
Methods for Assessing the Ecosystem Service of Honey Provisioning by the European Honey Bee (Apis mellifera L.): A Systematic Review


Abstract
1. Introduction
- What kind of models and methodologies are used in the literature to map or assess the HPCs of specific sites, landscapes or regions?
- What are the main methodological choices underlying the different models? How are these choices influenced by the availability of information (input data)?
- How do these choices influence the model outputs (the resulting HPC indicator)?
- How do the different approaches identify, describe and resolve uncertainties?
- How well are the different options aligned with the best practices and recommendations for ecosystem-service (ES) assessment studies?
2. Materials and Methods
2.1. Literature Search
2.2. Selection Criteria
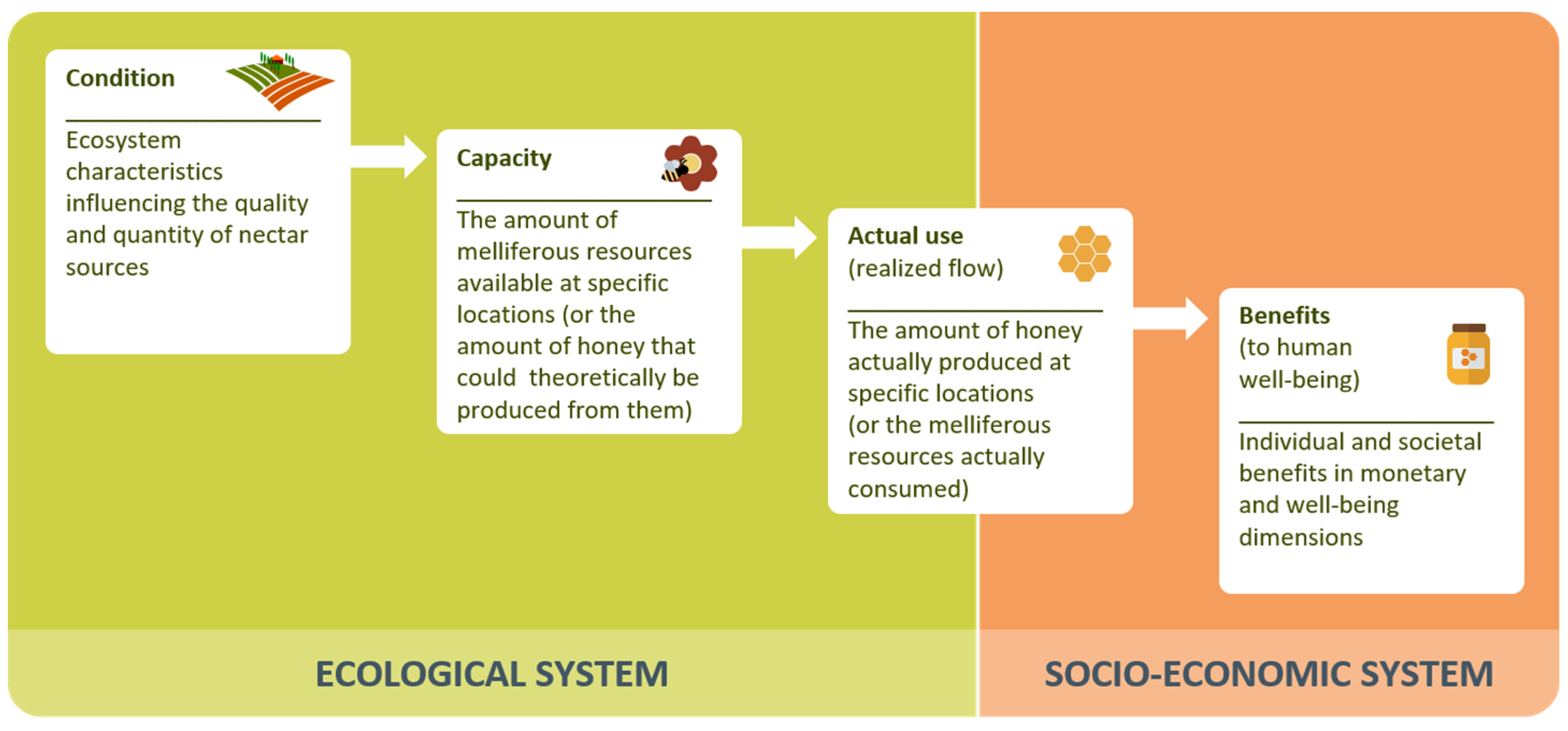
- The indicator characterizes the capacity of ecosystems to provide honey (level 1 or 2 of the ES cascade), quantifying one of the following:
- the potential supply (estimated multiannual mean or maximum ‘flow’) of honey or its main ‘ingredients’ (i.e., nectar or honeydew),
- the availability of melliferous resources (flowers, specific plant species or ecosystems) as ’proxy’ indicators for nectar potential, or
- the overall suitability of spatial locations for beekeeping.
- The study/indicator either focuses specifically on European honey bees (A. mellifera) or describes the landscape (availability of resources) from the perspective of a group of pollinators that explicitly includes honey bees.
