
Phenotypic Responses of Twenty Diverse Proso Millet (Panicum miliaceum L.) Accessions to Irrigation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Location
2.2. Experimental Design and Data Collection
2.3. Statistical Analysis
3. Results
3.1. Yield
3.2. Emergence and Plant Height
3.3. Days to Heading and Maturity
4. Discussion
4.1. Yield
4.2. Emergence and Plant Height
4.3. Days to Heading and Maturity
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ceccarelli, S.; Grando, S. Drought as a challenge for the plant breeder. In Drought Tolerance in Higher Plants: Genetical, Physiological and Molecular Biological Analysis; Springer: Dutch, The Netherlands, 1996; pp. 71–77. [Google Scholar]
- Shwabe, K.; Albiac-Mutillo, J.; Connor, J.D.; Hassan, R.; Meza, G.L. (Eds.) Drought in Arid and Semi-Arid Regions; Springer: Dutch, The Netherlands, 2013.
- Seghatoleslami, M.J.; Kafi, M.; Majidi, E. Effect of drought stress at different growth stages on yield and water use efficiency of five proso millet (Panicum miliaceum L.) genotypes. Pak. J. Bot. 2008, 40, 1427–1432. [Google Scholar]
- Padulosi, S.; Mal, B.; King, O.I.; Gotor, E. Minor millets as a central element for sustainably enhanced incomes, empowerment, and nutrition in rural India. Sustainability 2015, 7, 8904–8933. [Google Scholar] [CrossRef]
- Reddy, V.G.; Upadhyaya, H.D.; Gowda, C.L.L. Morphological characterization of world’s proso millet germplasm collection. J. SAT Agric. Res. 2007, 3, 4. [Google Scholar]
- Baltensperger, D.D. Progress with proso, pearl and other millets. In Trends in New Crops and New Uses; ASHS Press: Alexandria, VA, USA, 2002; pp. 100–103. [Google Scholar]
- Amadou, I.; Gounga, M.E.; Le, G.W. Millets: Nutritional composition, some health benefits and processing—A review. Emirates J. Food Agric. 2013, 25, 501–508. [Google Scholar]
- Baltensperger, D.; Lyon, D.J.; Anderson, R.; Holman, T.; Stymieste, C.; Shanahan, J.; Krall, J. EC95-137 Producing and Marketing Proso Millet in the High Plains; Historical Materials from University of Nebraska-Lincoln Extension; DigitalCommons@University of Nebraska–Lincoln: Lincoln, NE, USA, 1995; p. 709. [Google Scholar]
- USDA-NASS (USDA—National Agricultural Statistics Service). Crop Production 2015 Summary; USDA-NASS: Washington, DC, USA, 2016.
- USDA FAS. Increased US Millet Exports to The Netherlands; Global Agricultural Information Network Report Number: NL2009; USDA-Foreign Agricultural Service: Washington, DC, USA, 2012; pp. 1–2.
- McDonald, S.K.; Hofsteen, L.; Downey, L. Crop Profile for Proso Millet in Colorado; USDA Crop Profiles; Regional IPM Centers: Raleigh, NC, USA, 2003.
- Lyon, D.J.; Burgener, P.A.; De Boer, K.L.; Harveson, R.M.; Hein, G.L.; Hergert, G.W.; Holman, T.L.; Nelson, L.A.; Johnson, J.J.; Nleya, T.; et al. Producing and Marketing of Proso Millet in Great Plains (EC137); University of Nebraska–Lincoln Extension: Lincoln, NE, USA, 2008. [Google Scholar]
- Dipak, K.S. Proso Millet Varieties for Western Nebraska; Nebraska Guide, NIAR; University of Nebraska: Lincoln, NE, USA, 2013. [Google Scholar]
- Anderson, R.L. Planting date effect on no-till proso millet. J. Prod. Agric. 1994, 7, 454–458. [Google Scholar] [CrossRef]
- AgWeatherNet, Washington State University. Pullman Station Historical Data. 2016. Available online: http://weather.wsu.edu/index.php?p=88650 (accessed on 2 March 2017).
