Tree Death Not Resulting in Gap Creation: An Investigation of Canopy Dynamics of Northern Temperate Deciduous Forests


Abstract

1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Lidar Data Processing
2.3. Field Sampling
2.4. Statistical Analysis
3. Results
3.1. Comparing the Structures and Mechanisms Models
3.2. Tree Damage Mechanisms
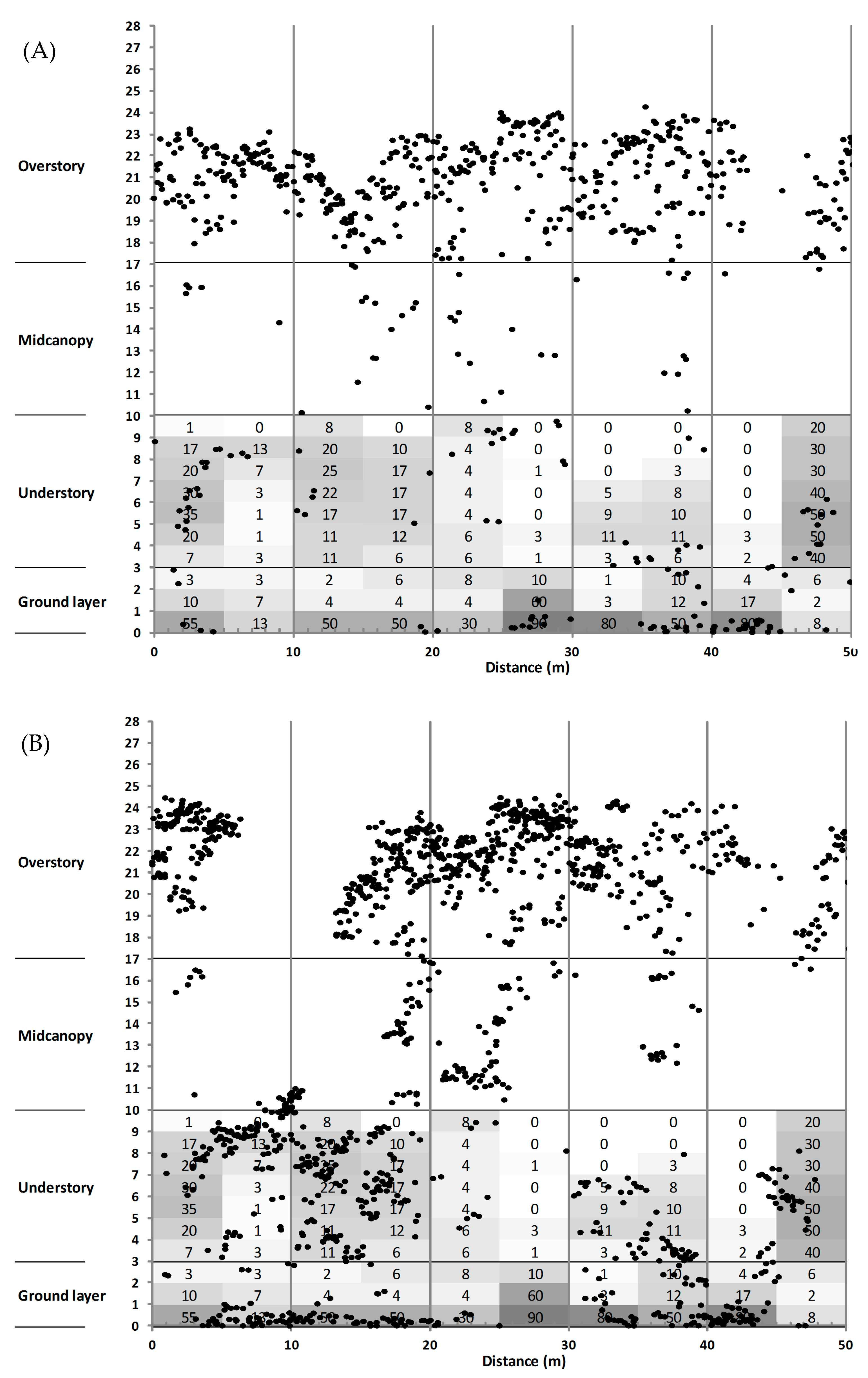
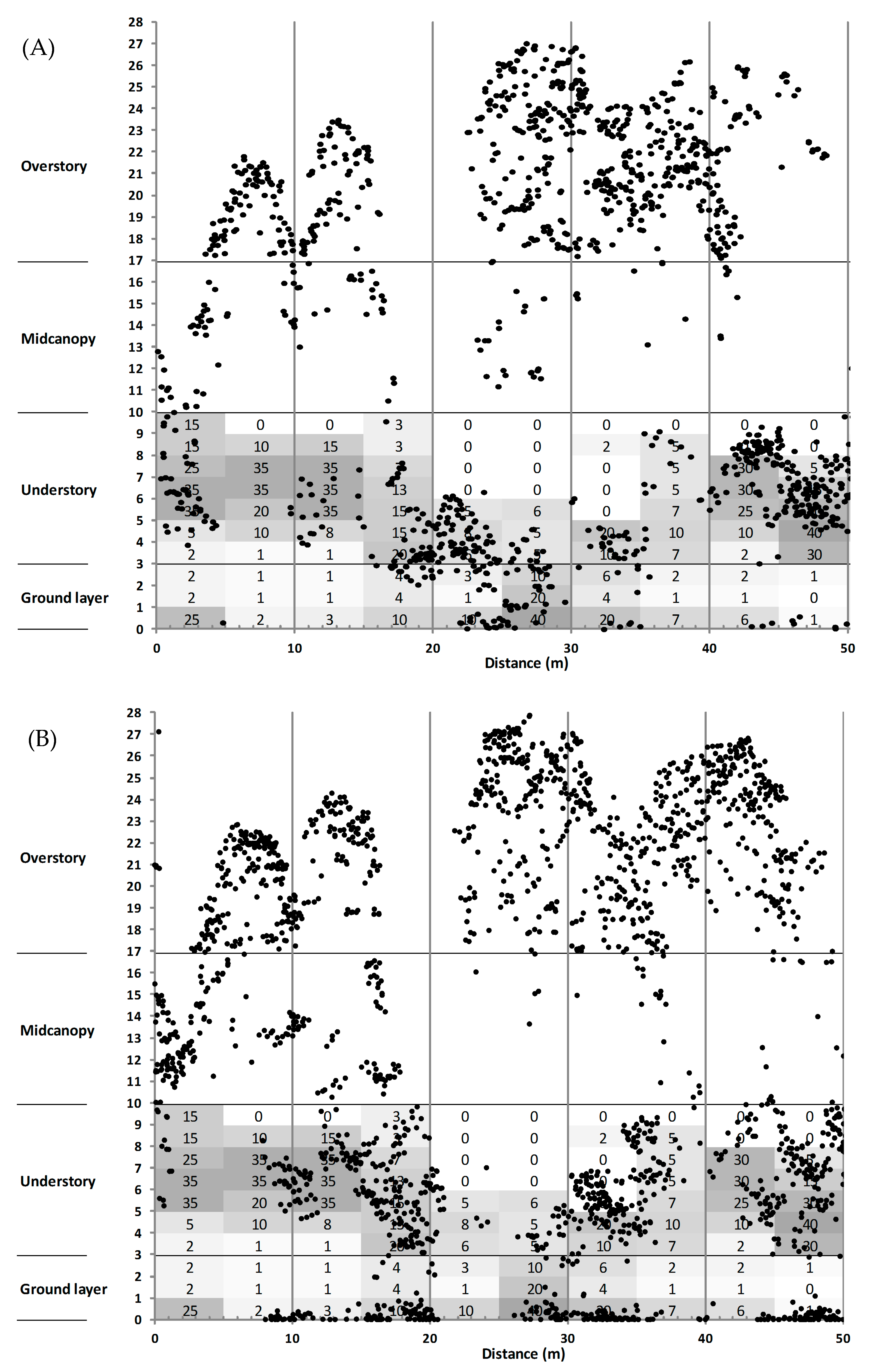
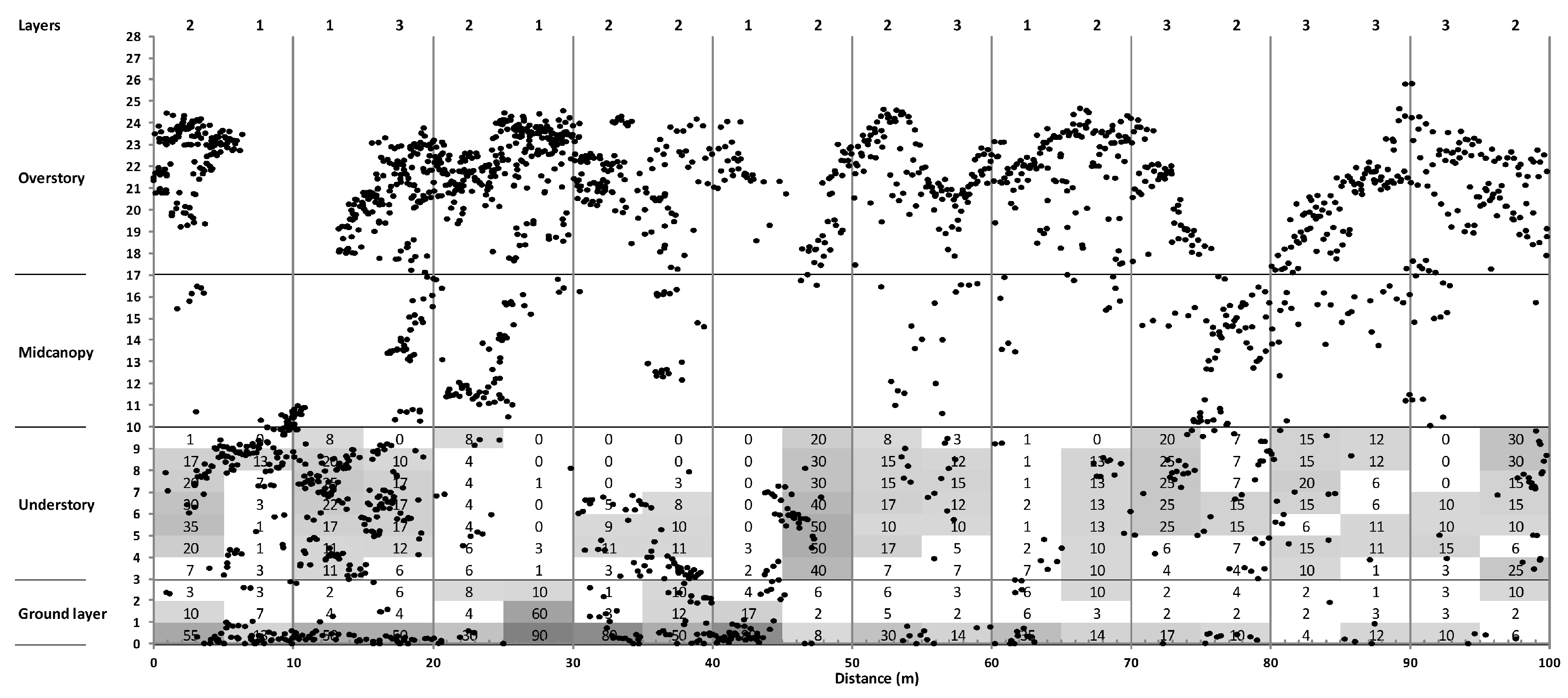
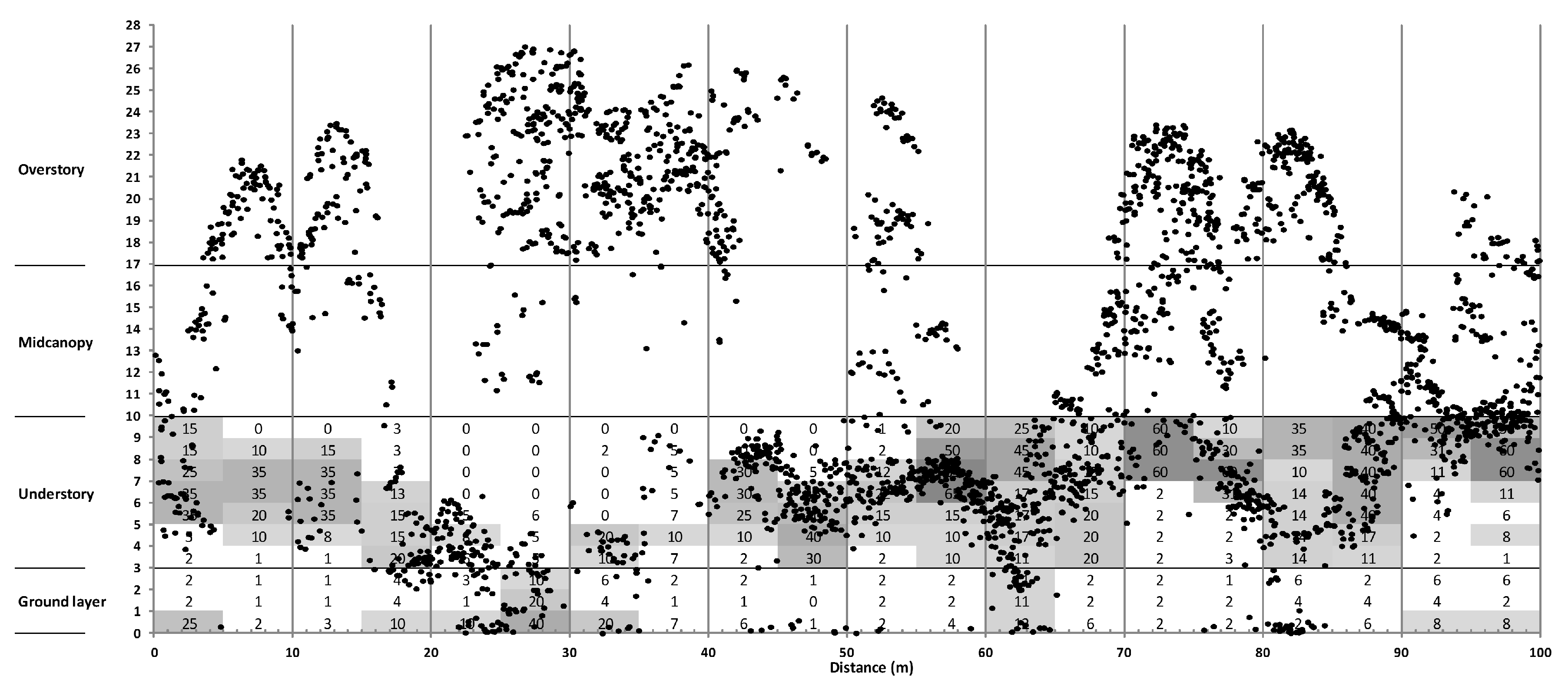
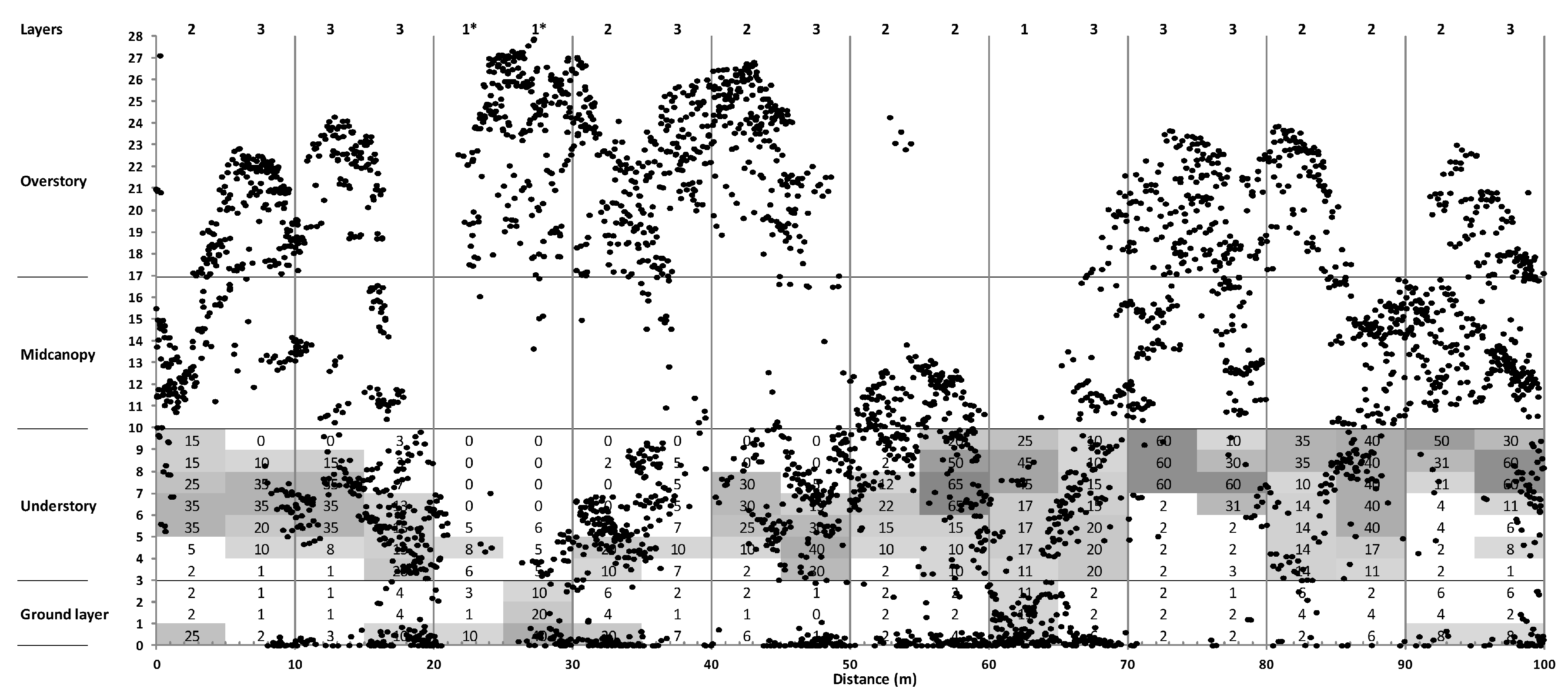
3.3. Canopy Structure
4. Discussion
4.1. Tree Damage Mechanisms
4.2. Sub-Canopy Structure as the Main Driver of New Gap Creation
4.3. Sylvicultural Implications
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A




References
- Yamamoto, S.I. Forest gap dynamics and tree regeneration. J. For. Res. 2000, 5, 223–229. [Google Scholar] [CrossRef]
- Beaudet, M.; Messier, C.; Leduc, A. Understorey light profiles in temperate deciduous forests: Recovery process following selection cutting. J. Ecol. 2004, 92, 328–338. [Google Scholar] [CrossRef]
- Runkle, J.R. Patterns of disturbance in some old-growth mesic forests of eastern north america. Ecology 1982, 63, 1533–1546. [Google Scholar] [CrossRef]
- Lertzman, K.P.; Sutherland, G.D.; Inselberg, A.; Saunders, S.C. Canopy gaps and the landscape mosaic in a coastal temperate rain forest. Ecology 1996, 77, 1254–1270. [Google Scholar] [CrossRef]
- McCarthy, J. Gap dynamics of forest trees: A review with particular attention to boreal forests. Environ. Rev. 2001, 9, 1–59. [Google Scholar] [CrossRef]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Schliemann, S.A.; Bockheim, J.G. Methods for studying treefall gaps: A review. For. Ecol. Manag. 2011, 261, 1143–1151. [Google Scholar] [CrossRef]
