



Mapping and Characterizing Eelgrass Meadows Using UAV Imagery in Placentia Bay and Trinity Bay, Newfoundland and Labrador, Canada

 , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Aerial Image Collection and Processing
2.3. Ground-Truth Data
2.4. OBIA and Classification
2.5. Classification Accuracy
2.6. Presence of Anthropogenic Stressors
3. Results
3.1. Eelgrass Distribution
3.2. Map Accuracy Assessment
3.3. Anthropogenic Disturbances
3.4. Semi-Quantitative Epiphyte Cover
4. Discussion
4.1. Presence of Anthropogenic Stressors
4.2. Image Segmentation
4.3. Monitoring Recommendations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Green, E.P.; Short, F.T. World Atlas of Seagrasses; University of California Press: Berkeley, CA, USA, 2004; Volume 41, ISBN 0520240472. [Google Scholar]
- Costanza, R.; D’Arge, R. The Value of the World’s Ecosystem Services and Natural Capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Nordlund, L.M.; Unsworth, R.K.F.; Gullström, M.; Cullen-Unsworth, L.C. Global Significance of Seagrass Fishery Activity. Fish Fish. 2018, 19, 399–412. [Google Scholar] [CrossRef]
- Unsworth, R.K.F.; Nordlund, L.M.; Cullen-Unsworth, L.C. Seagrass Meadows Support Global Fisheries Production. Conserv. Lett. 2019, 12, e12566. [Google Scholar] [CrossRef]
- Paul, M. The Protection of Sandy Shores—Can We Afford to Ignore the Contribution of Seagrass? Mar. Pollut. Bull. 2018, 134, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.M.; Losada, I.J.; Hendriks, I.E.; Mazarrasa, I.; Marbà, N. The Role of Coastal Plant Communities for Climate Change Mitigation and Adaptation. Nat. Clim. Chang. 2013, 3, 961–968. [Google Scholar] [CrossRef]
- Fourqurean, J.W.; Duarte, C.M.; Kennedy, H.; Marbà, N.; Holmer, M.; Mateo, M.A.; Apostolaki, E.T.; Kendrick, G.A.; Krause-Jensen, D.; McGlathery, K.J.; et al. Seagrass Ecosystems as a Globally Significant Carbon Stock. Nat. Geosci. 2012, 5, 505–509. [Google Scholar] [CrossRef]
- Nordlund, L.M.; Koch, E.W.; Barbier, E.B.; Creed, J.C. Seagrass Ecosystem Services and Their Variability across Genera and Geographical Regions. PLoS ONE 2016, 11, e0163091. [Google Scholar] [CrossRef] [PubMed]
- Borum, J.; Duarte, C.; Krause-Jensen, D.; Greve, T.M. European Seagrasses: An Introduction to Monitoring and Management; The M&M Project: Copenhagen, Denmark, 2004; ISBN 8789143213. [Google Scholar]
- Güreşen, A.; Güreşen, S.O.; Aktan, Y. Combined Synthetic and Biotic Indices of Posidonia Oceanica to Qualify the Status of Coastal Ecosystems in the North Aegean. Ecol. Indic. 2020, 113, 106149. [Google Scholar] [CrossRef]
- Kerninon, F.; Payri, C.E.; Le Loc’h, F.; Alcoverro, T.; Maréchal, J.P.; Chalifour, J.; Gréaux, S.; Mège, S.; Athanase, J.; Cordonnier, S.; et al. Selection of Parameters for Seagrass Management: Towards the Development of Integrated Indicators for French Antilles. Mar. Pollut. Bull. 2021, 170, 112646. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Crego, B.; Vergés, A.; Alcoverro, T.; Romero, J. Selection of Multiple Seagrass Indicators for Environmental Biomonitoring. Mar. Ecol. Prog. Ser. 2008, 361, 93–109. [Google Scholar] [CrossRef]
- Orth, R.J.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnick, S.; et al. A Global Crisis for Seagrass Ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; et al. Accelerating Loss of Seagrasses across the Globe Threatens Coastal Ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [PubMed]
