
Numerical Simulation of Wind and Sand Resistance in Three Typical Shrubs


Abstract
1. Introduction
2. Materials and Methods
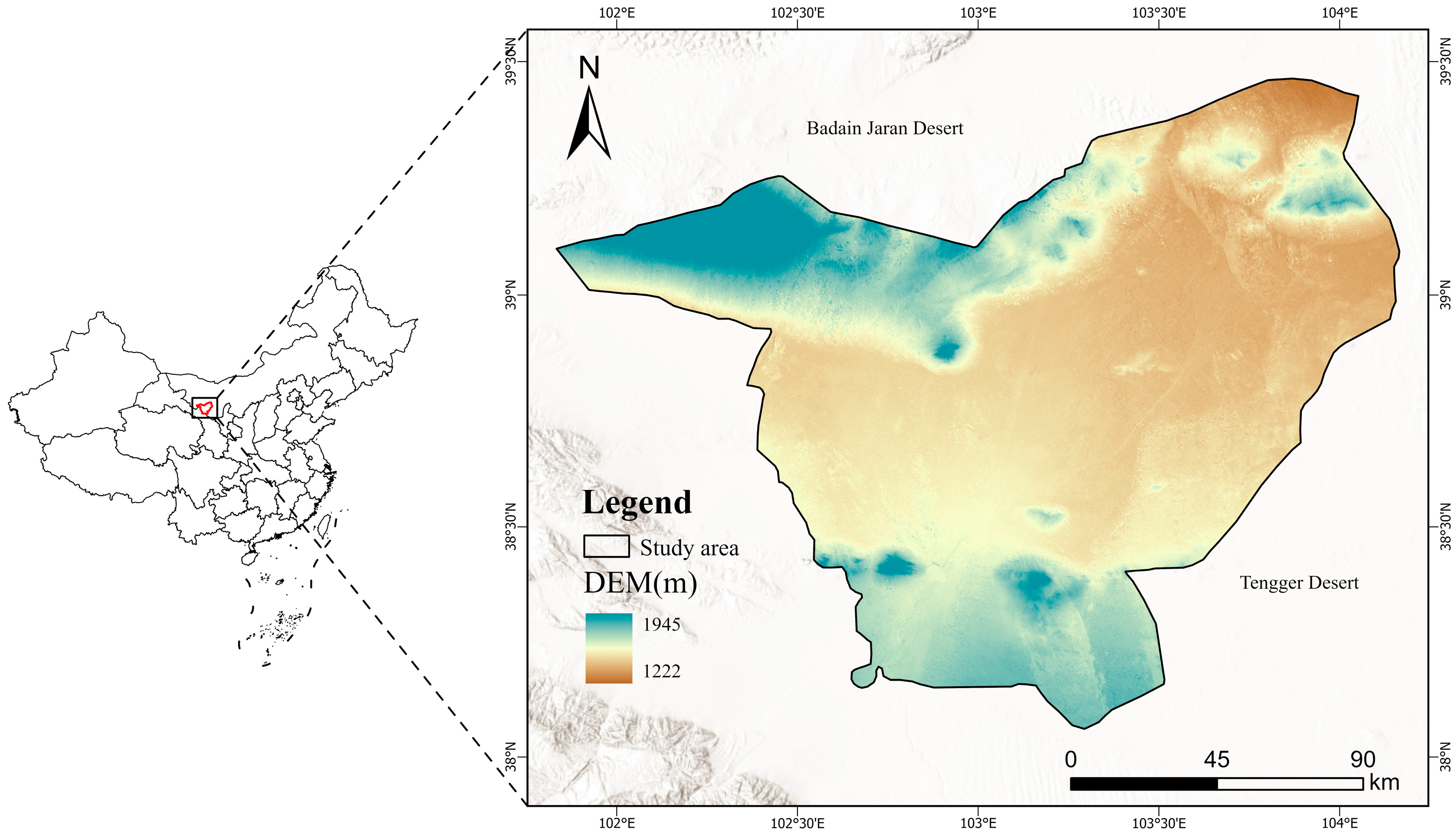
2.1. Objects of Study
2.2. Numerical Simulation
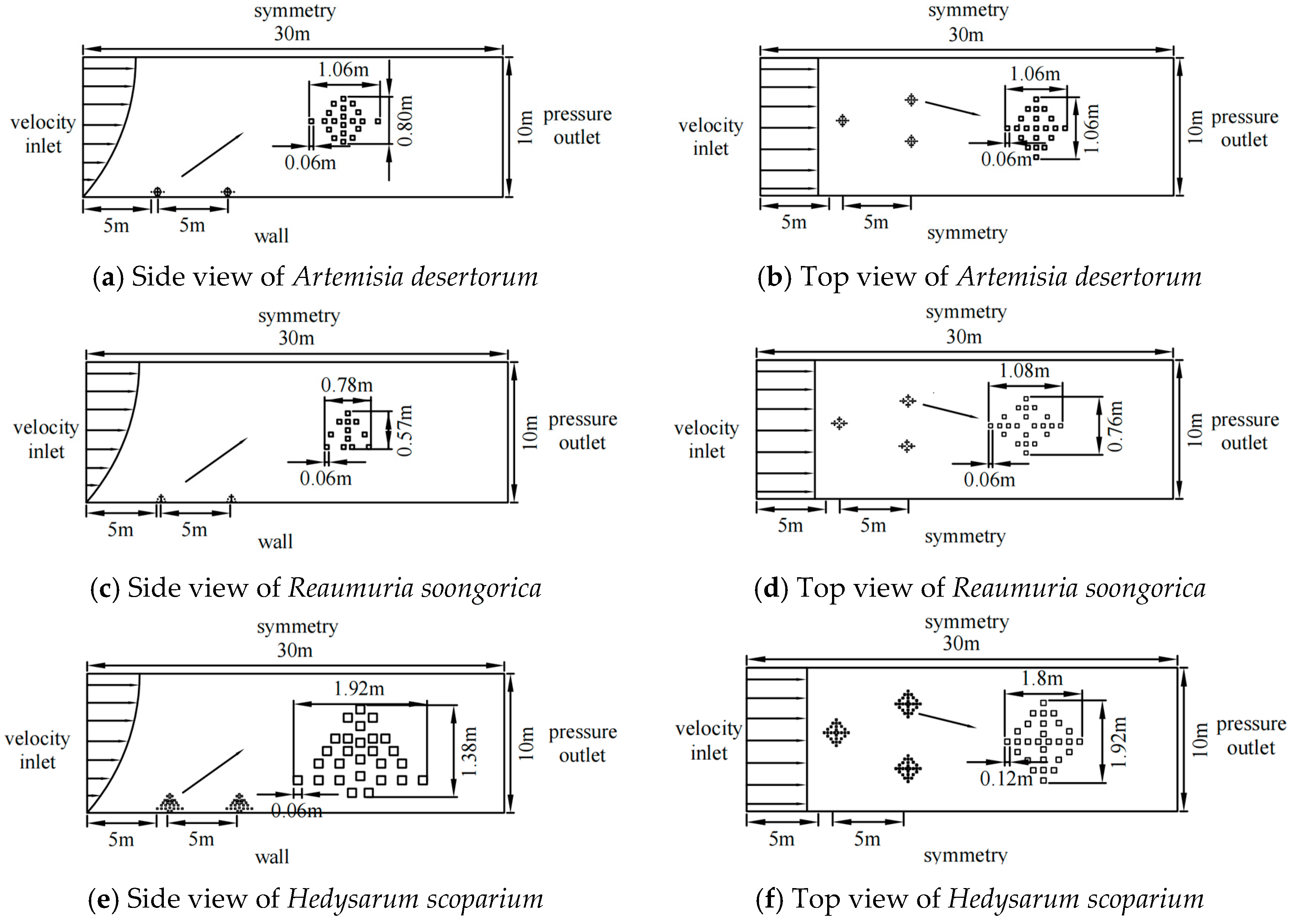
