
Effect of Agricultural Management Intensity on the Organic Carbon Fractions and Biological Properties of a Volcanic-Ash-Derived Soil

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site Description and Sampling
2.2. Soil Measurements
2.2.1. Soil Organic Carbon (SOC), Soluble Carbon (OC-sol), and Active Carbon (POXC)
2.2.2. Cellulase Activity and Autoclaved-Citrate-Extractable Protein
2.3. Earthworm Biomass and Density
2.4. Water Stability of Aggregates (WSA)
2.5. Statistical Analysis
3. Results
3.1. Changes in SOC, Soluble OC, and POXC Content According to Management Intensity
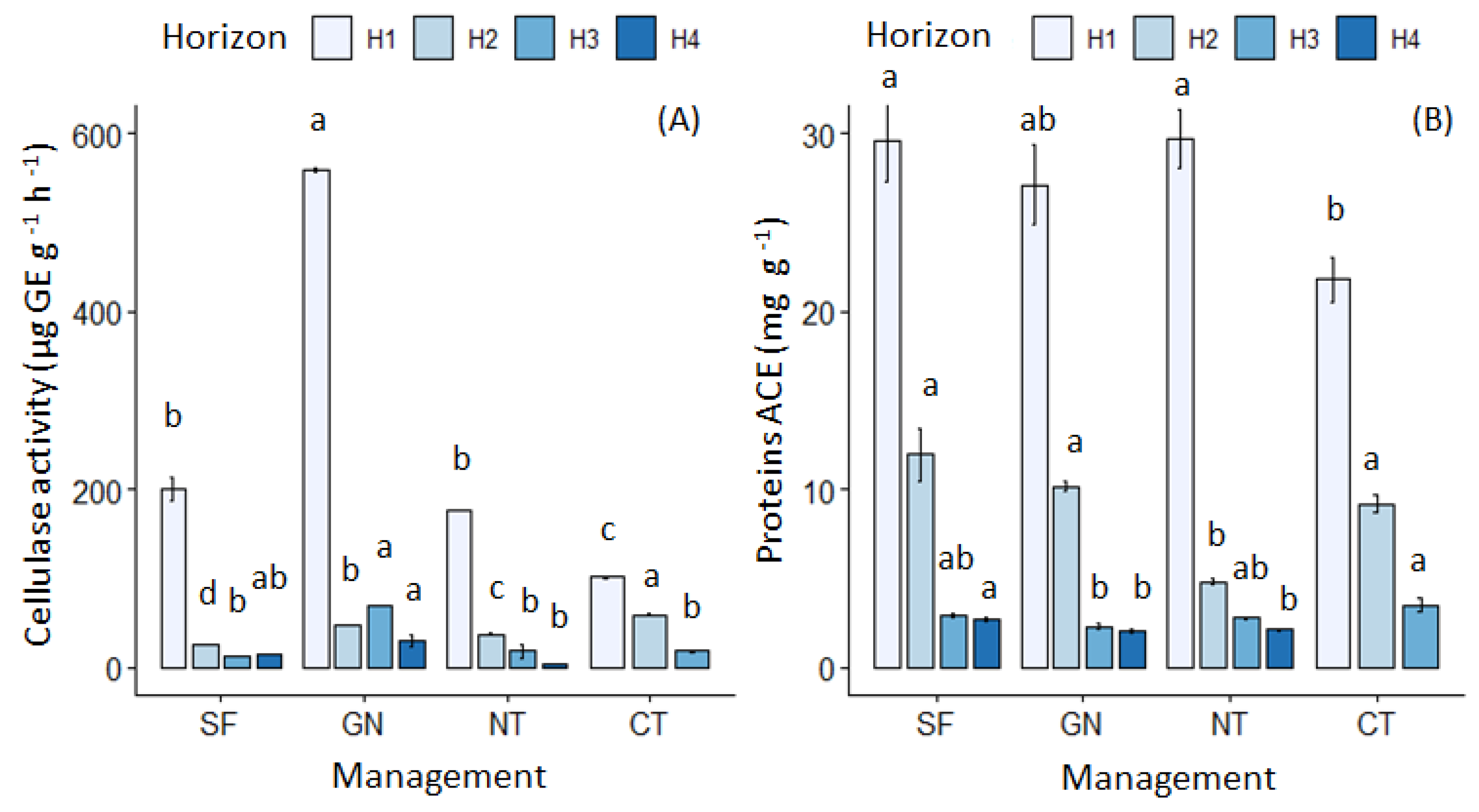
3.2. Changes in Biological Indicators According to Management Intensity
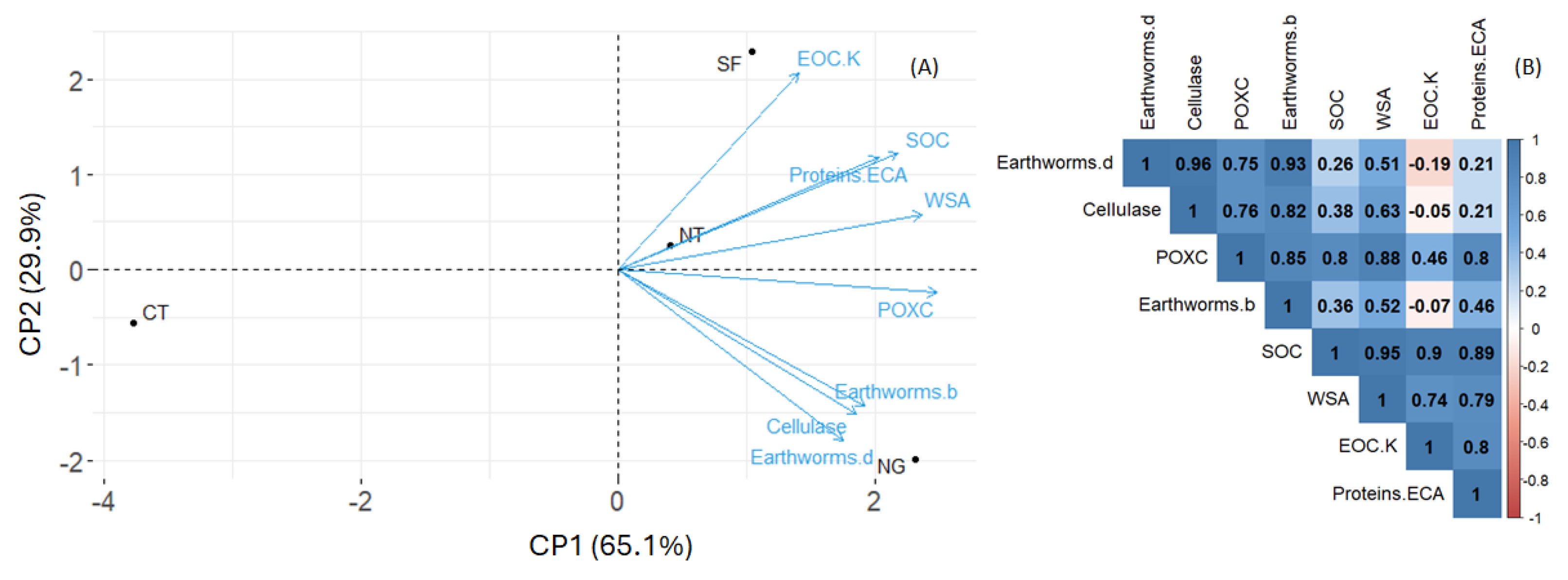
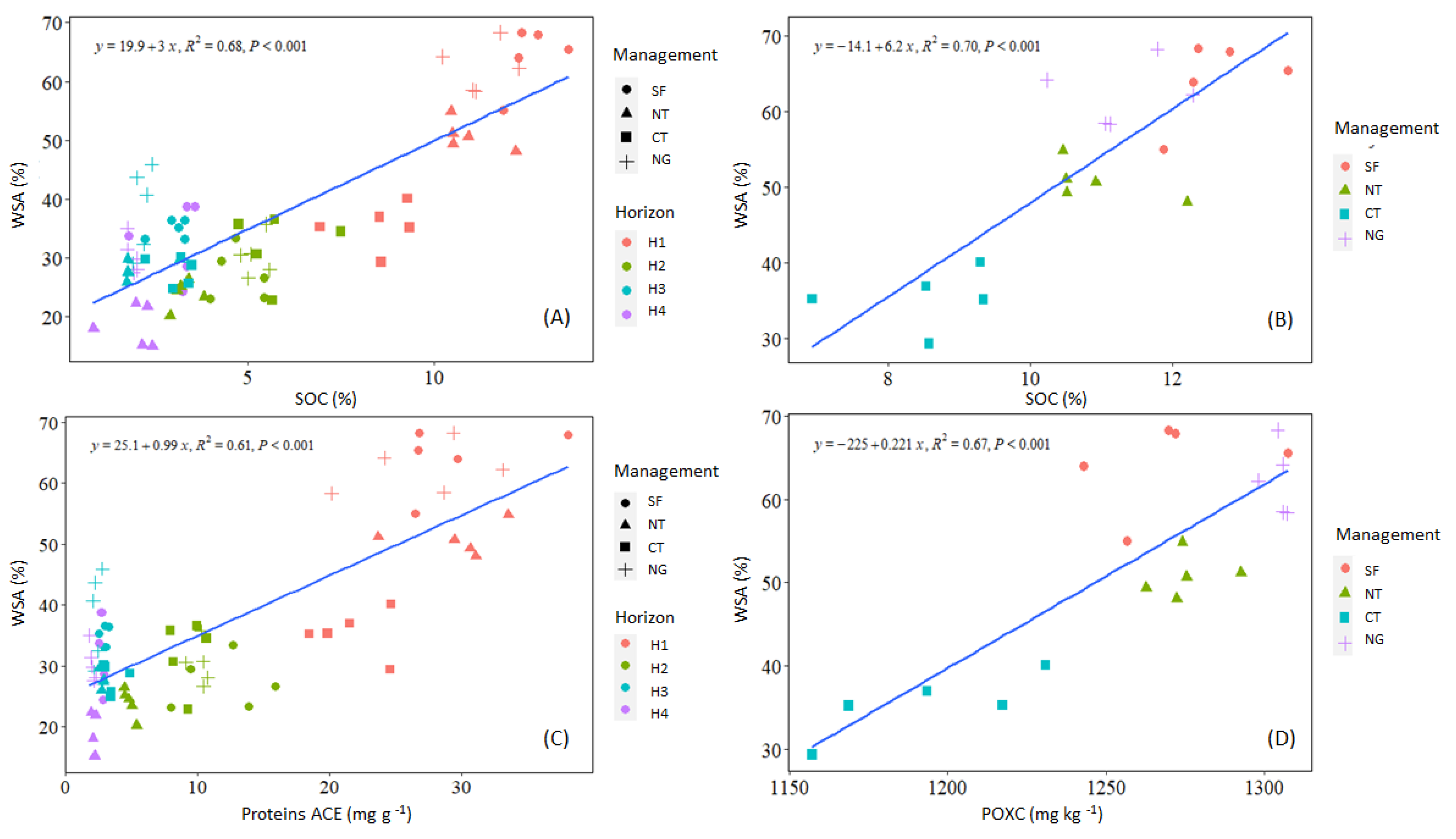
3.3. Relation Between Structural Stability, Organic Carbon Content, and Biological Indicators
4. Discussion
4.1. Changes in SOC Content and Labile Fractions According to Soil Management Intensity
4.2. Influence of Biological Indicators on C Content Under Different Soil Management Intensities
4.3. Influence of Aggregate Stability on SOC Persistence and Labile C Fractions
4.4. Importance of SOC Storage and Protection in Volcanic-Ash-Derived Soils
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lal, R.; Negassa, W.; Lorenz, K. Carbon sequestration in soil. Curr. Opin. Environ. Sustain. 2015, 15, 79–86. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Urbanski, L.; Hobley, E.; Lang, B.; von Lützow, M.; Marin-Spiotta, E.; van Wesemael, B.; Rabot, E.; Ließ, M.; Garcia-Franco, N.; et al. Soil organic carbon storage as a key function of soils-A review of drivers and indicators at various scales. Geoderma 2019, 333, 149–162. [Google Scholar] [CrossRef]
- Smith, P.; Soussana, J.F.; Angers, D.; Schipper, L.; Chenu, C.; Rasse, D.P.; Batjes, N.H.; Van Egmond, F.; McNeill, S.; Kuhnert, M.; et al. How to measure, report and verify soil carbon change to realize the potential of soil carbon sequestration for atmospheric greenhouse gas removal. Glob. Change Biol. 2020, 26, 219–241. [Google Scholar] [CrossRef]
- FAO. Carbono Orgánico del Suelo: El Potencial Oculto; Organización de las Naciones Unidas para la Alimentación y Agricultura: Roma, Italy, 2017; 90p. [Google Scholar]
- Cotrufo, M.F.; Lavallee, J.M. Soil organic matter formation, persistence, and functioning: A synthesis of current understanding to inform its conservation and regeneration. Adv. Agron. 2022, 172, 1–66. [Google Scholar] [CrossRef]
- Sanderman, J.; Hengl, T.; Fiske, G.J. Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. USA 2017, 114, 9575–9580. [Google Scholar] [CrossRef] [PubMed]