- Miller, D.T. Growth Stages of Wheat: Identification and Understanding Improve Crop Management; Potash & Phosphate Institute (PPI): Norcorss, GA, USA, 1992; pp. 1–4. [Google Scholar]
- Wise, K.; Johnson, B.; Mansfield, C.; Krupke, C. Managing Wheat by Growth Stage; Purdue Ext. ID-422; Purdue University: West Lafayette, IN, USA, 2011. [Google Scholar]
- Hommel, G. A stagewise rejective multiple test procedure based on a modified Bonferroni test. Biometrika 1988, 75, 383–386. [Google Scholar] [CrossRef]
- Wright, S.P. Adjusted p-values for simultaneous inference. Biometrics 1992, 48, 1005–1013. [Google Scholar] [CrossRef]
- Smiciklas, K.D.; Mullen, R.E.; Carlson, R.E.; Knapp, A.D. Soybean seed quality response to drought stress and pod position. Agron. J. 1992, 84, 166–170. [Google Scholar] [CrossRef]
- Ney, B.; Duthion, C.; Ture, O. Phonological responses of pea to water stress during reproductive development. Crop Sci. 1994, 34, 141–146. [Google Scholar]
- Westgate, M.E. Water status and development of the maize endosperm and embryo during drought. Crop Sci. 1994, 34, 76–83. [Google Scholar] [CrossRef]
- Richards, R.A.; Rebetzke, G.J.; Condon, A.G.; van Herwaarden, A.F. Breeding opportunities for increasing the efficiency of water use and crop yield in temperate cereals. Crop Sci. 2002, 42, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Umar, S. Alleviating adverse effects of water stress on yield of sorghum, mustard and groundnut by potassium application. Pak. J. Bot. 2006, 38, 1373–1380. [Google Scholar]
- Mahalakshmi, V.; Bidinger, F.R. Flowering response of Pearl millet to water stress during panicle development. Ann. Appl. Biol. 1985, 106, 571–578. [Google Scholar] [CrossRef]
- Yadav, R.S.; Hash, C.T.; Bidinger, F.R.; Dhanoa, M.S.; Howarth, C.J. Identification and utilization of quantitative trait loci to improve terminal drought tolerance in pearl millet (Pennisetum glaucum (L.) R. Br.). In Proceedings of the Workshop on Molecular Approaches for the Genetic Improvement of Cereals for Stable Production in Water-Limited Environments, CIMMYT, El Batan, Mexico, 21–25 June 1999; Ribaut, J.M., Poland, D., Eds.; pp. 108–114.
- Ludlow, M.M.; Muchow, R.C. A critical evaluations of traits for improving crop yields in water-limited environments. Adv. Agron. 1990, 43, 107–153. [Google Scholar]
- Hussain, M.; Malik, M.A.; Farooq, M.; Ashraf, M.Y.; Cheema, M.A. Improving drought tolerance by exogenous application of glycinebetaine and salicylic acid in sunflower. J. Agron. Crop Sci. 2008, 194, 193–199. [Google Scholar] [CrossRef]