- The study makes spatial predictions (to evaluate and compare different sites) in a specific case-study area based on the model/indicator.
- 4.
- Exclusion of ‘pseudo-replications’: if two or more studies by the same authors presented the same model in different study areas, only one study was kept.

2.3. Data Extraction and Analysis
- be spatially explicit (predicted values are assigned to concrete locations, describing the spatial variability of honey pastures)
- characterize the capacity of each location to provide honey, regardless of whether this capacity was eventually harvested (turned into “actual use”) or not.
3. Results
3.1. Model Types
3.2. Model Assumptions
3.3. Model Input Data
4. Discussion
4.1. Discussion of the Findings of the Systematic Review
4.1.1. Key Decision Points in the Modelling Process
4.1.2. Methodological Uncertainties
4.1.3. Applicability of HPC Model Outputs in Apicultural Practice
4.2. Reflections About the Methodology Used for the Systematic Review
5. Conclusions
5.1. Clear Definitions
5.2. Clear Methodology
- -
- spatial variability in nectar sources
- -
- temporal dynamics of nectar flow, including seasonal fluctuations within growing seasons, with a special emphasis on the dynamics of nectar shortage and flowering peak periods
- -
- effects of climatic conditions on nectar provision
5.3. Involvement of Beekeepers
5.4. Impacts of Land Use on Pollinator Health
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ES | ecosystem services |
| ET | ecosystem type |
| HPC | honey provisioning capacity |
References
- Daily, G.C. Nature’s Services: Societal Dependence on Natural Ecosystems (1997). In The Future of Nature; Robin, L., Sörlin, S., Warde, P., Eds.; Yale University Press: New Haven, CT, USA, 2017; pp. 454–464. ISBN 978-0-300-18847-9. [Google Scholar]
- Reid, W.V.; Mooney, H.A.; Cropper, A.; Capistrano, D.; Carpenter, S.R.; Chopra, K.; Dasgupta, P.; Dietz, T.; Duraiappah, A.K.; Hassan, R.; et al. Ecosystems and Human Well-Being: Synthesis; The Millennium Ecosystem Assessment Series; Island Press: Washington, DC, USA, 2005; ISBN 978-1-59726-040-4. [Google Scholar]
- Díaz, S.; Demissew, S.; Carabias, J.; Joly, C.; Lonsdale, M.; Ash, N.; Larigauderie, A.; Adhikari, J.R.; Arico, S.; Báldi, A.; et al. The IPBES Conceptual Framework—Connecting Nature and People. Curr. Opin. Environ. Sustain. 2015, 14, 1–16. [Google Scholar] [CrossRef]
- Papa, G.; Maier, R.; Durazzo, A.; Lucarini, M.; Karabagias, I.K.; Plutino, M.; Bianchetto, E.; Aromolo, R.; Pignatti, G.; Ambrogio, A.; et al. The Honey Bee Apis mellifera: An Insect at the Interface between Human and Ecosystem Health. Biology 2022, 11, 233. [Google Scholar] [CrossRef] [PubMed]
- Aizen, M.A.; Harder, L.D. The Global Stock of Domesticated Honey Bees Is Growing Slower Than Agricultural Demand for Pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef] [PubMed]
- Brittain, C.; Potts, S.G. The Potential Impacts of Insecticides on the Life-History Traits of Bees and the Consequences for Pollination. Basic Appl. Ecol. 2011, 12, 321–331. [Google Scholar] [CrossRef]
- IPBES. The Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on Pollinators, Pollination and Food Production; Potts, S.G., Imperatriz-Fonseca, V.L., Ngo, H.T., Eds.; Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2016. [Google Scholar] [CrossRef]
- Haines-Young, R.; Potschin, M.B. Common International Classification of Ecosystem Services (CICES) V5.2 and Guidance on the Application of the Revised Structure. Available online: https://cices.eu/ (accessed on 30 January 2025).