2.2.1. Modeling
2.2.2. Setting Parameters
3. Results
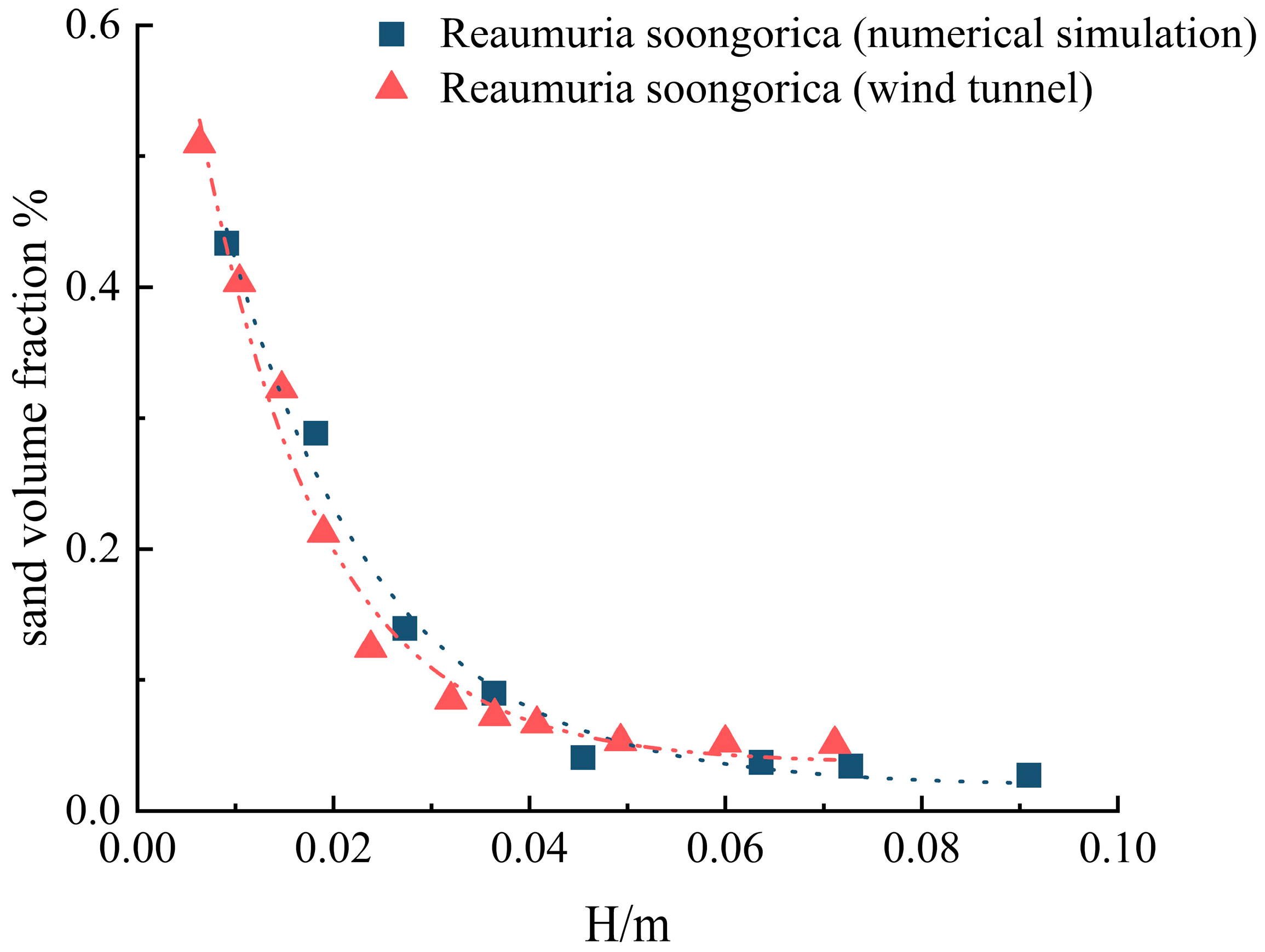
3.1. Wind Tunnel Test
3.2. The Shelter Effect of Different Shrubs
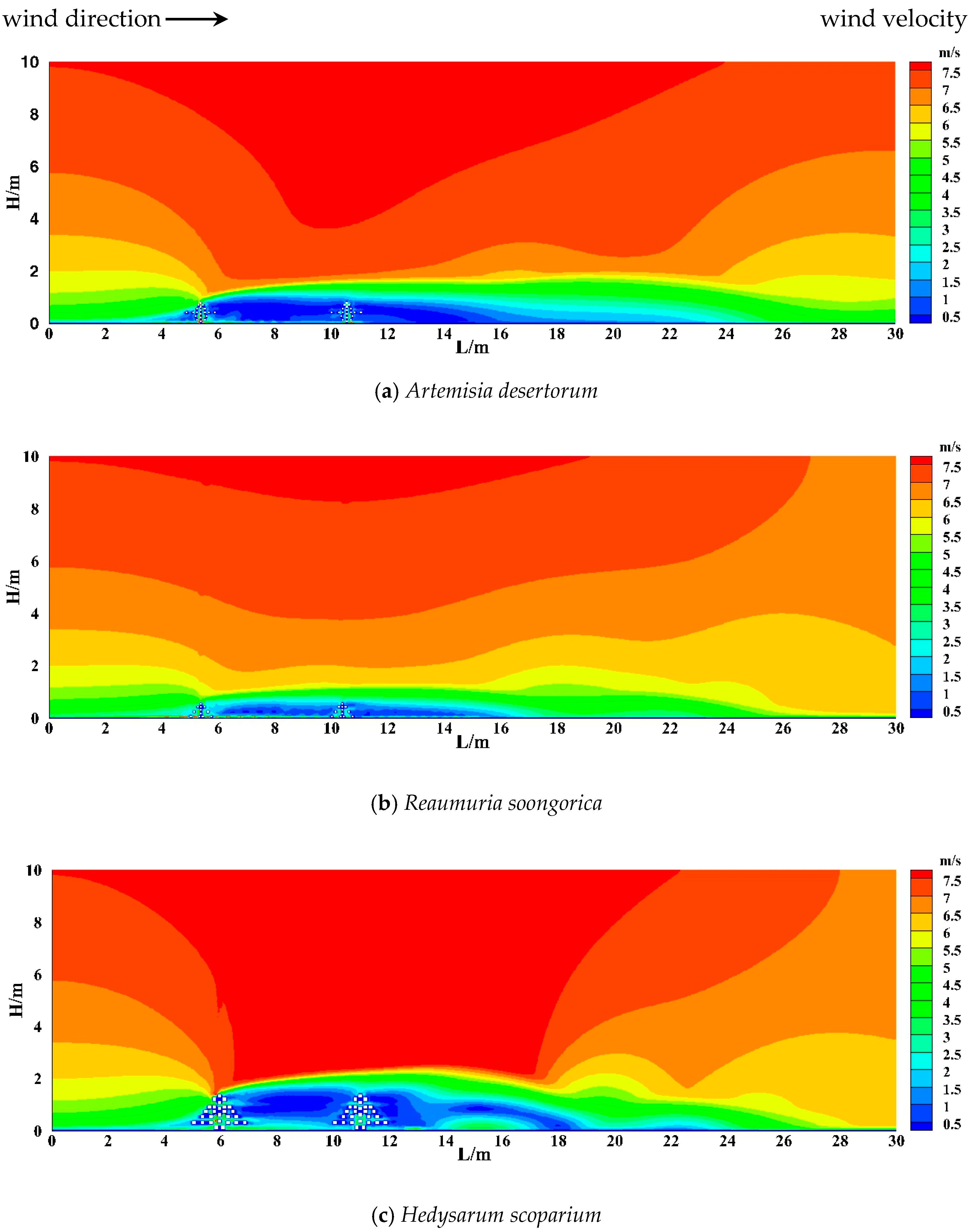
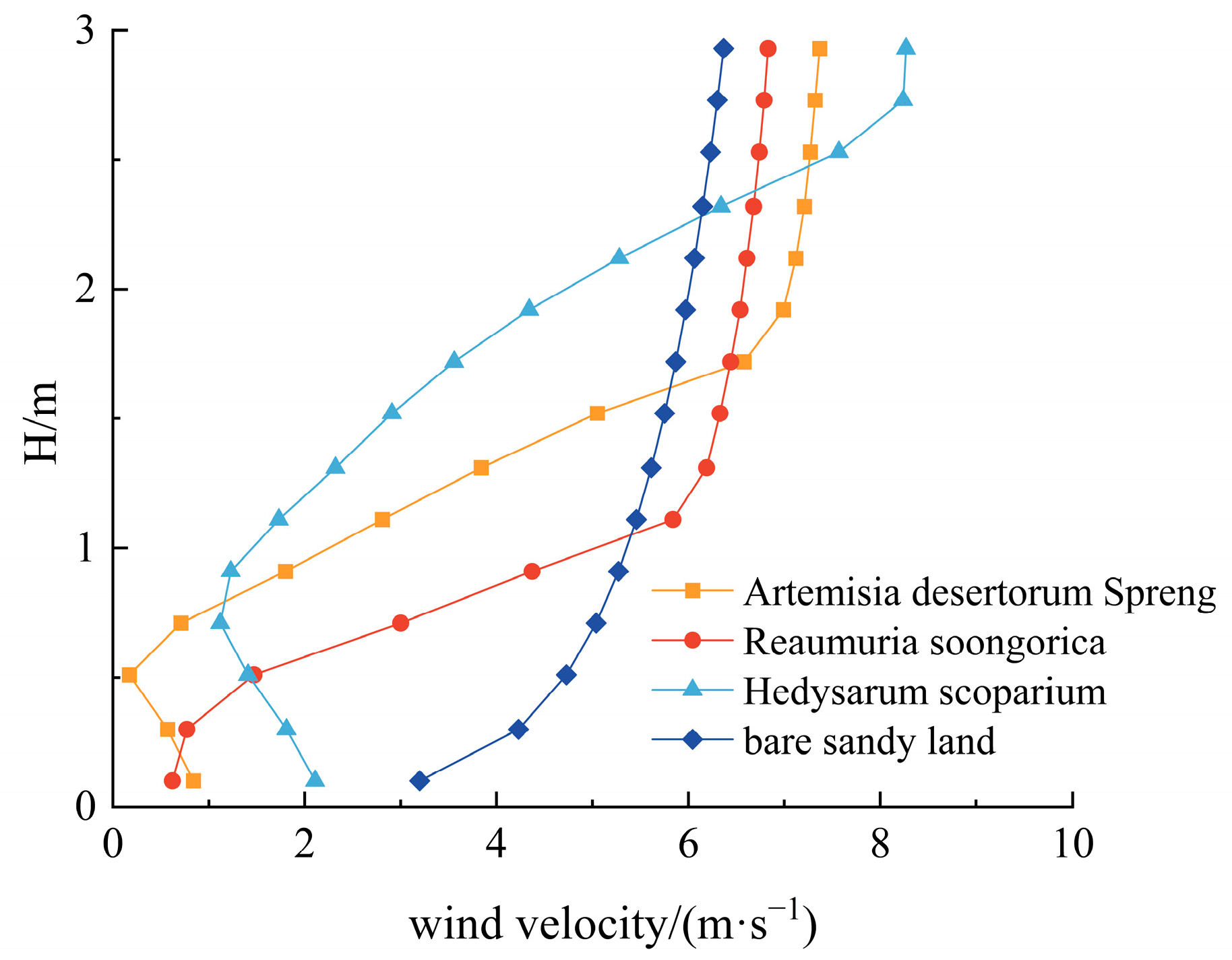