- de los Santos, C.B.; Krause-Jensen, D.; Alcoverro, T.; Marbà, N.; Duarte, C.M.; van Katwijk, M.M.; Pérez, M.; Romero, J.; Sánchez-Lizaso, J.L.; Roca, G.; et al. Recent Trend Reversal for Declining European Seagrass Meadows. Nat. Commun. 2019, 10, 3356. [Google Scholar] [CrossRef] [PubMed]
- Lefcheck, J.S.; Orth, R.J.; Dennison, W.C.; Wilcox, D.J.; Murphy, R.R.; Keisman, J.; Gurbisz, C.; Hannam, M.; Brooke Landry, J.; Moore, K.A.; et al. Long-Term Nutrient Reductions Lead to the Unprecedented Recovery of a Temperate Coastal Region. Proc. Natl. Acad. Sci. USA 2018, 115, 3658–3662. [Google Scholar] [CrossRef] [PubMed]
- Krause-Jensen, D.; Duarte, C.M.; Sand-Jensen, K.; Carstensen, J. Century-Long Records Reveal Shifting Challenges to Seagrass Recovery. Glob. Chang. Biol. 2020, 27, 563–575. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, L.J.; Nordlund, L.M.; Jones, B.L.; Cullen-Unsworth, L.C.; Roelfsema, C.; Unsworth, R.K.F. The Global Distribution of Seagrass Meadows. Environ. Res. Lett. 2020, 15, 074041. [Google Scholar] [CrossRef]
- Unsworth, R.K.F.; McKenzie, L.J.; Collier, C.J.; Cullen-Unsworth, L.C.; Duarte, C.M.; Eklöf, J.S.; Jarvis, J.C.; Jones, B.L.; Nordlund, L.M. Global Challenges for Seagrass Conservation. Ambio 2019, 48, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.E.P.; Dunic, J.C.; Adamczyk, E.M.; Bittick, S.J.; Côté, I.M.; Cristiani, J.; Geissinger, E.A.; Gregory, R.S.; Lotze, H.K.; O’Connor, M.I.; et al. From Coast to Coast to Coast: Ecology and Management of Seagrass Ecosystems across Canada. Facets 2021, 6, 139–179. [Google Scholar] [CrossRef]
- DFO. Does Eelgrass (Zostera marina) Meet the Criteria as an Ecologically Significant Species? CSAS Sci. Advis. Rep. 2009, 18, 11. [Google Scholar]
- Matheson, K.; McKenzie, C.H.; Gregory, R.S.; Robichaud, D.A.; Bradbury, I.R.; Snelgrove, P.V.R.; Rose, G.A. Linking Eelgrass Decline and Impacts on Associated Fish Communities to European Green Crab Carcinus Maenas Invasion. Mar. Ecol. Prog. Ser. 2016, 548, 31–45. [Google Scholar] [CrossRef]
- Blakeslee, A.M.H.; McKenzie, C.H.; Darling, J.A.; Byers, J.E.; Pringle, J.M.; Roman, J. A Hitchhiker’s Guide to the Maritimes: Anthropogenic Transport Facilitates Long-Distance Dispersal of an Invasive Marine Crab to Newfoundland. Divers. Distrib. 2010, 16, 879–891. [Google Scholar] [CrossRef]
- Davis, R.C.; Short, F.T.; Burdick, D.M. Quantifying the Effects of Green Crab Damage to Eelgrass Transplants. Restor. Ecol. 1998, 6, 297–302. [Google Scholar] [CrossRef]
- Warren, M.A.; Gregory, R.S.; Laurel, B.J.; Snelgrove, P.V.R. Increasing Density of Juvenile Atlantic (Gadus morhua) and Greenland Cod (G. ogac) in Association with Spatial Expansion and Recovery of Eelgrass (Zostera marina) in a Coastal Nursery Habitat. J. Exp. Mar. Biol. Ecol. 2010, 394, 154–160. [Google Scholar] [CrossRef]
- Rao, A.S.; Gregory, R.S.; Murray, G.; Ings, D.W.; Coughlan, E.J.; Newton, B.H. Eelgrass (Zostera marina) Locations in Newfoundland and Labrador; Fisheries and Oceans Canada: St. John’s, NL, Canada, 2014. [Google Scholar]
- Prystay, T.S.; Adams, G.; Favaro, B.; Gregory, R.S.; Le Bris, A. The Reproducibility of Remotely Piloted Aircraft Systems to Monitor Seasonal Variation in Submerged Seagrass and Estuarine Habitats. Facets 2023, 8, 1–22. [Google Scholar] [CrossRef]
- DFA. Issues Scan of Selected Coastal and Ocean Areas of Newfoundland and Labrador. 2007. Available online: https://www.gov.nl.ca/ffa/files/publications-archives-pdf-issues-scan-of-selected-coastal-areas-of-newfoundland-and-labrador.pdf (accessed on 17 April 2024).
