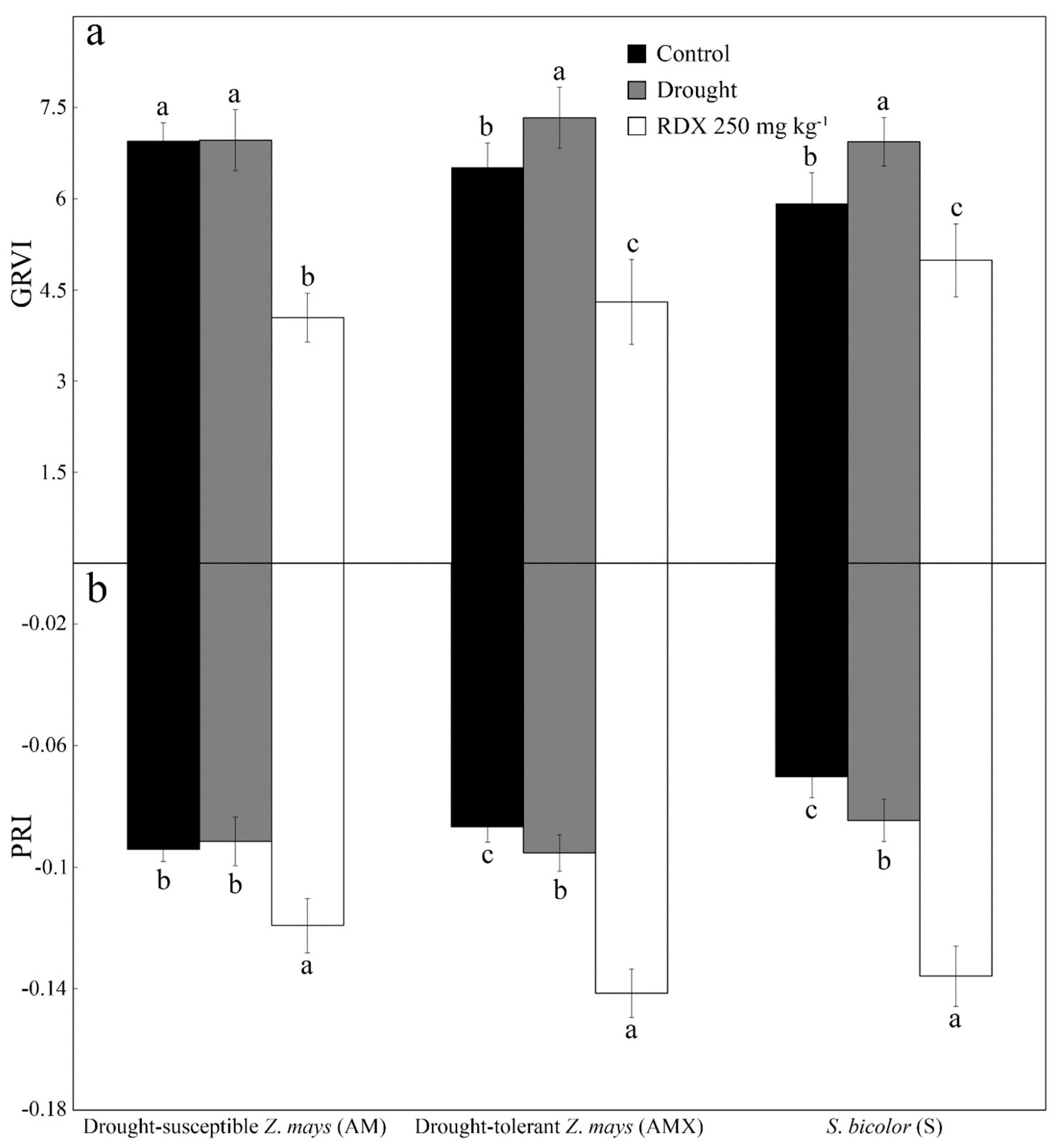
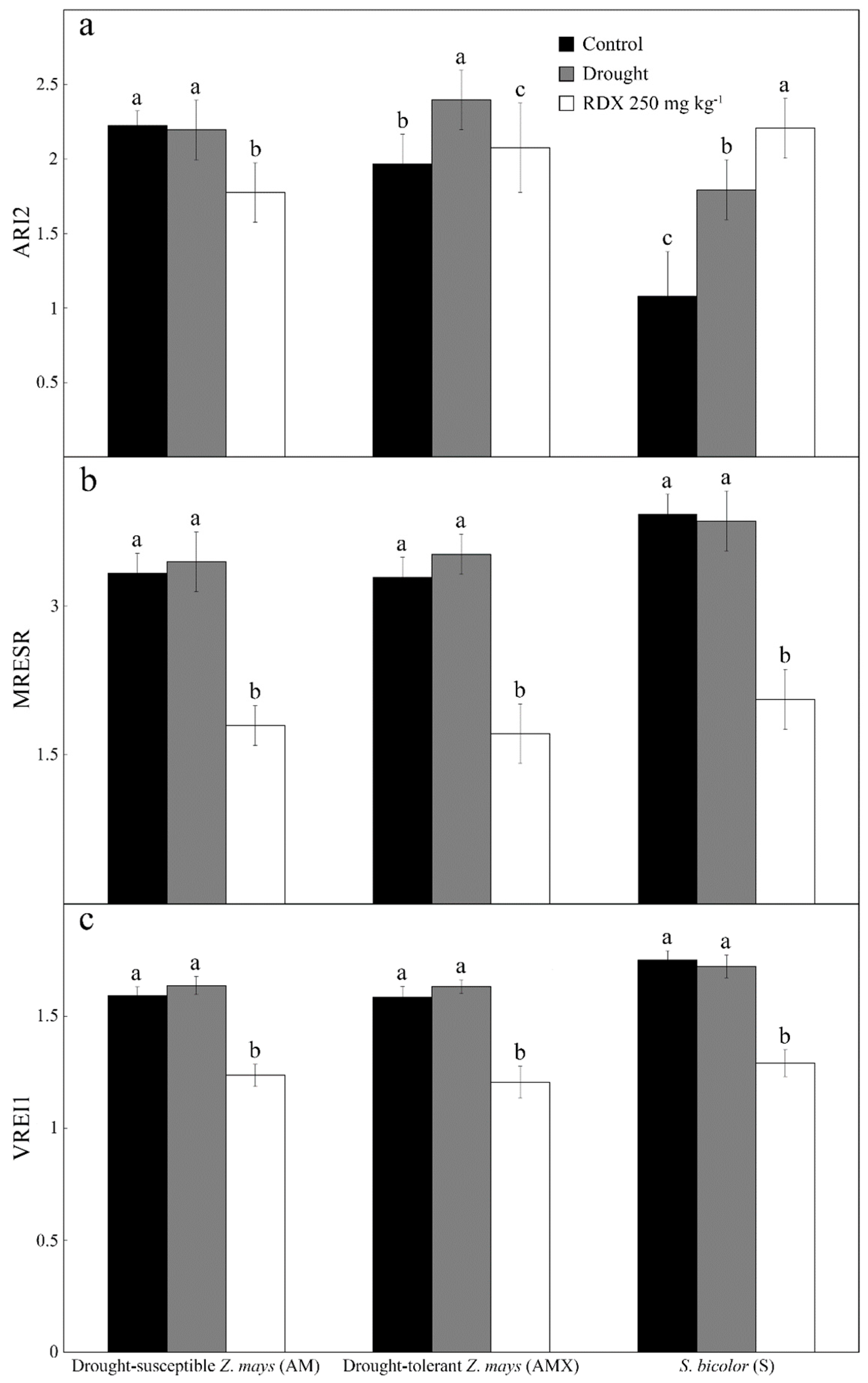
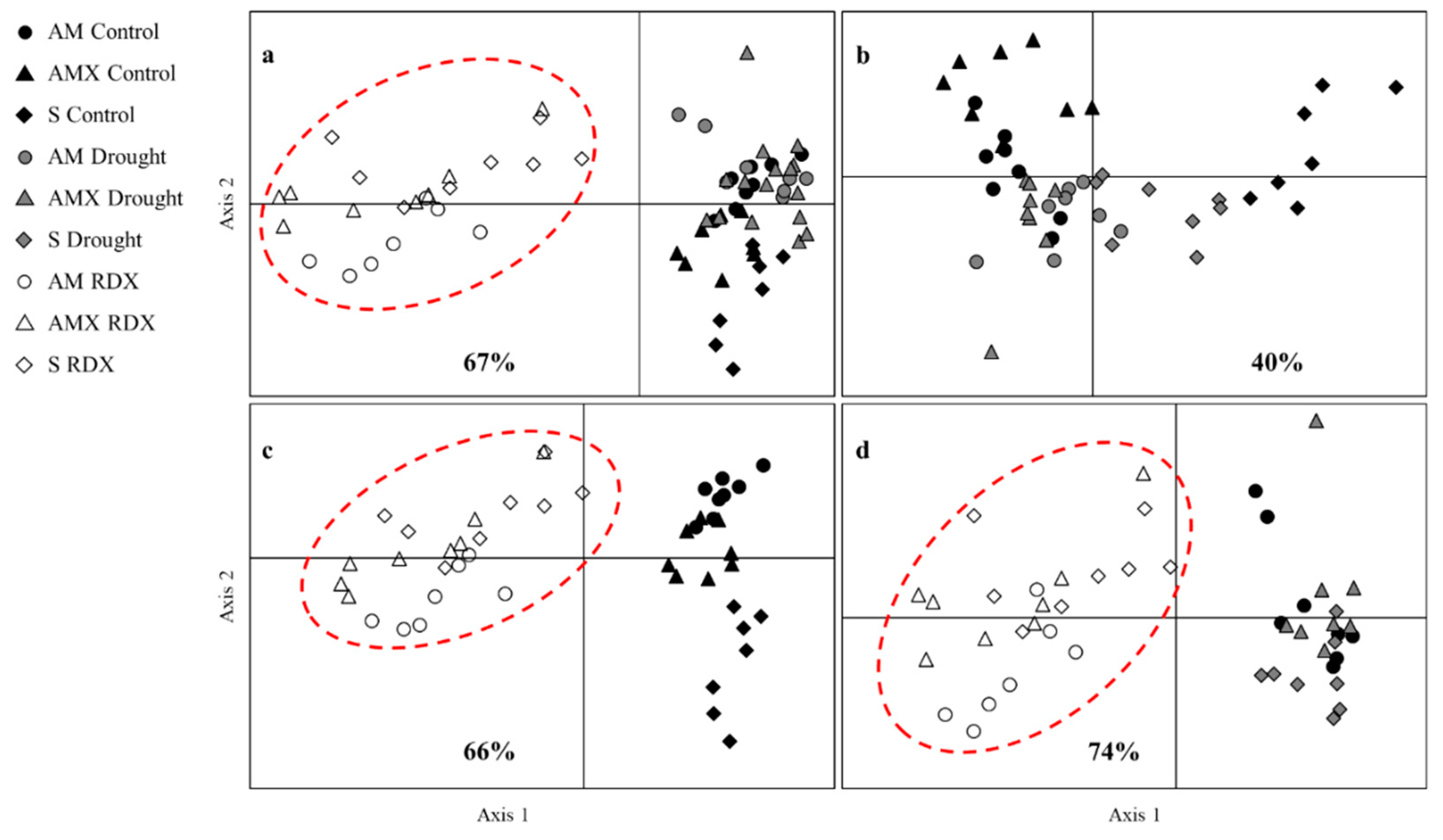