- Harris, D.; Tripathi, R.S.; Joshi, A. On-farm seed priming to improve crop establishment and yield in dry direct-seeded rice. In Direct Seeding: Research Strategies and Opportunities; Pandey, S., Mortimer, M., Wade, L., Tuong, T.P., Lopes, K., Hardy, B., Eds.; International Research Institute: Manila, Philippines, 2002; pp. 231–240. [Google Scholar]
- Kaya, M.D. The role of hull in germination and salinity tolerance in some sunflower (Helianthus annuus L.) cultivars. Afr. J. Biotechnol. 2009, 8, 597–600. [Google Scholar]
- Conover, D.G.; Sovonick-Dunford, S.A. Influence of water deficits on the water relations and growth of Echinochloa turneriana, Echinochloa crus-galli, and Pennisetum americanum. Funct. Plant Biol. 1989, 16, 291–304. [Google Scholar]
- Madakadze, I.C. Switchgrass biomass and chemical composition for biofuel in Eastern Canada. Agron. J. 1999, 97, 696–701. [Google Scholar] [CrossRef]
- Nonami, H. Plant water relations and control of cell elongation at low water potentials. J. Plant Res. 1998, 111, 373–382. [Google Scholar]
- Bidinger, F.R.; Mahalakshmi, V.; Rat, G.D.P. Assessment of drought resistance in pearl millet (Pennisetum americanum (L.) Leeke). I. Factors affecting yields under stress. Aust. J. Agric. Res. 1987, 38, 37–48. [Google Scholar] [CrossRef]
- Passioura, J.B. Drought and drought tolerance. Plant Growth Regul. 1996, 20, 79–83. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H. Grain filling of cereals under soil drying. New Phytol. 2006, 169, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Stone, P. The effects of heat stress on cereal yield and quality. In Crop Responses and Adaptations to Temperature Stress; Basra, A.S., Ed.; Food Products Press: Binghamton, NY, USA, 2001; pp. 243–291. [Google Scholar]
- Aspinall, D.; Nicholls, P.B.; May, L.H. The effects of soil moisture stress on the growth of barley: I. Vegetative development and grain yield. Aust. J. Agric. Res. 1964, 15, 729–745. [Google Scholar] [CrossRef]
- Aspinall, D. The effects of soil moisture stress on the growth of barley: II. Grain Growth. Aust. J. Agric. Res. 1965, 16, 265–275. [Google Scholar] [CrossRef]
- Wells, S.A.; Dubetz, S. Reaction of barley varieties to soil water stress. Can. J. Plant Sci. 1966, 46, 507–512. [Google Scholar] [CrossRef]
- Wopereis, M.C.S.; Kropff, M.J.; Maligaya, A.R.; Tuong, T.P. Drought-stress responses of two lowland rice cultivars to soil water status. Field Crops Res. 1996, 46, 21–39. [Google Scholar] [CrossRef]
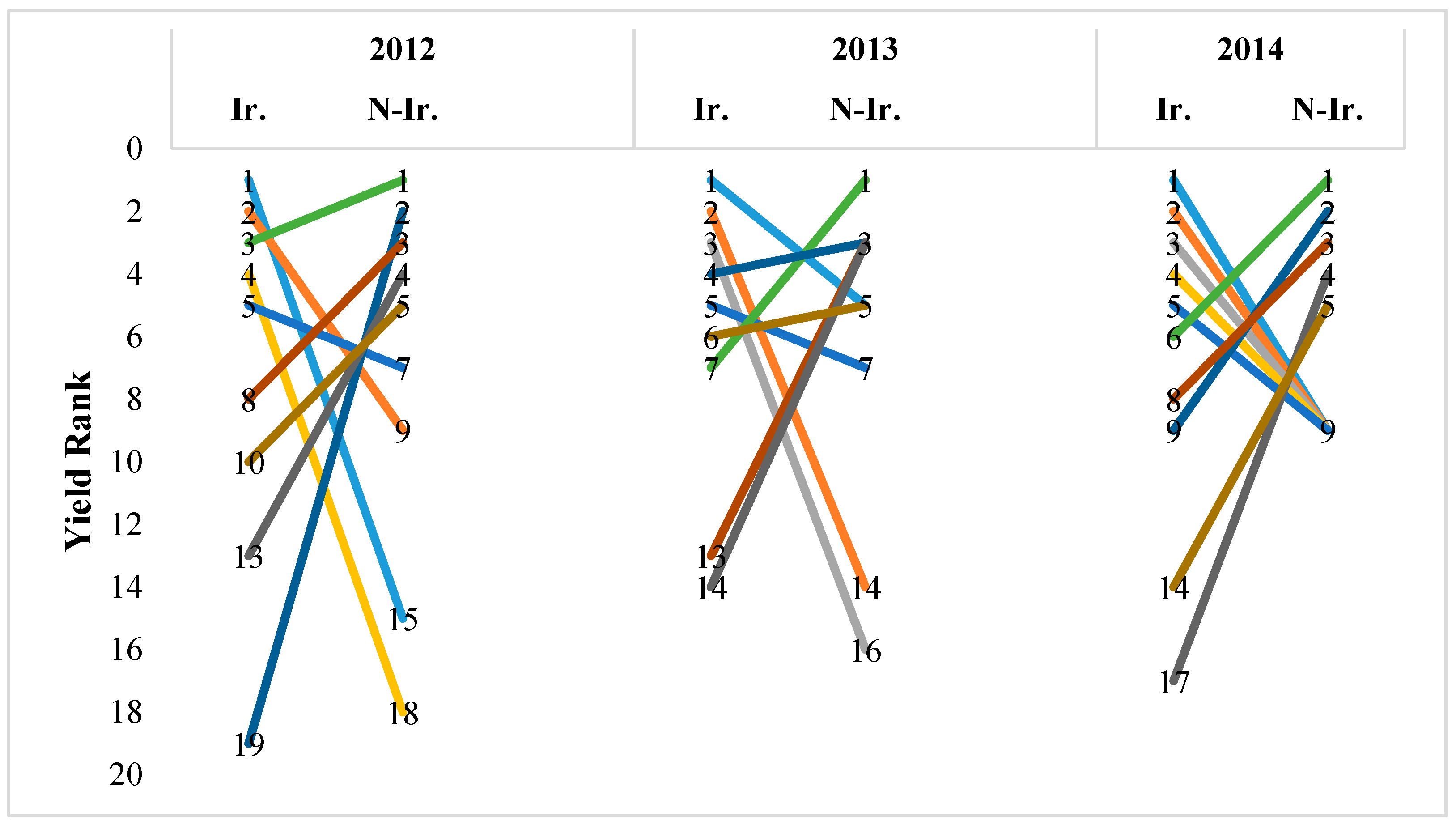