3.2.1. The Vertical Shelter Effect of Shrubs
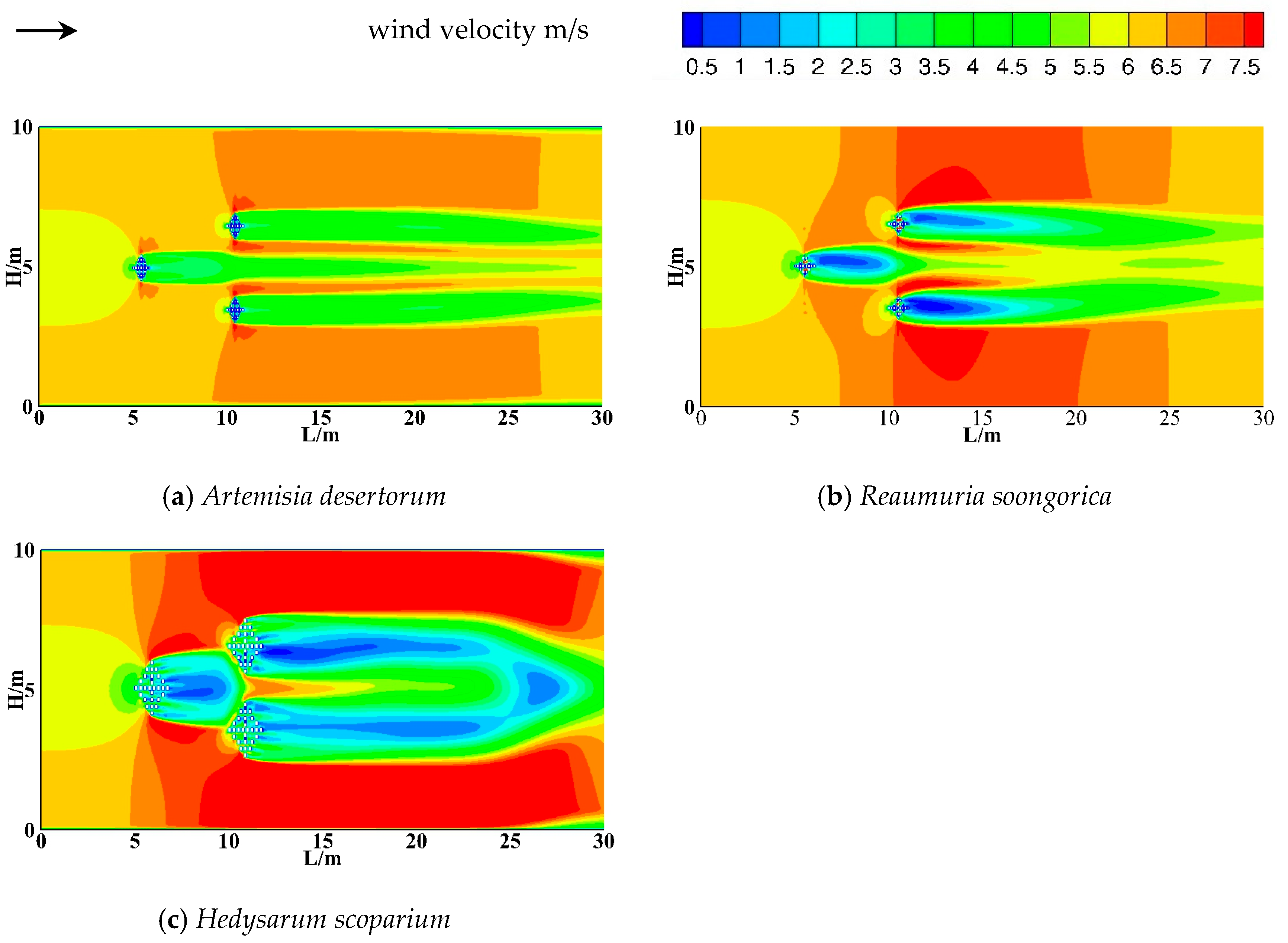
3.2.2. The Horizontal Shelter Effect of Shrubs
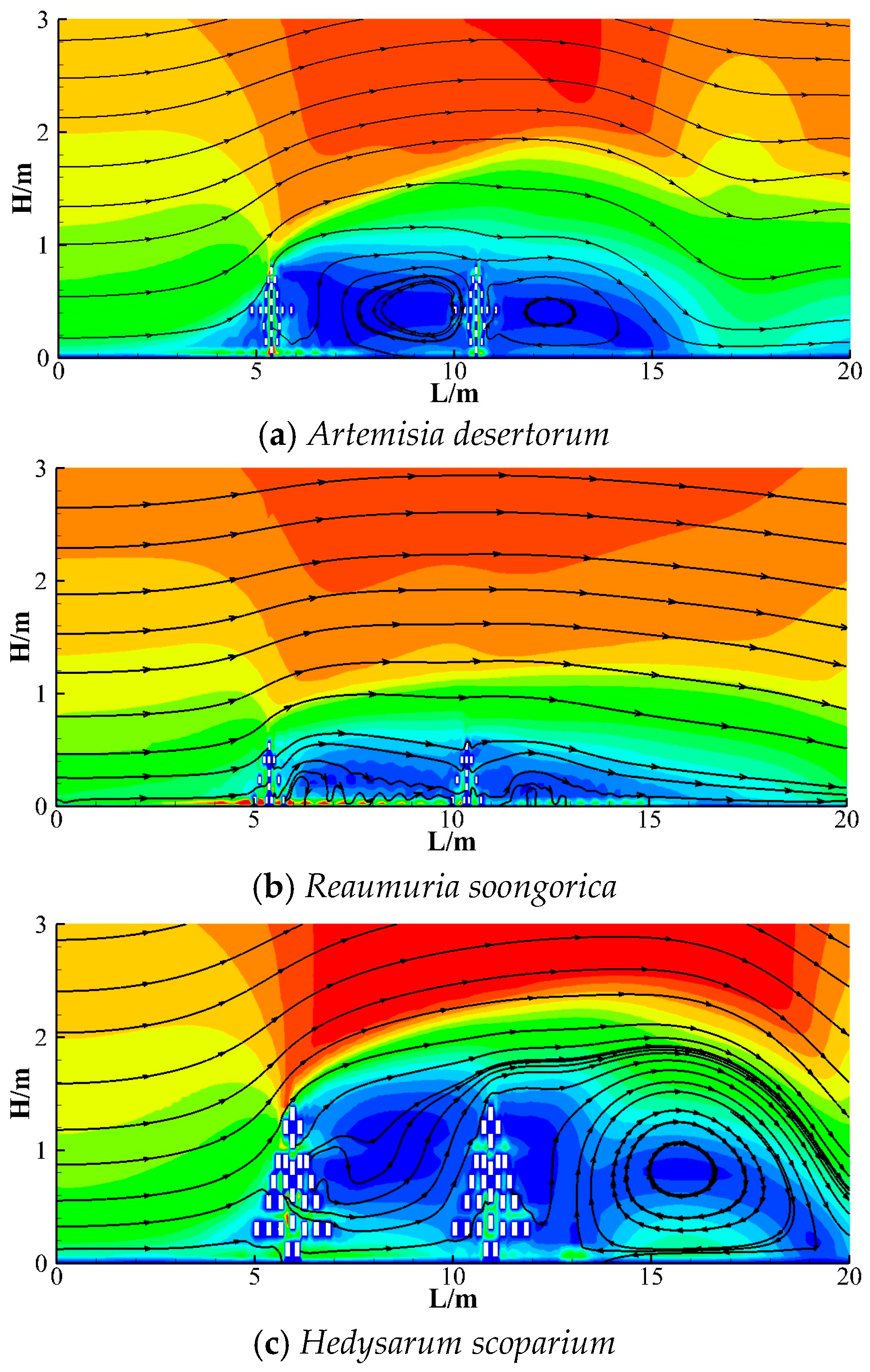
3.2.3. Vortex Analysis After Shrubs
3.2.4. Analysis of Shelter Effect of Different Shrubs