- Dignac, M.F.; Derrien, D.; Barré, P.; Barot, S.; Cécillon, L.; Chenu, C.; Chevallier, T.; Freschet, G.T.; Garnier, P.; Guenet, B.; et al. Increasing soil carbon storage: Mechanisms, effects of agricultural practices and proxies. A review. Agron. Sustain. Dev. 2017, 37, 14. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Soong, J.L.; Cotrufo, M.F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Change Biol. 2020, 26, 261–273. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Zamanian, K. Reviews and synthesis: Agropedogenesis-humankind as the sixth soil-forming factor and attractors of agricultural soil degradation. Biogeosciences 2019, 16, 4783–4803. [Google Scholar] [CrossRef]
- Lehman, J.; Bossio, D.; Kogel-Knabner, I.; Rillig, M. The concept and prospects of soil health. Nat. Rev. Earth Environ. 2020, 10, 544–553. [Google Scholar] [CrossRef]
- Matus, F.J.; Paz-Pellat, F.; Covaleda, S.; Etchevers, J.D.; Hidalgo, C.; Báez, A. Upper limit of mineral-associated organic carbon in temperate and sub-tropical soils: How far is it? Geoderma Reg. 2024, 37, e00811. [Google Scholar] [CrossRef]
- Angst, G.; Mueller, K.; Kögel-Knabner, I.; Freeman, K.; Mueller, C. Aggregation controls the stability of lignin and lipids in clay-sized particulate and mineral associated organic matter. Biogeochemistry 2017, 132, 307–324. [Google Scholar] [CrossRef]
- Chan, K.Y. An overview of some tillage impacts on earthworm population abundance and diversity-implications for functioning in soil. Soil Tillage Res. 2001, 57, 179–191. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Haddix, M.L.; Gregorich, E.G.; Helgason, B.L.; Janzen, H.; Ellert, B.H.; Cotrufo, M.F. Climate, carbon content, and soil texture control the independent formation and persistence of particulate and mineral-associated organic matter in soil. Geoderma 2020, 363, 114160. [Google Scholar] [CrossRef]
- Six, J.; Elliot, E.T.; Paustian, K. Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture. Soil Biol. Biochem. 2000, 32, 2099–2103. [Google Scholar] [CrossRef]
- Kan, Z.; Liu, W.X.; Liu, W.S.; Lal, R.; Dang, Y.; Zhao, X.; Zhang, H. Mechanisms of soil organic carbon stability and its response to no-till: A global synthesis and perspective. Glob. Change Biol. 2021, 8, 693–710. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Feng, X.; Yang, H.; Li, Y.; Kuzyakov, Y.; Liu, S.; Li, F. Effects of tillage on soil organic carbon and crop yield under straw return. Agric. Ecosyst. Environ. 2023, 354, 108543. [Google Scholar] [CrossRef]
- Brookes, P.; Chen, Y.; Chen, L.; Qiu, G.; Luo, Y.; Xu, J. Is the rate of mineralization of soil organic carbon under microbiological control? Soil Biol. Biochem. 2017, 112, 127–139. [Google Scholar] [CrossRef]
- Yudina, A.; Kuzyakov, Y. Dual nature of soil structure: The unity of aggregates and pores. Geoderma 2023, 434, 116478. [Google Scholar] [CrossRef]