{kind=link}
| Year | Months | Growing Degree Days at Planting | Total Precipitation [mm] | Average Maximum Day Temperature [°C] |
|---|---|---|---|---|
| 2012 | June | 748 | 42.93 | 19.8 |
| July | 0 | 27.8 | ||
| August | 0 | 29.5 | ||
| September | 0 | 24.6 | ||
| 2013 | June | 862 | 54.36 | 21.6 |
| July | 4.83 | 29.6 | ||
| August | 6.35 | 28.9 | ||
| September | 55.12 | 22.1 | ||
| 2014 | June | 881 | 19.05 | 21.2 |
| July | 11.18 | 30.8 | ||
| August | 9.14 | 28.8 | ||
| September | 4.06 | 24.2 |
| Item | Accession Number | Accession Name | Year Collected | Origin | Latitude | Longitude |
|---|---|---|---|---|---|---|
| 1 | PI 171727 | Dari | 1948 | Bolu, Turkey | 40.6792°N | 31.5583°E |
| 2 | PI 346937 | Tlicevskoje | 1969 | Former Soviet Union | N/A | N/A |
| 3 | PI 517017 | GR 658 | 1986 | Ouarzazate, Morocco | 30.9167°N | 6.9167°W |
| 4 | PI 517018 | GR 664 | 1986 | Ouarzazate, Morocco | 30.9335°N | 6.9370°W |
| 5 | PI 517019 | GR 665 | 1986 | Ouarzazate, Morocco | 30.9335°N | 6.9370°W |
| 6 | PI 531398 | Bolgar 159 | 1989 | Bulgaria | 42.7500°N | 25.5000°E |
| 7 | PI 531410 | Kamusinszkoe 67 | 1989 | Former Soviet Union | N/A | N/A |
| 8 | PI 531411 | Komsomolskoe 996 | 1989 | Former Soviet Union | N/A | N/A |
| 9 | PI 531412 | Kazanskoe 176 | 1989 | Former Soviet Union | N/A | N/A |
| 10 | PI 531429 | Tuvinskoe | 1989 | Former Soviet Union | N/A | N/A |
| 11 | PI 531430 | Veszelopodoljanszkoe 403 | 1989 | Former Soviet Union | N/A | N/A |
| 12 | PI 531431 | Unikum | 1989 | Czechoslovakia | 50.0833°N | 14.4167°E |
| 13 | PI 536011 | Sunup | 1989 | Nebraska, United States | 41.2324°N | 98.4160°W |
| 14 | PI 578073 | Earlybird | 1994 | Nebraska, United States | 41.2324°N | 98.4160°W |
| 15 | PI 578074 | Huntsman | 1994 | Nebraska, United States | 41.2324°N | 98.4160°W |
| 16 | PI 583347 | Sunrise | 1994 | Nebraska, United States | 41.2324°N | 98.4160°W |
| 17 | PI 649382 | Turghai | 1961 | North Dakota, United States | 47.0000°N | 100.0000°W |
| 18 | PI 649385 | Minsum | 1980 | Minnesota, United States | 46.0000°N | 94.0000°W |
| 19 | PI 654403 | TU-85-074-03 | 1986 | Bitlis, Turkey | 38.4000°N | 42.1083°E |
| 20 | PI 654404 | TU-85-087-01 | 1986 | Bitlis, Turkey | 38.4000°N | 42.1083°E |
| Years | Effect | DF | Emergence Rate | PH1 | PH2 | DH | DM | Yield |
|---|---|---|---|---|---|---|---|---|
| 2012 | Irrigation Variety Irrigation × Variety | 1 19 19 | N/A | 18.06 * 3.20 *** | 197.77 ** 3.34 *** 1.78 * | 46.94 *** | 12.87 *** | 81.89 *** 2.51 ** 2.30 ** |
