Limitations and Challenges of MODIS-Derived Phenological Metrics Across Different Landscapes in Pan-Arctic Regions

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data and Methods
2.2.1. EC Measurement-based model GPP
2.2.2. Land Surface Phenology from Standard MODIS Product
2.2.3. Indices Derived from MODIS Surface Reflectance Products
2.2.4. Satellite Chlorophyll Fluorescence from GOME-2
2.2.5. Extraction of Phenology Metrics
2.2.6. Analysis Approach
3. Results
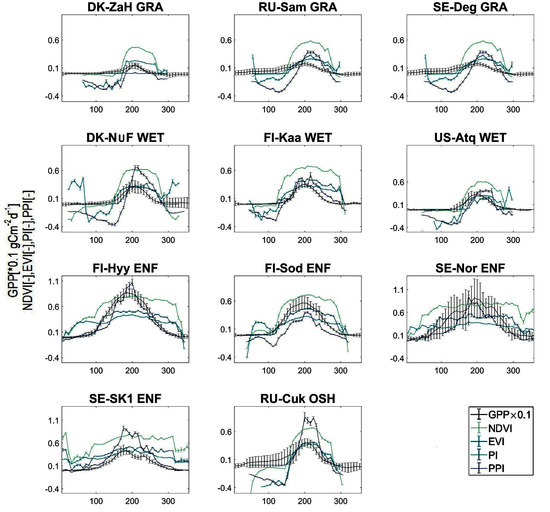
3.1. Seasonal Variations in Canopy Photosynthesis in Pan-Arctic Regions
3.2. Phenology Metrics Estimated by Different VIs
4. Discussion
4.1. Comparison of Different Vegetation Phenology Indices
4.2. Phenological Transitions in Different Landscapes
4.3. Future Prospects for Capturing Phenology Metrics in Pan-Arctic regions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

Appendix C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | LC | M value |
|---|---|---|
| DK-NuF | WET | 0.21 |
| DK-ZaH | GRA | 0.19 |
| FI-Hyy | ENF | 0.25 |
| FI-Haa | WET | 0.2 |
| FI-Sod | ENF | 0.2 |
| RU-Cuk | OSH | 0.23 |
| RU-Sam | GRA | 0.16 |
| SE-Deg | GRA | 0.25 |
| SE-Nor | ENF | 0.21 |
| SE-SK1 | ENF | 0.27 |
| US-Atq | WET | 0.21 |
| LC | MEAN | STD |
|---|---|---|
| WET | 0.207 | 0.006 |
| GRA | 0.2 | 0.046 |
| ENF | 0.233 | 0.033 |
| OSH | 0.23 | 0 |
Appendix D
| Metrics | LC | Indices | R2 | Equation | RMSE |
|---|---|---|---|---|---|
| SOS | GRA | NDVI | 0.31 | y = 0.68x + 31.59 | 94 |
| EVI | 0.41 | y = 0.47x + 84.959 | 41.02 | ||
| PI | 0.04 | y = 0.16x + 92 | 76.32 | ||
| PPI | 0.03 | y = 0.21x +105.7 | 33.17 | ||
| MCD12 | 0.4 | y = 0.99x − 19.16 | 20 | ||
| WET | NDVI | 0.43 | y = 0.58x + 64.06 | 10.4 | |
| EVI | 0.1 | y = 0.2379x + 116.09 | 59.02 | ||
| PI | 0.27 | y = 0.394x + 85.857 | 50.44 | ||
| PPI | 0.004 | y = −0.02x + 151.8 | 14.01 | ||
| MCD12 | 0.06 | y = −0.19x + 176.2 | 15.44 | ||
| ENF | NDVI | 0.17 | y = 0.24x + 68.29 | 81.6 | |
| EVI | 0.3 | y = −0.12x + 112.17 | 12.78 | ||
| PI | 0.15 | y = 0.27x + 64.21 | 83.54 | ||
| PPI | 0.07 | y = 0.2x + 68.91 | 18.27 | ||
| MCD12 | 0.22 | y = 0.62x + 16.11 | 16.75 | ||
| OSH | NDVI | 0.6 | y = -1.93x + 455.3 | 8.353 | |
| EVI | 0.13 | y = −0.29x + 200 | 13.42 | ||
| PI | 0.06 | y = −0.39x + 218.1 | 14.97 | ||
| PPI | 0.08 | y = 0.25x + 118.6 | 13.87 | ||
| MCD12 | 0.04 | y = −0.29x + 209.3 | 11.95 | ||
| EOS | GRA | NDVI | 0.19 | y = 0.41x + 155.8 | 32.81 |
| EVI | 0.11 | y = 0.25x + 199.5 | 54.31 | ||
| PI | 0.11 | y = 0.17x + 229.06 | 38.89 | ||
| PPI | 0.01 | y = −0.07x + 290.5 | 20.35 | ||
| MCD12 | 0.14 | y = 0.54x + 133.1 | 22.16 | ||
| WET | NDVI | 0.03 | y = −0.10x + 291.67 | 91.9 | |
| EVI | 0.09 | y = 0.18x + 206.6 | 77.25 | ||
| PI | 0.36 | y = 0.31x + 177.66 | 32.72 | ||