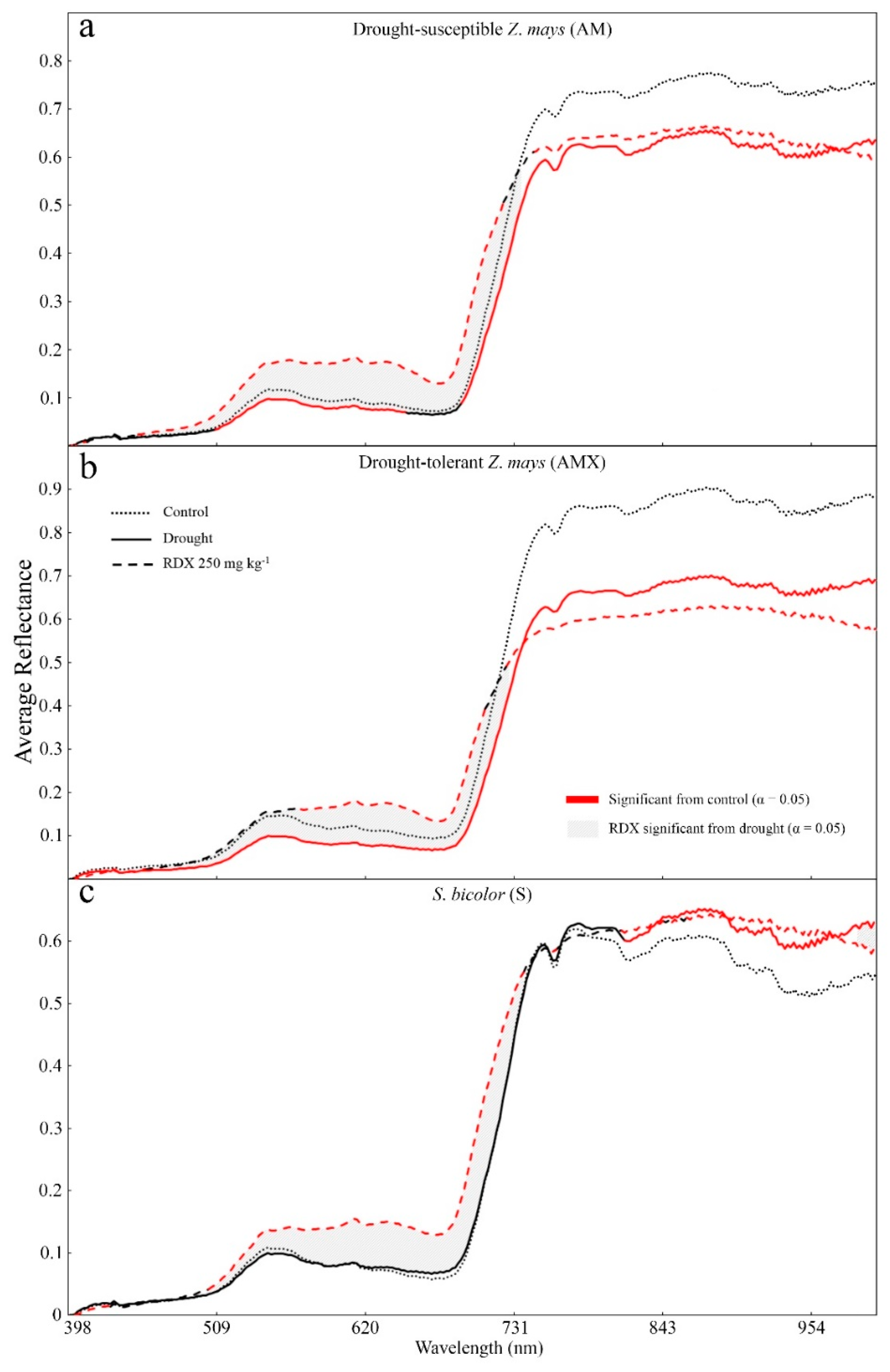
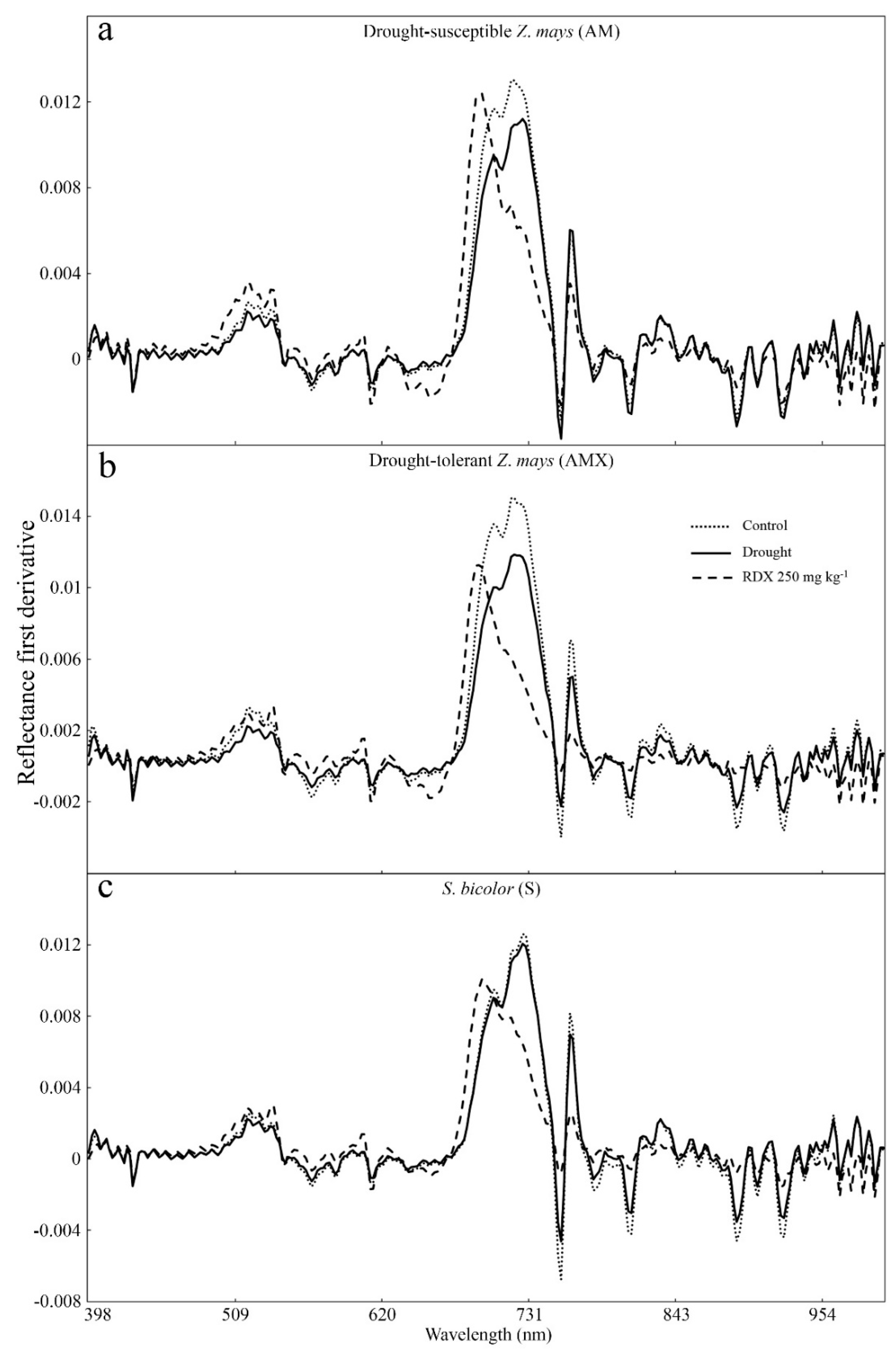
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- U.S. Environmental Protection Agency. Available online: https://cfpub.epa.gov/ncea/iris/iris_documents/documents/toxreviews/0313tr.pdf (accessed on 23 March 2019).