- Barden, L.S. Repeatability in forest gap research: Studies in the great smoky mountains. Ecology 1989, 70, 558–559. [Google Scholar] [CrossRef]
- Brokaw, N.V.L. The definition of treefall gap and its effect on measures of forest dynamics. Biotropica 1982, 14, 158–160. [Google Scholar] [CrossRef]
- Senecal, J.-F.; Doyon, F.; Messier, C. Management implications of varying gap detection height thresholds and other canopy dynamics processes in temperate deciduous forests. For. Ecol. Manag. 2018, 410, 84–94. [Google Scholar] [CrossRef]
- Bonnet, S.; Gaulton, R.; Lehaire, F.; Lejeune, P. Canopy gap mapping from airborne laser scanning: An assessment of the positional and geometrical accuracy. Remote Sens. 2015, 7, 11267–11294. [Google Scholar] [CrossRef]
- Hirata, Y.; Tanaka, H.; Furuya, N. Canopy and gap dynamics analysed using multi-temporal airborne laser scanner data in a temperate deciduous forest. In Proceedings of the International Conference on Lidar Applications in Forest Assessment and Inventory, Heriot-Watt University, Edinburgh, UK, 17–19 September 2008; pp. 144–150. [Google Scholar]
- St-Onge, B.; Vepakomma, U.; Sénécal, J.-F.; Kneeshaw, D.; Doyon, F. Canopy gap detection and analysis with airborne laser scanning. In Forestry Applications of Airborne Laser Scanning; Maltamo, M., Næsset, E., Vauhkonen, J., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 27, pp. 419–437. [Google Scholar]
- Vepakomma, U.; Kneeshaw, D.; Fortin, M.J. Spatial contiguity and continuity of canopy gaps in mixed wood boreal forests: Persistence, expansion, shrinkage and displacement. J. Ecol. 2012, 100, 1257–1268. [Google Scholar] [CrossRef]