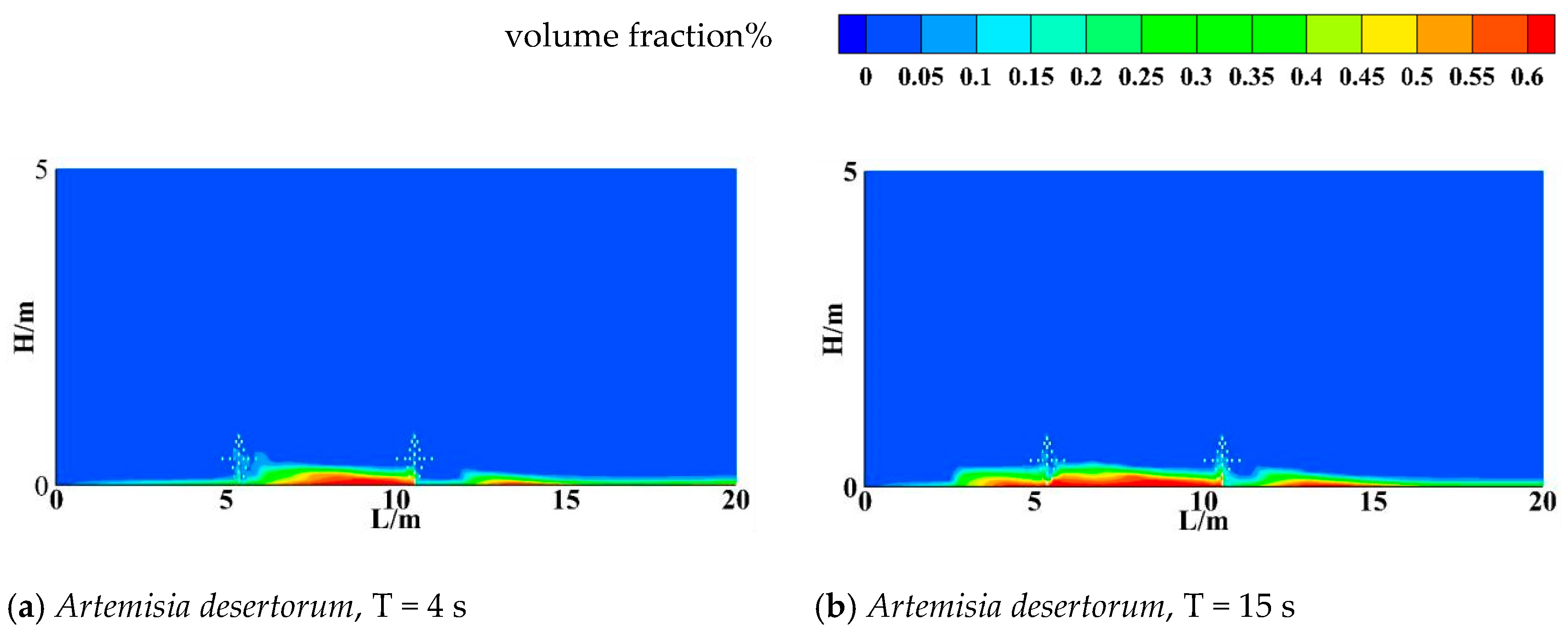
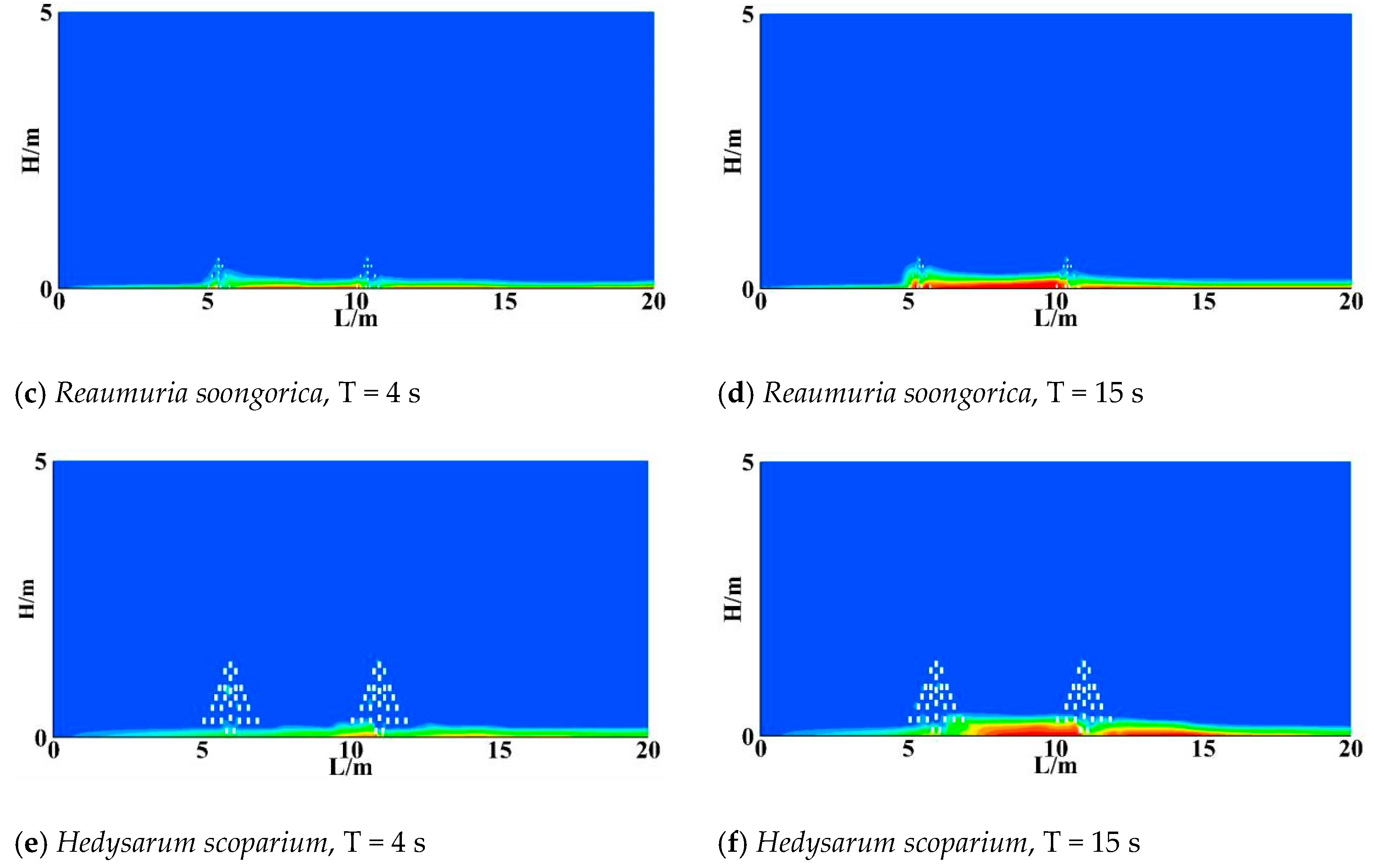
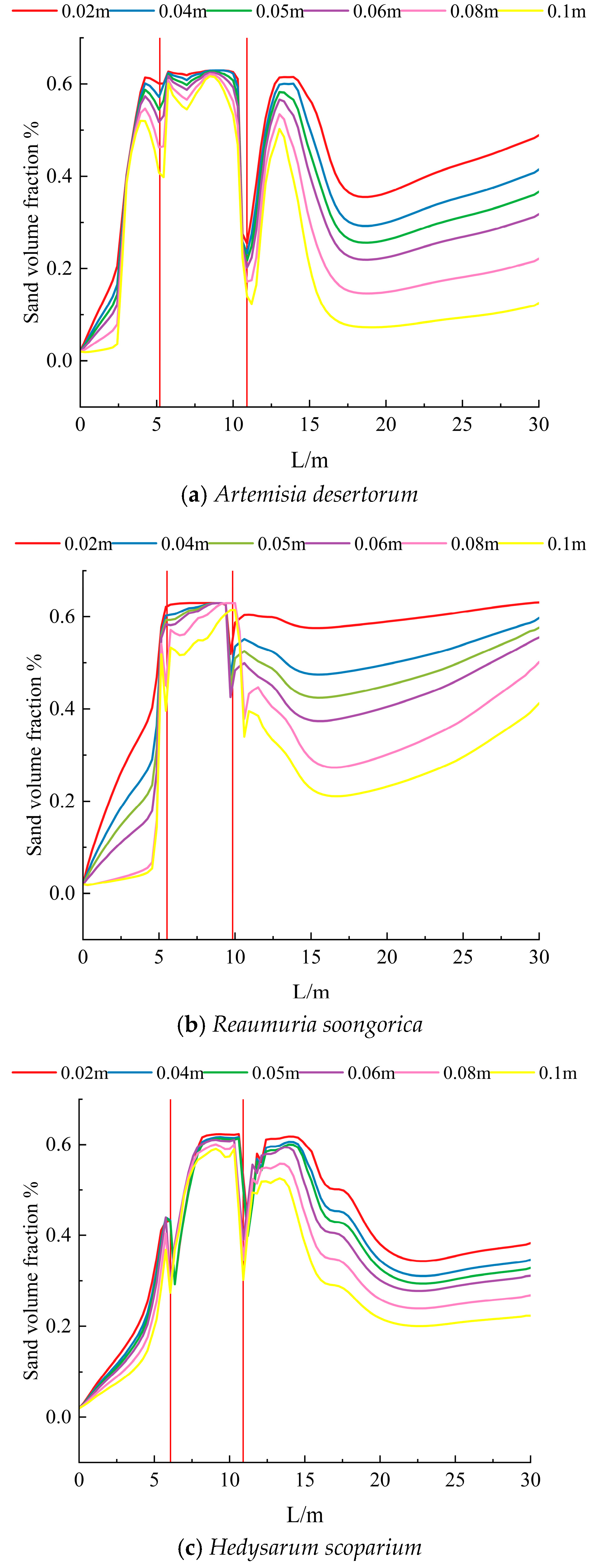
3.3. Sand Resistance of Different Shrubs
4. Discussion
5. Conclusions
- (1)
- The airflow field around the shrubs was divided into five distinct zones: the deceleration zone, acceleration zone, transition zone, vortex zone, and restoration zone. The effective wind speed reduction distances downwind of the three shrub species reached up to 7 H, 6 H, and 6 H, respectively. Fan-shaped deceleration zones were observed upwind of all three shrub species, with the maximum width of the deceleration zone reaching 5 m. The sheltering effect widths of the three shrubs were 3 m, 3 m, and 8 m, respectively.