- Walsh, N.E.; Walsh, W.S. Rehabilitation of landmine victims—The ultimate challenge. Bull. World Health Organ. 2003, 81, 665–670. [Google Scholar] [PubMed]
- Strada, G. The horror of land mines. Sci. Am. 1996, 274, 40–45. [Google Scholar] [CrossRef]
- Rylott, E.L.; Bruce, N.C. Plants disarm soil: Engineering plants for the phytoremediation of explosives. Trends Biotechnol. 2009, 27, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Rylott, E.L.; Budarina, M.V.; Barker, A.; Lorenz, A.; Strand, S.E.; Bruce, N.C. Engineering plants for the phytoremediation of RDX in the presence of the co-contaminating explosive TNT. New Phytol. 2011, 192, 405–413. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency. Available online: https://www.epa.gov/sites/production/files/documents/ifuxoctthandbook.pdf (accessed on 7 February 2019).
- MacDonald, J.A.; Small, M.J.; Granger Morgan, M. Quantifying the risks of unexploded ordnance at closed military bases. Environ. Sci. Technol. 2009, 43, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Bigl, S.; Packer, B. Condition of in situ unexploded ordnance. Sci. Total Environ. 2015, 505, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Limmer, M.; Burken, J. Phytovolatilization of organic contaminants. Environ. Sci. Technol. 2016, 50, 6632–6643. [Google Scholar] [CrossRef] [PubMed]
- Burken, J.G.; Vroblesky, D.A.; Balouet, J.C. Phytoforensics, dendrochemistry, and phytoscreening: New green tools for delineating contaminants from past and present. Environ. Sci. Technol. 2011, 45, 6218–6226. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Singh, S.P. A review on phytoremediation of heavy metals and utilization of it’s by products. Asian J. Energy Environ. 2005, 6, 214–231. [Google Scholar]
- Singh, S.N.; Mishra, S. Phytoremediation of TNT and RDX. In Biological Remediation of Explosive Residues; Springer: Cham, Switzerland; New York, NY, USA, 2014; pp. 371–392. [Google Scholar] [CrossRef]
- Vila, M.; Lorber-Pascal, S.; Laurent, F. Fate of RDX and TNT in agronomic plants. Environ. Pollut. 2007, 148, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Zinnert, J.C. Plants as phytosensors: Physiological responses of a woody plant in response to RDX exposure and potential for remote detection. Int. J. Plant Sci. 2012, 173, 1005–1014. [Google Scholar] [CrossRef]
- Barton, C.V.M. Advances in remote sensing of plant stress. Plant Soil 2012, 354, 41–44. [Google Scholar] [CrossRef]
- Zinnert, J.C.; Via, S.M.; Young, D.R. Distinguishing natural from anthropogenic stress in plants: Physiology, fluorescence and hyperspectral reflectance. Plant Soil 2013, 366, 133–141. [Google Scholar] [CrossRef]
- Chen, D.; Liu, Z.L.; Banwart, W. Concentration-dependent RDX uptake and remediation by crop plants. Environ. Sci. Pollut. Res. 2011, 18, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Naumann, J.C.; Anderson, J.E.; Young, D.R. Remote detection of plant physiological responses to TNT soil contamination. Plant Soil 2010, 329, 239–248. [Google Scholar] [CrossRef]
- Horler, D.N.H.; Barber, J.; Darch, J.P.; Ferns, D.C.; Barringer, A.R. Approaches to detention of geochemical stress in vegetation. Adv. Space Res. 1983, 3, 175–179. [Google Scholar] [CrossRef]
- Naumann, J.C.; Anderson, J.E.; Young, D.R. Linking physiological responses, chlorophyll fluorescence and hyperspectral imagery to detect salinity stress using the physiological reflectance index in the coastal shrub, Myrica cerifera. Remote Sens. Environ. 2008, 112, 3865–3875. [Google Scholar] [CrossRef]
- Sripada, R.P.; Heiniger, R.W.; White, J.G.; Meijer, A.D. Aerial color infrared photography for determining early in-season nitrogen requirements in corn. Agron. J. 2006, 98, 968–977. [Google Scholar] [CrossRef]