- Seymour, R.S.; White, A.S.; Maynadier, P.G. Natural disturbance regimes in northeastern north america: Evaluating silvicultural systems using natural scales and frequencies. For. Ecol. Manag. 2002, 155, 357–367. [Google Scholar] [CrossRef]
- Boose, E.R.; Foster, D.R.; Fluet, M. Hurricane impacts to tropical and temperate forest landscapes. Ecol. Monogr. 1994, 64, 369–400. [Google Scholar] [CrossRef]
- Jones, E. The structure and reproduction oe the virgin forest of the north temperate zone. New Phytol. 1945, 44, 130–148. [Google Scholar] [CrossRef]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef]
- Hooper, M.; Arii, K.; Lechowicz, M. Impact of a major ice storm on an old-growth hardwood forest. Botany 2001, 79, 70–75. [Google Scholar] [CrossRef]
- Lafon, C.W. Ice-storm disturbance and long-term forest dynamics in the adirondack mountains. J. Veg. Sci. 2004, 15, 267–276. [Google Scholar] [CrossRef]
- Pisaric, M.; King, D.; MacIntosh, A.; Bemrose, R. Impact of the 1998 ice storm on the health and growth of sugar maple (Acer saccharum marsh.) dominated forests in gatineau park, quebec 1. J. Torrey Bot. Soc. 2008, 135, 530–539. [Google Scholar] [CrossRef]
- Tremblay, M.; Messier, C.; Marceau, D. Analysis of deciduous tree species dynamics after a severe ice storm using sortie model simulations. Ecol. Model. 2005, 187, 297–313. [Google Scholar] [CrossRef]
- White, J. The plant as a metapopulation. Annu. Rev. Ecol. Syst. 1979, 10, 109–145. [Google Scholar] [CrossRef]
- Beaudet, M.; Brisson, J.; Gravel, D.; Messier, C. Effect of a major canopy disturbance on the coexistence of Acer saccharum and fagus grandifolia in the understorey of an old-growth forest. J. Ecol. 2007, 95, 458–467. [Google Scholar] [CrossRef]