- Panettieri, M.; Berns, A.E.; Knicker, H.; Murillo, J.M.; Madejón, E. Evaluation of seasonal variability of soil biogeochemical properties in aggregate-size fractioned soil under different tillages. Soil Tillage Res. 2015, 151, 39–49. [Google Scholar] [CrossRef]
- Bai, Y.; Cotrufo, M.F. Grassland soil carbon sequestration: Current understanding, challenges, and solutions. Science 2022, 377, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, E.; Silva, P. Agronomía de la Cero Labranza; Universidad de Chile: Santiago, Chile, 2003; Facultad de Ciencias Agronómicas. Serie Ciencias Agronómicas Nº 10; 118p, ISBN 956-19-0363-6. [Google Scholar]
- Bai, Z.; Caspari, T.; Ruiperez Gonzalez, M.; Batjes, N.H.; Mäder, P.; Bünemann, E.K.; de Goede, R.; Brussaard, L.; Xu, M.; Santos Ferreira, C.S.; et al. Effects of agricultural management practices on soil quality: A review of long-term experiments for Europe and China. Agric. Ecosyst. Environ. 2018, 265, 1–7. [Google Scholar] [CrossRef]
- Bongiorno, G.; Bünemann, E.; Oguejiofor, C.; Meier, J.; Gort, G.; Comans, R.; Mäder, P.; Brussard, L.; De Goede, R. Sensivity of labile carbon fractions to tillage and organic matter management and their potencial as comprehensive soil quality indicators across pedoclimatic conditions in Europe. Ecol. Indic. 2019, 99, 38–50. [Google Scholar] [CrossRef]
- Ghani, A.; Dexter, M.; Perrott, K.W. Hot-water extractable carbon in soils: A sensitive measurement for determining impacts of fertilization, grazing and cultivation. Soil Boil. Biochem. 2003, 35, 1231–1243. [Google Scholar] [CrossRef]
- Jangid, K.; Williams, M.; Franzluebbers, A.; Sanderlin, J.; Reeves, J.; Jenkins, M.; Endale, D.; Coleman, D.; Whitman, W. Relative impacts of land-use, management intensity and fertilization upon soil microbial community structure in agriculture systems. Soil Biol. Biochem. 2008, 40, 2843–2853. [Google Scholar] [CrossRef]
- Valle, S.R.; Carrasco, J. Soil quality indicator selection in Chilean volcanic soils formed under temperate and humid conditions. Catena 2018, 162, 386–395. [Google Scholar] [CrossRef]
- Zúñiga, F.; Dec, D.; Valle, S.; Thiers, O.; Paulino, L.; Martínez, O.; Seguel, O.; Casanova, M.; Pino, M.; Horn, R.; et al. The waterlogged volcanic ash soils of southern Chile. A review of the “Ñadi” soils. Catena 2019, 173, 99–113. [Google Scholar] [CrossRef]
- Bachman, J.; Goebel, M.O.; Krueger, J.; Fleige, H.; Woche, S.; Dörner, J.; Horn, R. Aggregate stability of south Chilean volcanic ash soils—A combined XPS, contact angle, and surface charge analysis. Geoderma 2020, 361, 114022. [Google Scholar] [CrossRef]
- Matus, F.; Rumpel, C.; Neculman, R.; Panichini, M.; Mora, M.L. Soil carbon storage and stabilisation in andic soils: A review. Catena 2014, 120, 102–110. [Google Scholar] [CrossRef]