| 2013 | Irrigation Variety Irrigation × Variety | 1 19 | 22.26 *** 248.74 *** 39.42 ** | 1304.54 *** 3.37 *** | 2062.74 *** 8.36 *** 4.46 *** | 13.31 17.59 *** 6.05 *** (DF = 11) | 101.94 *** 40.39 *** 3.13 ** (DF = 14)) | 3.35 *** 2.37 ** |
| 2014 | Irrigation Variety Irrigation × Variety | 1 19 14 | 739.35 *** 91.04 *** 76.58 *** | 608.05 *** | 122.48 *** 2.16 * | 96.97 *** | 13.27 *** 41.60 *** 6.35 *** | 43.03 *** 2.99 *** |
| Plant Name/Year | Emergence | PH1 | PH2 | DH | DM | Yield |
|---|---|---|---|---|---|---|
| 2012 | ||||||
| Dari | NDC | 22.98 * | 4.12 *** | NIE | NIE | 0.63 |
| Tlicevskoje | NDC | 4.45 *** | NIE | NIE | 1.68 | |
| GR 658 | NDC | 6.20 *** | NIE | NIE | 0.71 * | |
| GR 664 | NDC | 6.00 *** | NIE | NIE | 1.60 | |
| GR 665 | NDC | 8.16 *** | NIE | NIE | 2.76 ** | |
| Bolgar 159 | NDC | 4.00 *** | NIE | NIE | 1.18 | |
| Kamusinszkoe 67 | NDC | 4.85 *** | NIE | NIE | 4.00 *** | |
| Komsomolskoe 996 | NDC | 5.14 *** | NIE | NIE | 2.89 ** | |
| Kazanskoe 176 | NDC | 6.32 *** | NIE | NIE | 3.82 *** | |
| Tuvinskoe | NDC | 2.32 * | NIE | NIE | 0.05 | |
| Veszelopodoljanszkoe 403 | NDC | 5.18 *** | NIE | NIE | 4.11 *** | |
| Unikum | NDC | 5.55 *** | NIE | NIE | 4.00 *** | |
| Sunup | NDC | 3.18 ** | NIE | NIE | 3.80 *** | |
| Earlybird | NDC | 4.12 *** | NIE | NIE | 2.83 ** | |
| Huntsman | NDC | 5.38 *** | NIE | NIE | 2.17 * | |
| Sunrise | NDC | 3.34 ** | NIE | NIE | 3.21 ** | |
| Turghai | NDC | 4.40 *** | NIE | NIE | 0.35 | |
| Minsum | NDC | 5.10 *** | NIE | NIE | 0.29 | |
| TU-85-074-03 | NDC | 4.57 *** | NIE | NIE | −0.50 | |
| TU-85-087-01 | NDC | 5.55 *** | NIE | NIE | 1.26 | |
| 2013 | ||||||
| Dari | 1.04 | 14.39 *** | −2.86 ** | −2.71 ** | ||
| Tlicevskoje | 8.73 ** | n/a | n/a | −3.25 ** | ||
| GR 658 | 11.22 *** | 17.42 *** | 3.44 ** | n/a | ||
| GR 664 | 6.41 * | 17.43 *** | −0.42 | −3.83 *** | ||
| GR 665 | 1.93 | 19.35 *** | n/a | −4.75 *** | ||
| Bolgar 159 | 3.09 | 10.61 *** | 0.19 | 0.02 | ||
| Kamusinszkoe 67 | 6.87 ** | 9.73 *** | 0.07 | −0.21 | ||
| Komsomolskoe 996 | 3.83 | 16.20 *** | −3.23 ** | −4.53 *** | ||
| Kazanskoe 176 | 1.51 | 15.17 *** | −0.00 | −0.00 | ||
| Tuvinskoe | 2.52 | 63.32 *** | 17.05 *** | −3.64 *** | −6.61 *** | 95.04 *** |
| Veszelopodoljanszkoe 403 | 1.55 | 16.04 *** | −3.61 ** | −2.65 * | ||
| Unikum | 2.86 | 13.32 *** | −4.21 *** | −4.75 *** | ||
| Sunup | 0.07 | 13.44 *** | n/a | −2.10 * | ||
| Earlybird | 0.60 | 12.60 *** | n/a | −2.10 * | ||
| Huntsman | 0.13 | n/a | n/a | n/a | ||
| Sunrise | 1.14 | n/a | n/a | n/a | ||
| Turghai | 0.21 | 16.98 *** | n/a | n/a | ||
