1. Introduction
Forests provide valuable ecosystem goods and services for human well-being [
1]. Therefore, it is of great interest to gather information about the state of the forest ecosystems. Because forest inventories are costly, it is essential to develop cost-effective mapping methods to allow management of forests [
2]. In times of climate change, periodic assessments are in demand, as changes in phenology [
3,
4] as well as shifts in species composition [
5,
6] become apparent. It is important to discover efficient ways to continually monitor forested areas and subsequently gain knowledge about processes and changes.
Remote sensing offers valuable tools to implement mapping and monitoring approaches [
7]. Until now, mainly optical sensors have been used to monitor forests on a global scale, ranging from intra-annual studies [
8,
9] to others examining several years [
4,
10]. Using optical sensors however, two main disadvantages are apparent. First, optical sensors are not sensitive to vegetation structure. Second, temporally consistent data acquisition is challenging, as the optical sensor’s views are often impaired by cloud cover. To overcome these disadvantages, it is important to consider synthetic aperture radar (SAR). SAR is sensitive to vegetation structure [
11] and is acquired independent of daylight and weather, so it can more easily produce temporally consistent data [
12].
Until now, SAR data have often been used to estimate biomass [
13,
14] and map the extent of deforestation [
15] in tropical forests. For boreal forests, several methods working with SAR data have been applied, e.g., for mapping forest types [
16,
17] or estimating growing stock volume [
18]. Because of frequent cloud coverage, SAR has to be considered in the tropics. In the northern boreal forests, however, SAR is also relevant, as long periods with sparse or no daylight pose a further barrier to using optical remote sensing data. It is useful to assess the potential of SAR data for mixed temperate forests, as only a few studies assessing this potential have been conducted until now [
19,
20]. Given that significant structural changes due to phenology happen within a short time period in spring and autumn, reliable observations are critical.
The two recently launched Sentinel-1 (S-1) satellites, carrying C-band SAR sensors, offer new possibilities in the compilation of time series. Generally, they acquire backscatter in vertical-vertical (VV) and vertical-horizontal (VH) polarisation mode over landmasses [
21]. S-1 data have already been successfully applied to several land applications. Both VV and VH polarisations have been used to monitor crop growth of several cultures in Angola [
22] and of rice in Myanmar [
23]. Balzter et al. [
24] and Abdikan et al. [
25] showed that using VV and VH polarisation data leads to satisfying classification of land cover classes, applying random forest (RF) and support vector machine (SVM) classifiers, respectively. Other studies exhibited the potential of S-1 data for burnt area mapping in forests [
26], monitoring wetlands [
27] or snow wetness estimation in the Alps [
28,
29]. S-1 offers superior spatial and temporal coverage and much improved radiometric calibration stability in comparison to former spaceborne C-band sensors [
30].
Previous work has shown that SAR forest backscatter is very complex, being dependent on several sensor and object properties. Generally, sensors with high frequencies such as X- or C-band are less able to penetrate the crown than are lower frequencies such as S- or L-band [
31]. Thus, the dominance of the crown return is stronger at high frequencies. The polarisation used by the SAR sensor also impacts the backscatter. Cross-polarised backscatter from forested areas leads to higher correlation with biomass than co-polarised backscatter [
32]. Object properties impacting forest backscatter can be divided into two groups. The first group consists of properties that influence the dielectric constant of the trees, such as the moisture content of branches and leaves [
19], external moisture conditions [
33], and the temperature of the wood [
34,
35]. The second group consists of structural properties such as the size and orientation of branches and leaves [
36], the spatial pattern of trees [
37], and whether or not foliage is present [
38,
39]. The last is especially interesting, as it could allow the monitoring of phenology at a land surface scale.
Until now, most studies with the aim of mixed temperate forest characterisation have focused on extracting additional information by using fully polarimetric SAR data to discriminate different backscatter behaviours of forest types [
40,
41] or estimate forest density using Wishart supervised and SVM classifiers [
42]. Collecting multiple SAR acquisitions into longer time series has the potential to reveal temporal signatures, as shown by Sharma et al. [
33] with C-band horizontal-horizontal (HH) polarisation data for forest and Gamba et al. [
43] with C-band VV polarisation data for several land cover types. However, only few studies have analysed longer C-band time series of forested areas. Ahern et al. [
39] and Proisy et al. [
19] found phenology effects on C-band SAR backscatter for coniferous and deciduous forest types using HH and VV polarisations, respectively. A recent study using S-1 data showed the impact of phenology of deciduous forests to be much higher in cross-polarised than in co-polarised data [
20].
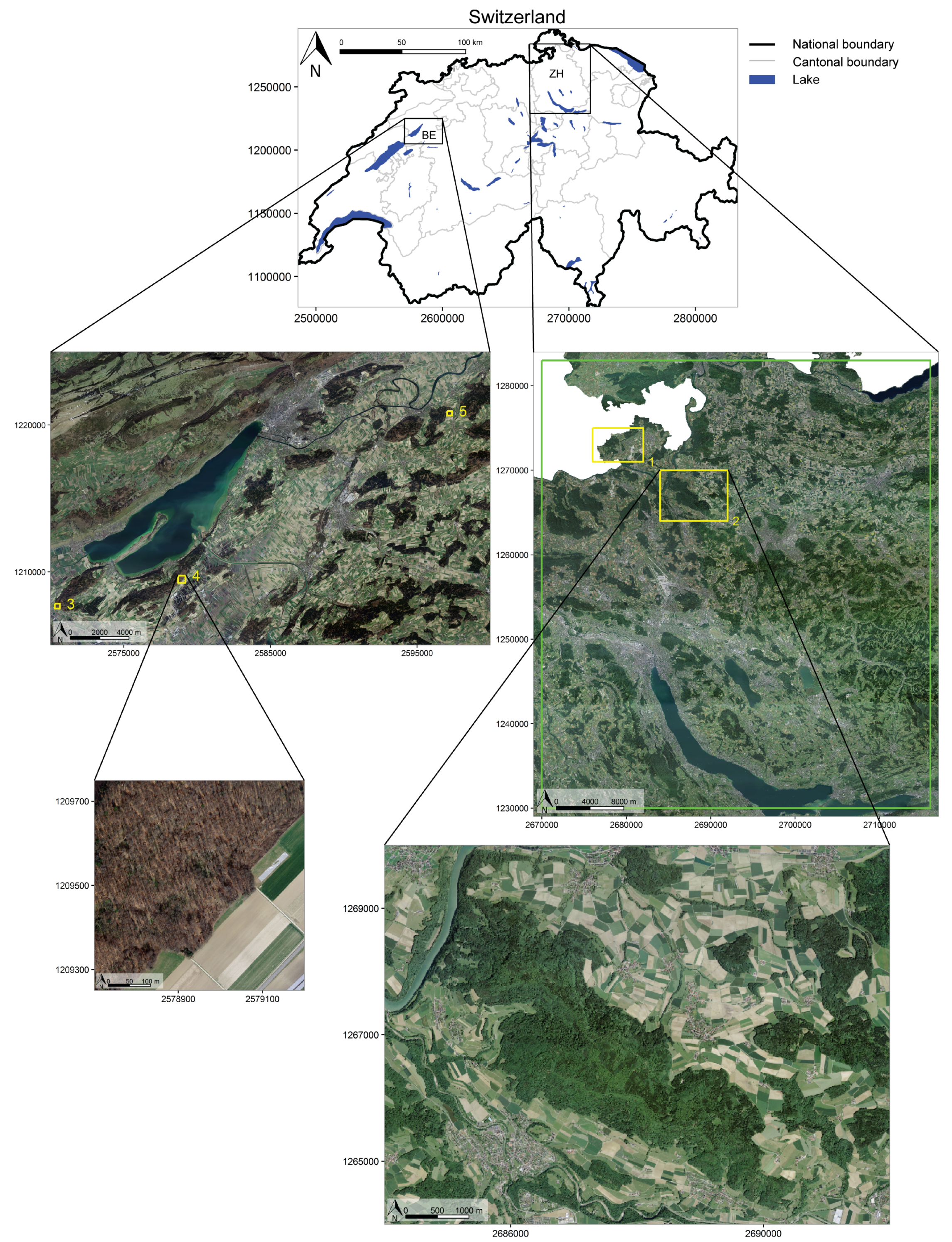
In this paper, we present an analysis of Sentinel-1 SAR radiometrically calibrated backscatter time series of an unprecedented set of VV and VH polarisation observations over mixed temperate forests in northern Switzerland. One coniferous and two broadleaved, deciduous species were investigated. We show how to build the C-band time series and to subsequently extract descriptors for the different forests investigated. A detailed analysis of these descriptors is provided, focusing on differences between tree species and between the investigated years. Finally, we show how our findings help to classify mixed temperate forests and how they could help in future monitoring of the seasonal vegetation cycles of forests.