- Mera, M.; Lizana, X.C.; Calderini, D.F. Chapter 6—Cropping systems in environments with high yield potential of southern Chile. In Crop Physiology, 2nd ed.; Sadras, V.O., Calderini, D.F., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 111–140. ISBN 9780124171046. [Google Scholar] [CrossRef]
- CIREN. Descripciones de Suelos y Materiales y Símbolos. Estudio Agrológico de la X Región; Publicación 123; Centro de Información de Recursos Naturales (CIREN): Santiago, Chile, 2003. [Google Scholar]
- Luzio, W.; Casanova, M.; Seguel, O. Suelos de Chile; Universidad de Chile: Santiago, Chile, 2010. [Google Scholar]
- Sadzawka, A.; Carrasco, M.; Grez, R.; Mora, M.; Flores, H.; Neaman, A. Métodos de Análisis Recomendados para los Suelos de Chile; Serie actas N°34; Instituto de Investigaciones Agropecuarias (INIA): Santiago, Chile, 2006; 164p. [Google Scholar]
- Haney, R.L.; Haney, L.B.; Smith, D.R.; Harmel, R.D.; White, M.J. The soil health tool—Theory and initial broad-scale application. Appl. Soil Ecol. 2018, 125, 162–168. [Google Scholar] [CrossRef]
- Weil, R.; Islam, K.; Stine, M.; Gruver, J.; Samson-Liebig, S. Estimating active carbon for soil quality assessment: A simplified method for laboratory and field use. Am. J. Altern. Agric. 2003, 18, 3–17. [Google Scholar] [CrossRef]
- Shinner, F.; Von Mersi, W. Xylanase, CM-cellulase and invertase activity in soil: An improved method. Soil Biol. Biochem. 1990, 22, 511–515. [Google Scholar] [CrossRef]
- Dick, R.; Burns, R. A Brief History of Soil Enzymology Research. In Methods of Soil Enzymology; Dick, R., Ed.; Soil Science Society of America: Madison, WI, USA, 2011; pp. 1–34. [Google Scholar] [CrossRef]
- Wright, S.; Upadhyaya, A. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci. 1996, 161, 575–586. [Google Scholar] [CrossRef]
- Clune, D.J. Glomalin: Its Relationship with Aggregate Stability, Response to Soil Management, Source and Quantification. Master’s Thesis, Cornell University, Ithaca, NY, USA, 2007. [Google Scholar]
- Moebius-Clune, B.N.; Moebius-Clune, D.J.; Gugino, B.; Idowu, O.; Schinedelbeck, R.; Ristow, A.; van Es, H.; Thies, J.; Shayler, H.; McBride, M.; et al. Comprehensive Assesment of Soil Health—The Cornell Framework, 3rd ed.; Cornell University: Geneva, Switzerland; New York, NY, USA, 2016. [Google Scholar]
- Bracamontes, L.; Fuentes, M.; Rodríguez, L.; Macedas, J. Manual de Indicadores Biológicos de la Salud del Suelo; Universidad Autónoma Metropolitana (UAM): Ciudad de México, Mexico, 2018; 93p. [Google Scholar]
- Edwards, C.; Bohlen, P. Biology and Ecology of Earthworms, 3rd ed.; Chapman & Hall: London, UK, 1996. [Google Scholar]
- Barrón, C.; Santelices, M.; Clunes, J.; Pinochet, D. Does earthworm density change the quality of degraded volcanic soil? Appl. Soil Ecol. 2024, 193, 105154. [Google Scholar] [CrossRef]