- LGL. Placentia Bay Atlantic Salmon Aquaculture Project Environmental Protection Plan (EPP): RAS Hatchery Operations. 2020. Available online: https://www.gov.nl.ca/ecc/files/FA0159-GriegNL-EPP-RAS-Hatchery-Operations-2-Oct.-2020.pdf (accessed on 17 April 2024).
- Catto, N.R.; Hooper, R.G.; Anderson, M.R.; Scruton, D.A.; Meade, J.D.; Ollerhead, L.M.N.; Williams, U.P. Biological and Geomorphological Classification of Placentia Bay: A Preliminary Assessment; Canadian Technical Report of Fisheries and Aquatic Sciences 1488-5379 No. 2289; Fisheries and Oceans Canada: St. John’s, NL, Canada, 1999. [Google Scholar]
- Cullain, N.; McIver, R.; Schmidt, A.L.; Milewski, I.; Lotze, H.K. Potential Impacts of Finfish Aquaculture on Eelgrass (Zostera marina) Beds and Possible Monitoring Metrics for Management: A Case Study in Atlantic Canada. PeerJ 2018, 6, e5630. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.L.; Cullen-Unsworth, L.C.; Unsworth, R.K.F. Tracking Nitrogen Source Using Δ15 N Reveals Human and Agricultural Drivers of Seagrass Degradation across the British Isles. Front. Plant Sci. 2018, 9, 329228. [Google Scholar] [CrossRef]
- Hallac, D.E.; Sadle, J.; Pearlstine, L.; Herling, F.; Shinde, D. Boating Impacts to Seagrass in Florida Bay, Everglades National Park, Florida, USA: Links with Physical and Visitor-Use Factors and Implications for Management. Mar. Freshw. Res. 2012, 63, 1117–1128. [Google Scholar] [CrossRef]
- Orth, R.J.; Lefcheck, J.S.; Wilcox, D.J. Boat Propeller Scarring of Seagrass Beds in Lower Chesapeake Bay, USA: Patterns, Causes, Recovery, and Management. Estuaries Coasts 2017, 40, 1666–1676. [Google Scholar] [CrossRef]
- Glasby, T.M.; West, G. Dragging the Chain: Quantifying Continued Losses of Seagrasses from Boat Moorings. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 383–394. [Google Scholar] [CrossRef]
- Unsworth, R.K.F.; Williams, B.; Jones, B.L.; Cullen-Unsworth, L.C. Rocking the Boat: Damage to Eelgrass by Swinging Boat Moorings. Front. Plant Sci. 2017, 8, 262774. [Google Scholar] [CrossRef] [PubMed]
- Eriander, L.; Laas, K.; Bergström, P.; Gipperth, L.; Moksnes, P.O. The Effects of Small-Scale Coastal Development on the Eelgrass (Zostera marina L.) Distribution along the Swedish West Coast—Ecological Impact and Legal Challenges. Ocean Coast. Manag. 2017, 148, 182–194. [Google Scholar] [CrossRef]
- Gladstone, W.; Courtenay, G. Impacts of Docks on Seagrass and Effects of Management Practices to Ameliorate These Impacts. Estuar. Coast. Shelf Sci. 2014, 136, 53–60. [Google Scholar] [CrossRef]
- Kelly, J.J.; Orr, D.; Takekawa, J.Y. Quantification of Damage to Eelgrass (Zostera marina) Beds and Evidence-Based Management Strategies for Boats Anchoring in San Francisco Bay. Environ. Manag. 2019, 64, 20–26. [Google Scholar] [CrossRef] [PubMed]
- La Manna, G.; Donno, Y.; Sarà, G.; Ceccherelli, G. The Detrimental Consequences for Seagrass of Ineffective Marine Park Management Related to Boat Anchoring. Mar. Pollut. Bull. 2015, 90, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.C.; Bravo, M.A.; Dowd, M. Ecological Dynamics of Zostera marina (Eelgrass) in Three Adjacent Bays in Atlantic Canada. Bot. Mar. 2013, 56, 413–424. [Google Scholar] [CrossRef]
- Nahirnick, N.K.; Reshitnyk, L.; Campbell, M.; Hessing-Lewis, M.; Costa, M.; Yakimishyn, J.; Lee, L. Mapping with Confidence; Delineating Seagrass Habitats Using Unoccupied Aerial Systems (UAS). Remote Sens. Ecol. Conserv. 2019, 5, 121–135. [Google Scholar] [CrossRef]
- Agisoft LLC Agisoft Metashape 2019. Agisoft LLC, St. Petersburg, Russia. Available online: https://www.agisoft.com/ (accessed on 17 April 2024).
