
Corridors of Suitable Distribution of Betula platyphylla Sukaczev Forest in China Under Climate Warming


 ,
,
Abstract
1. Introduction
2. Materials and Methods
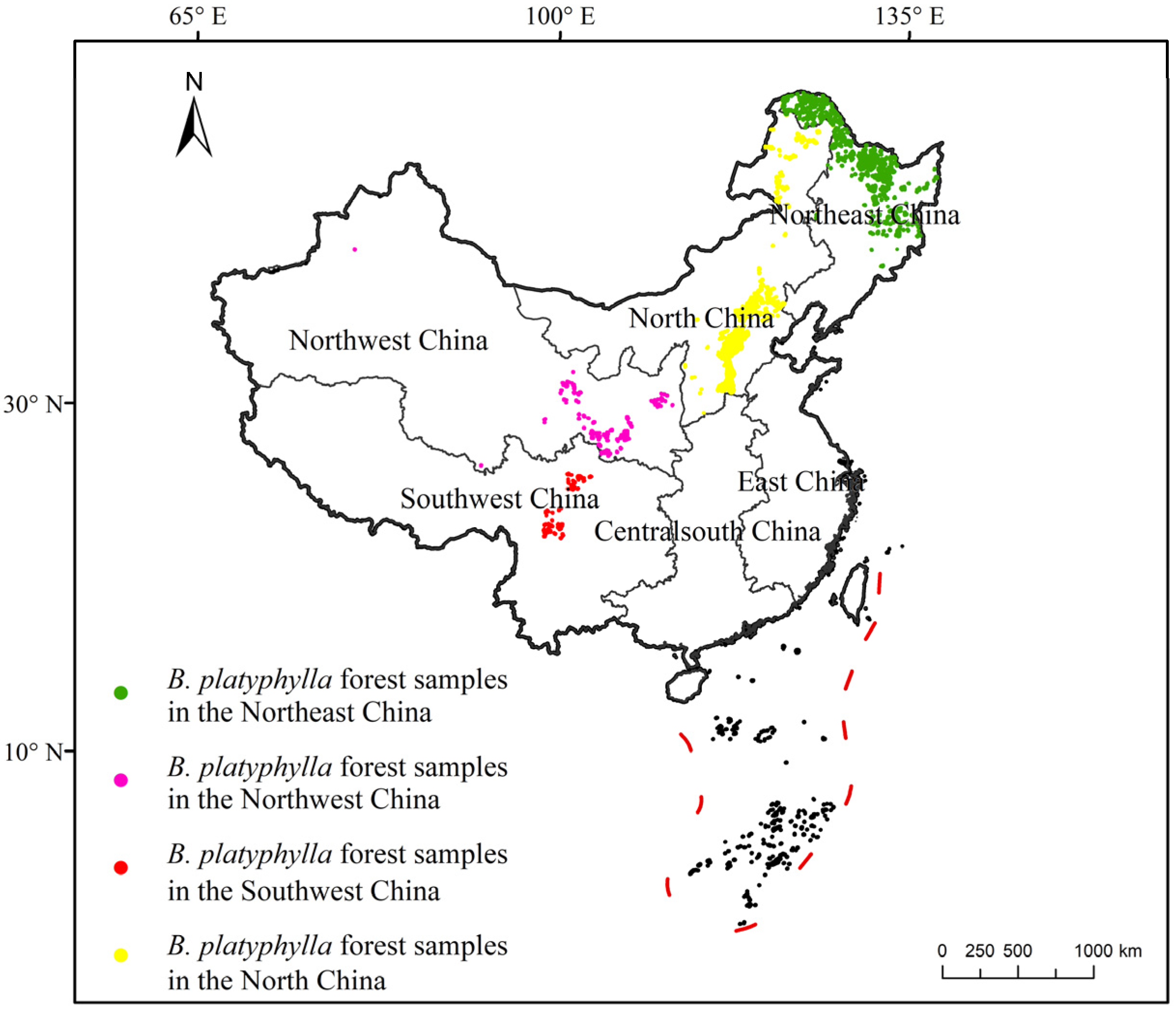
2.1. Data Sources and Data Processing
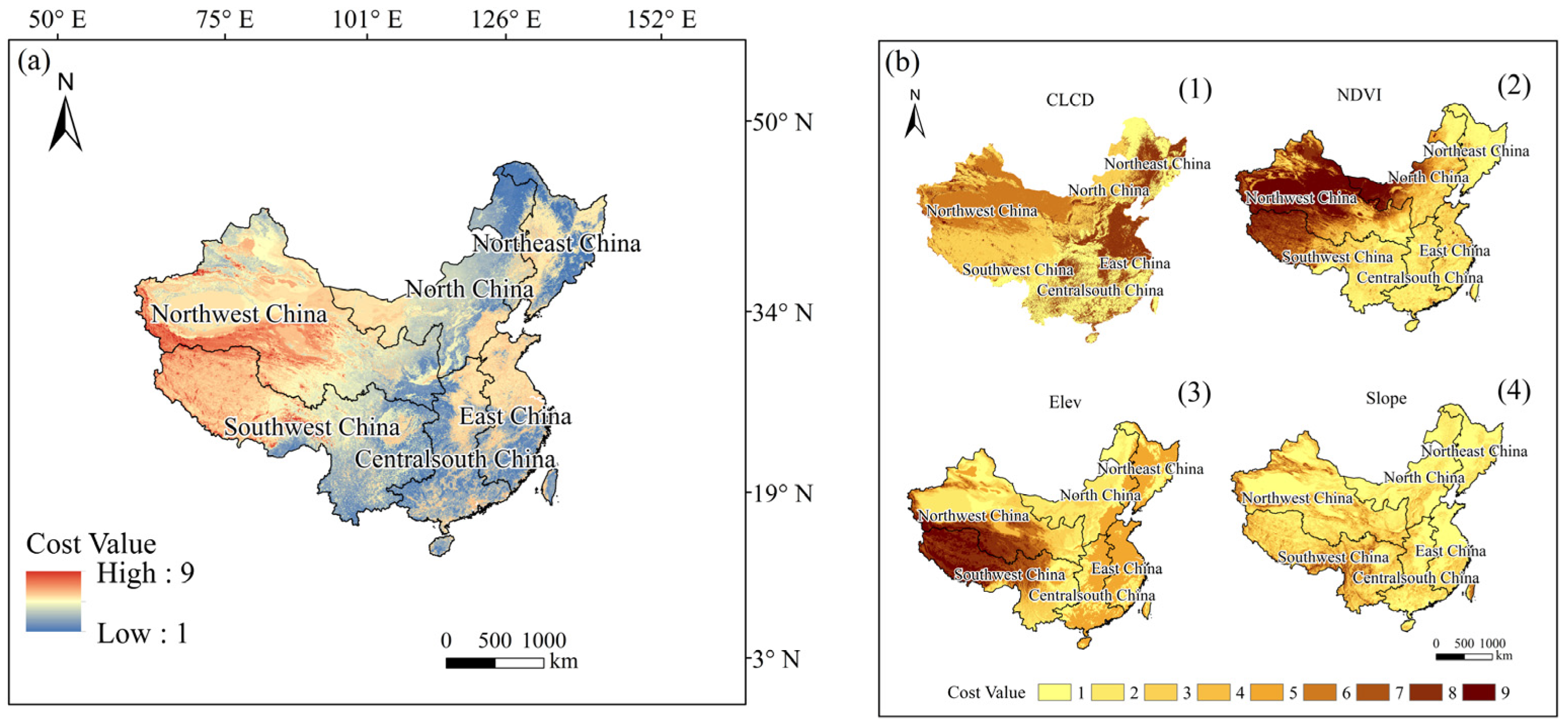
2.2. Calculation of Suitable Areas and Recognize of Driving Variables
2.3. Construct of Ecological Corridors