4. Discussion
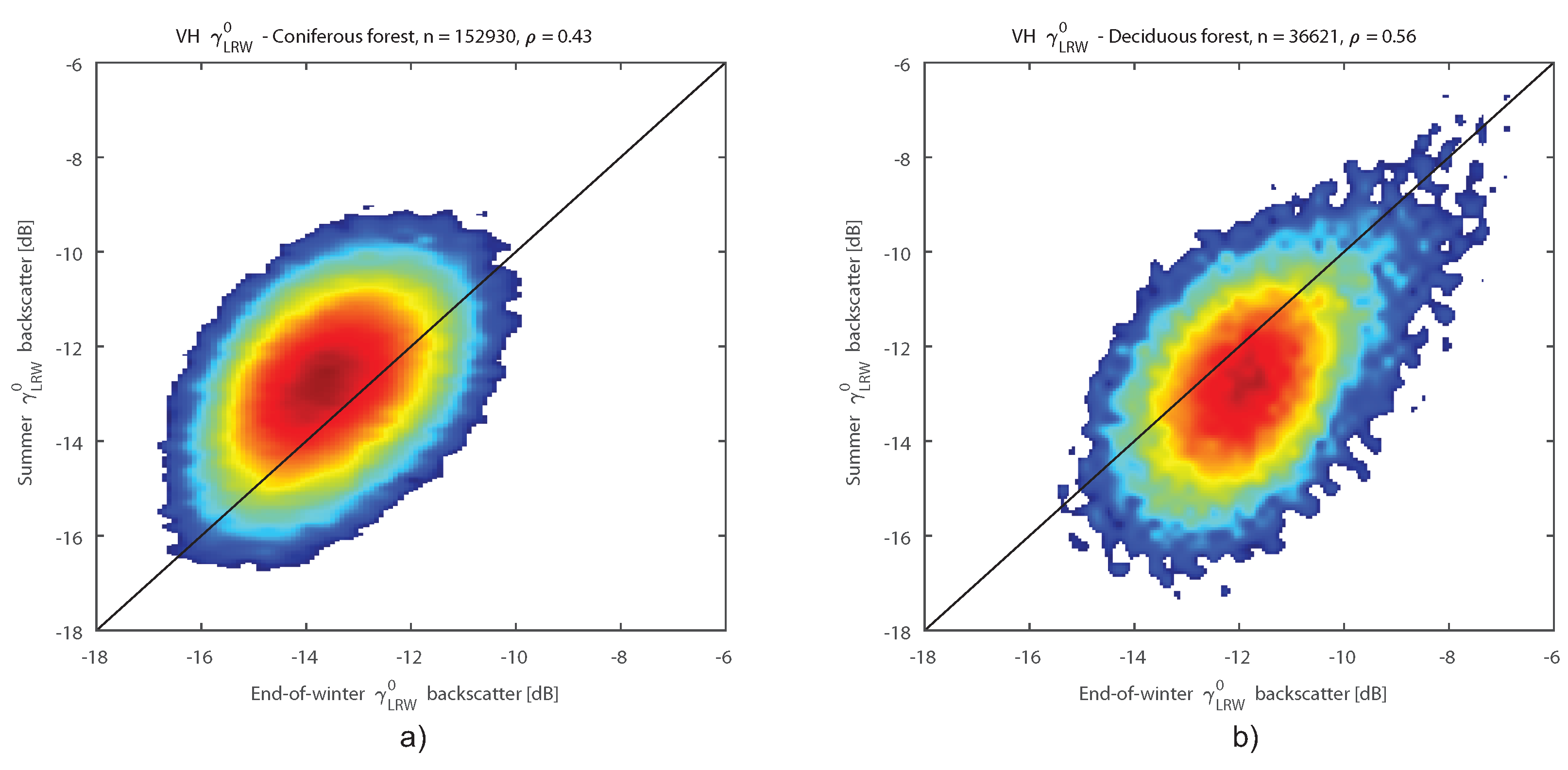
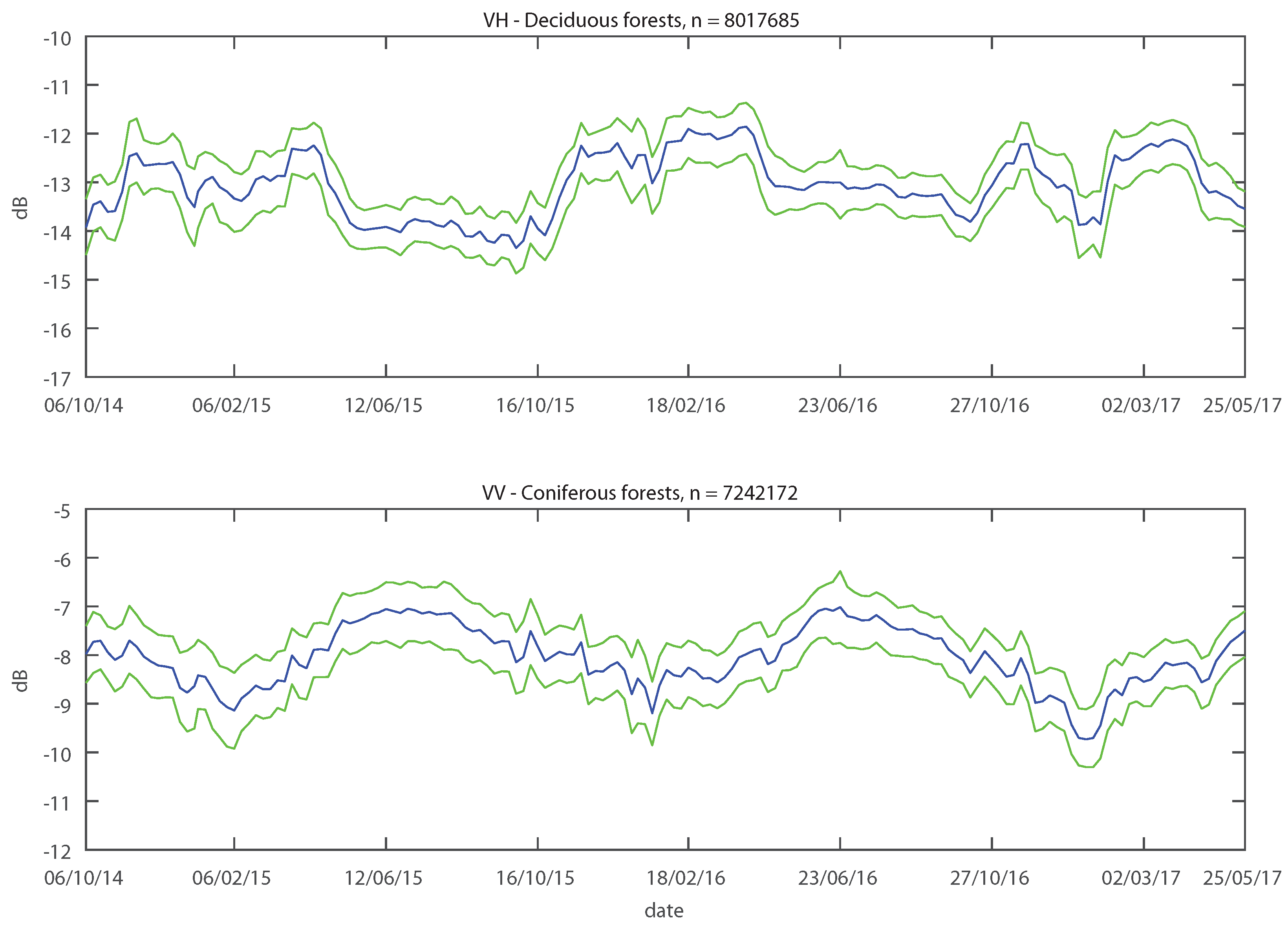
Analysis of the SAR C-band backscatter time series of forests showed that the observed backscatter behaviour was robust for multiple sites over multiple years. Generally, annual median backscatter of forests consisting of deciduous species was higher than backscatter of forests consisting of spruce. Both types of forests showed distinctive seasonal backscatter patterns. The deciduous species had higher backscatter in winter and lower in summer, while spruce exhibited the opposite behaviour.
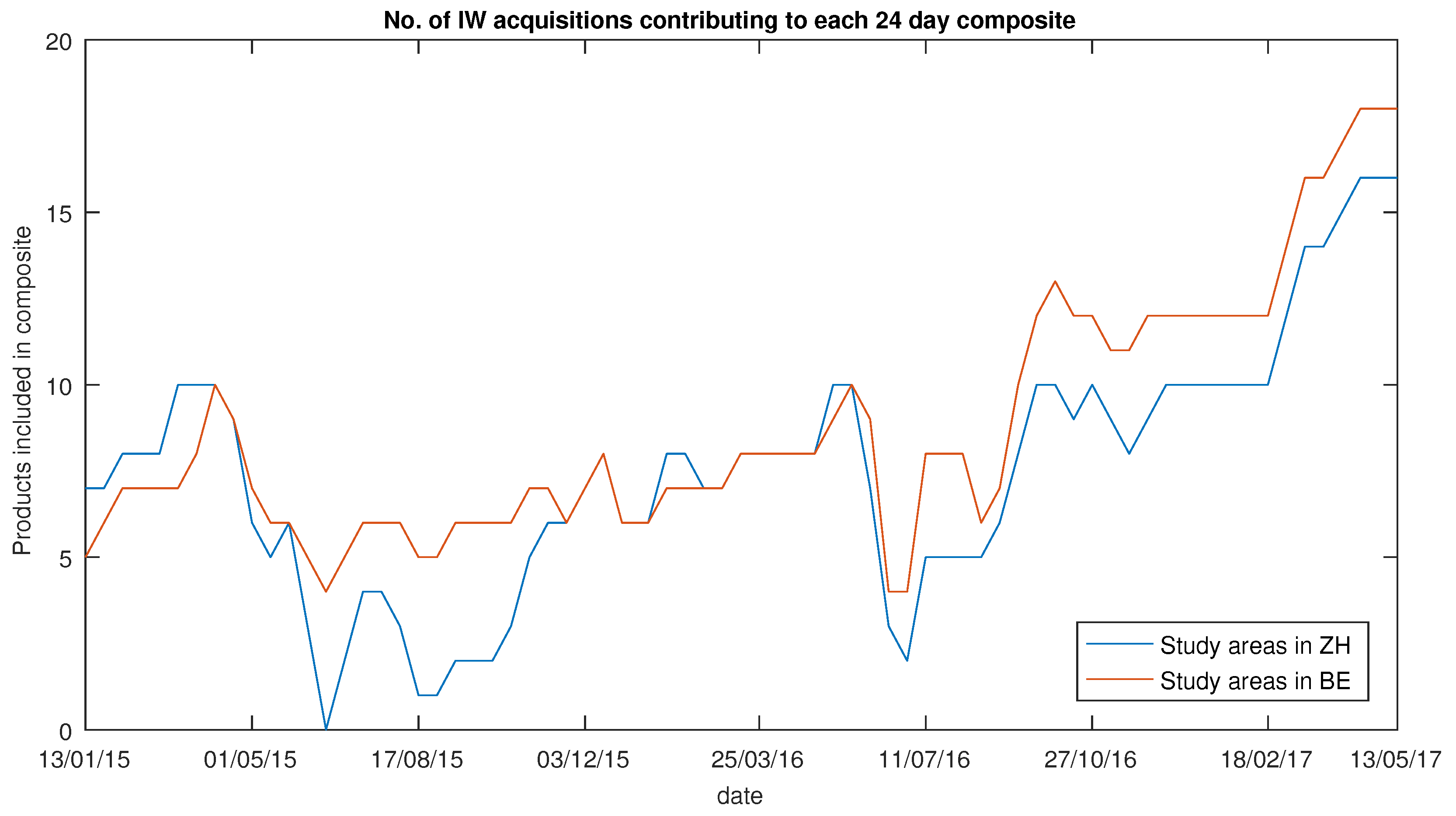
Temporal variation in SAR data is caused by signal noise, speckle and short- and long-term environmental changes. The former three were strongly mitigated because of the methodology used in the SAR data processing. The LRW approach combines the backscatter from multiple tracks (see
Figure 5) into one composite. Hence, the approach suppresses the influences of signal noise, speckle and short-term variations, such as rainfall events before single acquisitions. As a result, the annual variation can mainly be attributed to long-term environmental changes occurring within the study areas. Moreover, the decision to use terrain-flattened
images could contribute to better results as well. All study areas (bar Büren a.d.A.) investigated in this study are in hilly terrain.