- Boegh, E.; Soegaard, H.; Broge, N.; Hasager, C.; Jensen, N.; Schelde, K.; Thomsen, A. Airborne multi-spectral data for quantifying leaf area index, nitrogen concentration and photosynthetic efficiency in agriculture. Remote Sens. Environ. 2002, 81, 179–193. [Google Scholar] [CrossRef]
- Rouse, J.; Haas, R.; Schell, J.; Deering, D. Monitoring vegetation systems in the Great Plains with ERTS. In Proceedings of the Third ERTS Symposium, NASA, Washington, DC, USA, 10–14 December 1973; pp. 309–317. [Google Scholar]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Gamon, J.A.; Peñuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N.; Chivkunova, O.B. Optical properties and nondestructive estimation of anthocyanin content in plant leaves. Photochem. Photobiol. 2001, 71, 38–45. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing carotenoid content in plant leaves with reflectance spectroscopy. Photochem. Photobiol. 2002, 75, 272–281. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Remote estimation of chlorophyll content in higher plant leaves. Int. J. Remote Sens. 1997, 18, 2691–2697. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; Brown de Colstoun, E.; McMurtry, J.E., III. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Peñuelas, J.; Baret, F.; Filella, I. Semi-empirical indices to assess carotenoids/chlorophyll-a ratio from leaf spectral reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Vogelmann, J.E.; Rock, B.N.; Moss, D.M. Red edge spectral measurements from sugar maple leaves. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Rakitin, V.Y. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef]
- Curran, P.; Windham, W.; Gholz, H. Exploring the relationship between reflectance red edge and chlorophyll concentration in slash pine leaves. Tree Physiol. 1995, 15, 203–206. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Savé, R. The reflectance at the 950-970 nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Via, S.M.; Zinnert, J.C.; Young, D.R. Differential effects of two explosive compounds on seed germination and seedling morphology of a woody shrub, Morella cerifera. Ecotoxicology 2015, 24, 194–201. [Google Scholar] [CrossRef]
- Hallik, L.; Kazantsev, T.; Kuusk, A.; Galmes, J.; Tomas, M.; Niinemets, U. Generality of relationships between leaf pigment contents and spectral vegetation indices in Mallorca (Spain). Reg. Environ. Chang. 2017, 17, 2097–2109. [Google Scholar] [CrossRef]
- Gates, D.M.; Keegan, H.J.; Schleter, J.C.; Weidner, V.R. Spectral properties of plants. Appl. Opt. 1965, 4, 11–20. [Google Scholar] [CrossRef]
- Zagajewski, B.; Tommervik, H.; Bjerke, J.W.; Raczko, E.; Bochenek, Z.; Klos, A.; Jarocinska, A.; Lavender, S.; Ziolgkowski, D. Intraspecific differences in spectral reflectance curves as indicators of reduced vitality in high-arctic plants. Remote Sens. 2017, 9, 1289. [Google Scholar] [CrossRef]
- Chappelle, E.W.; Kim, M.S.; McMurtrey, J.E. The red edge shift: An explanation of its relationship to stress and the concentration of chlorophyll a. In Proceedings of the IGARSS’91 Remote Sensing: Global Monitoring for Earth Management, Espoo, Finland, 3–6 June 1991. [Google Scholar] [CrossRef]
- Sanches, I.D.; Souza Filho, C.R.; Magalhaes, L.A.; Quiterio, G.C.M.; Alves, M.N.; Oliveira, W.J. Assessing the impact of hydrocarbon leakages on vegetation using reflectance spectroscopy. ISPRS J. Photogramm. Remote Sens. 2013, 78, 85–101. [Google Scholar] [CrossRef]
- Colwell, J.E. Vegetation canopy reflectance. Remote Sens. Environ. 1974, 3, 175–183. [Google Scholar] [CrossRef]
- Curran, P.J. Remote sensing of foliar chemistry. Remote Sens. Environ. 1989, 30, 271–278. [Google Scholar] [CrossRef]