- ESRI ArcGIS 2019. Environmental Systems Research Institute, Redlands, California, USA. Available online: https://www.esri.com/en-us/home (accessed on 17 April 2024).
- Nahirnick, N.K.; Hunter, P.; Costa, M.; Schroeder, S.; Sharma, T. Benefits and Challenges of UAS Imagery for Eelgrass (Zostera marina) Mapping in Small Estuaries of the Canadian West Coast. J. Coast. Res. 2019, 35, 673–683. [Google Scholar] [CrossRef]
- Blaschke, T. Object Based Image Analysis for Remote Sensing. ISPRS J. Photogramm. Remote Sens. 2010, 65, 2–16. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing 2020. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 17 April 2024).
- Gonçalves, J.; Pôças, I.; Marcos, B.; Mücher, C.A.; Honrado, J.P. SegOptim—A New R Package for Optimizing Object-Based Image Analyses of High-Spatial Resolution Remotely-Sensed Data. Int. J. Appl. Earth Obs. Geoinf. 2019, 76, 218–230. [Google Scholar] [CrossRef]
- Comaniciu, D.; Meer, P. Mean Shift: A Robust Approach toward Feature Space Analysis. IEEE Trans. Pattern Anal. Mach. Intell. 2002, 24, 603–619. [Google Scholar] [CrossRef]
- Chabot, D.; Dillon, C.; Shemrock, A.; Weissflog, N.; Sager, E.P.S. An Object-Based Image Analysis Workflow for Monitoring Shallow-Water Aquatic Vegetation in Multispectral Drone Imagery. ISPRS Int. J. Geo-Inform. 2018, 7, 294. [Google Scholar] [CrossRef]
- Ellis, S.L.; Taylor, M.L.; Schiele, M.; Letessier, T.B. Influence of Altitude on Tropical Marine Habitat Classification Using Imagery from Fixed-Wing, Water-Landing UAVs. Remote Sens. Ecol. Conserv. 2020, 2, 50–63. [Google Scholar] [CrossRef]
- Oldeland, J.; Revermann, R.; Luther-Mosebach, J.; Buttschardt, T.; Lehmann, J.R.K. New Tools for Old Problems—Comparing Drone- and Field-Based Assessments of a Problematic Plant Species. Environ. Monit. Assess. 2021, 193, 90. [Google Scholar] [CrossRef] [PubMed]