- Lorimer, C.G.; Frelich, L.E. A methodology for estimating canopy disturbance frequency and intensity in dense temperate forests. Can. J. For. Res. 1989, 19, 651–663. [Google Scholar] [CrossRef]
- Krasny, M.E.; DiGregorio, L.M. Gap dynamics in allegheny northern hardwood forests in the presence of beech bark disease and gypsy moth disturbances. For. Ecol. Manag. 2001, 144, 265–274. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; White, P.S. The Ecology of Natural Disturbance and Patch Dynamics; Academic Press: San Diego, CA, USA, 1985; p. 475. [Google Scholar]
- Nadkarni, N.M.; McIntosh, A.C.S.; Cushing, J.B. A framework to categorize forest structure concepts. For. Ecol. Manag. 2008, 256, 872–882. [Google Scholar] [CrossRef]
- Parker, G.G. Structure and microclimate of forest canopies. In Forest Canopies, 1st ed.; Lowman, M., Nadkarni, N.M., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 73–106. [Google Scholar]
- Connell, J.; Lowman, M.; Noble, I. Subcanopy gaps in temperate and tropical forests. Aust. J. Ecol. Oxf. 1997, 22, 163–168. [Google Scholar] [CrossRef]
- Lieberman, M.; Lieberman, D.; Peralta, R. Forests are not just swiss cheese: Canopy stereogeometry of non-gaps in tropical forests. Ecology 1989, 70, 550–552. [Google Scholar] [CrossRef]
- Majcen, Z. Historique des coupes de jardinage dans les forêts inéquiennes au Québec. Rev. For. Fr. 1994, 46, 9. [Google Scholar] [CrossRef]
- Robitaille, A.; Saucier, J.-P. Paysages Régionaux du Québec Méridional; Publications du Québec: Quebec City, QC, Canada, 1998; p. 213. [Google Scholar]
- Chabot, M. Dommages Causés à la Forêt par le Verglas de Janvier 1998: Résultats Préliminaires d'une Reconnaissance Aérienne Réalisée Entre le 19 Janvier et le 4 Février 1998; Direction de la Conservation des Forêts: Quebec City, QC, Canada, 1998; p. 16. [Google Scholar]