3. Results
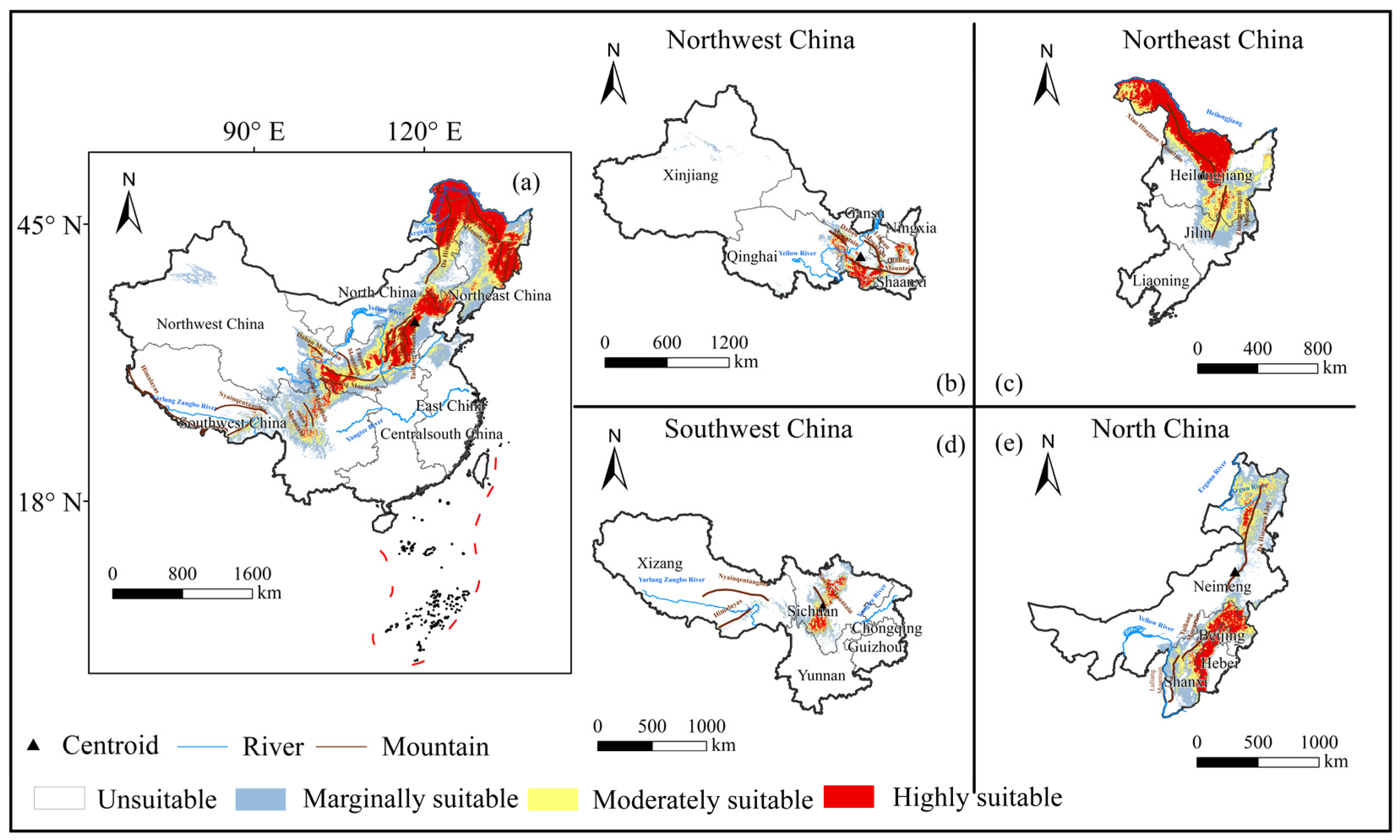
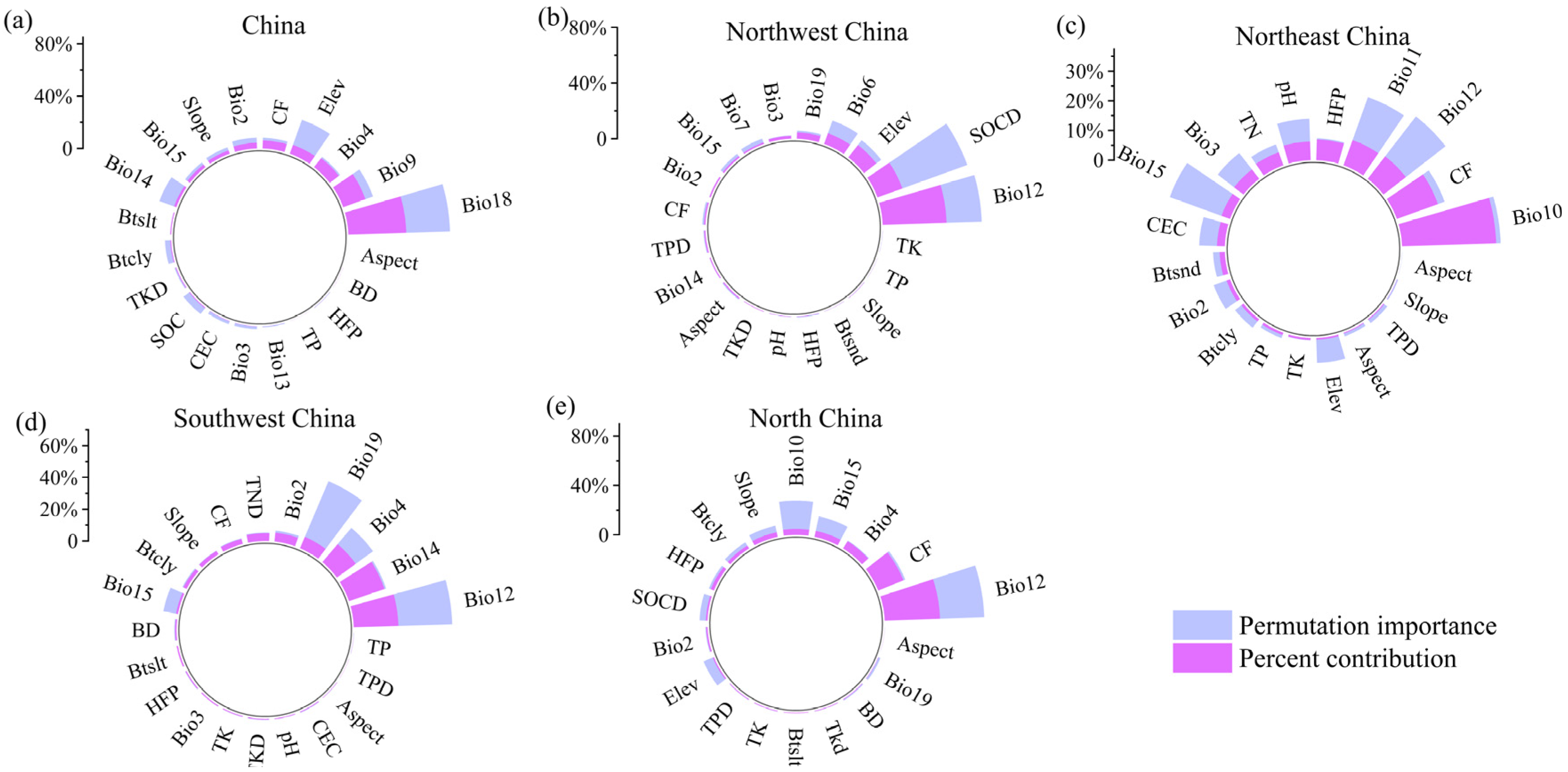
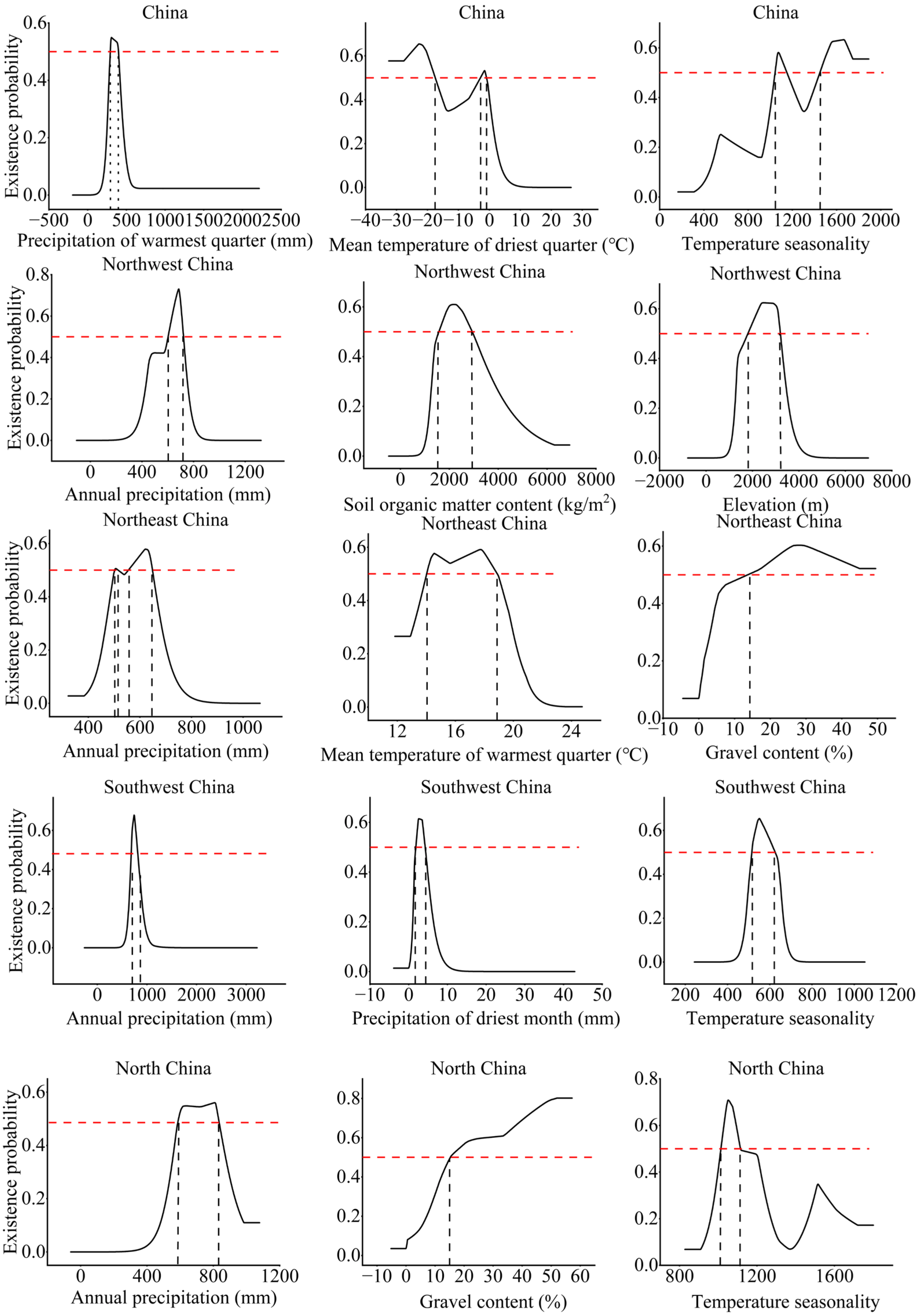
3.1. Suitable Distribution and Driving Factors
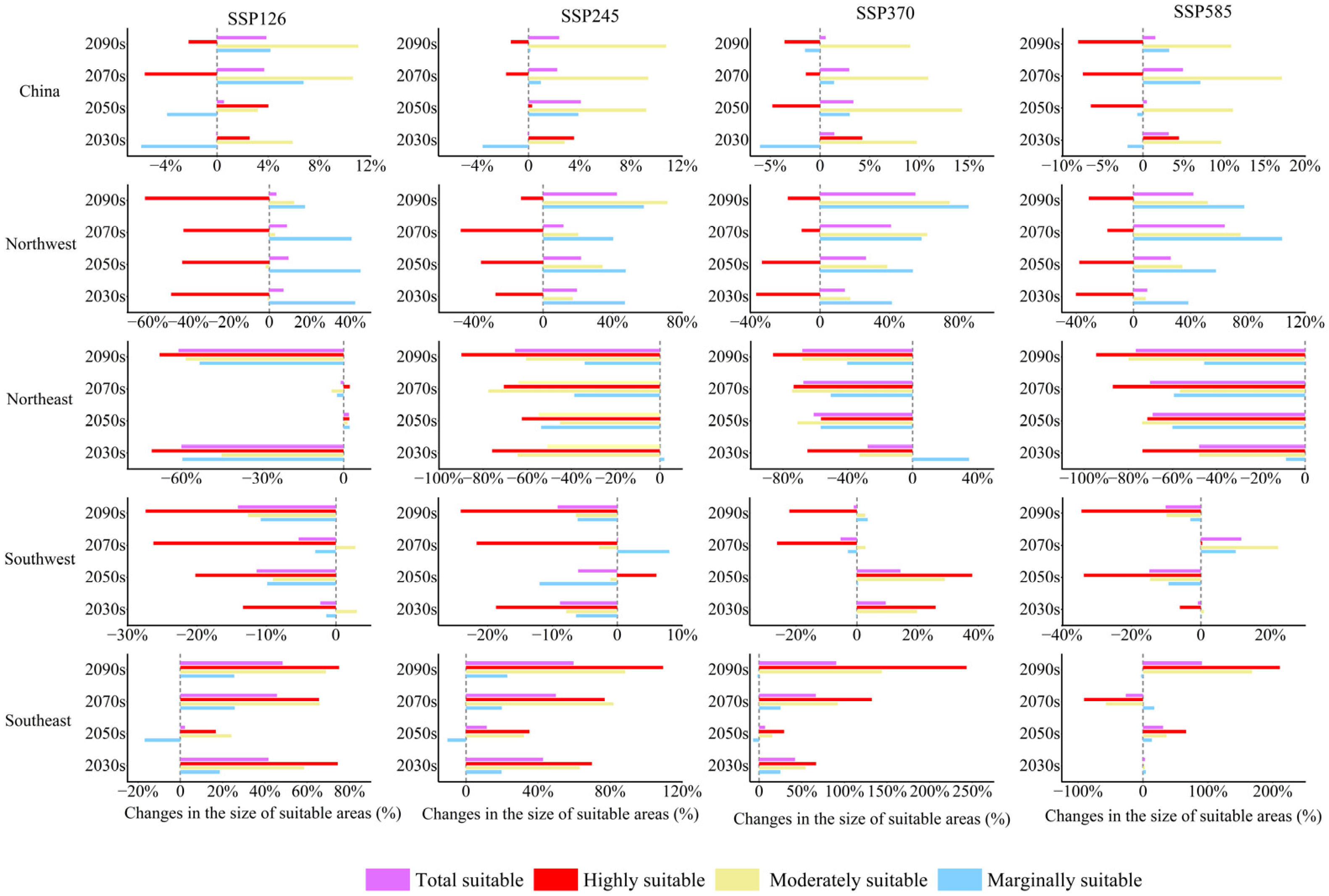
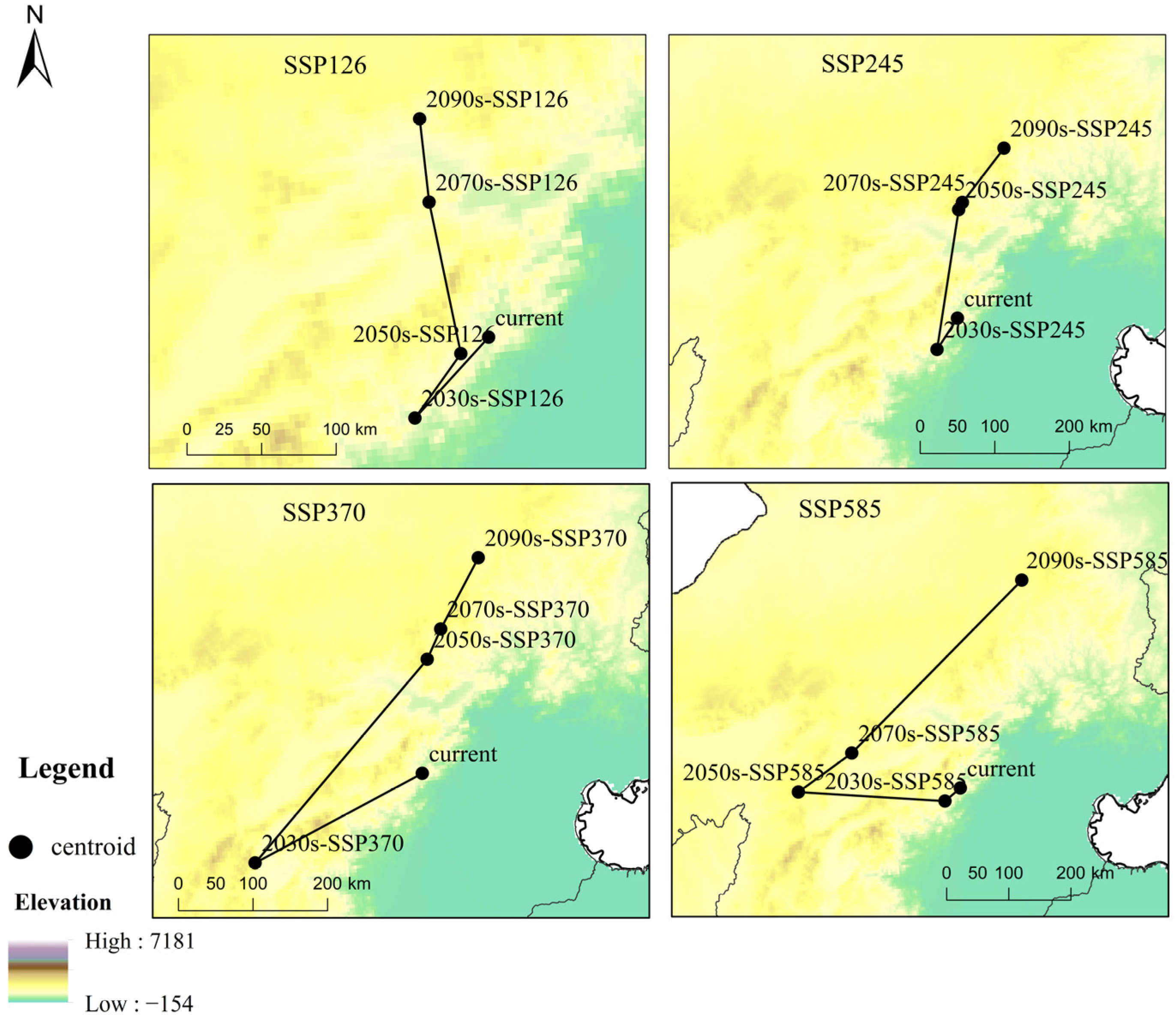
3.2. The Contraction and Expansion of the Suitable Areas
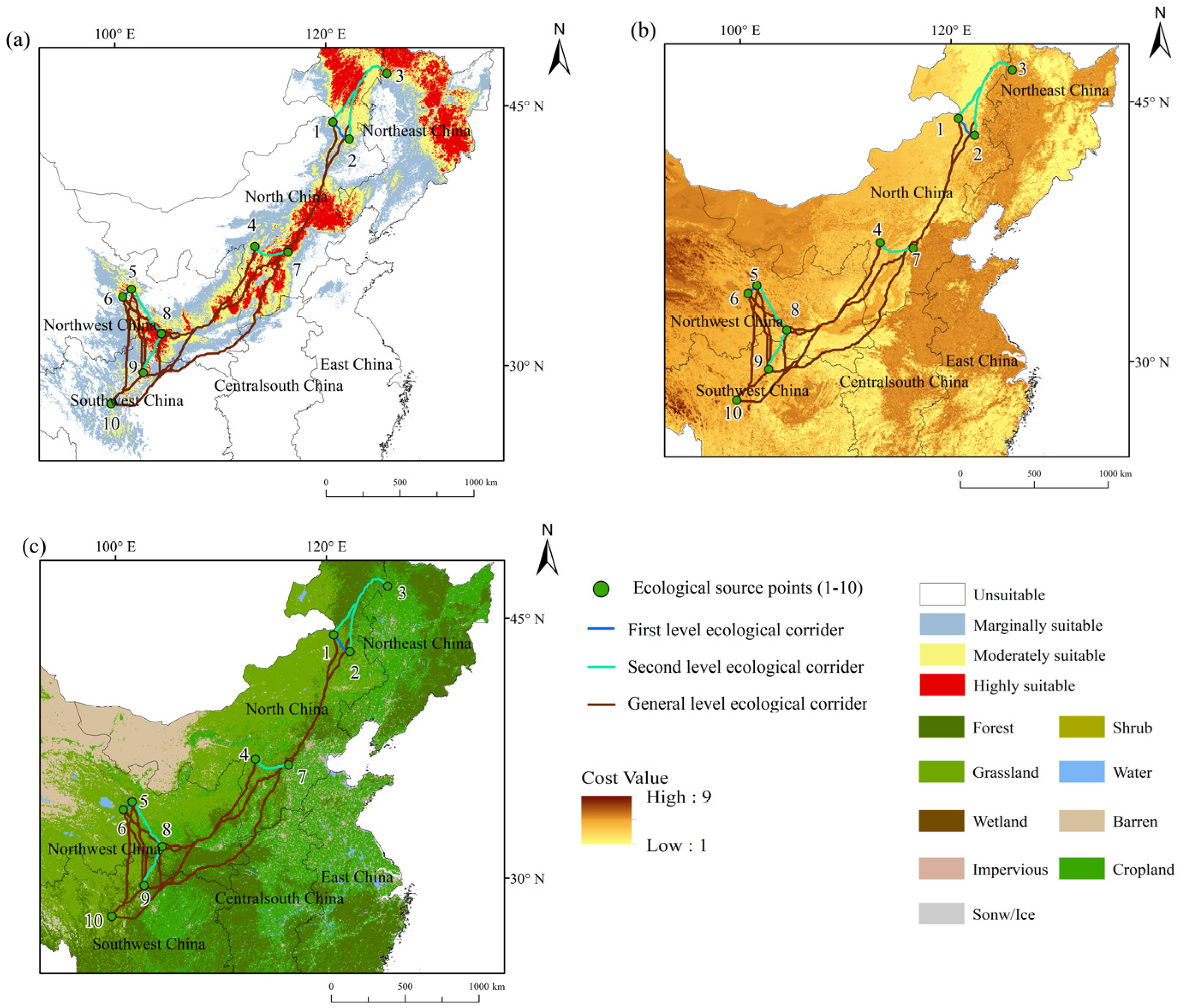
3.3. Construction of Ecological Corridors
4. Discussion
5. Conclusions
- (1)
- The highly suitable areas for B. platyphylla forest distributed in China are mainly concentrated in the Northeast and North China ecoregions, and precipitation in warmest quarter was the main environmental factor. In Northwest China, the highly suitable areas for B. platyphylla forest are in Gansu and Shaanxi Provinces; in Southwest China, in Sichuan Province; in North China, in Hebei Province and Inner Mongolia Autonomous Region. Annual precipitation is the main environmental factor in these three ecoregions. In Northeast Chin, the highly suitable areas for B. platyphylla forest are in Heilongjiang Province, and the mean temperature of the warmest quarter is the main factor affecting its distribution.