- Ausseil, A.E.; Dymond, J.R.; Newstrom, L. Mapping Floral Resources for Honey Bees in New Zealand at the Catchment Scale. Ecol. Appl. 2018, 28, 1182–1196. [Google Scholar] [CrossRef]
- Lehébel-Péron, A.; Sidawy, P.; Dounias, E.; Schatz, B. Attuning Local and Scientific Knowledge in the Context of Global Change: The Case of Heather Honey Production in Southern France. J. Rural Stud. 2016, 44, 132–142. [Google Scholar] [CrossRef]
- Keresztesi, B. Breeding and Cultivation of Black Locust, Robinia Pseudoacacia, in Hungary. For. Ecol. Manag. 1983, 6, 217–244. [Google Scholar] [CrossRef]
- Southwood, T.R.E. Entomology and Mankind: Insects over the Ages Have Greatly Affected Man’s Health and Food Supply and Have Played an Important Role as Religious and Cultural Symbols. Am. Sci. 1977, 65, 30–39. [Google Scholar]
- Seeley, T.D. The Lives of Bees: The Untold Story of the Honey Bee in the Wild; Princeton University Press: Princeton, NJ, USA, 2019; ISBN 978-0-691-16676-6. [Google Scholar]
- Klein, A.-M.; Boreux, V.; Fornoff, F.; Mupepele, A.-C.; Pufal, G. Relevance of Wild and Managed Bees for Human Well-Being. Curr. Opin. Insect Sci. 2018, 26, 82–88. [Google Scholar] [CrossRef]
- Potschin, M.B.; Haines-Young, R.H. Ecosystem Services: Exploring a Geographical Perspective. Prog. Phys. Geogr. Earth Environ. 2011, 35, 575–594. [Google Scholar] [CrossRef]
- Heink, U.; Jax, K. Going Upstream—How the Purpose of a Conceptual Framework for Ecosystem Services Determines Its Structure. Ecol. Econ. 2019, 156, 264–271. [Google Scholar] [CrossRef]
- Czúcz, B.; Haines-Young, R.; Kiss, M.; Bereczki, K.; Kertész, M.; Vári, Á.; Potschin-Young, M.; Arany, I. Ecosystem Service Indicators along the Cascade: How Do Assessment and Mapping Studies Position Their Indicators? Ecol. Indic. 2020, 118, 106729. [Google Scholar] [CrossRef]
- Wright, G.A.; Nicolson, S.W.; Shafir, S. Nutritional Physiology and Ecology of Honey Bees. Annu. Rev. Entomol. 2018, 63, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Munro, J.A. A Study of the Factors Influencing the Yield of Honey during the Main Nectar Flow. J. Econ. Entomol. 1929, 22, 518–521. [Google Scholar] [CrossRef]
- Wielgolaski, F.E. Phenological Modifications in Plants by Various Edaphic Factors. Int. J. Biometeorol. 2001, 45, 196–202. [Google Scholar] [CrossRef]
- Dahlgren, J.P.; Von Zeipel, H.; Ehrlén, J. Variation in Vegetative and Flowering Phenology in a Forest Herb Caused by Environmental Heterogeneity. Am. J. Bot. 2007, 94, 1570–1576. [Google Scholar] [CrossRef]
- Holmes, W. The Influence of Weather on Annual Yields of Honey. J. Agric. Sci. 2002, 139, 95–102. [Google Scholar] [CrossRef]
- Corbet, S.A.; Willmer, P.G.; Beament, J.W.L.; Unwin, D.M.; Prŷs-Jones, O.E. Post-secretory Determinants of Sugar Concentration in Nectar. Plant Cell Environ. 1979, 2, 293–308. [Google Scholar] [CrossRef]
- Park, W. The Storing and Ripening of Honey by Honeybees. J. Econ. Entomol. 1925, 18, 405–410. [Google Scholar] [CrossRef]
- Lee, W.R. Relation of Distance to Foraging Intensity of Honey Bees (Apis mellifera) on Natural Food Sources1. Ann. Entomol. Soc. Am. 1965, 58, 94–100. [Google Scholar] [CrossRef]
- Robinson, S.V.J.; Hoover, S.E.; Pernal, S.F.; Cartar, R.V. Optimal Distributions of Central-Place Foragers: Honey Bee Foraging in a Mass Flowering Crop. Behav. Ecol. 2022, 33, 386–397. [Google Scholar] [CrossRef]
- Couvillon, M.J.; Riddell Pearce, F.C.; Accleton, C.; Fensome, K.A.; Quah, S.K.L.; Taylor, E.L.; Ratnieks, F.L.W. Honey Bee Foraging Distance Depends on Month and Forage Type. Apidologie 2015, 46, 61–70. [Google Scholar] [CrossRef]
- Schmickl, T.; Karsai, I. How Regulation Based on a Common Stomach Leads to Economic Optimization of Honeybee Foraging. J. Theor. Biol. 2016, 389, 274–286. [Google Scholar] [CrossRef]
- Örösi, P.Z. Méhek Között; Mezőgazdasági Kiadó: Budapest, Hungary, 1955. [Google Scholar]
- Ludányi, I.; Csaba, G. Méhész Leszek; Herman Ottó Intézet Nonprofit Kft.: Budapest, Hungary, 2020; Volume 1, ISBN 978-963-309-121-0. [Google Scholar]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.; et al. Country-Specific Effects of Neonicotinoid Pesticides on Honey Bees and Wild Bees. Science 2017, 356, 1393–1395. [Google Scholar] [CrossRef]