- Belgiu, M.; Drăgu, L. Random Forest in Remote Sensing: A Review of Applications and Future Directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Millard, K.; Richardson, M. On the Importance of Training Data Sample Selection in Random Forest Image Classification: A Case Study in Peatland Ecosystem Mapping. Remote Sens. 2015, 7, 8489–8515. [Google Scholar] [CrossRef]
- Papakonstantinou, A.; Stamati, C.; Topouzelis, K. Comparison of True-Color and Multispectral Unmanned Aerial Systems Imagery for Marine Habitat Mapping Using Object-Based Image Analysis. Remote Sens. 2020, 12, 554. [Google Scholar] [CrossRef]
- Moran, P. Notes on Continuous Stochastic Phenomena. Biometrika 1950, 37, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Sim, J.; Wright, C.C. The Kappa Statistic in Reliability Studies: Use, Interpretation, and Sample Size Requirements. Phys. Ther. 2005, 85, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W.G. Development of an Epiphyte Indicator of Nutrient Enrichment: A Critical Evaluation of Observational and Experimental Studies. Ecol. Indic. 2017, 79, 207–227. [Google Scholar] [CrossRef] [PubMed]
- Kelleway, J. Ecological Impacts of Recreational Vehicle Use on Saltmarshes of the Georges River, Sydney. Wetl. Aust. 2006, 22, 52. [Google Scholar] [CrossRef][Green Version]
- Martin, S.R.; Onuf, C.P.; Dunton, K.H. Assessment of Propeller and Off-Road Vehicle Scarring in Seagrass Beds and Wind-Tidal Flats of the Southwestern Gulf of Mexico. Bot. Mar. 2008, 51, 79–91. [Google Scholar] [CrossRef]
- Evans, S.M.; Griffin, K.J.; Blick, R.A.J.; Poore, A.G.B.; Vergés, A. Seagrass on the Brink: Decline of Threatened Seagrass Posidonia Australis Continues Following Protection. PLoS ONE 2018, 13, e0190370. [Google Scholar] [CrossRef] [PubMed]
- Canada, S. Census Profile, 2016 Census Placentia, Town [Census Subdivision], Newfoundland and Labrador and Division No. 1, Census Division [Census Division], Newfoundland and Labrador. Available online: https://www12.statcan.gc.ca/census-recensement/2016/dp-pd/prof/details/page.cfm?Lang=E&Geo1=CSD&Code1=1001240&Geo2=CD&Code2=1001&SearchText=Placentia&SearchType=Begins&SearchPR=01&B1=All&TABID=1&type=0 (accessed on 17 April 2024).