- Kemper, W.; Rosenau, R. Aggregate Stability and Size Distribution. In Methods of Soil Analysis, Part 1: Physical and Mineralogical Methods, 2nd ed.; Karaca, A., Ed.; Monography Agronomy: Madison, WI, USA, 1986; pp. 415–442. [Google Scholar]
- Luo, Z.; Wang, E.; Sun, O.J. Can no-tillage stimulate carbon sequestration in agricultural soils? A meta analysis of paired experiments. Agric. Ecosyst. Environ. 2010, 139, 224–231. [Google Scholar] [CrossRef]
- Begum, R.; Jahangir, M.; Jahiruddin, M.; Islam, M.; Bokhtiar, S.; Islam, K. Reduce tillage with residue retentiton improves soil labile carbon pools and carbon lability and management indices in a seven-year trial with wheat-mung bean-rice rotation. Pedosphere 2022, 32, 916–927. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Wang, J.; You, Y.; Yang, Y.; Shi, Z.; Huang, X.; Zheng, L.; Li, Z.; Ming, A.; et al. Mixed-species plantation with Pinus massoniana and Castanopsis hystrix accelerates C loss in recalcitrant coniferous litter but slows C loss in labile broadleaf litter in southern China. For. Ecol. Manag. 2018, 422, 207–213. [Google Scholar] [CrossRef]
- Yuan, Z.; Jin, X.; Xiao, W.; Wang, L.; Sun, Y.; Guan, Q.; Ofori, A. Comparing soil organic carbon stock and fractions under natural secondary forest and Pinus massoniana plantation in subtropical China. Catena 2022, 212, 106092. [Google Scholar] [CrossRef]
- Conant, R.; Paustian, K. Potential soil carbon sequestration in overgrazed grassland ecosystems. Glob. Biogeochem. Cycles 2002, 16, 1143. [Google Scholar] [CrossRef]
- Lugato, E.; Lavallee, J.M.; Haddix, M.L.; Panagos, P.; Cotrufo, M.F. Different climate sensitivity of particulate and mineral-associated soil organic matter. Nat. Geosci. 2021, 14, 295–300. [Google Scholar] [CrossRef]
- Rumpel, C.; Kögel-Knabner, I. Deep soil organic matter—A key but poorly understood component of terrestrial C cycle. Plant Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Montesdeoca, F.; Ávila, M.; Quisphe, J.; Borie, F.; Cornejo, P.; Aguilera, P.; Alvarado, S.; Espinosa, J. Early changes in the transition from conventional to no-tillage in a volcanic cultivated with beans (Phaseolus vulgaris L.). Chil. J. Agric. Anim. Sci. 2020, 36, 181–189. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration to mitigate climate change. Geoderma 2004, 123, 1–22. [Google Scholar] [CrossRef]
- Jat, H.; Datta, A.; Choundhary, M.; Shrama, P.; Dixit, B.; Jat, M. Soil enzymes activity: Effect of climate smart agriculture on rhizosphere and bulk soil under cereal based systems of north-west India. Eur. J. Soil Biol. 2021, 103, 103–292. [Google Scholar] [CrossRef]
- Mirzavand, J.; Asadi-Rahmani, H.; Moradi-Talebbeigi, R. Biological indicators of soil quality under conventional, reduced, and no-tillage systems. Arch. Agron. Soil Sci. 2022, 68, 311–324. [Google Scholar] [CrossRef]