- Best, E.P.H.; Smith, T.; Hagen, F.L. Candidate Herbaceous Plants for Phytoremediation of Energetics on Ranges; Technical Report ERDC TR-07-11; U.S. Army Engineer Research and Development Center: Vicksburg, MS, USA, 2007. [Google Scholar]
- Taylor, S.; Lever, J.H.; Bostick, B.; Walsh, M.R.; Walsh, M.E.; Packer, B. Underground UXO: Are They a Significant Source of Explosives in Soil Compared to Low- and High-Order Detonation; ERDC/CRREL Technical Report TR-04-23; Cold Regions Research and Engineering Laboratory: Hanover, NH, USA, 2004. [Google Scholar]
- Rylott, E.L.; Jackson, R.G.; Edwards, J.; Womack, G.L.; Seth-Smith, H.M.B.; Rathbone, D.A.; Strand, S.E.; Bruce, N.C. An explosive-degrading cytochrome P450 activity and its targeted application for the phytoremediation of RDX. Nat. Biotechnol. 2006, 24, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Panz, K.; Miksch, K.; Sojka, T. Synergetic toxic effect of an explosive material mixture in soil. Bull. Environ. Contam. Toxicol. 2013, 91, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Haboudane, D.; Miller, J.R.; Pattev, E.; Zarco-Tejada, P.J.; Stachan, I.B. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Slonecker, T.; Fisher, G.B.; Aiello, D.P.; Haack, B. Visible and infrared remote imaging of hazardous waste: A review. Remote Sens. 2010, 2, 2474–2508. [Google Scholar] [CrossRef]
- Leeneer, I.; Pastijn, H. Selecting land mine detection strategies by means of outranking MCDM techniques. Eur. J. Oper. Res. 2002, 139, 327–338. [Google Scholar] [CrossRef]
- Habib, M.K. Controlled biological and biomimetic systems for landmine detection. Biosens. Bioelectron. 2007, 23, 1–18. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control – Drought | Plant Type | |||||
|---|---|---|---|---|---|---|
| Index | Acronym | Relates to | References | AM | AMX | S |
| Green Difference Vegetation Index | GDVI | Biomass | [21] | 0.0157 | 0.0001 | 0.0057 |
| Leaf Area Index | LAI | [22] | 0.0711 | 0.0415 | 0.8420 | |
| Normalized Difference Vegetation Index | NDVI | [23] | 0.6455 | 0.8991 | 0.9290 | |
| Green Ratio Vegetation Index | GRVI | Photosynthesis | [21] | 0.9968 | 0.0176 | 0.0027 |
| Modified Red Edge Simple Ratio | MRESR | [24] | 0.5161 | 0.1252 | 0.8249 | |
| Photochemical Reflectance Index | PRI | [25] | 0.7440 | 0.0340 | 0.0070 | |
| Anthocyanin Reflectance Index 1 | ARI1 | Pigments | [26] | 0.1553 | 0.0008 | 0.0002 |
| Anthocyanin Reflectance Index 2 | ARI2 | [26] | 0.9573 | 0.0056 | <0.0001 | |
| Carotenoid Reflectance Index 1 | CRI1 | [27] | 0.9999 | 0.0002 | 0.9985 | |
| Carotenoid Reflectance Index 2 | CRI2 | [27] | 0.9219 | 0.0002 | 0.7921 | |
| Green Normalized Difference Vegetation Index | GNDVI | [28] | 0.9999 | 0.1213 | 0.0396 | |
| Modified Chlorophyll Absorption Ratio Index | MCARI | [29] | 0.0994 | 0.0080 | 0.5899 | |
| Structure-Insensitive Pigment Index | SIPI | [30] | 0.9933 | 0.8080 | 0.9129 | |
| Vogelmann Red Edge Index 1 | VREI1 | [31] | 0.1114 | 0.1947 | 0.4924 | |
| Vogelmann Red Edge Index 2 | VREI2 | [31] | 0.0039 | 0.0602 | 0.9844 | |
| Plant Senescence Reflectance Index | PSRI | Stress | [32] | 0.4803 | 0.9719 | 0.5158 |
| Red Edge Position Index | REPI | [33] | 0.3607 | 0.6276 | 0.7286 | |
| Water Band Index | WBI | Water Content | [34] | 0.9125 | 0.8755 | <0.0001 |
| Control – RDX | Plant Type | |||||
|---|---|---|---|---|---|---|
| Index | Acronym | Relates to | References | AM | AMX | S |
| Green Difference Vegetation Index | GDVI | Biomass | [21] | 0.0002 | <0.0001 | 0.9979 |
| Leaf Area Index | LAI | [22] | <0.0001 | <0.0001 | <0.0001 | |
| Normalized Difference Vegetation Index | NDVI | [23] | <0.0001 | <0.0001 | <0.0001 | |
| Green Ratio Vegetation Index | GRVI | Photosynthesis | [21] | <0.0001 | <0.0001 | 0.0048 |
| Modified Red Edge Simple Ratio | MRESR | [24] | <0.0001 | <0.0001 | <0.0001 | |