- (2)
- All three shrub species generated vortices in the wake region behind the vegetation. However, the intensity of these vortices varied depending on differences in plant architecture and canopy size. The effective wind sheltering ranges were as follows: Artemisia desertorum (0.5–4 H), Reaumuria soongorica (0.5–3 H), and Hedysarum scoparium (0.5–2 H). Additionally, Artemisia desertorum generated an intermediate vortex between the two rows of shrubs, extending from 1 H to 4 H behind the first row.
- (3)
- The wind velocity profiles behind the shrubs exhibited a characteristic vertical “V” shape. The maximum reduction in wind speed occurred at the same height stratum as the maximum upwind projected area of the shrubs. The optimal wind resistance heights varied among shrub species: Reaumuria soongorica, Artemisia desertorum, and Hedysarum scoparium achieved the most effective wind sheltering at low (0–0.2 m), medium (0.2–0.7 m), and high (0.7–2.2 m) height layers, respectively.
- (4)
- All three shrub species exhibited wind-driven sand deposition between the two shrub rows. The wind accumulation ranges were 7.5–8.5 m, 9–10 m, and 7–10 m for R. soongorica, A. desertorum, and H. scoparium, respectively. Hedysarum scoparium additionally exhibited aeolian sediment deposition on the leeward side of the shrubs. Overall, the three shrub species reduced wind erosion both upwind and downwind of the windbreak zones. Hedysarum scoparium exhibited the strongest sand resistance, followed by A. desertorum, while Reaumuria soongorica showed the weakest performance.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zheng, X.; Zhu, J.; Xing, Z. Assessment of the effects of shelterbelts on crop yields at the regional scale in Northeast China. Agric. Syst. 2016, 143, 49–60. [Google Scholar] [CrossRef]
- Zhang, K.; Qu, J.; Zhang, X.; Zhao, L.; Li, S. Protective efficiency of railway arbor-shrub windbreak forest belts in Gobi regions: Numerical simulation and wind tunnel tests. Front. Environ. Sci. 2022, 10, 885070. [Google Scholar] [CrossRef]
- Song, C.; Yu, Q.; Wang, R.; Cui, G. Radiating Benefit of Windbreak and Sand Fixation in the Baijitan Nature Reserve of Lingwu, Ningxia, China. Sustainability 2021, 13, 3508. [Google Scholar] [CrossRef]
- Cheng, H.; Liu, C.; Zou, X.; Li, H.; Kang, L.; Liu, B.; Li, J. Wind erosion rate for vegetated soil cover: A prediction model based on surface shear strength. Catena 2020, 187, 104398. [Google Scholar]
- Zhao, Y.; Wu, J.; He, C.; Ding, G. Linking wind erosion to ecosystem services in drylands: A landscape ecological approach. Landsc. Ecol. 2017, 32, 2399–2417. [Google Scholar] [CrossRef]
- San, B.; Wang, Y.; Qiu, Y. Numerical simulation and optimization study of the wind flow through a porous fence. Environ. Fluid Mech. 2018, 18, 1057–1075. [Google Scholar] [CrossRef]
- Wu, Y.; Jin, A.; Jiang, J. Numerical Simulation and Parameter Optimization of a New Slant Insertion-Opening Combination Sand Fence. Sustainability 2024, 16, 8651. [Google Scholar] [CrossRef]
- Huang, H. Modeling the inhibition effect of straw checkerboard barriers on wind-blown sand. Earth Surf. Dyn. 2023, 11, 167–181. [Google Scholar] [CrossRef]
- Okin, G.S. A new model of wind erosion in the presence of vegetation. J. Geophys. Res. Earth Surf. 2008, 113. [Google Scholar] [CrossRef]
- Liu, Y.; Yin, Z.; Yan, Q.; Zhang, C. Numerical simulation of the windbreak and sand-fixation effects of Tamarix chinensis. Arid Zone Res. 2024, 41, 1887–1897. (In Chinese) [Google Scholar]
- Yan, Q.; Li, J.; Yin, Z.; Liu, J.; Liu, H. Numerical simulation of the influence of typical shrub types on wind-sand flow field. Arid Zone Res. 2023, 40, 785–797. [Google Scholar]
- Liu, J.; Li, J.; Yin, Z.; Liu, H. Numerical simulation of the impact of typical plant types of sandy shrubs on wind and sand flow fields. Arid Reg. Res. 2022, 39, 1514–1525. [Google Scholar]
- Huang, S.; Ma, T.; Jiang, F.; Nie, F.; Wang, X.; Ma, T. Numerical simulation and field study on predicting wind-blown sand accumulation in sand mitigation measures of the Ganquan railway. Front. Earth Sci. 2024, 12, 1443030. [Google Scholar] [CrossRef]
- Sun, D.; Dawson, R.; Li, B. Agricultural causes of desertification risk in Minqin, China. J. Environ. Manag. 2006, 79, 348–356. [Google Scholar]
- Chang, Z.; Duan, X.; Han, F.; Zhong, S.; Wang, Q.; Zhang, J. Stability and Ecological Effects of Main Plant Communities in Minqin Desert Area. Anim. Husb. Feed. Sci. 2015, 7, 191. [Google Scholar]
- Liu, H.; Yuan, H.; Wang, D.; Liu, S.; Guo, C.; Ma, R.; Li, X.; Liu, K.; Wan, X.; Li, J. Field observation on windbreak effect of two simulated sand fixing shrubs. Acta Bot. Boreali–Occident. Sin. 2014, 34, 155–159. [Google Scholar]