- Zimmer, J.; Poggere, G.; Studart, R.; Hungria, M.; Carvalho, I. Soil enzymatic activity in Brazilian biomes under native vegetation and contrasting cropping and management. Appl. Soil Ecol. 2023, 190, 105014. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Zobeck, T.M.; Gill, T.E.; Kennedy, A.C. Enzyme activities and microbial community structure in semiarid agricultural soils. Biol. Fertil. Soils 2003, 38, 216–227. [Google Scholar] [CrossRef]
- Alvear, M.; Pino, M.; Castillo, C.; Trasar Ce peda, C.; Gil Sotres, F. Efecto de la cero labranza sobre algunas actividades biológicas en un Alfisol del sur de Chile. Rev. Cienc. Suelo Nutr. Veg. 2006, 6, 38–53. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; Burns, R.G. Ecology of extracellular enzyme activities and organic matter degradation in soil: A complex community-driven process. In Methods of Soil Enzymology; Dick, R.P., Ed.; Soil Science Society of America: Madison, WI, USA, 2011; pp. 35–56. [Google Scholar]
- Wu, L. Sequestering organic carbon in soils through land use change and agricultural practices: A review. Front. Agric. Sci. Eng. 2023, 10, 210–225. [Google Scholar] [CrossRef]
- Li, J.Y.; Chen, P.; Li, Z.G.; Li, L.Y.; Zhang, R.Q.; Hu, W.; Liu, Y. Soil aggregate-associated organic carbon mineralization and its driving factors in rhizosphere soil. Soil Biol. Biochem. 2023, 186, 109182. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.; Marianari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Mencel, J.; Mocek-Płóciniak, A.; Kryszak, A. Soil Microbial Community and Enzymatic Activity of Grasslands under Different Use Practices: A Review. Agronomy 2022, 12, 1136. [Google Scholar] [CrossRef]
- Xu, H.; You, C.; Tan, B.; Xu, L.; Liu, Y.; Wang, M.; Xu, Z.; Sardans, J.; Peñuelas, J. Effects of livestock grazing on the relationships between soil microbial community and soil carbon in grassland ecosystems. Sci. Total Environ. 2023, 881, 163416. [Google Scholar] [CrossRef]
- Blum, W.E.H. Functions of Soil for Society and the Environment. Rev. Environ. Sci. Biotechnol. 2005, 4, 75–79. [Google Scholar] [CrossRef]
- DeLuca, T.H.; Pingree, M.R.A.; Gao, S. Assessing soil biological health in forest soils. In Developments in Soil Science; Elsevier: Amsterdam, The Netherlands, 2019; Volume 36, pp. 397–426. [Google Scholar] [CrossRef]
- Huang, B.; Yan, G.; Liu, G.; Sun, X.; Wang, X.; Xing, Y.; Wang, Q. Effects of long-term nitrogen addition and precipitation reduction on glomalin-related soil protein and soil aggregate stability in a temperate forest. Catena 2022, 214, 106284. [Google Scholar] [CrossRef]
- Situ, G.; Zhao, Y.; Zhang, L.; Yang, X.; Chen, D.; Li, S.; Wu, Q.; Xu, Q.; Chen, J.; Qin, H. Linking the chemical nature of soil organic carbon and biological binding agent in aggregates to soil aggregate stability following biochar amendment in a rice paddy. Sci. Total Environ. 2022, 847, 157460. [Google Scholar] [CrossRef]