| PPI | 0.02 | y = −0.06x + 275.6 | 11.99 | ||
| MCD12 | 0.13 | y = 0.25x + 193.7 | 11.49 | ||
| ENF | NDVI | 0.24 | y = −0.23x + 357.75 | 9.07 | |
| EVI | 0.08 | y = −0.13x + 324.9 | 55.48 | ||
| PI | 0.28 | y = −0.20x + 349.16 | 6.99 | ||
| PPI | 0.04 | y = −0.08x + 313.5 | 12.4 | ||
| MCD12 | 0.09 | y = 0.23x + 232.1 | 14.2 | ||
| OSH | NDVI | 0.09 | y = 0.4x + 155.2 | 14.4 | |
| EVI | 0.01 | y = 0.03x + 255.3 | 14.9 | ||
| PI | 0.11 | y = 0.16x + 221.8 | 13.73 | ||
| PPI | 0.68 | y = 0.43x + 157.5 | 7.73 | ||
| MCD12 | 0.04 | y = −0.37x + 358.2 | 13.77 |
References
- Peñuelas, J.; Filella, I. Phenology feedbacks on climate change. Science 2009, 324, 887–888. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.F. Phenology: Spring greening in a warming world. Nature 2015, 526, 48–49. [Google Scholar] [CrossRef] [PubMed]
- Hogda, K.A.; Tommervik, H.; Karlsen, S.R. Trends in the Start of the Growing Season in Fennoscandia 1982–2011. Remote. Sens. 2013, 5, 4304–4318. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M.; Price, D.T.; Kurz, W.A.; Wu, C. Land surface phenology from optical satellite measurement and CO2eddy covariance technique. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M. Circumpolar vegetation dynamics product for global change study. Remote. Sens. Environ. 2016, 182, 13–26. [Google Scholar] [CrossRef]
- Tang, J.; Körner, C.; Muraoka, H.; Piao, S.; Shen, M.; Thackeray, S.J.; Yang, X. Emerging opportunities and challenges in phenology: A review. Ecosphere 2016, 7, e01436. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hufkens, K.; Milliman, T.; Frolking, S. Intercomparison of phenological transition dates derived from the PhenoCam Dataset V1.0 and MODIS satellite remote sensing. Sci. Rep. 2018, 8, 5679. [Google Scholar] [CrossRef] [PubMed]
- D’Odorico, P.; Gonsamo, A.; Gough, C.M.; Bohrer, G.; Morison, J.; Wilkinson, M.; Hanson, P.J.; Gianelle, D.; Fuentes, J.D.; Buchmann, N. The match and mismatch between photosynthesis and land surface phenology of deciduous forests. Agric. For. Meteorol. 2015, 214–215, 25–38. [Google Scholar] [CrossRef]
- Henebry, G.M.; de Beurs, K.M. Remote Sensing of Land Surface Phenology: A Prospectus; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Zhang, X.; Wang, J.; Gao, F.; Liu, Y.; Schaaf, C.; Friedl, M.; Yu, Y.; Jayavelu, S.; Gray, J.; Liu, L.; et al. Exploration of scaling effects on coarse resolution land surface phenology. Remote. Sens. Environ. 2017, 190, 318–330. [Google Scholar] [CrossRef]
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. Boil. Sci. 2010, 365, 3227–3246. [Google Scholar] [CrossRef] [PubMed]