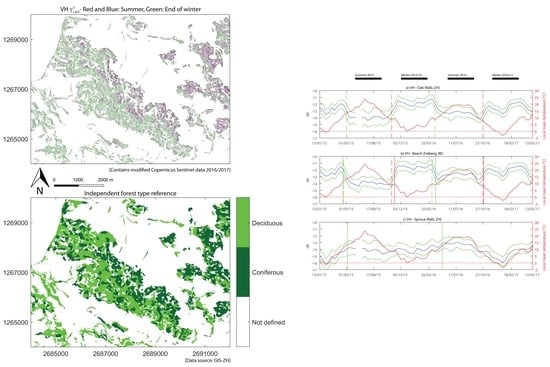
4.1. Seasonal Signature of Deciduous Species
Both deciduous species showed seasonal changes in VH backscatter behaviour. Backscatter values dropped in the spring from a high winter level to a low summer level, rising again in the autumn, resulting in positive
values. The predominant long-term change in deciduous forests is the seasonal vegetation cycle, manifested in the annual growth of trees and presence of foliage. The presence of foliage occurred at the time with the lower backscatter values observed in the summer months: a dependence of the backscatter on leaf presence was observed [
19,
20].
Potential causes for the changing cross-pol scattering behaviour remain uncertain. One reason might be the changing size of the scatterers present in the tree crown. In summer, leaves substitute branches as main scatterers. C-band is primarily sensitive to small branches and only secondarily to leaves [
71]. Leaves have been reported to act not only as scatterers but also as attenuators for C-band [
32,
72]. Hence, leaf emergence in spring leads to a combination of effects, resulting in lower backscatter during the leaf-on period. (a) A scatterer substitution, where the sensitivity of C-band is reduced and (b) an attenuation of the microwave energy by the leaves. Leaf fall in autumn otherwise exposes the small branches again, leading to higher backscatter.
Another possible reason for higher backscatter values during the leaf-off period is the influence of ground scattering [
73]. With lower attenuation in winter than in summer, ground scattering contributes more strongly to the measured backscatter. Proisy et al. [
19] showed that in winter, the contribution of soil and branches were comparable, whereas in summer, the contribution of the soil dropped because of the attenuation by the leaves.
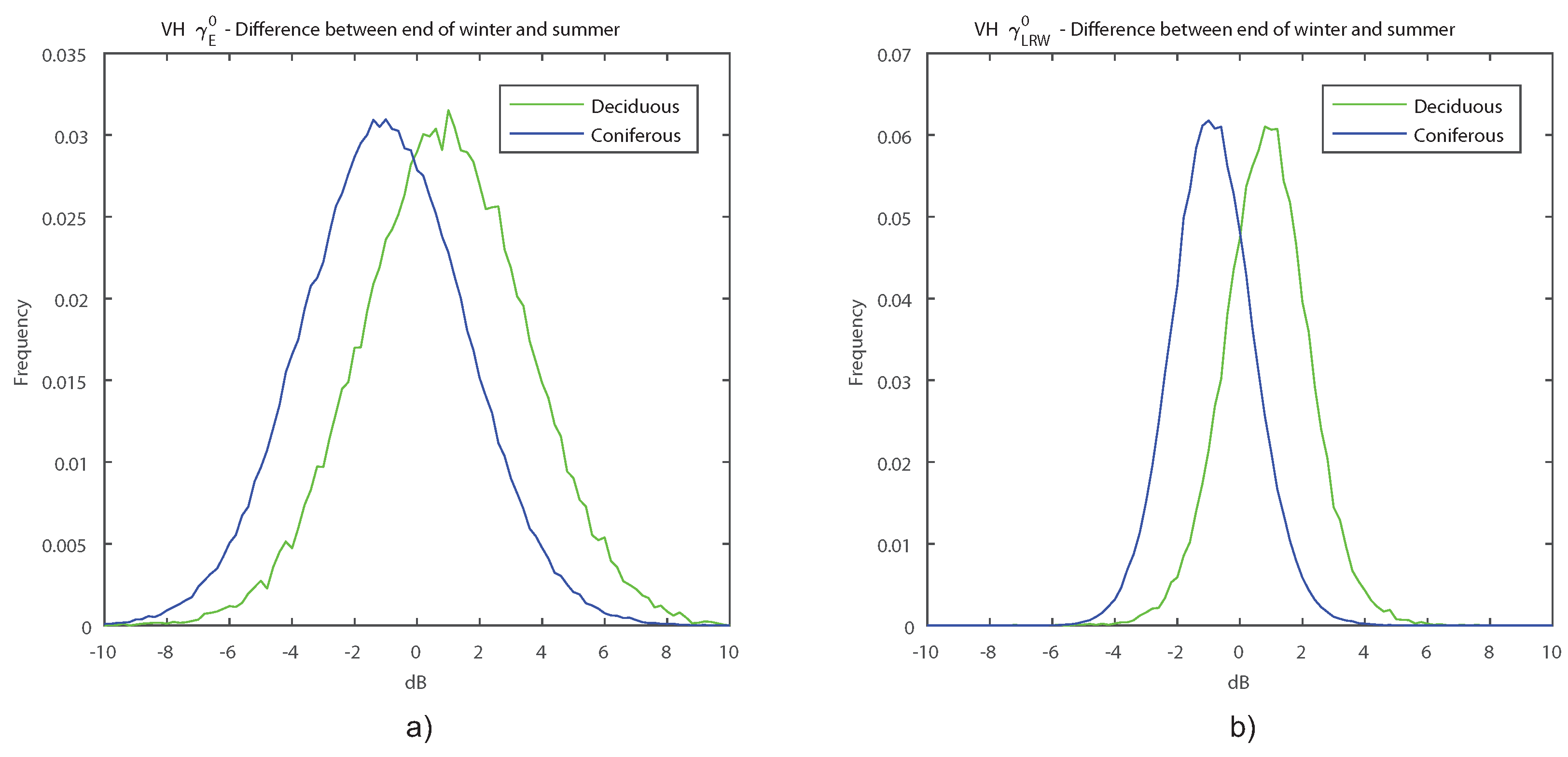
Surprisingly, VV backscatter did not show the same seasonal behaviour as VH. Although slightly higher backscatter was observed in winter, the
s show that there was a less defined seasonal signature. An interesting feature was the constant decrease in backscatter during the leaf-on season. This could be attributed to the slow drying of leaves in summer [
74].