| Minsum | 3.88* | 9.70 *** | n/a | −4.64 *** | ||
| TU-85-074-03 | 0.07 | n/a | −3.49 ** | −1.88 | ||
| 2014 | ||||||
| Dari | 18.43 *** | NIE | −0.41 | |||
| Tlicevskoje | 1720.2 *** | NIE | n/a | |||
| GR 658 | 32.63 *** | NIE | −0.65 | |||
| GR 664 | 21.14 *** | NIE | −0.00 | |||
| GR 665 | 34.63 *** | NIE | −0.65 | |||
| Bolgar 159 | 1554.7 *** | NIE | n/a | |||
| Kamusinszkoe 67 | 1516.2 *** | NIE | n/a | |||
| Komsomolskoe 996 | 27.58 *** | NIE | −6.86 *** | |||
| Kazanskoe 176 | 33.89 *** | NIE | −7.03 *** | |||
| Tuvinskoe | 12.93 *** | 2.47 *** | 65.49 *** | NIE | 0.87 * | 3.37 *** |
| Veszelopodoljanszkoe 403 | 29.20 *** | NIE | −1.31 * | |||
| Unikum | 40.80 *** | NIE | −0.46 | |||
| Sunup | 14.59 *** | NIE | 0.44 | |||
| Earlybird | 14.89 *** | NIE | 1.23 * | |||
| Huntsman | 8.65 ** | NIE | 0.00 | |||
| Sunrise | 16.22 *** | NIE | 0.00 | |||
| Turghai | 1.59 | NIE | 0.55 | |||
| Minsum | 1431.1 *** | NIE | n/a | |||
| TU-85-074-03 | 1521.0 *** | NIE | n/a | |||
| TU-85-087-01 | 15.86 *** | NIE | 0.00 |
| Plant Name/Year | Emergence Rate (%) | PH1 (cm) | PH2 (cm) | DH (Day) | DM (Day) | Yield (g/Plot) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ir. | N-Ir. | Ir. | N-Ir. | Ir. | N-Ir. | Ir. | N-Ir. | Ir. | N-Ir. | Ir. | N-Ir. | |
| 2012 | ||||||||||||
| Dari | n/a | n/a | 93 | 68 | 135 | 101 | 47 | 46 | 90 | 88 | 28 | 16 |
| Tlicevskoje | n/a | n/a | 74 | 58 | 129 | 93 | 71 | 75 | 110 | 105 | 71 | 25 |
| GR 658 | n/a | n/a | 71 | 56 | 145 | 95 | 69 | 73 | 108 | 108 | 54 | 29 |
| GR 664 | n/a | n/a | 80 | 57 | 153 | 104 | 72 | 69 | 110 | 108 | 69 | 21 |
| GR 665 | n/a | n/a | 69 | 48 | 149 | 82 | 71 | 73 | 108 | 110 | 95 | 19 |
| Bolgar 159 | n/a | n/a | 85 | 59 | 127 | 95 | 44 | 45 | 81 | 82 | 68 | 26 |
| Kamusinszkoe 67 | n/a | n/a | 86 | 65 | 129 | 89 | 50 | 50 | 80 | 88 | 35 | 2 |
| Komsomolskoe 996 | n/a | n/a | 90 | 70 | 135 | 93 | 50 | 49 | 81 | 90 | 30 | 6 |
| Kazanskoe 176 | n/a | n/a | 92 | 59 | 124 | 72 | 45 | 44 | 79 | 90 | 40 | 4 |
| Tuvinskoe | n/a | n/a | 92 | 81 | 129 | 110 | 50 | 48 | 82 | 79 | 67 | 38 |
| Veszelopodoljanszkoe 403 | n/a | n/a | 84 | 63 | 134 | 92 | 50 | 49 | 81 | 90 | 100 | 6 |
| Unikum | n/a | n/a | 83 | 47 | 130 | 85 | 48 | 51 | 84 | 85 | 72 | 4 |
| Sunup | n/a | n/a | 90 | 68 | 132 | 106 | 54 | 52 | 88 | 98 | 61 | 5 |
| Earlybird | n/a | n/a | 79 | 60 | 128 | 94 | 58 | 52 | 91 | 91 | 49 | 12 |
| Huntsman | n/a | n/a | 75 | 53 | 127 | 83 | 66 | 62 | 95 | 101 | 53 | 11 |
| Sunrise | n/a | n/a | 79 | 60 | 119 | 91 | 59 | 56 | 91 | 93 | 31 | 5 |