- Garrity, S.R.; Bohrer, G.; Maurer, K.D.; Mueller, K.L.; Vogel, C.S.; Curtis, P.S. A comparison of multiple phenology data sources for estimating seasonal transitions in deciduous forest carbon exchange. Agric. For. Meteorol. 2011, 151, 1741–1752. [Google Scholar] [CrossRef]
- Noormets, A.; Chen, J.; Gu, L.; Desai, A. The Phenology of Gross Ecosystem Productivity and Ecosystem Respiration in Temperate Hardwood and Conifer Chronosequences. In Phenology of Ecosystem Processes: Applications in Global Change Research; Noormets, A., Ed.; Springer: New York, NY, USA, 2009; pp. 59–85. [Google Scholar]
- White, K.; Pontius, J.; Schaberg, P. Remote sensing of spring phenology in northeastern forests: A comparison of methods, field metrics and sources of uncertainty. Remote. Sens. Environ. 2014, 148, 97–107. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, Z.; Liu, Z.; Zeng, Z.; Ciais, P.; Huang, M.; Liu, Y.; Piao, S. Seasonal and interannual changes in vegetation activity of tropical forests in Southeast Asia. Agric. For. Meteorol. 2016, 224, 1–10. [Google Scholar] [CrossRef]
- Luo, X.; Chen, X.; Wang, L.; Xu, L.; Tian, Y. Modeling and predicting spring land surface phenology of the deciduous broadleaf forest in northern China. Agric. For. Meteorol. 2014, 198–199, 33–41. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Joiner, J.; Yoshida, Y.; Vasilkov, A.P.; Schaefer, K.; Jung, M.; Guanter, L.; Zhang, Y.; Garrity, S.; Middleton, E.M.; Huemmrich, K.F.; et al. The seasonal cycle of satellite chlorophyll fluorescence observations and its relationship to vegetation phenology and ecosystem atmosphere carbon exchange. Remote. Sens. Environ. 2014, 152, 375–391. [Google Scholar] [CrossRef]
- Walther, S.; Voigt, M.; Thum, T.; Gonsamo, A.; Zhang, Y.; Kohler, P.; Jung, M.; Varlagin, A.; Guanter, L. Satellite chlorophyll fluorescence measurements reveal large-scale decoupling of photosynthesis and greenness dynamics in boreal evergreen forests. Glob. Chang. Boil. 2016, 22, 2979–2996. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Cheng, X.; Li, X.; Chen, J.; Sun, M.; Ji, M.; He, H.; Wang, S.; Li, S.; Tang, J. Seasonal patterns of canopy photosynthesis captured by remotely sensed sun-induced fluorescence and vegetation indexes in mid-to-high latitude forests: A cross-platform comparison. Sci. Total. Environ. 2018, 644, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sensing of Environment. Remote. Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Nagai, S.; Nasahara, K.N.; Muraoka, H.; Akiyama, T.; Tsuchida, S. Field experiments to test the use of the normalized-difference vegetation index for phenology detection. Agric. For. Meteorol. 2010, 150, 152–160. [Google Scholar] [CrossRef]
- Wu, C.; Peng, D.; Soudani, K.; Siebicke, L.; Gough, C.M.; Arain, M.A.; Bohrer, G.; Lafleur, P.M.; Peichl, M.; Gonsamo, A.; et al. Land surface phenology derived from normalized difference vegetation index (NDVI) at global FLUXNET sites. Agric. For. Meteorol. 2017, 233, 171–182. [Google Scholar] [CrossRef]