- Lavery, P.S.; Reid, T.; Hyndes, G.A.; Van Elven, B.R. Effect of Leaf Movement on Epiphytic Algal Biomass of Seagrass Leaves. Mar. Ecol. Prog. Ser. 2007, 338, 97–106. [Google Scholar] [CrossRef]
- Strand, J.A.; Weisner, S.E.B. Wave Exposure Related Growth of Epiphyton: Implications for the Distribution of Submerged Macrophytes in Eutrophic Lakes. Hydrobiologia 1996, 325, 113–119. [Google Scholar] [CrossRef]
- Orth, R.J.; Williams, M.R.; Marion, S.R.; Wilcox, D.J.; Carruthers, T.J.B.; Moore, K.A.; Kemp, W.M.; Dennison, W.C.; Rybicki, N.; Bergstrom, P.; et al. Long-Term Trends in Submersed Aquatic Vegetation (SAV) in Chesapeake Bay, USA, Related to Water Quality. Estuaries Coasts 2010, 33, 1144–1163. [Google Scholar] [CrossRef]
- Murphy, G.E.P.; Wong, M.C.; Lotze, H.K. A Human Impact Metric for Coastal Ecosystems with Application to Seagrass Beds in Atlantic Canada. Facets 2019, 4, 210–237. [Google Scholar] [CrossRef]
- Sagerman, J.; Hansen, J.P.; Wikström, S.A. Effects of Boat Traffic and Mooring Infrastructure on Aquatic Vegetation: A Systematic Review and Meta-Analysis. Ambio 2020, 49, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, P.; Teodoro, A.C.; Gonçalves, J.A.; Honrado, J.P.; Cunha, M.; Sillero, N. Assessing the Performance of Different OBIA Software Approaches for Mapping Invasive Alien Plants along Roads with Remote Sensing Data. Int. J. Appl. Earth Obs. Geoinf. 2021, 95, 102263. [Google Scholar] [CrossRef]
- Kavzoglu, T.; Tonbul, H. An Experimental Comparison of Multi-Resolution Segmentation, SLIC and K-Means Clustering for Object-Based Classification of VHR Imagery. Int. J. Remote Sens. 2018, 39, 6020–6036. [Google Scholar] [CrossRef]