- Houston, D.R. Major new tree disease epidemics: Beech bark disease. Annu. Rev. Phytopathol. 1994, 32, 75–87. [Google Scholar] [CrossRef]
- Vepakomma, U.; St-Onge, B.; Kneeshaw, D. Spatially explicit characterization of boreal forest gap dynamics using multi-temporal lidar data. Remote Sens. Environ. 2008, 112, 2326–2340. [Google Scholar] [CrossRef]
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H.; Fischer, E.; Gerlitz, L.; Wehberg, J.; Wichmann, V.; Böhner, J. System for automated geoscientific analyses (saga). Geosci. Model Dev. 2015, 8, 1991–2007. [Google Scholar] [CrossRef]
- Anderson, D.R. Model Based Inference in the Life Sciences: A Primer on Evidence; Springer: New York, NY, USA, 2008. [Google Scholar]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression; Wiley-Interscience: Hoboken, NJ, USA, 2000; Volume 354, p. 392. [Google Scholar]
- Senecal, J.-F. Dynamique Spatio-Temporelle des Trouées en Forêt Feuillue Tempérée par Télédétection Lidar; Université du Québec à Montréal: Montréal, QC, Canada, 2011. [Google Scholar]
- DiGregorio, L.M.; Krasny, M.E.; Fahey, T.J. Radial growth trends of sugar maple (Acer saccharum) in an allegheny northern hardwood forest affected by beech bark disease. J. Torrey Bot. Soc. 1999, 126, 245–254. [Google Scholar] [CrossRef]
- Armesto, J.; Mitchell, J.; Villagran, C. A comparison of spatial patterns of trees in some tropical and temperate forests. Biotropica 1986, 18, 1–11. [Google Scholar] [CrossRef]
- Everham, E.M.; Brokaw, N.V. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 113–185. [Google Scholar] [CrossRef]