| Turghai | n/a | n/a | 87 | 65 | 143 | 107 | 50 | 47 | 81 | 79 | 46 | 29 |
| Minsum | n/a | n/a | 85 | 62 | 140 | 98 | 53 | 53 | 85 | 86 | 76 | 50 |
| TU-85-074-03 | n/a | n/a | 84 | 64 | 134 | 96 | 56 | 55 | 91 | 84 | 26 | 45 |
| TU-85-087-01 | n/a | n/a | 86 | 66 | 136 | 91 | 52 | 51 | 91 | 85 | 25 | 9 |
| Mean | 83 | 61 | 134 | 94 | 56 | 55 | 90 | 92 | 55 | 18 | ||
| LSD (p < 0.05) | 2.5 | 5 | 4 | 8.1 | 11.5 | |||||||
| 2013 | ||||||||||||
| Dari | 43 | 31.7 | 95 | 22 | 141 | 15 | 60 | 70 | 86 | 98 | 164 | 9 |
| Tlicevskoje | 83.3 | 45 | 78 | 23 | 157 | 37 | 76 | n/a | 114 | 125 | 238 | 1 |
| GR 658 | 63.3 | 21.7 | 86 | 28 | 164 | n/a | 76 | 66 | 108 | n/a | 180 | n/a |
| GR 664 | 83.3 | 50 | 92 | 28 | 168 | 49 | 74 | 76 | 108 | 121 | 279 | 5 |
| GR 665 | 66.7 | 83.3 | 91 | 31 | 161 | 43 | 76 | n/a | 108 | 122 | 290 | 9 |
| Bolgar 159 | 48.3 | 28.3 | 82 | 27 | 121 | 29 | 60 | 60 | 83 | 82 | 89 | 1 |
| Kamusinszkoe 67 | 41.7 | 13.3 | 88 | 33 | 129 | 45 | 66 | 66 | 90 | 91 | 166 | 9 |
| Komsomolskoe 996 | 66.7 | 43.3 | 92 | 30 | 137 | 37 | 66 | 73 | 91 | 105 | 271 | 8 |
| Kazanskoe 176 | 60 | 45 | 92 | 34 | 125 | 32 | 60 | 60 | 83 | 83 | 184 | 2 |
| Tuvinskoe | 38.3 | 21.7 | 86 | 28 | 143 | 26 | 64 | 76 | 86 | 108 | 88 | 2 |
| Veszelopodoljanszkoe 403 | 66.7 | 50 | 92 | 32 | 136 | 37 | 64 | 73 | 91 | 99 | 198 | 3 |
| Unikum | 93.3 | 78.3 | 96 | 37 | 130 | 49 | 60 | 69 | 88 | 102 | 268 | 14 |
| Sunup | 28.3 | 30 | 71 | 22 | 151 | 33 | 74 | n/a | 108 | 117 | 344 | 1 |
| Earlybird | 40 | 30 | 83 | 24 | 146 | 36 | 72 | n/a | 108 | 117 | 365 | 8 |
| Huntsman | 11.7 | 15 | 64 | n/a | 140 | n/a | 76 | n/a | 108 | n/a | 155 | n/a |
| Sunrise | 35 | 25 | 79 | n/a | 155 | n/a | 74 | n/a | 108 | n/a | 336 | n/a |
| Turghai | 10 | 6.7 | 72 | n/a | 146 | n/a | 66 | n/a | 90 | n/a | 64 | n/a |
| Minsum | 30 | 53.3 | 79 | 27 | 152 | 35 | 69 | n/a | 101 | 117 | 227 | 3 |
| TU-85-074-03 | 26.7 | 31.7 | 78 | 31 | 124 | 38 | 66 | 76 | 90 | 98 | 129 | 3 |
| TU-85-087-01 | 21.7 | 8.3 | 93 | 33 | 121 | n/a | 64 | 66 | 87 | n/a | 97 | n/a |
| Mean | 48 | 35 | 84 | 29 | 142 | 36 | 68 | 69 | 97 | 106 | 206 | 5 |
| LSD (p < 0.05) | 25 | 3 | 8 | 4 | 3 | 43 | ||||||
| 2014 | ||||||||||||
| Dari | 48 | 12 | 90 | 19 | 96 | 32 | 49 | 49 | 92 | 93 | 5 | 2 |
| Tlicevskoje | 83 | n/a | 75 | n/a | 75 | n/a | 74 | n/a | 113 | n/a | 19 | n/a |
| GR 658 | 74 | 12 | 78 | 17 | 100 | 17 | 73 | 75 | 111 | 113 | 26 | n/a |
| GR 664 | 85 | 3 | 81 | n/a | 94 | n/a | 79 | 80 | 113 | 113 | 28 | n/a |
| GR 665 | 74 | 15 | 87 | 7 | 98 | 11 | 71 | 70 | 111 | 113 | 112 | n/a |