- Luo, X.; Chen, X.; Xu, L.; Myneni, R.; Zhu, Z. Assessing Performance of NDVI and NDVI3g in Monitoring LeafUnfolding Dates of the Deciduous Broadleaf Forest in Northern China. Remote. Sens. 2013, 5, 845–861. [Google Scholar] [CrossRef]
- Rees, W.G.; Golubeva, E.I.; Williams, M. Are vegetation indices useful in the Arctic? Pol. Rec. 2009, 34, 333. [Google Scholar] [CrossRef]
- Luus, K.A.; Commane, R.; Parazoo, N.C.; Benmergui, J.; Euskirchen, E.S.; Frankenberg, C.; Joiner, J.; Lindaas, J.; Miller, C.E.; Oechel, W.C.; et al. Tundra photosynthesis captured by satellite-observed solar-induced chlorophyll fluorescence. Geophys. Res. Lett. 2017, 44, 1564–1573. [Google Scholar] [CrossRef]
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky–Golay filter. Remote. Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Elmore, A.J.; Guinn, S.M.; Minsley, B.J.; Richardson, A.D. Landscape controls on the timing of spring, autumn, and growing season length in mid-Atlantic forests. Glob. Chang. Boil. 2012, 18, 656–674. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M.; D’Odorico, P. Deriving land surface phenology indicators from CO2 eddy covariance measurements. Ecol. Indic. 2013, 29, 203–207. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using MODIS NDVI. Remote. Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Walker, M.A.; Fja, D.; Evander, M. Circumpolar arctic vegetation: Introduction and perspectives. J. Veg. Sci. 2010, 5, 757–764. [Google Scholar] [CrossRef]
- Høye, T.T.; Post, E.; Schmidt, N.M.; Trøjelsgaard, K.; Forchhammer, M.C. Shorter flowering seasons and declining abundance of flower visitors in a warmer Arctic. Nat. Clim. Chang. 2013, 3, 759. [Google Scholar] [CrossRef]
- Schimel, D.; Pavlick, R.; Fisher, J.B.; Asner, G.P.; Saatchi, S.; Townsend, P.; Miller, C.; Frankenberg, C.; Hibbard, K.; Cox, P. Observing terrestrial ecosystems and the carbon cycle from space. Glob. Chang. Boil. 2015, 21, 1762–1776. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.A.; Raynolds, M.K.; Einarsson, E.; Elvebakk, A.; Gould, W.A.; Talbot, S.S.; Yurtsev, B.A. The Circumpolar Arctic Vegetation Map. J. Veg. Sci. 2010, 16, 267–282. [Google Scholar] [CrossRef]
- Lund, M.; Falk, J.M.; Friborg, T.; Mbufong, H.N.; Sigsgaard, C.; Soegaard, H.; Tamstorf, M.P. Trends in CO2 exchange in a high Arctic tundra heath, 2000–2010. J. Geophys. Res. Biogeosci. 2015, 117, 136. [Google Scholar]
- Kutzbach, L.; Wille, C.; Pfeiffer, E.M. Heat, water and carbon exchange between arctic tundra and the atmospheric boundary layer - the eddy covariance method. Available online: http://epic.awi.de/11002/ (accessed on 20 October 2018).