- Proulx, O.; Greene, D. The relationship between ice thickness and northern hardwood tree damage during ice storms. Can. J. For. Res. 2001, 31, 1758–1767. [Google Scholar] [CrossRef]
- Schaetzl, R.J.; Burns, S.F.; Johnson, D.L.; Small, T.W. Tree uprooting: Review of impacts on forest ecology. Vegetatio 1988, 79, 165–176. [Google Scholar] [CrossRef]
- Barden, L.S. Forest development in canopy gaps of a diverse hardwood forest of the southern appalachian mountains. Oikos 1981, 37, 205–209. [Google Scholar] [CrossRef]
- Bater, C.W.; Wulder, M.A.; Coops, N.C.; Nelson, R.F.; Hilker, T.; Nasset, E. Stability of sample-based scanning-lidar-derived vegetation metrics for forest monitoring. IEEE Trans. Geosci. Remote Sens. 2011, 49, 2385–2392. [Google Scholar] [CrossRef]
- Liu, J.; Skidmore, A.K.; Jones, S.; Wang, T.; Heurich, M.; Zhu, X.; Shi, Y. Large off-nadir scan angle of airborne lidar can severely affect the estimates of forest structure metrics. ISPRS J. Photogramm. Remote Sens. 2018, 136, 13–25. [Google Scholar] [CrossRef]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef]
- Nolet, P.; Doyon, F.; Messier, C. A new silvicultural approach to the management of uneven-aged northern hardwoods: Frequent low-intensity harvesting. Forestry 2013, 87, 39–48. [Google Scholar] [CrossRef]
- Runkle, J.R. Disturbance regimes in temperate forests. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1985. [Google Scholar]
- Woods, K.D. Intermediate disturbance in a late-successional hemlock-northern hardwood forest. J. Ecol. 2004, 92, 464–476. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Kolasa, J.; Armesto, J.J.; Collins, S.L. The ecological concept of disturbance and its expression at various hierarchical levels. Oikos 1989, 54, 129–136. [Google Scholar] [CrossRef]