- Molan, P.C. Authenticity of Honey. In Food Authentication; Ashurst, P.R., Dennis, M.J., Eds.; Springer: Boston, MA, USA, 1996; pp. 259–303. ISBN 978-1-4612-8426-0. [Google Scholar]
- La Notte, A. Ecologically Intermediate and Economically Final: The Role of the Ecosystem Services Framework in Measuring Sustainability in Agri-Food Systems. Land 2022, 11, 84. [Google Scholar] [CrossRef]
- Decourtye, A.; Mader, E.; Desneux, N. Landscape Enhancement of Floral Resources for Honey Bees in Agro-Ecosystems. Apidologie 2010, 41, 264–277. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Tooker, J.F.; Grozinger, C.M.; Patch, H.M. Bee Nutrition and Floral Resource Restoration. Curr. Opin. Insect Sci. 2015, 10, 133–141. [Google Scholar] [CrossRef]
- Halmágyi, L.; Keresztesi, B.; Bujtás, Z. A Méhlegelő, 2nd ed.; Akadémiai Kiadó: Budapest, Hungary, 1991; ISBN 978-963-05-6017-7. [Google Scholar]
- Kovács-Hostyánszki, A.; Szigeti, V.; Miholcsa, Z.; Sándor, D.; Soltész, Z.; Török, E.; Fenesi, A. Threats and Benefits of Invasive Alien Plant Species on Pollinators. Basic Appl. Ecol. 2022, 64, 89–102. [Google Scholar] [CrossRef]
- Rollin, O.; Bretagnolle, V.; Decourtye, A.; Aptel, J.; Michel, N.; Vaissière, B.E.; Henry, M. Differences of Floral Resource Use between Honey Bees and Wild Bees in an Intensive Farming System. Agric. Ecosyst. Environ. 2013, 179, 78–86. [Google Scholar] [CrossRef]
- Requier, F.; Odoux, J.-F.; Tamic, T.; Moreau, N.; Henry, M.; Decourtye, A.; Bretagnolle, V. Honey Bee Diet in Intensive Farmland Habitats Reveals an Unexpectedly High Flower Richness and a Major Role of Weeds. Ecol. Appl. 2015, 25, 881–890. [Google Scholar] [CrossRef]
- Garbuzov, M.; Balfour, N.J.; Shackleton, K.; Al Toufailia, H.; Scandian, L.; Ratnieks, F.L.W. Multiple Methods of Assessing Nectar Foraging Conditions Indicate Peak Foraging Difficulty in Late Season. Insect Conserv. Divers. 2020, 13, 532–542. [Google Scholar] [CrossRef]
- Pilati, L.; Daris, R.; Prestamburgo, M.; Sgroi, F. Modeling Sequential Production: The Migratory Beekeeper Case. Qual. Access Success 2018, 19, 146–154. [Google Scholar]
- Crane, E. A Short History of Knowledge about Honey Bees (Apis) up to 1800. Bee World 2004, 85, 6–11. [Google Scholar] [CrossRef]
- Phillips, C. Telling Times: More-than-Human Temporalities in Beekeeping. Geoforum 2020, 108, 315–324. [Google Scholar] [CrossRef]
- Maderson, S. There’s More Than One Way To Know A Bee: Beekeepers’ Environmental Knowledge, and Its Potential Role in Governing for Sustainability. Geoforum 2023, 139, 103690. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Dixon-Woods, M.; Bonas, S.; Booth, A.; Jones, D.R.; Miller, T.; Sutton, A.J.; Shaw, R.L.; Smith, J.A.; Young, B. How Can Systematic Reviews Incorporate Qualitative Research? A Critical Perspective. Qual. Res. 2006, 6, 27–44. [Google Scholar] [CrossRef]
- Dixon-Woods, M.; Cavers, D.; Agarwal, S.; Annandale, E.; Arthur, A.; Harvey, J.; Hsu, R.; Katbamna, S.; Olsen, R.; Smith, L.; et al. Conducting a Critical Interpretive Synthesis of the Literature on Access to Healthcare by Vulnerable Groups. BMC Med. Res. Methodol. 2006, 6, 35. [Google Scholar] [CrossRef]
- Estoque, R.C.; Murayama, Y. Suitability Analysis for Beekeeping Sites in La Union, Philippines, Using GIS and Multi-Criteria Evaluation Techniques. Res. J. Appl. Sci. 2010, 5, 242–253. [Google Scholar] [CrossRef]
- Maris, N.M.N.; Mansor, S.; Shafri, H.Z.M. Apicultural Site Zonation Using GIS and Multi-Criteria Decision Analysis. Pertanika J. Trop. Agric. Sci. 2008, 31, 147–162. [Google Scholar]
- Smith, D.; Davis, A.; Hitaj, C.; Hellerstein, D.; Preslicka, A.; Kogge, E.; Mushet, D.; Lonsdorf, E. The Contribution of Land Cover Change to the Decline of Honey Yields in the Northern Great Plains. Environ. Res. Lett. 2021, 16, 064050. [Google Scholar] [CrossRef]
- ISO 3166-1:2020(en); Codes for the Representation of Names of Countries and Their Subdivisions. International Organization for Standardization: Geneva, Switzerland, 2020.