| Photochemical Reflectance Index | PRI | [25] | <0.0001 | <0.0001 | <0.0001 | |
| Anthocyanin Reflectance Index 1 | ARI1 | Pigments | [26] | 0.5534 | 0.0054 | <0.0001 |
| Anthocyanin Reflectance Index 2 | ARI2 | [26] | 0.0008 | 0.6521 | <0.0001 | |
| Carotenoid Reflectance Index 1 | CRI1 | [27] | <0.0001 | 0.0525 | 0.0953 | |
| Carotenoid Reflectance Index 2 | CRI2 | [27] | <0.0001 | 0.4028 | 0.5756 | |
| Green Normalized Difference Vegetation Index | GNDVI | [28] | <0.0001 | <0.0001 | 0.0040 | |
| Modified Chlorophyll Absorption Ratio Index | MCARI | [29] | 0.0008 | 0.0322 | <0.0001 | |
| Structure-Insensitive Pigment Index | SIPI | [30] | 0.0017 | 0.0009 | 0.0012 | |
| Vogelmann Red Edge Index 1 | VREI1 | [31] | <0.0001 | <0.0001 | <0.0001 | |
| Vogelmann Red Edge Index 2 | VREI2 | [31] | <0.0001 | <0.0001 | <0.0001 | |
| Plant Senescence Reflectance Index | PSRI | Stress | [32] | <0.0001 | <0.0001 | <0.0001 |
| Red Edge Position Index | REPI | [33] | <0.0001 | <0.0001 | <0.0001 | |
| Water Band Index | WBI | Water Content | [34] | <0.0001 | <0.0001 | <0.0001 |
| Drought – RDX | Plant Type | |||||
|---|---|---|---|---|---|---|
| Index | Acronym | Relates to | References | AM | AMX | S |
| Green Difference Vegetation Index | GDVI | Biomass | [21] | 0.1478 | 0.0015 | 0.0050 |
| Leaf Area Index | LAI | [22] | <0.0001 | <0.0001 | <0.0001 | |
| Normalized Difference Vegetation Index | NDVI | [23] | <0.0001 | <0.0001 | <0.0001 | |
| Green Ratio Vegetation Index | GRVI | Photosynthesis | [21] | <0.0001 | <0.0001 | <0.0001 |
| Modified Red Edge Simple Ratio | MRESR | [24] | <0.0001 | <0.0001 | <0.0001 | |
| Photochemical Reflectance Index | PRI | [25] | <0.0001 | <0.0001 | <0.0001 | |
| Anthocyanin Reflectance Index 1 | ARI1 | Pigments | [26] | 0.0220 | 0.7118 | 0.0093 |
| Anthocyanin Reflectance Index 2 | ARI2 | [26] | 0.0016 | 0.0406 | 0.0064 | |
| Carotenoid Reflectance Index 1 | CRI1 | [27] | <0.0001 | <0.0001 | 0.0740 | |
| Carotenoid Reflectance Index 2 | CRI2 | [27] | <0.0001 | <0.0001 | 0.2193 | |
| Green Normalized Difference Vegetation Index | GNDVI | [28] | <0.0001 | <0.0001 | <0.0001 | |
| Modified Chlorophyll Absorption Ratio Index | MCARI | [29] | <0.0001 | <0.0001 | <0.0001 | |
| Structure-Insensitive Pigment Index | SIPI | [30] | <0.0001 | 0.0002 | 0.0022 | |
| Vogelmann Red Edge Index 1 | VREI1 | [31] | <0.0001 | <0.0001 | <0.0001 | |
| Vogelmann Red Edge Index 2 | VREI2 | [31] | <0.0001 | <0.0001 | <0.0001 | |
| Plant Senescence Reflectance Index | PSRI | Stress | [32] | <0.0001 | <0.0001 | 0.0002 |
| Red Edge Position Index | REPI | [33] | <0.0001 | <0.0001 | <0.0001 | |
| Water Band Index | WBI | Water Content | [34] | 0.1478 | 0.0015 | 0.0050 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manley, P.V.; Sagan, V.; Fritschi, F.B.; Burken, J.G. Remote Sensing of Explosives-Induced Stress in Plants: Hyperspectral Imaging Analysis for Remote Detection of Unexploded Threats. Remote Sens. 2019, 11, 1827. https://doi.org/10.3390/rs11151827
Manley PV, Sagan V, Fritschi FB, Burken JG. Remote Sensing of Explosives-Induced Stress in Plants: Hyperspectral Imaging Analysis for Remote Detection of Unexploded Threats. Remote Sensing. 2019; 11(15):1827. https://doi.org/10.3390/rs11151827
Chicago/Turabian StyleManley, Paul V., Vasit Sagan, Felix B. Fritschi, and Joel G. Burken. 2019. "Remote Sensing of Explosives-Induced Stress in Plants: Hyperspectral Imaging Analysis for Remote Detection of Unexploded Threats" Remote Sensing 11, no. 15: 1827. https://doi.org/10.3390/rs11151827
APA StyleManley, P. V., Sagan, V., Fritschi, F. B., & Burken, J. G. (2019). Remote Sensing of Explosives-Induced Stress in Plants: Hyperspectral Imaging Analysis for Remote Detection of Unexploded Threats. Remote Sensing, 11(15), 1827. https://doi.org/10.3390/rs11151827