- Teodoro, A.C. Comparison of Performance of Object-Based Image Analysis Techniques Available in Open Source Software (Spring and Orfeo Toolbox/Monteverdi) Considering Very High Spatial Resolution Data. J. Appl. Remote Sens. 2016, 10, 016011. [Google Scholar] [CrossRef]
- Traganos, D.; Aggarwal, B.; Poursanidis, D.; Topouzelis, K.; Chrysoulakis, N.; Reinartz, P. Towards Global-Scale Seagrass Mapping and Monitoring Using Sentinel-2 on Google Earth Engine: The Case Study of the Aegean and Ionian Seas. Remote Sens. 2018, 10, 1227. [Google Scholar] [CrossRef]
- Neckles, H.A.; Kopp, B.S.; Peterson, B.J.; Pooler, P.S. Integrating Scales of Seagrass Monitoring to Meet Conservation Needs. Estuaries Coasts 2012, 35, 23–46. [Google Scholar] [CrossRef]
- Neckles, H.A. Loss of Eelgrass in Casco Bay, Maine, Linked to Green Crab Disturbance. Northeast. Nat. 2015, 22, 478–500. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
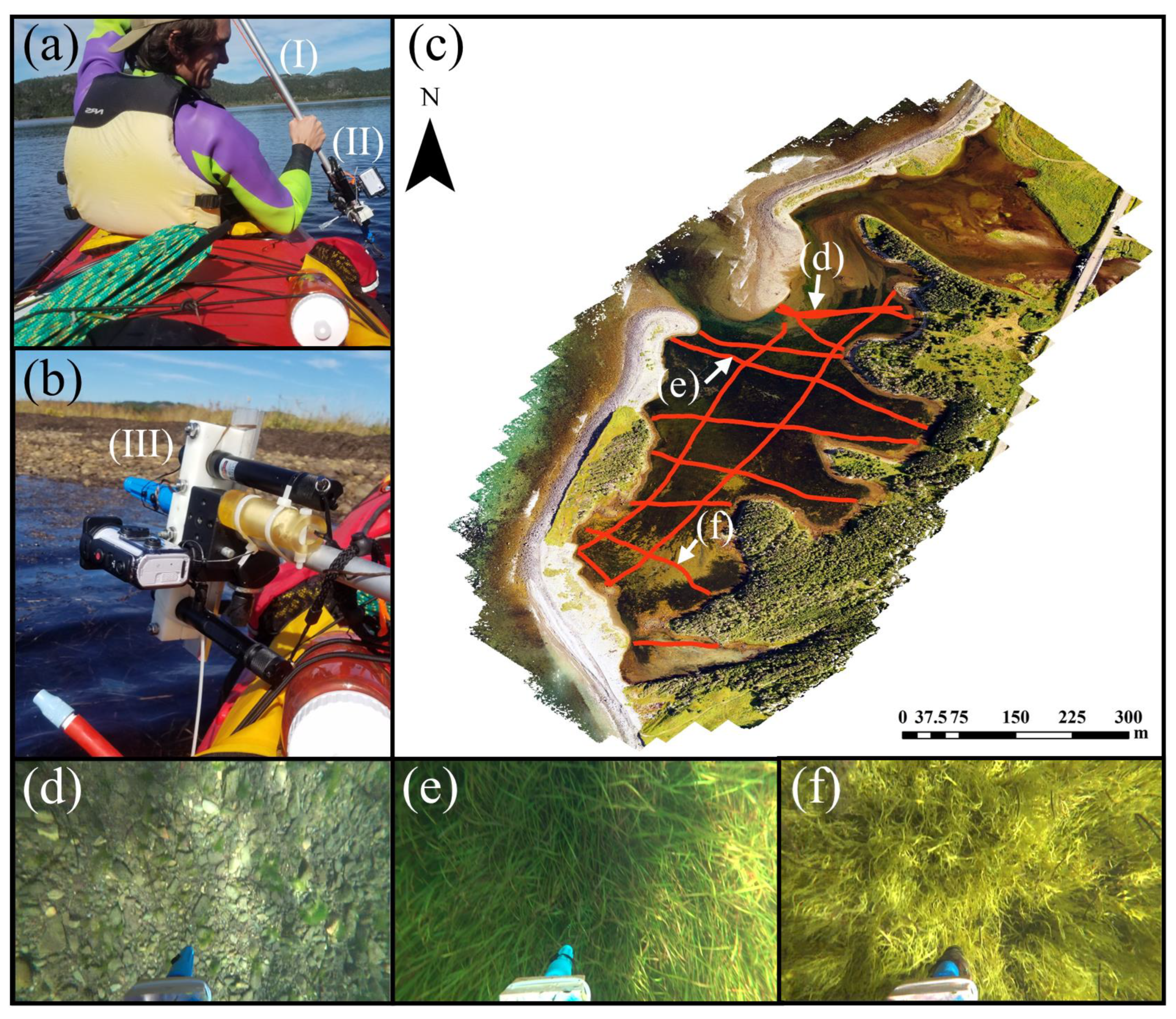{kind=link}
{kind=link}
{kind=link}
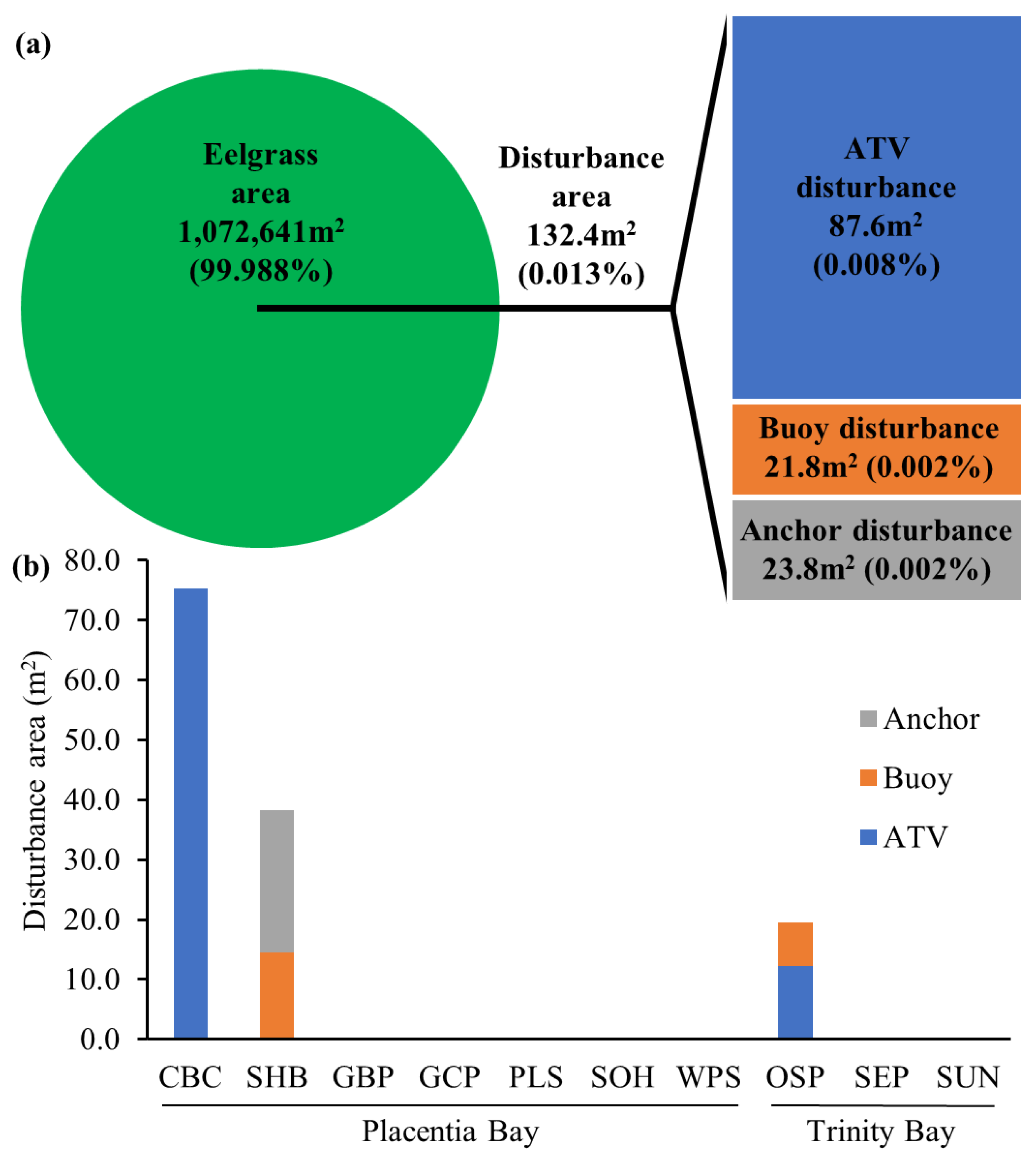{kind=link}
{kind=link}
| Study Site | Mean Kappa | Overall Accuracy (%) | Eelgrass Producer’s Accuracy (%) | Eelgrass User’s Accuracy (%) | Eelgrass Area (km2) |
|---|---|---|---|---|---|
| Come By Chance Gut | 0.71 ± 0.08 | 82.2 ± 4.9 | 89.9 ± 9.0 | 87.9 ± 7.3 | 0.1310 |
| Glennons Cove Pond | 0.67 ± 0.08 | 82.2 ± 3.5 | 60.0 ± 43.5 | 73.3 ± 43.5 | 0.0013 |
| Great Barasway Pond | 0.81 ± 0.04 | 89.3 ± 2.8 | 98.4 ± 2.3 | 95.1 ± 0.4 | 0.0597 |
| Old Shop Pond | 0.73 ± 0.10 | 82.0 ± 6.9 | 88.5 ± 6.5 | 89.2 ± 9.7 | 0.0687 |
| Placentia Swans | 0.46 ± 0.07 | 69.7 ± 4.1 | 56.0 ± 5.2 | 62.1 ± 8.3 | 0.0916 |
| Ship Harbour | 0.72 ± 0.08 | 82.2 ± 5.1 | 91.7 ± 7.3 | 77.2 ± 10.9 | 0.0587 |
| Southern Harbour | 0.61 ± 0.15 | 75.8 ± 9.9 | 23.3 ± 14.9 | 48.8 ± 36.6 | 0.0054 |
| Spread Eagle Pond | 0.65 ± 0.14 | 84.7 ± 6.3 | 95.4 ± 3.8 | 90.0 ± 3.7 | 0.1967 |
| Sunny Side | 0.69 ± 0.10 | 80.3 ± 6.3 | 89.8 ± 5.6 | 81.0 ± 5.2 | 0.1265 |
| Western Placentia Southeast Arm | 0.22 ± 0.23 | 86.3 ± 3.1 | 99.2 ± 1.1 | 87.5 ± 3.0 | 0.3331 |