- (2)
- The total suitable areas of B. platyphylla forest showed an expanding trend in China, as well as in the ecoregions of North China and Northwest China, and a declining trend in the ecoregions of Northeast China and Southwest China. Therefore, ecological buffer zones should be designated to prevent excessive expansion of B. platyphylla forest from encroaching on the habitats of other rare tree species in the North China and Northwest China. In Northeast and Southwest China, logging and human activities should be strictly restricted to enhance the forest’s adaptability.
- (3)
- In total, 45 ecological corridors were identified, including 2 first-level corridors, 5 second-level corridors, and 38 general corridors. During ecological corridor construction, the existing patterns of land use must be carefully considered. It is advisable to primarily utilize forest land, with grassland as a supplement, and ensure that the ratio of construction to cultivated land within the corridors remains minimal.
6. Future Work
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, Y.Y.; Zheng, J.; Gao, J.; He, X.R.; Liu, X.L.; Chen, Y.Y.; Liu, J.C.; Li, C.X. Functional Diversity Explains Ecosystem Carbon Storage in Subtropical Forests. Glob. Change Biol. 2025, 31, e70120. [Google Scholar] [CrossRef]
- Deng, J.J.; Zhou, W.M.; Dai, L.M.; Yuan, Q.; Zhou, L.; Qi, L.; Yu, D.P. The Effects of Shrub Removal on Soil Microbial Communities in Primary Forest, Secondary Forest and Plantation Forest on Changbai Mountain. Microb. Ecol. 2023, 85, 642–658. [Google Scholar] [CrossRef]
- Taubert, F.; Fischer, R.; Groeneveld, J.; Lehmann, S.; Müller, M.S.; Rödig, E.; Wiegand, T.; Huth, A. Global patterns of tropical forest fragmentation. Nature 2018, 554, 519–522. [Google Scholar] [CrossRef]
- Ma, J.; Li, J.W.; Wu, W.B.; Liu, J.J. Global forest fragmentation change from 2000 to 2020. Nat. Commun. 2023, 14, 3752. [Google Scholar] [CrossRef]
- Liu, L.; Wang, L.J.; Song, L.K.; Sheng, M.Y. Carbon sequestration law by phytoliths in the bamboo forests: Insights for the management of phytolith carbon sink. Glob. Ecol. Conserv. 2025, 58, e03491. [Google Scholar] [CrossRef]
- Khatiwala, S.; Primeau, F.; Hall, T. Reconstruction of the history of anthropogenic CO2 concentrations in the ocean. Nature 2009, 462, 346-U110. [Google Scholar] [CrossRef] [PubMed]
- Gruber, N.; Bakker, D.C.E.; DeVries, T.; Gregor, L.; Hauck, J.; Landschuetzer, P.; McKinley, G.A.; Mueller, J.D. Trends and variability in the ocean carbon sink. Nat. Rev. Earth Environ. 2023, 4, 119–134. [Google Scholar] [CrossRef]
- Samaras, D.A.; Damianidis, C.; Fotiadis, G.; Tsiftsis, S. Effect of climate change on fir forest communities in the mountains of South-Central Greece. Eur. J. Environ. Sci. 2022, 12, 39–50. [Google Scholar] [CrossRef]
- Taylor, C.; Lindenmayer, D.B. Temporal fragmentation of a critically endangered forest ecosystem. Austral Ecol. 2020, 45, 340–354. [Google Scholar] [CrossRef]
- Lin, X.; Zhen, S.Y.; Zhao, Q.; Hu, X.S. A New Paradigm for Assessing Detailed Dynamics of Forest Landscape Fragmentation. Forests 2024, 15, 1212. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Ballantyne, A.P.; Smith, W.K.; Majkut, J.; Rabin, S.; Beaulieu, C.; Birdsey, R.; Dunne, J.P.; Houghton, R.A.; Myneni, R.B.; et al. Tropical nighttime warming as a dominant driver of variability in the terrestrial carbon sink. Proc. Natl. Acad. Sci. USA 2015, 112, 15591–15596. [Google Scholar] [CrossRef] [PubMed]
- Duffy, K.A.; Schwalm, C.R.; Arcus, V.L.; Koch, G.W.; Liang, L.Y.L.; Schipper, L.A. How close are we to the temperature tipping point of the terrestrial biosphere? Sci. Adv. 2021, 7, eaay1052. [Google Scholar] [CrossRef]
- Sitch, S.; Huntingford, C.; Gedney, N.; Levy, P.E.; Lomas, M.; Piao, S.L.; Betts, R.; Ciais, P.; Cox, P.; Friedlingstein, P.; et al. Evaluation of the terrestrial carbon cycle, future plant geography and climate-carbon cycle feedbacks using five Dynamic Global Vegetation Models (DGVMs). Glob. Change Biol. 2008, 14, 2015–2039. [Google Scholar] [CrossRef]
- Shen, X.Y.; Rezaei, T.; Kachenchart, B.; Tanhan, P.; Chaiyarat, R. Optimal region connection: Establishing effective ecological corridors for biodiversity conservation in Yunnan Province, China. Ecol. Indic. 2024, 169, 112918. [Google Scholar] [CrossRef]
- Beita, C.M.; Murillo, L.F.S.; Alvarado, L.D.A. Ecological corridors in Costa Rica: An evaluation applying landscape structure, fragmentation-connectivity process, and climate adaptation. Conserv. Sci. Pract. 2021, 3, e475. [Google Scholar] [CrossRef]
- Liu, D.; An, Y.; Li, Z.; Wang, Z.H.; Zhao, Y.H.; Wang, X.C. Differences and similarities in radial growth of Betula species to climate change. J. For. Res. 2024, 35, 40. [Google Scholar] [CrossRef]
- Gao, R.M.; Shi, X.D.; Wang, J.R. Comparative studies of the response of larch and birch seedlings from two origins to water deficit. New Zeal. J. For. Sci. 2017, 47, 14. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Ngatia, J.N.; Song, R.X.; Farooq, U.; Somadona, S.; Lestari, A.T.; Chen, S. Abiotic stresses induced physiological, biochemical, and molecular changes in Betula platyphylla: A review. Silva Fenn. 2021, 55, 10516. [Google Scholar] [CrossRef]
- Liu, L.F.; Qin, F.C.; Liu, Y.; Hu, Y.N.; Wang, W.F.; Duan, H.; Li, M.Y. Forecast of potential suitable areas for forest resources in Inner Mongolia under the Shared Socioeconomic Pathway 245 scenario. Ecol. Indic. 2024, 167, 112694. [Google Scholar] [CrossRef]
- Dolezal, J.; Ishii, H.; Kyncl, T.; Takahashi, K.; Vetrova, V.P.; Homma, K.; Sumida, A.; Hara, T. Climatic factors affecting radial growth of Betula ermanii and Betula platypylla in Kamchatka. Can. J. For. Res. 2010, 40, 273–285. [Google Scholar] [CrossRef]