- Arany, I.; Vari, A.; Aszalos, R.; Kelemen, K.; Kelemen, M.A.; Bone, G.; Lellei-Kovacz, E.; Czusz, B. Diversity of Flower-Rich Habitats as a Persistent Source of Healthy Diet for Honey Bees. Eur. J. Geogr. 2019, 10, 89–106. [Google Scholar]
- Tredennick, A.T.; Hooker, G.; Ellner, S.P.; Adler, P.B. A Practical Guide to Selecting Models for Exploration, Inference, and Prediction in Ecology. Ecology 2021, 102, e03336. [Google Scholar] [CrossRef] [PubMed]
- Affek, A.N. Indicators of Ecosystem Potential for Pollination and Honey Production. Ecol. Indic. 2018, 94, 33–45. [Google Scholar] [CrossRef]
- Baude, M.; Kunin, W.E.; Boatman, N.D.; Conyers, S.; Davies, N.; Gillespie, M.A.K.; Morton, R.D.; Smart, S.M.; Memmott, J. Historical Nectar Assessment Reveals the Fall and Rise of Floral Resources in Britain. Nature 2016, 530, 85–88. [Google Scholar] [CrossRef]
- Balzan, M.V.; Caruana, J.; Zammit, A. Assessing the Capacity and Flow of Ecosystem Services in Multifunctional Landscapes: Evidence of a Rural-Urban Gradient in a Mediterranean Small Island State. Land Use Policy 2018, 75, 711–725. [Google Scholar] [CrossRef]
- Sari, F.; Kandemir, İ.; Ceylan, D.A.; Gül, A. Using AHP and PROMETHEE Multi-Criteria Decision Making Methods to Define Suitable Apiary Locations. J. Apic. Res. 2020, 59, 546–557. [Google Scholar] [CrossRef]
- Gallant, A.L.; Euliss, N.H.; Browning, Z. Mapping Large-Area Landscape Suitability for Honey Bees to Assess the Influence of Land-Use Change on Sustainability of National Pollination Services. PLoS ONE 2014, 9, e99268. [Google Scholar] [CrossRef]
- Otto, C.R.V.; Zheng, H.; Gallant, A.L.; Iovanna, R.; Carlson, B.L.; Smart, M.D.; Hyberg, S. Past Role and Future Outlook of the Conservation Reserve Program for Supporting Honey Bees in the Great Plains. Proc. Natl. Acad. Sci. USA 2018, 115, 7629–7634. [Google Scholar] [CrossRef]
- Norton, L.R.; Smart, S.M.; Maskell, L.C.; Henrys, P.A.; Wood, C.M.; Keith, A.M.; Emmett, B.A.; Cosby, B.J.; Thomas, A.; Scholefield, P.A.; et al. Identifying Effective Approaches for Monitoring National Natural Capital for Policy Use. Ecosyst. Serv. 2018, 30, 98–106. [Google Scholar] [CrossRef]
- Becher, M.A.; Grimm, V.; Thorbek, P.; Horn, J.; Kennedy, P.J.; Osborne, J.L. BEEHAVE: A Systems Model of Honeybee Colony Dynamics and Foraging to Explore Multifactorial Causes of Colony Failure. J. Appl. Ecol. 2014, 51, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Horn, J.; Becher, M.A.; Kennedy, P.J.; Osborne, J.L.; Grimm, V. Multiple Stressors: Using the Honeybee Model BEEHAVE to Explore How Spatial and Temporal Forage Stress Affects Colony Resilience. Oikos 2016, 125, 1001–1016. [Google Scholar] [CrossRef]
- Becher, M.A.; Grimm, V.; Knapp, J.; Horn, J.; Twiston-Davies, G.; Osborne, J.L. BEESCOUT: A Model of Bee Scouting Behaviour and a Software Tool for Characterizing Nectar/Pollen Landscapes for BEEHAVE. Ecol. Model. 2016, 340, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Amiri, F.; Mohamed Shariff, A.R.B.; Arekhi, S. An Approach for Rangeland Suitability Analysis to Apiculture Planning in Gharah Aghach Region, Isfahan-Iran. World Appl. Sci. J. 2011, 12, 962–972. [Google Scholar]
- Zoccali, P.; Malacrinò, A.; Campolo, O.; Laudani, F.; Algeri, G.M.; Giunti, G.; Strano, C.P.; Benelli, G.; Palmeri, V. A Novel GIS-Based Approach to Assess Beekeeping Suitability of Mediterranean Lands. Saudi J. Biol. Sci. 2017, 24, 1045–1050. [Google Scholar] [CrossRef]
- Campana, C.; Peralta, C.; Cecconello, J.C.; Pons, D.H.; Uranga, J.; Scavuzzo, M.C.; Ferral, A. Geospatial Tools Applied to the Generation of an Aptitude Map for the Development of Beekeeping Activity in San Javier, Córdoba, Argentina. In Proceedings of the 2019 XVIII Workshop on Information Processing and Control (RPIC), Bahía Blanca, Argentina, 18–20 September 2019; pp. 298–302. [Google Scholar]
- Vanbergen, A.J. The Insect Pollinators Initiative Threats to an Ecosystem Service: Pressures on Pollinators. Front. Ecol. Environ. 2013, 11, 251–259. [Google Scholar] [CrossRef]
- Pettis, J.S.; Lichtenberg, E.M.; Andree, M.; Stitzinger, J.; Rose, R.; van Engelsdorp, D. Crop Pollination Exposes Honey Bees to Pesticides Which Alters Their Susceptibility to the Gut Pathogen Nosema ceranae. PLoS ONE 2013, 8, e70182. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Goulson, D.; Pennacchio, F.; Nazzi, F.; Goka, K.; Desneux, N. Are Bee Diseases Linked to Pesticides?—A Brief Review. Environ. Int. 2016, 89–90, 7–11. [Google Scholar] [CrossRef]