- Van der Molen, M.K.; Van Huissteden, J.; Parmentier, F.J.W.; Petrescu, A.M.R.; Dolman, A.J.; Maximov, T.C.; Kononov, A.V.; Karsanaev, S.V.; Suzdalov, D.A. The growing season greenhouse gas balance of a continental tundra site in the Indigirka lowlands, NE Siberia. Biogeosciences 2007, 4, 985–1003. [Google Scholar] [CrossRef]
- Laurila, T.; Thum, T.; Aurela, M.; Lohila, A. Carbon Dioxide Fluxes between the Scots Pine Forest and the Atmosphere, Meteorology and Biomass Data during SIFLEX-2002; Final Report of SIFLEX-2002 Project; European Space Agency: Paris, France, 2003. [Google Scholar]
- Westergaard-Nielsen, A.; Lund, M.; Hansen, B.U.; Tamstorf, M.P. Camera derived vegetation greenness index as proxy for gross primary production in a low Arctic wetland area. ISPRS J. Photogramm. Remote. Sens. 2013, 86, 89–99. [Google Scholar] [CrossRef]
- Suni, T.; Rinne, J.; Reissell, A.; Altimir, N.; Keronen, P.; Rannik, Ü.; Dal Maso, M.; Kulmala, M.; Vesala, T. Long-term measurements of surface fluxes above a Scots pine forest in Hyytiälä, southern Finland, 1996–2001. Boreal Environ. Res. 2003, 8, 287–301. [Google Scholar]
- Kolari, P.; Kuimaia, L.; Pumpanen, J.; Launiainen, S.; Ilvesniemi, H.; Han, P.; Nikinmaa, E. CO2 exchange and component CO2 fluxes of a boreal Scots pine forest. Boreal Environ. Res. 2009, 14, 761–783. [Google Scholar]
- Aurela, M.; Laurila, T.; Tuovinen, J.P. Annual CO2 balance of a subarctic fen in northern Europe: Importance of the wintertime efflux. J. Geophys. Res. Atmos. 2002, 107, ACH–17. [Google Scholar] [CrossRef]
- Peichl, M.; Öquist, M.; Ottosson Löfvenius, M.; Ilstedt, U.; Sagerfors, J.; Grelle, A.; Lindroth, A.; Nilsson, M.B. A 12-year record reveals pre-growing season temperature and water table level threshold effects on the net carbon dioxide exchange in a boreal fen. Environ. Res. Lett. 2014, 9, 055006. [Google Scholar] [CrossRef]
- Nilsson, M.; Sagerfors, J.; Buffam, I.; Laudon, H.; Eriksson, T.; Grelle, A.; Klemedtsson, L.; Weslien, P.E.R.; Lindroth, A. Contemporary carbon accumulation in a boreal oligotrophic minerogenic mire—A significant sink after accounting for all C-fluxes. Glob. Chang. Boil. 2008, 14, 2317–2332. [Google Scholar] [CrossRef]
- Lindroth, A.; Lagergren, F.; Aurela, M.; Bjarnadottir, B.; Christensen, T.; Dellwik, E.; Grelle, A.; Ibrom, A.; Johansson, T.; Lankreijer, H.; et al. Leaf area index is the principal scaling parameter for both gross photosynthesis and ecosystem respiration of Northern deciduous and coniferous forests. Tellus B 2008, 60, 129–142. [Google Scholar] [CrossRef]
- Chen, B.; Coops, N.C.; Fu, D.; Margolis, H.A.; Amiro, B.D.; Black, T.A.; Arain, M.A.; Barr, A.G.; Bourque, C.P.A.; Flanagan, L.B.; et al. Characterizing spatial representativeness of flux tower eddy-covariance measurements across the Canadian Carbon Program Network using remote sensing and footprint analysis. Remote. Sens. Environ. 2012, 124, 742–755. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote. Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Delbart, N.; Kergoat, L.; Le Toan, T.; Lhermitte, J.; Picard, G. Determination of phenological dates in boreal regions using normalized difference water index. Remote. Sens. Environ. 2005, 97, 26–38. [Google Scholar] [CrossRef]
- Delbart, N.; Toan, T.L.; Kergoat, L.; Fedotova, V. Remote sensing of spring phenology in boreal regions: A free of snow-effect method using NOAA-AVHRR and SPOT-VGT data (1982–2004). Remote. Sens. Environ. 2008, 101, 52–62. [Google Scholar] [CrossRef]