- Takahashi, K.; Azuma, H.; Yasue, K. Effects of climate on the radial growth of tree species in the upper and lower distribution limits of an altitudinal ecotone on Mount Norikura, central Japan. Ecol. Res. 2003, 18, 549–558. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Pei, X.N.; Xu, L.F.; Lu, X.B.; Wen, B.Y.; Li, Y.L.; Wang, L.K.; Dong, G.Z.; Shi, W.L.; Hu, X.Q.; et al. Genetic Improvement of Betula platyphylla Suk. in China: A Review. Phyton-Int. J. Exp. Bot. 2022, 91, 1585–1599. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, J.J.; Qu, G.Z.; Chen, S. The cold-responsive C-repeat binding factors in Betula platyphylla Suk. positively regulate cold tolerance. Plant Sci. 2024, 341, 112012. [Google Scholar] [CrossRef]
- Lei, X.J.; Liu, Z.Y.; Li, X.P.; Tan, B.; Wu, J.; Gao, C.Q. Screening and functional identification of salt tolerance HMG genes in Betula platyphylla. Environ. Exp. Bot. 2021, 181, 104235. [Google Scholar] [CrossRef]
- Geng, W.L.; Li, Y.Y.; Sun, D.Q.; Li, B.; Zhang, P.Y.; Chang, H.; Rong, T.Q.; Liu, Y.; Shao, J.W.; Liu, Z.Y.; et al. Prediction of the potential geographical distribution of Betula platyphylla Suk. in China under climate change scenarios. PLoS ONE 2022, 17, e0262540. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.N.; Zheng, Y.T.; Wang, W.; Wang, Z.; Zhang, Q.F.; Liu, J.C.; Zhang, C.T. Habitat Suitability Evaluation of Different Forest Species in Lvliang Mountain by Combining Prior Knowledge and MaxEnt Model. Forests 2023, 14, 438. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Djordjevic, V.; Tsiripidis, I. Neottia cordata (Orchidaceae) at its southernmost distribution border in Europe: Threat status and effectiveness of Natura 2000 Network for its conservation. J. Nat. Conserv. 2019, 48, 27–35. [Google Scholar] [CrossRef]
- Jha, A.; Praveen, J.; Nameer, P.O. Contrasting occupancy models with presence-only models: Does accounting for detection lead to better predictions? Ecol. Model. 2022, 472, 110105. [Google Scholar] [CrossRef]
- Ahmadi, M.; Hemami, M.R.; Kaboli, M.; Shabani, F. MaxEnt brings comparable results when the input data are being completed; Model parameterization of four species distribution models. Ecol. Evol. 2023, 13, e9827. [Google Scholar] [CrossRef]
- Men, D.; Pan, J.H. Integrating key species distribution and ecosystem service flows to build directed ecological network: Evidence from the Shiyang River Basin, China. J. Environ. Manag. 2025, 381, 125183. [Google Scholar] [CrossRef]
- Yagoobi, S.; Sharma, N.; Traulsen, A. Categorizing update mechanisms for graph-structured metapopulations. J. R. Soc. Interface 2023, 20, 20220769. [Google Scholar] [CrossRef]
- Wu, Y.H.; Qin, F.C.; Li, L.; Dong, X.Y. Construction and optimisation of watershed scale ecological network: A case study of kuye river basin. Front. Environ. Sci. 2024, 12, 1364568. [Google Scholar] [CrossRef]
- Yuan, Q.; Li, R. The negative impacts of human activities on the ecological corridor in the karst highly urbanized area are gradually diminishing: A case study from the karst mountain cities in Southwest China. Ecol. Indic. 2023, 157, 111257. [Google Scholar] [CrossRef]
- Li, P.X.; Liu, C.G.; Sun, W. Quantifying changes of landscape connectivity based on ecological process simulation in a rapidly urbanized city: Nanjing, China. Environ. Earth Sci. 2021, 80, 644. [Google Scholar] [CrossRef]
- Liu, M.X.; Li, L.; Wang, S.Y.; Xiao, S.R.; Mi, J.L. Forecasting the future suitable growth areas and constructing ecological corridors for the vulnerable species Ephedra sinica in China. J. Nat. Conserv. 2023, 73, 126401. [Google Scholar] [CrossRef]
- Fu, H.X.; Zhang, T.; Wang, J.G. Evaluating suitability of development and construction with of minimum cumulative resistance model for a mountain scenic area in Jinyun Xiandu, China. Ecol. Eng. 2024, 202, 107240. [Google Scholar] [CrossRef]
- Luo, L.Z.; Yang, C.X.; Chen, R.R.; Liu, W.P. Comprehensive Land Consolidation Zoning Based on Minimum Cumulative Resistance Model-A Case Study of Chongqing, Southwest China. Land 2023, 12, 1935. [Google Scholar] [CrossRef]
- Hou, X.Y. Vegetation Atlas of China; Chinese Academy of Science, the Editorial Board of Vegetation Map of China Scientific Press: Beijing, China, 2001. [Google Scholar]
- Warren, D.L.; Beaumont, L.J.; Dinnage, R.; Baumgartner, J.B. New methods for measuring ENM breadth and overlap in environmental space. Ecography 2019, 42, 444–446. [Google Scholar] [CrossRef]
- Mu, H.W.; Li, X.C.; Wen, Y.A.; Huang, J.X.; Du, P.J.; Su, W.; Miao, S.X.; Geng, M.Q. A global record of annual terrestrial Human Footprint dataset from 2000 to 2018. Sci. Data 2022, 9, 176. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Huang, X. The 30 m annual land cover dataset and its dynamics in China from 1990 to 2019. Earth Syst. Sci. Data 2021, 13, 3907–3925. [Google Scholar] [CrossRef]
- Tokarska, K.B.; Stolpe, M.B.; Sippel, S.; Fischer, E.M.; Smith, C.J.; Lehner, F.; Knutti, R. Past warming trend constrains future warming in CMIP6 models. Sci. Adv. 2020, 6, eaaz9549. [Google Scholar] [CrossRef]
- Sang, Y.H.; Ren, H.L.; Shi, X.L.; Xu, X.F.; Chen, H.S. Improvement of Soil Moisture Simulation in Eurasia by the Beijing Climate Center Climate System Model from CMIP5 to CMIP6. Adv. Atmos. Sci. 2021, 38, 237–252. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Zhou, Y.; Ji, X.D.; Wang, Z.Y.; Liu, Z. Climate Change Drives the Adaptive Distribution and Habitat Fragmentation of Betula albosinensis Forests in China. Forests 2025, 16, 184. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Y.; Yang, Y.S.; Wang, T.X.; Wu, C.; Zhang, X.J. Prediction of Historical, Current, and Future Configuration of Tibetan Medicinal Herb Gymnadenia orchidis Based on the Optimized MaxEnt in the Qinghai-Tibet Plateau. Plants-Basel 2024, 13, 645. [Google Scholar] [CrossRef]