Leckie and Ranson [
32] did not mention a difference in temporal signature between VH and VV backscatter in their comprehensive review of SAR backscatter of forest. Potential reasons for our observed differences between the two polarisations are discussed in the following. Volume scattering becomes more important when the scatterers present are more random and complex [
75]. Volume scattering is also known to have a larger influence on VH than VV polarisation [
31,
32]. Leaf emergence probably leads to a more complex arrangement of the scatterers, leading to a stronger influence on the VH than the VV backscatter. Our empirical observation of the difference between leaf-off and leaf-on conditions in C-band backscatter of deciduous forests corresponded well with the simulation made by Chuah and Tan [
38]. The S-1 IW acquisitions processed in our study were measured at incident angles between 31° and 46°. Their modelled C-band backscatter values at 50° incident angle were different at both polarisations, but they also showed a positive difference at VH and almost no difference at VV between leaf-off and leaf-on conditions.
4.2. Break Date Extraction to Monitor Phenology
The break date extraction yielded promising predictions, bearing in mind that the time series had a temporal resolution of 24 days. Even finer details such as differences between species were observable. The observed tendency of the oak stands to have their leaves emerge later than the beech stands agrees with the common theory concerning the timing of leaf emergence of the two species [
76]. However, it is important to consider that the independent ground validation data were produced by observations from volunteers. Hence, the reported values have to be treated with caution.
We showed that with our approach, the phenological cycles of deciduous forests can be monitored on a landscape scale. With longer time series available in the future, it will be possible to analyse phenology trends with S-1 data. The same type of phenology trend analyses that have usually used MODIS Normalised Difference Vegetation Index (NDVI) such as White et al. [
77] in North America or Hamunyela et al. [
78] in Europe could be conducted. Using S-1 instead of MODIS data, an enhanced spatial and temporal resolution would be available.
The results also showed that there are still some issues using the breakpoints algorithm for the extraction of the break dates. The problems encountered for the two study areas can be explained by the weak dependency of the backscatter on foliage within each respective year. Other influences (e.g., exceptional freezing temperature) on the backscatter seemed to disturb the seasonal signature to such an extent that a smooth break date extraction was not always possible.
4.3. Seasonal Signature of Spruce
Backscatter from spruce stands did not show similarly abrupt changes. At both polarisations, their seasonal signatures were less pronounced and more gradual. Lower values were observed in winter and higher in summer, resulting in negative
values. Because of their perpetual foliage, no strong influence of changing foliage or ground scattering was expected [
39]. The backscatter of coniferous forests is mainly dependent on the number of needles and small branches [
11]. Because the leaf area index (LAI) of coniferous forests is usually highest in July [
74], the highest backscatter can therefore be expected in that month. Dostálová et al. [
20] argued that in addition to the larger number of needles, the development of the understory in summer might also influence backscatter in coniferous forests.
The observed lower annual VH median backscatter of spruce stands compared to beech and oak stands was mainly due to the higher backscatter observed in winter for the latter, caused by the factors mentioned in
Section 4.1. At VV, this effect was not observed to the same extent. Nevertheless, higher backscatter in winter relative to the spruce stands was apparent. This observation confirms previous studies that investigated differences in backscatter between deciduous and coniferous species [
19,
39,
79].
4.4. Differences between the Investigated Years
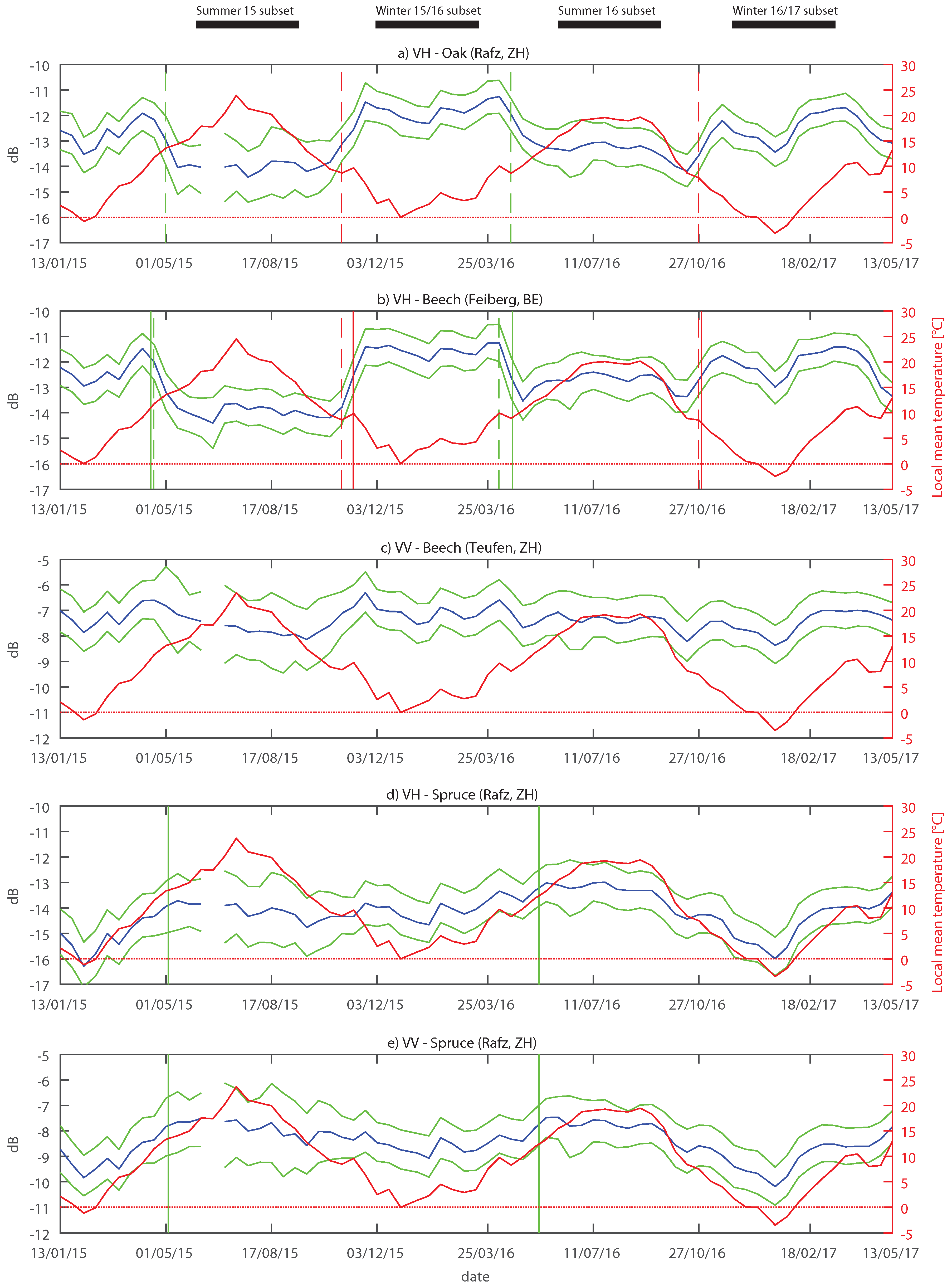
Clear differences were observed within the timespan investigated. Inter-annual differences were most probably due to different meteorological conditions. A clear influence of temperature on the backscatter can be seen in
Figure 9. In summer, higher VH backscatter was observed when temperatures were also higher. In winter, there was an even stronger influence of the temperature. When the temperature fell below the freezing point, the backscatter dropped significantly. This was mainly observed in January 2017, when temperatures in Switzerland were very low for a long period, but also in January 2015. Temperature is known to influence the dielectric properties of the scatterers [
35]. Especially when temperatures are below 0 °C, dielectric properties of the frozen scatterers change fundamentally, resulting in lower backscatter [
73].