| Bolgar 159 | 56 | n/a | 69 | n/a | 69 | n/a | 45 | n/a | 94 | n/a | 11 | n/a |
| Kamusinszkoe 67 | 47 | n/a | 83 | n/a | 80 | n/a | 51 | n/a | 93 | n/a | 4 | n/a |
| Komsomolskoe 996 | 66 | 12 | 77 | 5 | 91 | 17 | 51 | 51 | 92 | 113 | 8 | 1 |
| Kazanskoe 176 | 73 | 8 | 92 | 28 | 91 | 31 | 48 | 48 | 86 | 103 | 6 | n/a |
| Tuvinskoe | 36 | 12 | 78 | 10 | 67 | 13 | 51 | n/a | 89 | 86 | n/a | n/a |
| Veszelopodoljanszkoe 403 | 65 | 18 | 90 | 19 | 74 | 16 | 51 | 51 | 93 | 97 | 12 | n/a |
| Unikum | 96 | 15 | 94 | 16 | 69 | 14 | 49 | 49 | 92 | 93 | 16 | n/a |
| Sunup | 40 | 18 | 73 | 9 | 99 | 24 | 54 | 54 | 94 | 93 | 19 | 12 |
| Earlybird | 49 | 14 | 82 | 10 | 86 | 25 | 55 | 51 | 96 | 93 | 24 | 4 |
| Huntsman | 26 | 9 | 66 | 10 | 90 | 9 | 70 | 70 | 113 | 113 | 26 | n/a |
| Sunrise | 45 | 9 | 75 | 5 | 96 | 7 | 65 | 65 | 113 | 113 | 39 | n/a |
| Turghai | 17 | 14 | 80 | 13 | 113 | 60 | 51 | 51 | 94 | 93 | 26 | 12 |
| Minsum | 30 | n/a | 78 | n/a | 109 | n/a | 54 | n/a | 93 | n/a | 34 | n/a |
| TU-85-074-03 | 48 | n/a | 75 | n/a | 47 | n/a | 54 | n/a | 96 | n/a | 4 | n/a |
| TU-85-087-01 | 42 | 15 | 79 | 16 | 62 | 18 | 55 | 55 | 86 | 86 | 7 | n/a |
| Mean | 55 | 12.4 | 80 | 13 | 85 | 21 | 57 | 58 | 98 | 101 | 22 | 6.2 |
| LSD (p < 0.05) | 12 | 3 | 8 | 1 | 3 | 7 | ||||||
| PH1 | PH2 | DH | DM | |
|---|---|---|---|---|
| PH1 | ||||
| PH2 | 0.86 *** | |||
| DH | −0.12 | 0.09 | ||
| DM | −0.37 *** | −0.23 * | 0.76 *** | |
| Yield | 0.45 *** | 0.65 *** | 0.42 *** | 0.11 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habiyaremye, C.; Barth, V.; Highet, K.; Coffey, T.; Murphy, K.M. Phenotypic Responses of Twenty Diverse Proso Millet (Panicum miliaceum L.) Accessions to Irrigation. Sustainability 2017, 9, 389. https://doi.org/10.3390/su9030389
Habiyaremye C, Barth V, Highet K, Coffey T, Murphy KM. Phenotypic Responses of Twenty Diverse Proso Millet (Panicum miliaceum L.) Accessions to Irrigation. Sustainability. 2017; 9(3):389. https://doi.org/10.3390/su9030389
Chicago/Turabian StyleHabiyaremye, Cedric, Victoria Barth, Kelsey Highet, Todd Coffey, and Kevin M. Murphy. 2017. "Phenotypic Responses of Twenty Diverse Proso Millet (Panicum miliaceum L.) Accessions to Irrigation" Sustainability 9, no. 3: 389. https://doi.org/10.3390/su9030389
APA StyleHabiyaremye, C., Barth, V., Highet, K., Coffey, T., & Murphy, K. M. (2017). Phenotypic Responses of Twenty Diverse Proso Millet (Panicum miliaceum L.) Accessions to Irrigation. Sustainability, 9(3), 389. https://doi.org/10.3390/su9030389