- Yue, X. Flow Field and Sand Particle Phase Movement Characteristics on the Downwind Side of Windbreak and Sand–Fixing Walls. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2022. [Google Scholar]
- Zhang, X.; Xie, S.; Pang, Y. Numerical simulation on wind-sand flow field around railway embankment with different wind angles. Front. Environ. Sci. 2023, 10, 1073257. [Google Scholar] [CrossRef]
- Lu, W. Research on the Windbreak and Sand Control Benefits of Typical Individual Shrubs in the Ulan Buh Deser. Master’s Thesis, Beijing Forestry University, Beijing, China, 2019. [Google Scholar]
- Morris, H.M., Jr. Flow in rough conduits. Trans. Am. Soc. Civ. Eng. 1955, 120, 373–398. [Google Scholar] [CrossRef]
- Liu, J.; Kimura, R.; Miyawaki, M.; Kinugasa, T. Effects of plants with different shapes and coverage on the blown-sand flux and roughness length examined by wind tunnel experiments. Catena 2021, 197, 104976. [Google Scholar] [CrossRef]
- Wolfe, S.A.; Nickling, W.G. The protective role of sparse vegetation in wind erosion. Prog. Phys. Geogr. 1993, 17, 50–68. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, H.; Wang, W.; Sun, J.; Ma, X. Research Progress on the Correlation between Urban Morphological Indicators and Urban Ventilation at Mesoscopic and Microscopic Scales. Ecol. Sci. 2023, 42, 252–262. [Google Scholar]
- Wei, Z. Analysis and Research on the Movement Laws of Windblown Sand. Soil Water Conserv. China 2012, 5, 44–46. [Google Scholar]
- Wu, X.; Zou, X.; Zhou, N.; Zhang, C.; Shi, S. Deceleration efficiencies of shrub windbreaks in a wind tunnel. Aeolian Res. 2015, 16, 11–23. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Jiang, Z.; Chai, W. Spatial Structure and Windbreak Effects of Main Sand Control and Afforestation Tree Species in Minqin County. Res. Soil Water Conserv. 2008, 3, 121–124. [Google Scholar]
- Lang, L.; Wang, X.; Hasi, E.; Hua, T. Nebkha (coppice dune) formation and significance to environmental change reconstructions in arid and semiarid areas. J. Geogr. Sci. 2013, 23, 344–358. [Google Scholar] [CrossRef]
- Liu, S.; Ha, Z. Prediction of random packing limit for multimodal particle mixtures. Powder Technol. 2002, 126, 283–296. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, Z.; Cheng, H.; Zou, X. Airflow around single and multiple plants. Agric. For. Meteorol. 2018, 252, 27–38. [Google Scholar] [CrossRef]
- Guo, Z.; Yang, X.; Wu, X.; Zou, X.; Zhang, C.; Fang, H.; Xiang, H. Optimal design for vegetative windbreaks using 3D numerical simulations. Agric. For. Meteorol. 2021, 298, 108290. [Google Scholar] [CrossRef]
- Yang, W.; Zhao, A.; Wang, J.; Yao, J.; Tian, Y.; Hu, X.; Yang, H. Study on the Horizontal Configuration Structure and Windbreak and Sand–Fixation Effects of Low–Coverage Artemisia ordosica Clusters. J. Desert Res. 2006, 1, 108–112. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H1/m | C/m | H2/m | S/m | A/m2 | |
|---|---|---|---|---|---|
| A. desertorum | 0.848 | 0.95–1.04 | 0.08 | 0.2~0.4 | 0.045 |
| R. soongorica | 0.553 | 0.83–1.07 | 0.03 | 0~0.1 | 0.033 |
| H. scoparium | 1.417 | 1.95–2.03 | 0.12 | 0.2~0.4 | 0.091 |
| H1/m | C/m | H2/m | S/m | A/m2 | |
|---|---|---|---|---|---|
| A. desertorum | 0.80 | 1.06–1.06 | 0.08 | 0.2~0.4 | 0.042 |
| R. soongorica | 0.57 | 0.76–1.08 | 0.03 | 0~0.1 | 0.039 |
| H. scoparium | 1.38 | 1.80–1.92 | 0.12 | 0.2~0.4 | 0.120 |
| 1 H | 2 H | |||
|---|---|---|---|---|
| Average Wind Speed Reduction % | Maximum Wind Speed Reduction % | Average Wind Speed Reduction % | Maximum Wind Speed Reduction % | |
| A. desertorum | 55.75 | 93.85 | 53.95 | 93.18 |
| R. soongorica | 45.76 | 87.24 | 43.17 | 81.94 |
| H. scoparium | 58.09 | 82.90 | 51.48 | 82.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Pei, L.; Li, J.; Wang, F.; Yin, Z. Numerical Simulation of Wind and Sand Resistance in Three Typical Shrubs. Sustainability 2025, 17, 5481. https://doi.org/10.3390/su17125481
Zhang H, Pei L, Li J, Wang F, Yin Z. Numerical Simulation of Wind and Sand Resistance in Three Typical Shrubs. Sustainability. 2025; 17(12):5481. https://doi.org/10.3390/su17125481
Chicago/Turabian StyleZhang, Huimin, Liang Pei, Juyan Li, Fan Wang, and Zhongdong Yin. 2025. "Numerical Simulation of Wind and Sand Resistance in Three Typical Shrubs" Sustainability 17, no. 12: 5481. https://doi.org/10.3390/su17125481
APA StyleZhang, H., Pei, L., Li, J., Wang, F., & Yin, Z. (2025). Numerical Simulation of Wind and Sand Resistance in Three Typical Shrubs. Sustainability, 17(12), 5481. https://doi.org/10.3390/su17125481