- Jin, H.; Eklundh, L. A physically based vegetation index for improved monitoring of plant phenology. Remote. Sens. Environ. 2014, 152, 512–525. [Google Scholar] [CrossRef]
- Salomonson, V.V.; Appel, I. Estimating fractional snow cover from MODIS using the normalized difference snow index. Remote. Sens. Environ. 2004, 89, 351–360. [Google Scholar] [CrossRef]
- White, M.A.; De Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Chang. Boil. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Sekhon, N.S.; Hassan, Q.K.; Sleep, R.W. Evaluating potential of MODIS-based indices in determining “Snow Gone” stage over forest-dominant regions. Remote. Sens. 2010, 2, 1348–1363. [Google Scholar] [CrossRef]
- Karkauskaite, P.; Tagesson, T.; Fensholt, R. Evaluation of the Plant Phenology Index (PPI), NDVI and EVI for Start-of-Season Trend Analysis of the Northern Hemisphere Boreal Zone. Remote. Sens. 2017, 9, 485. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; Vanderzee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Gamon, J.A.; Huemmrich, K.F.; Wong, C.Y.; Ensminger, I.; Garrity, S.; Hollinger, D.Y.; Noormets, A.; Penuelas, J. A remotely sensed pigment index reveals photosynthetic phenology in evergreen conifers. Proc. Natl. Acad. Sci. USA 2016, 113, 13087–13092. [Google Scholar] [CrossRef] [PubMed]
- Dye, D.G.; Tucker, C.J. Seasonality and trends of snow-cover, vegetation index, and temperature in northern Eurasia. Geophys. Res. Lett. 2003, 30. [Google Scholar] [CrossRef]
- Shabanov, N.V.; Zhou, L.; Knyazikhin, Y.; Myneni, R.B. Analysis of interannual changes in northern vegetation activity observed in AVHRR data from 1981 to 1994. IEEE Trans. Geosci. Remote. Sens. 2002, 40, 115–130. [Google Scholar] [CrossRef]
- Beurs, K.M.D.; Wright, C.K.; Henebry, G.M. Dual scale trend analysis for evaluating climatic and anthropogenic effects on the vegetated land surface in Russia and Kazakhstan. Environ. Res. Lett. 2009, 4, 940–941. [Google Scholar] [CrossRef]
- Hird, J.N.; McDermid, G.J. Noise reduction of NDVI time series: An empirical comparison of selected techniques. Remote. Sens. Environ. 2009, 113, 248–258. [Google Scholar] [CrossRef]
- Eastman, R.; Warren, S.G. Arctic cloud changes from surface and satellite observations. J. Clim. 2009, 23, 4233–4242. [Google Scholar] [CrossRef]
- Fletcher, B.J.; Gornall, J.L.; Poyatos, R.; Press, M.C.; Stoy, P.C.; Huntley, B.; Baxter, R.; Phoenix, G.K. Photosynthesis and productivity in heterogeneous arctic tundra: Consequences for ecosystem function of mixing vegetation types at stand edges. J. Ecol. 2012, 100, 441–451. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhu, W. Uncertainty of Remote Sensing Data in Monitoring Vegetation Phenology: A Comparison of MODIS C5 and C6 Vegetation Index Products on the Tibetan Plateau. Remote. Sens. 2017, 9, 1288. [Google Scholar] [CrossRef]
- Joiner, J.; Yoshida, Y.; Vasilkov, A.P.; Yoshida, Y.; Corp, L.A.; Middleton, E.M. First observations of global and seasonal terrestrial chlorophyll fluorescence from space. Biogeosci. Discuss. 2011, 8, 637–651. [Google Scholar] [CrossRef]
- Yang, X.; Tang, J.; Mustard, J.F.; Lee, J.E.; Rossini, M.; Joiner, J.; Munger, J.W.; Kornfeld, A.; Richardson, A.D. Solar-induced chlorophyll fluorescence that correlates with canopy photosynthesis on diurnal and seasonal scales in a temperate deciduous forest. Geophys. Res. Lett. 2015, 42, 2977–2987. [Google Scholar] [CrossRef]