- Borie, F.; Rubio, R.; Rouanet, J.; Morales, A.; Borie, G.; Rojas, C. Effects of tillage systems on soil characteristics, glomalin and mycorrhizal propagules in a Chilean Ultisol. Soil Tillage Res. 2006, 88, 253–261. [Google Scholar] [CrossRef]
- Kavdir, Y.; Ílay, R. Earthworms and Soil Structure. Chapter 3. In Biology of Earthworms; Karaca, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 39–50. [Google Scholar]
- Barthod, J.; Dignac, M.; Rumpel, C. Effect of descomposition products produced in the presence or absence of epigeic earthworms and minerals on soil carbon stabilization. Soil Biol. Biochem. 2021, 160, 108308. [Google Scholar] [CrossRef]
- Bossuyt, H.; Six, J.; Hendrix, P. Protection of soil carbon by microaggregates within earthworm casts. Soil Biol. Biochem. 2005, 37, 251–258. [Google Scholar] [CrossRef]
- Bartz, M.L.C.; Brown, G.G.; da Rosa, M.G.; Klauberg Filho, O.; James, S.W.; Decaëns, T.; Baretta, D. Earthworm richness in land-use systems in Santa Catarina, Brazil. Appl. Soil Ecol. 2014, 83, 59–70. [Google Scholar] [CrossRef]
- Stellin, F.; Gavinelli, F.; Stevanato, P.; Concheri, G.; Squartini, A.; Paoletti, M. Effects of different concentrations of glyphosate (Roundup 360) on earthworms (Octodrilus complanatus, Lumbricus terrestris and Aporrectodea caliginosa) in veneyards in the North-East of Italy. Appl. Soil Ecol. 2018, 123, 802–808. [Google Scholar] [CrossRef]
- Stevenson, B.A.; Sarmah, A.K.; Smernik, R.; Hunter, D.W.F.; Fraser, S. Soil carbon characterization and nutrient ratios across land uses on two contrasting soils: Their relationships to microbial biomass and function. Soil Biol. Biochem. 2016, 97, 50–62. [Google Scholar] [CrossRef]
- Stamati, F.; Nikolaidis, N.; Banwart, S.; Blum, W. A coupled carbon, aggregation, and structure turnover (CAST) model for topsoils. Geoderma 2013, 211–212, 51–64. [Google Scholar] [CrossRef]
- Vera-Peters, V.; Zúñiga, F.; Valle, S.R.; Dec, D.; Clunes, J.; Dörner, J. Shrinkage behavior of Aquands along a longitudinal climatic gradient in Southern Chile. J. Soil Sci. Plant Nutr. 2022, 23, 638–650. [Google Scholar] [CrossRef]
- Lal, R. Soil Health and carbon management. Food Energy Secur. 2016, 5, 212–222. [Google Scholar] [CrossRef]
- Gong, N.; Hong, H.; Huff, W.D.; Fang, Q.; Bae, C.J.; Wang, C.; Yin, K.; Chen, S. Influences of Sedimentary Environments and Volcanic Sources on Diagenetic Alteration of Volcanic Tuffs in South China. Sci. Rep. 2018, 8, 7616. [Google Scholar] [CrossRef] [PubMed]
- ODEPA. Panorama de la Agricultura Chilena; ODEPA: Bhubaneswar, India, 2019; 152p, ISBN 978-956-7244-31-7. [Google Scholar]