| Site | Bay | Eelgrass Area (km2) | Disturbance Area (m2) | Source of Disturbance | Signs of Eutrophication |
|---|---|---|---|---|---|
| Come By Chance | Placentia Bay | 0.1310 | 75.4 | ATV | / |
| Glennons Cove Pond | Placentia Bay | 0.0013 | / | / | / |
| Great Barasway Pond | Placentia Bay | 0.0597 | / | / | / |
| Old Shop Pond | Trinity Bay | 0.0687 | 19.6 | ATV, buoy | / |
| Placentia Swans | Placentia Bay | 0.0916 | / | / | proliferation of epiphytes |
| Ship Harbour | Placentia Bay | 0.0587 | 38.3 | buoy, anchor | / |
| Southern Harbour | Placentia Bay | 0.0054 | / | / | / |
| Spread Eagle Pond | Trinity Bay | 0.1967 | / | / | / |
| Sunny Side | Trinity Bay | 0.1265 | / | / | / |
| Western Placentia Southeast Arm | Placentia Bay | 0.3331 | / | / | floating algal mats |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sneep, A.; Devillers, R.; Robert, K.; Le Bris, A.; Edinger, E. Mapping and Characterizing Eelgrass Meadows Using UAV Imagery in Placentia Bay and Trinity Bay, Newfoundland and Labrador, Canada. Sustainability 2024, 16, 3471. https://doi.org/10.3390/su16083471
Sneep A, Devillers R, Robert K, Le Bris A, Edinger E. Mapping and Characterizing Eelgrass Meadows Using UAV Imagery in Placentia Bay and Trinity Bay, Newfoundland and Labrador, Canada. Sustainability. 2024; 16(8):3471. https://doi.org/10.3390/su16083471
Chicago/Turabian StyleSneep, Aaron, Rodolphe Devillers, Katleen Robert, Arnault Le Bris, and Evan Edinger. 2024. "Mapping and Characterizing Eelgrass Meadows Using UAV Imagery in Placentia Bay and Trinity Bay, Newfoundland and Labrador, Canada" Sustainability 16, no. 8: 3471. https://doi.org/10.3390/su16083471
APA StyleSneep, A., Devillers, R., Robert, K., Le Bris, A., & Edinger, E. (2024). Mapping and Characterizing Eelgrass Meadows Using UAV Imagery in Placentia Bay and Trinity Bay, Newfoundland and Labrador, Canada. Sustainability, 16(8), 3471. https://doi.org/10.3390/su16083471