- Damm, A.; Guanter, L.; Paul-Limoges, E.; Van der Tol, C.; Hueni, A.; Buchmann, N.; Eugster, W.; Ammann, C.; Schaepman, M.E. Far-red sun-induced chlorophyll fluorescence shows ecosystem-specific relationships to gross primary production: An assessment based on observational and modeling approaches. Remote. Sens. Environ. 2015, 166, 91–105. [Google Scholar] [CrossRef]
- Lu, X.; Cheng, X.; Li, X.; Tang, J. Opportunities and challenges of applications of satellite-derived sun-induced fluorescence at relatively high spatial resolution. Sci. Total. Environ. 2018, 619–620, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xiao, J.; He, B.; Altaf Arain, M.; Beringer, J.; Desai, A.R.; Emmel, C.; Hollinger, D.Y.; Krasnova, A.; Mammarella, I.; et al. Solar-induced chlorophyll fluorescence is strongly correlated with terrestrial photosynthesis for a wide variety of biomes: First global analysis based on OCO-2 and flux tower observations. Glob. Chang. Boil. 2018. [Google Scholar] [CrossRef] [PubMed]
- Collingwood, A.; Charbonneau, F.; Shang, C.; Treitz, P. Spatiotemporal Variability of Arctic Soil Moisture Detected from High-Resolution RADARSAT-2 SAR Data. Adv. Meteorol. 2018, 2018, 1–17. [Google Scholar] [CrossRef]








| Site | Site name | Latitude | Longitude | IGBP | GPP duration | Reference |
|---|---|---|---|---|---|---|
| DK-ZaH | Zackenberg Heath | 74.47 | −20.55 | GRA | 2000–2014 | [35] |
| RU-Sam | Samoylov | 72.37 | 126.50 | GRA | 2002–2014 | [36] |
| US-Atq | Atqasuk | 70.47 | −157.41 | WET | 2004–2008 | [35] |
| RU-Cok | Chokurdakh | 70.83 | 147.50 | OSH | 2003–2014 | [37] |
| FI-Sod | Sodankyla | 67.36 | 26.64 | ENF | 2001 -–2014 | [38] |
| DK-NuF | Nuuk Fen | 64.13 | −51.39 | WET | 2008–2014 | [39] |
| FI-Hyy | Hyytiala | 61.85 | 24.30 | ENF | 2000–2014 | [40,41] |
| FI-Kaa | Kaamanen | 69.14 | 27.30 | WET | 2000–2008 | [42] |
| SE-Deg | Degerö | 64.18 | 19.56 | GRA | 2001–2006 | [43,44] |
| SE-Nor | Norunda | 60.09 | 17.48 | ENF | 2003–2007 | [45] |
| SE-Sk1 | Skyttorp 1 | 60.13 | 17.92 | ENF | 2005–2008 | [45] |
| Collection 6 | Collection 5 | ||||
|---|---|---|---|---|---|
| Metrics | Indices | R2 | RMSE | R2 | RMSE |
| SOS | NDVI | 0.001 | 10.73 | 0.38 | 7.68 |
| EVI | 0.39 | 9.392 | 0.09 | 12.54 | |
| PI | 0.08 | 9.705 | 0.14 | 8.08 | |
| PPI | 0.14 | 8.175 | 0.2 | 8.14 | |
| EOS | NDVI | 0.02 | 18.37 | 0.23 | 19.34 |
| EVI | 0.07 | 19.07 | 0.1 | 17.78 | |
| PI | 0.08 | 17.91 | 0.01 | 15.87 | |
| PPI | 0.03 | 16.11 | 0.14 | 14.35 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Lu, X.; Cheng, X.; Li, X.; Peichl, M.; Mammarella, I. Limitations and Challenges of MODIS-Derived Phenological Metrics Across Different Landscapes in Pan-Arctic Regions. Remote Sens. 2018, 10, 1784. https://doi.org/10.3390/rs10111784
Wang S, Lu X, Cheng X, Li X, Peichl M, Mammarella I. Limitations and Challenges of MODIS-Derived Phenological Metrics Across Different Landscapes in Pan-Arctic Regions. Remote Sensing. 2018; 10(11):1784. https://doi.org/10.3390/rs10111784
Chicago/Turabian StyleWang, Siyu, Xinchen Lu, Xiao Cheng, Xianglan Li, Matthias Peichl, and Ivan Mammarella. 2018. "Limitations and Challenges of MODIS-Derived Phenological Metrics Across Different Landscapes in Pan-Arctic Regions" Remote Sensing 10, no. 11: 1784. https://doi.org/10.3390/rs10111784
APA StyleWang, S., Lu, X., Cheng, X., Li, X., Peichl, M., & Mammarella, I. (2018). Limitations and Challenges of MODIS-Derived Phenological Metrics Across Different Landscapes in Pan-Arctic Regions. Remote Sensing, 10(11), 1784. https://doi.org/10.3390/rs10111784