- Greenhalgh, T.; Peacock, R. Effectiveness and Efficiency of Search Methods in Systematic Reviews of Complex Evidence: Audit of Primary Sources. BMJ 2005, 331, 1064–1065. [Google Scholar] [CrossRef]
- Saunders, B.; Sim, J.; Kingstone, T.; Baker, S.; Waterfield, J.; Bartlam, B.; Burroughs, H.; Jinks, C. Saturation in Qualitative Research: Exploring Its Conceptualization and Operationalization. Qual. Quant. 2018, 52, 1893–1907. [Google Scholar] [CrossRef]
- Davis, L.F.; Ramírez-Andreotta, M.D. Participatory Research for Environmental Justice: A Critical Interpretive Synthesis. Environ. Health Perspect. 2021, 129, 026001. [Google Scholar] [CrossRef] [PubMed]
- Hemmelgarn, H.L.; Munsell, J.F. Exploring ‘beyond-Food’ Opportunities for Biocultural Conservation in Urban Forest Gardens. Urban Agric. Reg. Food Syst. 2021, 6, e20009. [Google Scholar] [CrossRef]
- Perevochtchikova, M.; Castro-Díaz, R.; Langle-Flores, A.; Von Thaden Ugalde, J.J. A Systematic Review of Scientific Publications on the Effects of Payments for Ecosystem Services in Latin America, 2000–2020. Ecosyst. Serv. 2021, 49, 101270. [Google Scholar] [CrossRef]
- House, J.; Kleiber, D.; Steenbergen, D.J.; Stacey, N. Participatory Monitoring in Community-Based Fisheries Management through a Gender Lens. Ambio 2023, 52, 300–318. [Google Scholar] [CrossRef]
- Himes, A.; Muraca, B.; Anderson, C.B.; Athayde, S.; Beery, T.; Cantú-Fernández, M.; González-Jiménez, D.; Gould, R.K.; Hejnowicz, A.P.; Kenter, J.; et al. Why Nature Matters: A Systematic Review of Intrinsic, Instrumental, and Relational Values. BioScience 2024, 74, 25–43. [Google Scholar] [CrossRef]
- Muller, J.; McEwan, K.; Gorczynski, P.; Weston, N. Defining Contemporary Outdoor Physical Activity: A Critical Interpretive Synthesis. J. Outdoor Recreat. Tour. 2024, 47, 100799. [Google Scholar] [CrossRef]
- Macura, B.; Suškevičs, M.; Garside, R.; Hannes, K.; Rees, R.; Rodela, R. Systematic Reviews of Qualitative Evidence for Environmental Policy and Management: An Overview of Different Methodological Options. Environ. Evid. 2019, 8, 24. [Google Scholar] [CrossRef]
- Chen, H.; Sloggy, M.R.; Dhiaulhaq, A.; Escobedo, F.J.; Rasheed, A.R.; Sánchez, J.J.; Yang, W.; Yu, F.; Meng, Z. Boundary of Ecosystem Services: Guiding Future Development and Application of the Ecosystem Service Concepts. J. Environ. Manag. 2023, 344, 118752. [Google Scholar] [CrossRef]
- Schutter, M.S.; Hicks, C.C. Speaking across Boundaries to Explore the Potential for Interdisciplinarity in Ecosystem Services Knowledge Production. Conserv. Biol. 2021, 35, 1198–1209. [Google Scholar] [CrossRef]
- Steger, C.; Hirsch, S.; Evers, C.; Branoff, B.; Petrova, M.; Nielsen-Pincus, M.; Wardropper, C.; Van Riper, C.J. Ecosystem Services as Boundary Objects for Transdisciplinary Collaboration. Ecol. Econ. 2018, 143, 153–160. [Google Scholar] [CrossRef]
- Bagstad, K.J.; Balbi, S.; Adamo, G.; Athanasiadis, I.N.; Affinito, F.; Willcock, S.; Magrach, A.; Hayashi, K.; Harmáčková, Z.V.; Niamir, A.; et al. Interoperability for Ecosystem Service Assessments: Why, How, Who, and for Whom? Ecosyst. Serv. 2025, 72, 101705. [Google Scholar] [CrossRef]
- Polasky, S.; Tallis, H.; Reyers, B. Setting the Bar: Standards for Ecosystem Services. Proc. Natl. Acad. Sci. USA 2015, 112, 7356–7361. [Google Scholar] [CrossRef] [PubMed]
- Bistline, J.; Budolfson, M.; Francis, B. Deepening Transparency about Value-Laden Assumptions in Energy and Environmental Modelling: Improving Best Practices for Both Modellers and Non-Modellers. Clim. Policy 2021, 21, 1–15. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A: Case Study Area | ||
| A1 | Country (The country of the study)
| |
| A2 | Spatial scale (The spatial extent of the study area expressed in size classes)
| |
| A2.1 | Spatial location (Geographic location of the study area)
| |
| A2.2 | Spatial extent (Size of the study area in km2)
| |
| B: Model Assumptions | ||
| B1 | Human inputs (Are model assumptions related to the colonies’ own honey consumption, and is supplementary feeding declared/discussed?)