- Puettmann, K.J.; Coates, K.D.; Messier, C.C. A Critique of Silviculture: Managing for Complexity; Island Press: Washington, DC, USA, 2012. [Google Scholar]
- Woods, K.D.; Hicks, D.J.; Schultz, J. Losses in understory diversity over three decades in an old-growth cool-temperate forest in Michigan, USA. Can. J. For. Res. 2012, 42, 532–549. [Google Scholar] [CrossRef]
- Franklin, J.F.; Shugart, H.H.; Harmon, M.E. Tree death as an ecological process. BioScience 1987, 37, 550–556. [Google Scholar] [CrossRef]
- Canham, C.D. Suppression and release during canopy recruitment in Acer saccharum. Bull. Torrey Bot. Club 1985, 112, 134–145. [Google Scholar] [CrossRef]
- Canham, C.D. Suppression and release during canopy recruitment in fagus grandifolia. Bull. Torrey Bot. Club 1990, 117, 1–7. [Google Scholar] [CrossRef]
- Poulson, T.; Platt, W. Replacement patterns of beech and sugar maple in warren woods, Michigan. Ecology 1996, 77, 1234–1253. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acquisition Time | September 2007 | August 2013 |
|---|---|---|
| Sensor model | Optech ALTM 3100 | Optech ALTM Gemini |
| Average point density | >3.0 pts/m2 | >4 pts/m2 |
| Max. half-scan angle | 20° | 18° |
| Scan rate | 41 Hz | 55 Hz |
| Altitude | 1300 m | 650 m |
| Line spacing | 750 m | 261 m |
| Target overlap | 50% | 50% |
| Model | Formula |
|---|---|
| MECHANISMS | GAP~Standing + Bent + Broken + Branch |
| STRUCTURE | GAP~AreaHRA + AdjacencyGap |
| Model | K | AICc | Delta AICc | AICc Weight | Log L. |
|---|---|---|---|---|---|
| STRUCTURE | 4 | 89.89 | 0 | 1 | −40.95 |
| MECHANISMS | 6 | 120.44 | 30.55 | 0 | −54.22 |
| Broken bole | Tree bent | |||||