- Yang, W.J.; Sun, S.X.; Wang, N.X.; Fan, P.X.; You, C.; Wang, R.Q.; Zheng, P.M.; Wang, H. Dynamics of the distribution of invasive alien plants (Asteraceae) in China under climate change. Sci. Total Environ. 2023, 903, 166260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Li, J.P.; Zou, H.C.; Wang, Z.Y.; Zhu, X.Y.; Zhang, Y.H.; Liu, Z. Distribution Pattern of Suitable Areas and Corridor Identification of Endangered Ephedra Species in China. Plants 2024, 13, 890. [Google Scholar] [CrossRef] [PubMed]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Peterson, A.T.; Cohoon, K.P. Sensitivity of distributional prediction algorithms to geographic data completeness. Ecol. Model. 1999, 117, 159–164. [Google Scholar] [CrossRef]
- Yang, M.; Zhao, H.X.; Xian, X.Q.; Liu, H.; Li, J.Y.; Chen, L.; Liu, W.X. Potential global geographical distribution of Lolium temulentum L. under climate change. Front. Plant Sci. 2022, 13, 1024635. [Google Scholar] [CrossRef]
- Goncharenko, I.; Krakhmalnyi, M.; Velikova, V.; Ascencio, E.; Krakhmalnyi, A. Ecological niche modeling of toxic dinoflagellate Prorocentrum cordatum in the Black Sea. Ecohydrol. Hydrobiol. 2021, 21, 747–759. [Google Scholar] [CrossRef]
- Feng, M.; Zhao, W.M.; Zhang, T. Construction and Optimization Strategy of County Ecological Infrastructure Network Based on MCR and Gravity Model-A Case Study of Langzhong County in Sichuan Province. Sustainability 2023, 15, 8478. [Google Scholar] [CrossRef]
- Liang, C.; Zeng, J.; Zhang, R.C.; Wang, Q.W. Connecting urban area with rural hinterland: A stepwise ecological security network construction approach in the urban-rural fringe. Ecol. Indic. 2022, 138, 108794. [Google Scholar] [CrossRef]
- Zhao, S.M.; Ma, Y.F.; Wang, J.L.; You, X.Y. Landscape pattern analysis and ecological network planning of Tianjin City. Urban For. Urban Green. 2019, 46, 126479. [Google Scholar] [CrossRef]
- Wang, X.K.; Xie, X.Q.; Wang, Z.F.; Lin, H.; Liu, Y.; Xie, H.L.; Liu, X.Z. Construction and Optimization of an Ecological Security Pattern Based on the MCR Model: A Case Study of the Minjiang River Basin in Eastern China. Int. J. Environ. Res. Public Health 2022, 19, 8370. [Google Scholar] [CrossRef]
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with Egyptian medicinal plants. Ecol. Inform. 2020, 60, 101150. [Google Scholar] [CrossRef]
- Ramírez-Rodríguez, R.; Rocha, J.; Crespí, A.L.; Amich, F. Modelling the present potential habitat distribution of the near-threatened endemic species Silene marizii: Implications for conservation. Plant Biosyst. 2025, 159, 154–163. [Google Scholar] [CrossRef]
- He, P.; Li, J.Y.; Li, Y.F.; Xu, N.; Gao, Y.; Guo, L.F.; Huo, T.T.; Peng, C.; Meng, F.Y. Habitat protection and planning for three Ephedra using the MaxEnt and Marxan models. Ecol. Indic. 2021, 133, 108399. [Google Scholar] [CrossRef]
- Grieger, R.; Capon, S.J.; Hadwen, W.L.; Mackey, B. Spatial variation and drivers of vegetation structure and composition in coastal freshwater wetlands of subtropical Australia. Mar. Freshw. Res. 2021, 72, 1746–1759. [Google Scholar] [CrossRef]
- Zhao, D.S.; Zhu, Y.; Wu, S.H.; Zheng, D. Projection of vegetation distribution to 1.5 °C and 2 °C of global warming on the Tibetan Plateau. Glob. Planet. Change 2021, 202, 103525. [Google Scholar] [CrossRef]
- Koike, T.; Kitao, M.; Quoreshi, A.M.; Matsuura, Y. Growth characteristics of root-shoot relations of three birch seedlings raised under different water regimes. Plant Soil 2003, 255, 303–310. [Google Scholar] [CrossRef]
- Xiao, C.W.; Sang, W.G.; Wang, R.Z. Fine root dynamics and turnover rate in an Asia white birch forest of Donglingshan Mountain, China. For. Ecol. Manag. 2008, 255, 765–773. [Google Scholar] [CrossRef]
- Ranney, T.G.; Bir, R.E.; Skroch, W.A. Comparative drought resistance among 6 species of birch (Betula)—Influence of mild water-stress on water relations and leaf gas-exchange. Tree Physiol. 1991, 8, 351–360. [Google Scholar] [CrossRef]
- Schmitt, C.B.; Senbeta, F.; Woldemariam, T.; Rudner, M.; Denich, M. Importance of regional climates for plant species distribution patterns in moist Afromontane forest. J. Veg. Sci. 2013, 24, 553–568. [Google Scholar] [CrossRef]
- Pinilla-Buitrago, G.E. Predicting potential range shifts using climatic time series and niche models: A Neotropical montane shrew’s case. Ecol. Inform. 2023, 77, 102212. [Google Scholar] [CrossRef]
- Li, H.; Xu, F.; Li, Z.C.; You, N.S.; Zhou, H.; Zhou, Y.; Chen, B.Q.; Qin, Y.W.; Xiao, X.M.; Dong, J.W. Forest Changes by Precipitation Zones in Northern China after the Three-North Shelterbelt Forest Program in China. Remote Sens. 2021, 13, 543. [Google Scholar] [CrossRef]
- Yang, J.H.; Li, Y.Q.; Zhou, L.; Zhang, Z.Q.; Zhou, H.K.; Wu, J.J. Effects of temperature and precipitation on drought trends in Xinjiang, China. J. Arid Land 2024, 16, 1098–1117. [Google Scholar] [CrossRef]
- Wang, B.; Dong, X.G.; Wang, Z.H.; Qin, G.Q. Characterizing Spatiotemporal Variations of Soil Salinization and Its Relationship with Eco-Hydrological Parameters at the Regional Scale in the Kashi Area of Xinjiang, China from 2000 to 2017. Water 2021, 13, 1075. [Google Scholar] [CrossRef]
- Li, J.; Gong, Q.; Zhao, L.W. Climatic features of summer temperature in Northeast China under warming background. Chin. Geogr. Sci. 2005, 15, 337–342. [Google Scholar] [CrossRef]
- Yuan, D.Y.; Zhu, L.J.; Cherubini, P.; Li, Z.S.; Zhang, Y.D.; Wang, X.C. Species-specific indication of 13 tree species growth on climate warming in temperate forest community of northeast China. Ecol. Indic. 2021, 133, 108389. [Google Scholar] [CrossRef]
- Yu, J.; Wu, C.J.; Jia, C.J.; Xu, W.Y. Study of variation of pore properties in gravel soil under triaxial loading based on discrete element method. Curr. Sci. 2021, 121, 801–809. [Google Scholar] [CrossRef]
- Harvey, B.J.; Cook, P.; Shaffrey, L.C.; Schiemann, R. The Response of the Northern Hemisphere Storm Tracks and Jet Streams to Climate Change in the CMIP3, CMIP5, and CMIP6 Climate Models. J. Geophys. Res. Atmos. 2020, 125, e2020JD032701. [Google Scholar] [CrossRef]