| |
| B2 | Interannual fluctuations (Are such fluctuations considered? If yes, how?)
| |
| B3 | Seasonal fluctuations (Are such fluctuations considered? If yes, how?)
| |
| B4 | Foraging behaviour (Are the foraging ranges/movements of the bees considered? If yes, how?)
| |
| B5 | Foraging distance (Radius of the foraging ranges used in the model in km)
| |
| B6 | Floral resource types (Which floral resource types (pollen, nectar, honeydew) are considered/distinguished?)
| |
| C: Model Input Data | ||
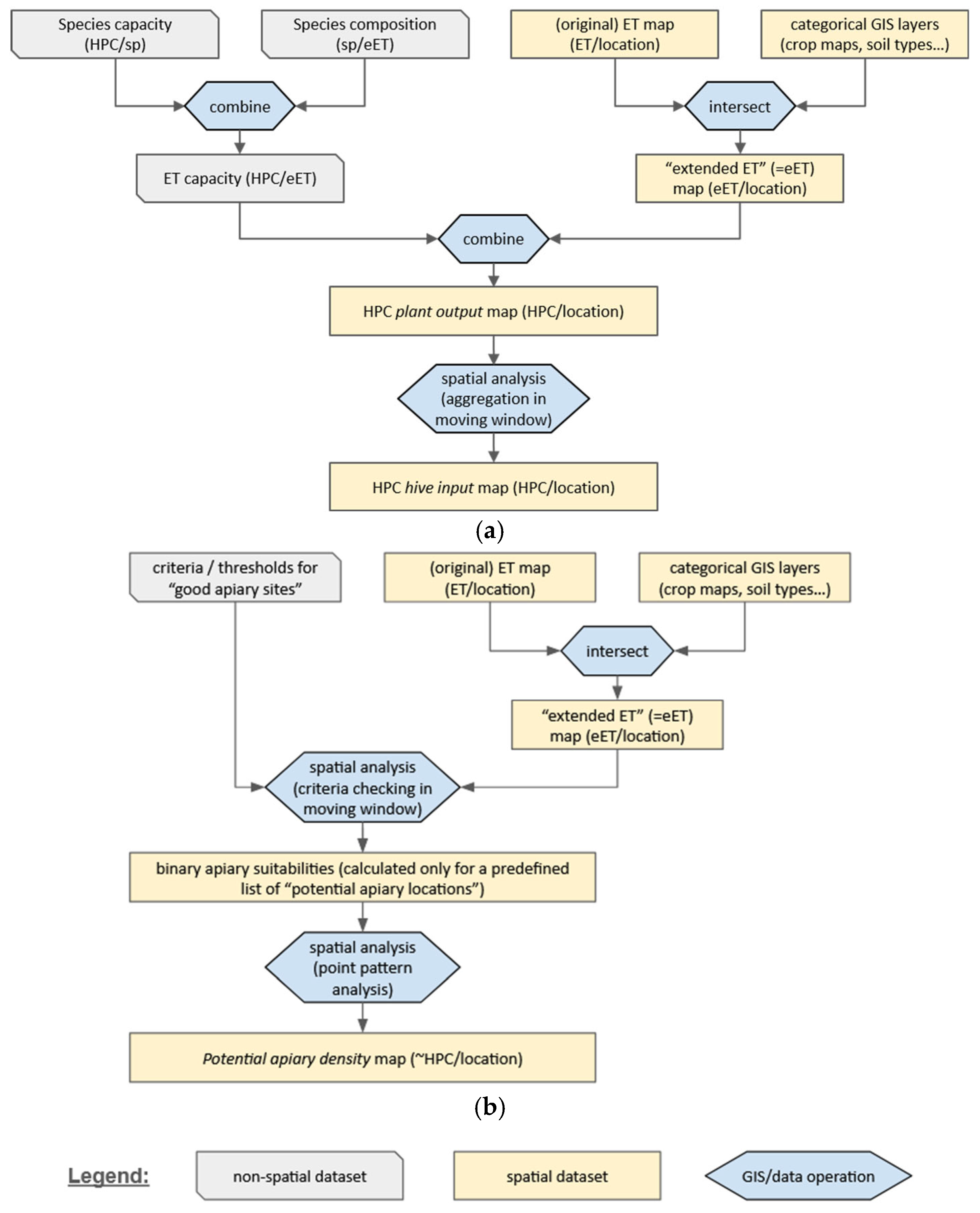
| C1 | Spatial data (Spatially explicit input datasets, multiple selection) et: ecosystem type (ET), including land use, land cover, habitat type, or other categorical variable assigned to each location reflecting the distinct combinations of abiotic and biotic environmental parameters hs: spatial distribution of honey species (based on species level occurrence data) cr: spatial distribution of agricultural crops (assigning the dominant crop species to each location within agricultural ETs) tr: spatial distribution of trees (assigning the dominant tree species to each location, typically only within forested ETs) vc: vegetation cover (of any vegetation layer/canopy level, typically in %); so: soil types (typically categorical) or other soil variables el: elevation: altitude above sea level (possibly as categories) dw: distance to surface water (water availability) dr: distance to roads (accessibility) dc: distance to urban zones ct: temperature (any annual or seasonal, mean or extreme climatic value) cp: precipitation (any annual or seasonal, mean or extreme climatic value) | |
| C2 | Ecosystem type-level data (Non-spatial input data assigned to ETs, multiple selection)
| |
| C3 | Species-level characteristics (Non-spatial input data assigned to plant species, multiple selection) mo: original biophysical measurements ml: values/measurements from literature sc: estimated scores | |
| D: Model Output (HPC Indicator) | ||
| D1 | Indicator name (The name of the HPC indicator (model output) as used in the original study)
| |
| D2 | Indicator type (What kind of units is the final HPC indicator expressed in?) NH: nectar or honey yield (expressed in mass/area/time “flow” units) FP: density of relevant plant species XX: dimensionless score | |
| D2.1 | Measurement scale (A concise textual description of the measurement scale/units)
| |
| D3 | Validation (Were the model outputs validated in some way? If yes, how?)