Disparities were observed in the typical summer backscatter reductions of the deciduous forests at VH. Compared to 2016, the drop in 2015 was larger, which is well depicted in the plot of the study area Feiberg (
Figure 9b). This can probably be attributed to different weather conditions in the springs and summers of 2015 and 2016. Compared to 2016, temperatures, sunshine duration and the amount of precipitation in the study areas in spring 2015 were higher than normal [
80,
81]. These favourable growing conditions might have caused a better development of the tree crowns for the deciduous species [
82]. According to the points mentioned in
Section 4.1, a well developed crown with a high LAI would lead to a higher attenuation of the microwave energy. Thus, a larger drop would be expected and was observed.
Although precipitation can have an influence on backscatter [
33], no strong influence on short-term backscatter changes was detected in our data. As mentioned above, only a general influence on the growing season coupled with other meteorological factors was observed. This may be due to the applied compositing approach, which mitigates variations caused by short-term precipitation events.
Last, a possible influence of the backscatter calibration issue in summer 2015 cannot be disregarded. To correct the wrong backscatter values, a simple correction was executed by subtracting a fixed value in the affected images [
57,
58]. Residual errors may have been larger in these earlier data, contributing to the differences between summer 2015 and summer 2016. However, the observed disparities were not the same for the different study areas and caused no “step” in the VV data, so the likelihood of radiometric miscalibration appears negligible.
4.5. Comparisons to Other Studies
The results of this study do not contradict previously published results but extend it with an unprecedented set of C-band cross-pol observations of mixed temperate forests. Similar backscatter values were retrieved in a simulation study of sensitivity of ERS-1 C-band VV backscatter over forest [
73]. The non linear dependence of backscatter on temperature below the freezing point had also been simulated in that study. Proisy et al. [
19] analysed ERS-1/2 C-band VV backscatter of a mixed temperate forest for the timespan between April 1994 and February 1997. They presented similar VV backscatter ranges from deciduous and coniferous stands. Their time series consisted of one image per month and had no VH measurements. Although they used a lower temporal resolution, the shape of their temporal backscatter plot of a beech stand throughout the year was similar to the one shown in this study. A recent study, analysing intra-annual S-1 data of an Austrian mixed temperate forest between February 2015 and 2016, yielded results that were in line with ours [
20]. The ranges of backscatter observed from deciduous and coniferous forests at VV and VH as well as the seasonal signatures shown by the two forest types at both polarisations were consistent.
4.6. Classification of Forest Types and Species
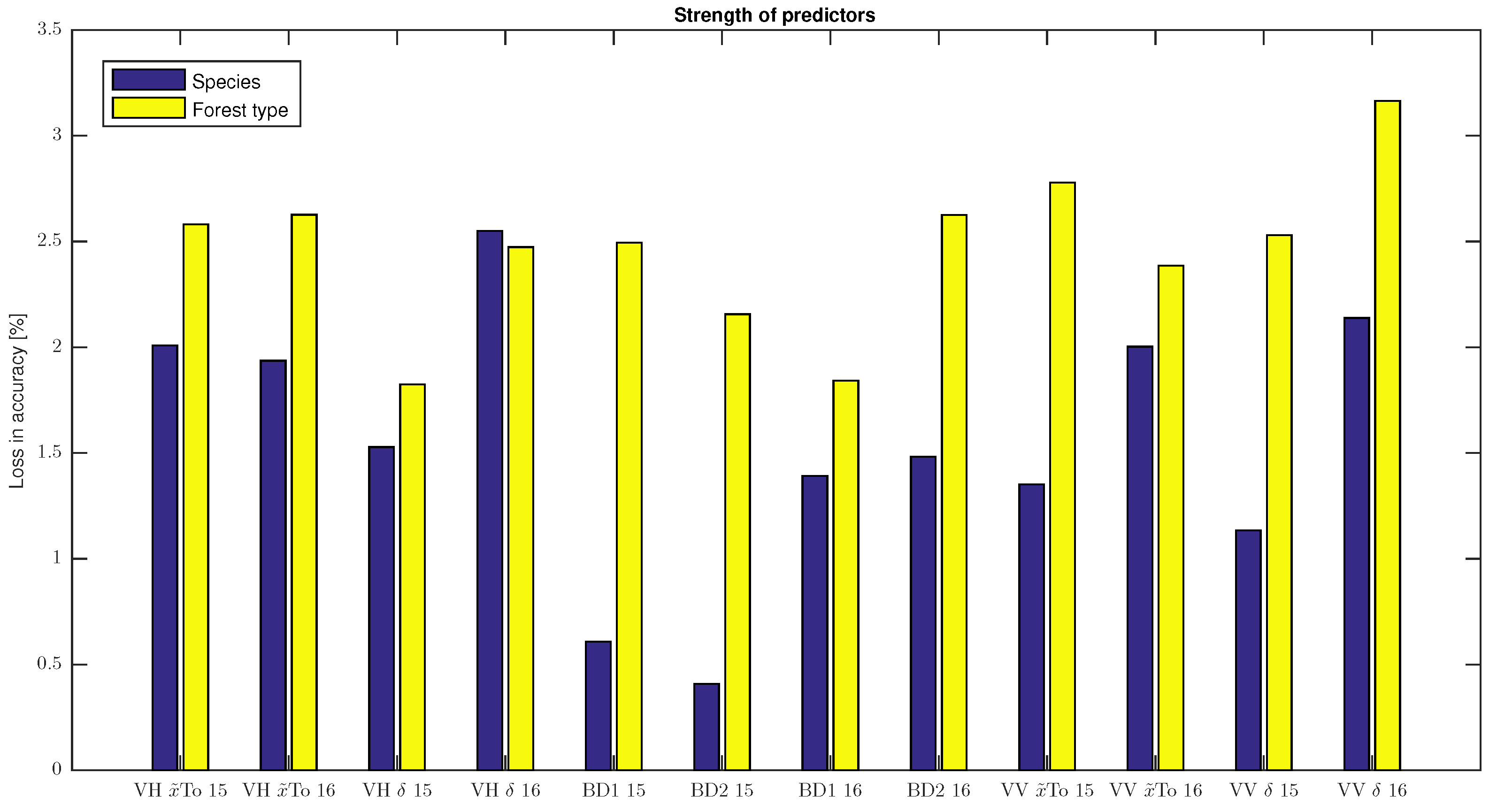
The classification results show that the observed backscatter behaviour allows segmentation into two different forest types or even into three different species. Not only the VH data but also VV contributed to achieving the good classification performances. This was surprising, as the seasonal backscatter behaviour of deciduous forests was less distinct at VV than at VH polarisation.