- Yao, W.H.; Wang, Z.H.; Fan, Y.; Liu, D.Y.; Ding, Z.Y.; Zhou, Y.M.; Hu, S.Y.; Zhang, W.; Ou, J. Prediction of Potential Habitat Distributions and Climate Change Impacts on the Rare Species Woonyoungia septentrionalis (Magnoliaceae) in China Based on MaxEnt. Plants 2025, 14, 86. [Google Scholar] [CrossRef]
- Hadinejad, M.; Naghipour, A.A.; Ebrahimi, A.; Naimi, B. Modeling the effect of climate change on the distribution of plant communities in Zayandeh-Rud basin, Iran. Environ. Monit. Assess. 2025, 197, 479. [Google Scholar] [CrossRef]
- Li, X.X.; Yang, D.S.; Wang, J.J.; Pan, G. Prediction of the change in suitable growth area of Sabina tibetica on the Qinghai-Tibetan plateau using MaxEnt model. Front. For. Glob. Change 2025, 8, 1465416. [Google Scholar] [CrossRef]
- Zhang, M.G.; Zhou, Z.K.; Chen, W.Y.; Cannon, C.H.; Raes, N.; Slik, J.W.F. Major declines of woody plant species ranges under climate change in Yunnan, China. Divers. Distrib. 2014, 20, 405–415. [Google Scholar] [CrossRef]
- Lin, X.K.; Chang, B.L.; Huang, Y.Q.; Jin, X. Predicting the impact of climate change and land use change on the potential distribution of two economic forest trees in Northeastern China. Front. Plant Sci. 2024, 15, 1407867. [Google Scholar] [CrossRef] [PubMed]
- Raghunathan, N.; Francois, L.; Dury, M.; Hambuckers, A. Contrasting climate risks predicted by dynamic vegetation and ecological niche-based models applied to tree species in the Brazilian Atlantic Forest. Reg. Environ. Change 2019, 19, 219–232. [Google Scholar] [CrossRef]
- Lemordant, L.; Gentine, P. Vegetation Response to Rising CO2 Impacts Extreme Temperatures. Geophys. Res. Lett. 2019, 46, 1383–1392. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Q.; Li, S.H.; Zhang, Y.W.; Xiang, Y.Z.; Yang, Y.; Zhang, J.X. Spatiotemporal Dynamics of Ilex macrocarpa Distribution Under Future Climate Scenarios: Implications for Conservation Planning. Forests 2025, 16, 370. [Google Scholar] [CrossRef]
- Dong, P.B.; Wang, L.J.; Qiu, D.Y.; Liang, W.; Cheng, J.L.; Wang, H.Y.; Guo, F.X.; Chen, Y. Evaluation of the environmental factors influencing the quality of Astragalus membranaceus var. mongholicus based on HPLC and the Maxent model. BMC Plant Biol. 2024, 24, 697. [Google Scholar] [CrossRef]
- Wu, C.W.; Xu, X.X.; Zhang, G.J.; Cheng, B.B.; Han, S. Predicting the potential suitable habitat for Tamarix chinensis under climate change based on CMIP6 in China. Appl. Ecol. Environ. Res. 2022, 20, 2845–2863. [Google Scholar] [CrossRef]
- Bai, J.Y.; Wang, H.C.; Hu, Y.K. Prediction of Potential Suitable Distribution of Liriodendron chinense (Hemsl.) Sarg. in China Based on Future Climate Change Using the Optimized MaxEnt Model. Forests 2024, 15, 988. [Google Scholar] [CrossRef]
- Khan, Z.; Ali, S.A.; Parvin, F.; Mohsin, M.; Shamim, S.K.; Ahmad, A. Predicting the effects of climate change on prospective Banj oak (Quercus leucotrichophora) dispersal in Kumaun region of Uttarakhand using machine learning algorithms. Model. Earth Syst. Environ. 2023, 9, 145–156. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Symbol | Environmental Variables | Unit |
|---|---|---|---|
| Bioclimatic | Bio1 | Annual mean temperature | °C |
| Bio2 | Mean diurnal range | °C | |
| Bio3 | Isothermality | \ | |
| Bio4 | Temperature seasonality | \ | |
| Bio5 | Max temperature of warmest month | °C | |
| Bio6 | Min temperature of coldest month | °C | |
| Bio7 | Temperature annual range | °C | |
| Bio8 | Mean temperature of wettest quarter | °C | |
| Bio9 | Mean temperature of driest quarter | °C | |
| Bio10 | Mean temperature of warmest quarter | °C | |
| Bio11 | Mean temperature of coldest quarter | °C | |
| Bio12 | Annual precipitation | mm | |
| Bio13 | Precipitation of wettest month | mm | |
| Bio14 | Precipitation of driest month | mm | |
| Bio15 | Precipitation seasonality | \ | |
| Bio16 | Precipitation of wettest quarter | mm | |
| Bio17 | Precipitation of driest quarter | mm | |
| Bio18 | Precipitation of warmest quarter | mm | |
| Bio19 | Precipitation of coldest quarter | mm | |
| Soil | BD | Soil bulk density | g/cm3 |
| Btcly | Clay content | g/kg | |
| Btslt | Silt content | g/kg | |
| Btsnd | Sand content | g/kg | |
| CEC | Cation exchange capacity | cmol/kg | |
| CF | Gravel content | % | |
| pH | Power of hydrogen | \ | |
| SOC | Soil organic matter | g/kg | |
| SOCD | Soil organic matter density | kg/m2 | |
| TK | Total potassium | g/kg | |
| TKD | Total potassium content | kg/m2 | |
| TN | Total nitrogen | g/kg | |
| TND | Total nitrogen content | kg/m2 | |
| TP | Total phosphorus | g/kg | |
| TPD | Total phosphorus content | kg/m2 | |
| Terrain | Elev | Elevation | m |
| Aspect | Aspect | ° | |
| Slope | Slope | ° | |
| HFP | HFP | Human Footprint | \ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, B.; Zhang, H.; Ji, X.; Zhao, B.; Wei, Y.; Peng, Y.; Liu, Z. Corridors of Suitable Distribution of Betula platyphylla Sukaczev Forest in China Under Climate Warming. Sustainability 2025, 17, 6937. https://doi.org/10.3390/su17156937
Xie B, Zhang H, Ji X, Zhao B, Wei Y, Peng Y, Liu Z. Corridors of Suitable Distribution of Betula platyphylla Sukaczev Forest in China Under Climate Warming. Sustainability. 2025; 17(15):6937. https://doi.org/10.3390/su17156937
Chicago/Turabian StyleXie, Bingying, Huayong Zhang, Xiande Ji, Bingjian Zhao, Yanan Wei, Yijie Peng, and Zhao Liu. 2025. "Corridors of Suitable Distribution of Betula platyphylla Sukaczev Forest in China Under Climate Warming" Sustainability 17, no. 15: 6937. https://doi.org/10.3390/su17156937
APA StyleXie, B., Zhang, H., Ji, X., Zhao, B., Wei, Y., Peng, Y., & Liu, Z. (2025). Corridors of Suitable Distribution of Betula platyphylla Sukaczev Forest in China Under Climate Warming. Sustainability, 17(15), 6937. https://doi.org/10.3390/su17156937