| |
| E: Model Type | ||
| MT | Model types (The model type identified in this study, synthetic construct)
| |
| Variables Considered | No. of HPC Models |
|---|---|
| B1 Human Inputs | |
| X: no assumptions declared | 13 |
| B: the bees’ own consumption considered | 1 |
| H: assumptions about human inputs | 2 |
| U: unclear | 2 |
| B2 Interannual Fluctuations | |
| O: optimal year assumed | 3 |
| X: question avoided | 15 |
| B3 Seasonal Fluctuations | |
| A: aggregated for the whole year | 15 |
| S: honey seasons are distinguished | 3 |
| B4 Foraging Behavior | |
| 0: movements not considered | 14 |
| 1: foraging ranges considered | 4 |
| B6 Floral Resource Types | |
| S: separate values for nectar and pollen | 4 |
| J: a single joint value | 6 |
| F: flat rate; melliferous species are counted | 8 |
| Data Incorporated in the HPC Model | No. of HPC Models |
|---|---|
| Spatially Explicit Data Sets | |
| et: ecosystem type | 18 |
| hs: honey species locations | 1 |
| cr: agricultural crop map | 2 |
| tr: forest/tree map | 1 |
| vc: vegetation cover | 1 |
| so: soil variables | 3 |
| el: elevation | 7 |
| dw: distance to water | 7 |
| dr: distance to roads | 7 |
| dc: distance to settlements | 2 |
| ct: temperature | 3 |
| cp: precipitation | 3 |
| ET-Level Data | |
| hp: floristic composition | 7 |
| su: successional stage classes | 1 |
| sl: length of flowering season | 1 |
| Species-Level Data | |
| mo: original measurements | 1 |
| ml: literature data | 2 |
| sc: estimated scores | 4 |
| Source of Uncertainty | Suggestion to Manage Uncertainty |
|---|---|
| Variation in species composition within ETs | Make direct use of species-distribution data Refine the species composition of ETs using species-distribution models |
| The nectar yield of certain plant species is unknown | Carry out a biophysical estimate of nectar yield based on field measurements in the sample area |
| Uncertainty of the sugar content of nectar (honey/nectar ratio) | Carefully select the honey/nectar ratio and accurately document the calculation Calibrate/validate the model with actual honey yields |
| High seasonality of nectar yield | Assume low-input stationary beekeeping when estimating honey-provisioning capacity for the whole growing season or define a reference beekeeping practice Separately assess each nectar-flow period |
| Variation in nectar yield due to weather variability | Modify biophysical nectar-yield estimates with weather variables Calibrate/validate the model with actual honey yields |
| Variation in nectar yield due to site conditions | Modify the value of biophysical nectar-yield estimates with site variables Calibrate/validate the model with actual honey yields |
| Effects of ecosystem type and land use on bee health | Integrate variables describing plant communities and management practices that can affect (support or harm) bee health in the model |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arany, I.; Czúcz, B. Methods for Assessing the Ecosystem Service of Honey Provisioning by the European Honey Bee (Apis mellifera L.): A Systematic Review. Sustainability 2025, 17, 4533. https://doi.org/10.3390/su17104533
Arany I, Czúcz B. Methods for Assessing the Ecosystem Service of Honey Provisioning by the European Honey Bee (Apis mellifera L.): A Systematic Review. Sustainability. 2025; 17(10):4533. https://doi.org/10.3390/su17104533
Chicago/Turabian StyleArany, Ildikó, and Bálint Czúcz. 2025. "Methods for Assessing the Ecosystem Service of Honey Provisioning by the European Honey Bee (Apis mellifera L.): A Systematic Review" Sustainability 17, no. 10: 4533. https://doi.org/10.3390/su17104533
APA StyleArany, I., & Czúcz, B. (2025). Methods for Assessing the Ecosystem Service of Honey Provisioning by the European Honey Bee (Apis mellifera L.): A Systematic Review. Sustainability, 17(10), 4533. https://doi.org/10.3390/su17104533