As expected, accuracies were lower when classifying species vs. forest type. Substantial confusion was seen between the beech and oak class, whereas in the forest type classification, they are both part of the deciduous class. The accuracies for spruce, however, were almost the same as for the coniferous class. This was not surprising as the coniferous class is mainly composed of spruce. The high accuracies were attained due to the backscatter behaviour of spruce being substantially different than that of the two deciduous species. Spruce was also the only coniferous species that could be examined. Future studies might include more species and try to use their seasonal backscatter patterns for classification approaches.
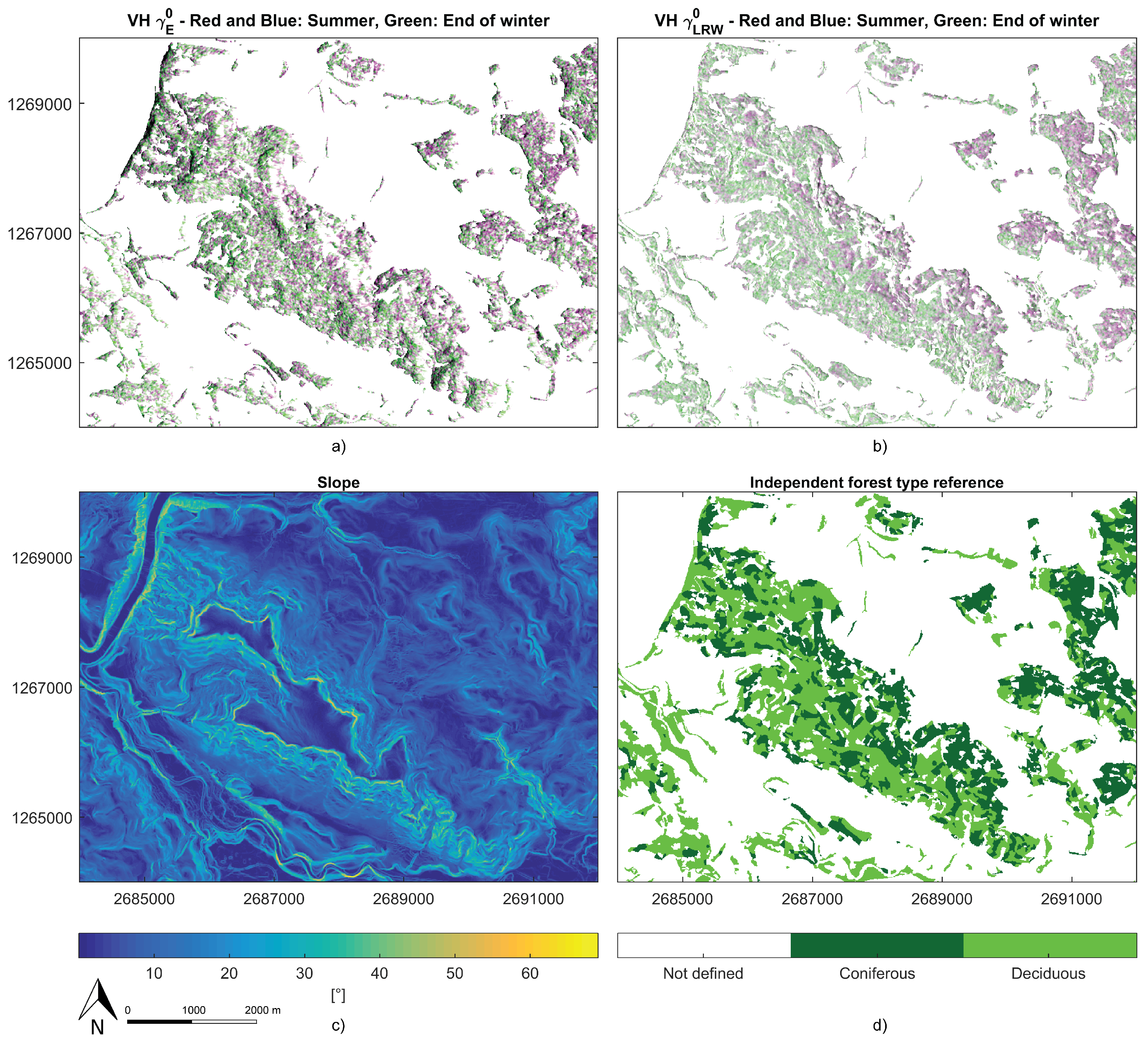
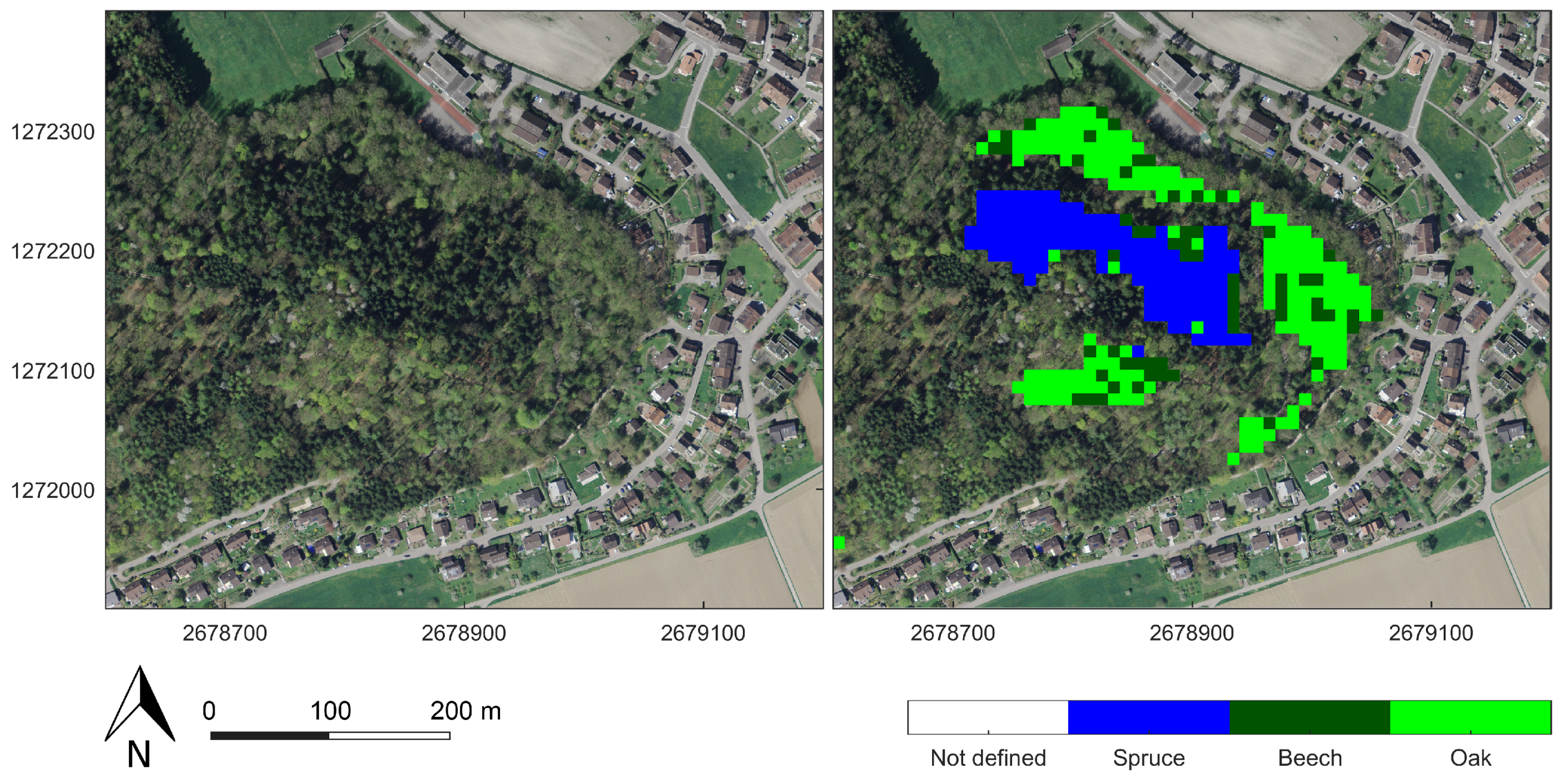
In this study, only stands with species homogeneity higher than 80% were considered for the classification. Hence, it remains unclear how the classifiers perform on more heterogeneous mixed forests. Future studies could test how the classifiers perform in mixed forests. However, when we compared the classification results to aerial images, we observed that the classifier was able to detect small patches of clustered spruce trees within a deciduous stand. Therefore, the classifiers might even outperform the aggregated forest stand-based independent ground reference. With a sample interval of 10 m, the classifiers operate at a high spatial resolution, whereas the information of the ground reference is based on larger homogenised areal extents. Therefore, many misclassifications may have been caused by the nature of the independent ground reference information. This might be verified in the future using a ground reference with a higher spatial resolution.