| Absence | Presence | Absence | Presence | |||
| No gap | 55.5% | 21.1% | No gap | 58.6% | 18.0% | |
| Gap | 9.4% | 14.1% | Gap | 19.5% | 3.9% | |
| Dead standing | Broken branch | |||||
| Absence | Presence | Absence | Presence | |||
| No gap | 43.0% | 33.6% | No gap | 50.0% | 26.6% | |
| Gap | 13.3% | 10.2% | Gap | 14.8% | 8.6% | |
| Uprooted | Adjacency to older gap | |||||
| Absence | Presence | Absence | Presence | |||
| No gap | 75.8% | 0.8% | No gap | 66.4% | 10.2% | |
| Gap | 21.1% | 2.3% | Gap | 7.0% | 16.4% | |
| Site | 1 Layer | 2 Layers | 3 Layers |
|---|---|---|---|
| Unmanaged forest | 25 (15) | 45 | 30 |
| Forest cut in 1993 | 15 (10) | 40 | 45 |
| Temperate forest 39°N 1 | 23 | 47 | 28 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senécal, J.-F.; Doyon, F.; Messier, C. Tree Death Not Resulting in Gap Creation: An Investigation of Canopy Dynamics of Northern Temperate Deciduous Forests. Remote Sens. 2018, 10, 121. https://doi.org/10.3390/rs10010121
Senécal J-F, Doyon F, Messier C. Tree Death Not Resulting in Gap Creation: An Investigation of Canopy Dynamics of Northern Temperate Deciduous Forests. Remote Sensing. 2018; 10(1):121. https://doi.org/10.3390/rs10010121
Chicago/Turabian StyleSenécal, Jean-Francois, Frédérik Doyon, and Christian Messier. 2018. "Tree Death Not Resulting in Gap Creation: An Investigation of Canopy Dynamics of Northern Temperate Deciduous Forests" Remote Sensing 10, no. 1: 121. https://doi.org/10.3390/rs10010121
APA StyleSenécal, J.-F., Doyon, F., & Messier, C. (2018). Tree Death Not Resulting in Gap Creation: An Investigation of Canopy Dynamics of Northern Temperate Deciduous Forests. Remote Sensing, 10(1), 121. https://doi.org/10.3390/rs10010121