- Moscoso, C.; Urrutia, N. Overview of the Forage Land-Use in Southern Chile in a Thirty-Year Period. Crop Forage Turfgrass Manag. 2017, 3, 1–4. [Google Scholar] [CrossRef]
- Ovalle, C. Un Nama Agrícola para Chile Mediante el Secuestro de Carbono Orgánico en el Suelo; Serie Actas INIA N°59; Instituto de Investigaciones Agropecuarias, Centro Regional de Investigación La Cruz, Región de Valparaíso: Santiago, Chile, 2020; 94p. [Google Scholar]
- Valle, S.R.; Dörner, J.; Zúñiga, F.; Dec, D. Seasonal dynamics of the physical quality of volcanic ash soils under different land uses in southern Chile. Soil Tillage Res. 2018, 182, 25–34. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Use | Physical Intervention | Vegetation | Residues Inputs | Time |
|---|---|---|---|---|
| CT | Plowing (0–20 cm) | Wheat monoculture | A portion of the residues is incorporated | 20 years |
| NT | Without plowing (last 2 years, surface movement: 2 cm) | Wheat–vetch–rape rotations, sometimes incorporating peas | Residues are kept on the ground | >25 years |
| NG | Grazing (high loads in short periods) | Grasses (Agrostis capillaris L., Arrhenatherum elatius spp., Holcus lanatus L., Hypochaeris radicata). Legumes (Trifolium pratense and Locus pedunculatus) | Maintenance of vegetation cover and grazing | 20 years |
| SF | No intervention | Maqui (Aristotelia chilensis), Arrayán (Luma apiculata), Roble (Nothofagus obliqua), Quila (Chusquea quila), Ulmo (Eucryphia cordifolia) | Maintenance of vegetation cover | >20 years |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aravena, C.; Valle, S.R.; Vergara, R.; González Chang, M.; Martínez, O.; Clunes, J.; Caurapán, B.; Asenjo, J. Effect of Agricultural Management Intensity on the Organic Carbon Fractions and Biological Properties of a Volcanic-Ash-Derived Soil. Sustainability 2025, 17, 2704. https://doi.org/10.3390/su17062704
Aravena C, Valle SR, Vergara R, González Chang M, Martínez O, Clunes J, Caurapán B, Asenjo J. Effect of Agricultural Management Intensity on the Organic Carbon Fractions and Biological Properties of a Volcanic-Ash-Derived Soil. Sustainability. 2025; 17(6):2704. https://doi.org/10.3390/su17062704
Chicago/Turabian StyleAravena, Camila, Susana R. Valle, Rodrigo Vergara, Mauricio González Chang, Oscar Martínez, John Clunes, Belén Caurapán, and Joel Asenjo. 2025. "Effect of Agricultural Management Intensity on the Organic Carbon Fractions and Biological Properties of a Volcanic-Ash-Derived Soil" Sustainability 17, no. 6: 2704. https://doi.org/10.3390/su17062704
APA StyleAravena, C., Valle, S. R., Vergara, R., González Chang, M., Martínez, O., Clunes, J., Caurapán, B., & Asenjo, J. (2025). Effect of Agricultural Management Intensity on the Organic Carbon Fractions and Biological Properties of a Volcanic-Ash-Derived Soil. Sustainability, 17(6), 2704. https://doi.org/10.3390/su17062704