There was a significant time difference between the ground reference and the SAR data acquisition. The ground reference was produced in 2010, so a considerable time gap of at least 5 years was a further possible source of errors. Logging activities during that period likely degraded the classification performance. Clear-cut logged spruce areas tended to be classified as deciduous (see
Figure 12), most probably due to the high ground backscatter.
It is difficult to perform an in-depth comparison to other approaches using SAR but also different remote sensing technologies to classify mixed temperate forests. Study areas, the species investigated, and the number of training and validation samples differed substantially between the studies. Nevertheless, our achieved accuracies were comparable to other studies classifying deciduous and coniferous forests using even fully polarimetric C-band SAR data [
16,
83,
84]. Compared with other technologies such as airborne laser scanning (OA of 89–96% and
= 0.61 − 0.92) [
85,
86,
87] or imaging spectrometer data (OA of 83–99% and
= 0.73 − 0.98) [
88,
89,
90], our
forest type classification performance (OA of 86% and
= 0.73) was not as competitive. This could be due to the lower spatial resolution used in this instance.
4.7. Implications of the Findings
If one is aware of the mentioned different influences on C-band backscatter, C-band data can be used for several applications. Its potential for the classification of mixed temperate forests has been presented. Thus, a repetitive classification of an area allows change detection over time. For ecological applications such as seasonal vegetation cycle monitoring, the use of C-band data will become progressively more interesting, as longer time series of data become available. Nevertheless, the capability of the data to observe subtle differences between the years has been shown using only 2.5 years of data. Because the production of S-1 data is promised until at least 2030 [
91], longer time series of data should be available in the future. Using them, changes over time, such as the shift of phenophases due to climate change, could be detected and quantified.
Compared with state-of-the-art methods using optical data, SAR has valuable advantages. The sensitivity to structural elements adds an additional independent dimension of information about forests. This structural information helps to improve the understanding of natural processes in forests. Furthermore, the high temporal and spatial resolutions offered by Sentinel-1 allow a much denser sampling of data. Since the end of the commissioning phase of Sentinel-1B in September 2016 [
56], the number of products has been high enough to generate LRW composites with over twice the temporal resolution of those used in this study. An LRW composite every six days would offer even more insights into the temporal evolution of the backscatter of forests. To further enhance the temporal resolution, efforts should be made to combine S-1 and RADARSAT Constellation Mission data, due for launch in 2018 [
92].
With high temporal resolution available, there lies also great potential in analysing larger spatial extents. The backscatter behaviour of forests described in this study was also observed over the whole Alpine region (Lat/Lon: 43.5–49° N/5.5–17.5° E). Applying the same methodology as described in
Section 2.3 but with a temporal resolution of the LRW composites of 12 days, descriptors were calculated within the Coordination of Information on the Environment (CORINE) land cover classes for deciduous and coniferous forest [
93]. Plotted in the temporal domain, the same seasonal patterns are apparent at both polarisations and for both forest types, as shown in
Figure 13.
Future research could focus on more detailed analysis of the backscatter and the understanding of scattering processes. An in-depth study of the observed summer drop in VH backscatter might result in interesting new insights, especially with a higher temporal resolution. Differences, for example, between the deciduous species are hypothesized, but could not be tested in this study. A dependence of the magnitude of the observed drop to LAI could also be tested. If the drop in backscatter is caused by the attenuation of the leaves, a dependence is credible. This could be tested by correlating VH backscatter time series with periodic LAI estimations derived from hemispherical images of deciduous forest stands in spring or autumn [
94].
This leads to the question of to what extent the findings of this study could be transferred to other ecosystems for time series analysis. In tropical or boreal ecosystems, where the use of SAR data is already well established, the use of multitemporal C-band data might contribute valuable new insights into the temporal evolution of these ecosystems.
5. Conclusions
This study showed that multitemporal C-band radiometrically calibrated and flattened backscatter data have the potential to supplement optical remote sensing data for use in forest mapping or ecological applications. Forests consisting of broadleaved, deciduous species, such as beech and oak, produce stronger backscatter in both polarisations than forests consisting of spruce. Typical seasonal phenological signatures were also observed within forested areas. In VH, areas consisting of deciduous forests showed a consistent and distinct intra-annual backscatter pattern with higher backscatter in winter and lower in summer. In coniferous forests, however, the opposite behaviour with lower backscatter in winter and higher in summer was observed. The same behaviour, but even more distinctive, was observed in coniferous forests at VV, whereas deciduous forests showed almost no seasonal cycle in VV.
These temporal signatures simplify the classification of mixed temperate forests because of the striking differences between the temporal signatures of deciduous vs. coniferous forests. Good classification performances were achieved for the forest types (OA of 86% and = 0.73) and for three different species (OA of 72% and = 0.58). The strong seasonal signal in VH allows the monitoring of phenology in deciduous forests. The dates independently reported for leaf emergence and leaf fall in three different study areas were satisfyingly matched. Almost all extracted break dates were within the error margin ( days) of the available temporal sampling.
These findings show that using multitemporal C-band data offers several new possibilities. Periodic and cost-effective mapping of mixed temperate forests will ease forest management to a great extent. In the near future, longer time series of S-1 and RADARSAT Constellation Mission data over several years will be available with high temporal resolution. Using them, monitoring at different sets of spatial and temporal scales of different ecosystems will be possible. These might lead to interesting ecological insights, such as an assessment of the shift of phenophases or a quantification of changes in species composition due to climate change.
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}