


Biological Activity of Four Trichoderma Species Confers Protection against Rhizoctonia solani, the Causal Agent of Cucumber Damping-Off and Root Rot Diseases




Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of R. solani and Trichoderma
2.2. Molecular Characterization of R. solani and Trichoderma Species
2.3. Pathogenicity Test
2.3.1. Preparing of Inoculum
2.3.2. Preparation of Soil and Pots
2.3.3. Inoculation Procedure
2.3.4. Disease Assessment
- (a)
- Pre-emergence damping-off percentage was estimated after 15 days:Pre-emergence (%) = Number of non-germinated seeds/Number of sown seeds × 100
- (b)
- Post-emergence damping-off percentage was estimated after 30 days:Post-emergence (%) = Number of dead seedlings/Number of survival plants × 100
- (c)
- Survival seedlings (%) = Number of survived plants/ Number of sown seeds × 100
- (d)
- Disease incidence (DI) percentage was estimated after 45 days of seeding:DI (%) = Number of infected plants/ Total number of examined plants × 100.
2.4. Evaluation of Trichoderma Species against R. solani In Vitro
- I = [(C − T)/C] × 100;
- I = inhibition of radial mycelia growth;
- C = the pathogen radial growth measurement in control;
- T = the pathogen radial growth in the existence of Trichoderma isolates.
2.5. Scanning Electron Microscopy (SEM) Examination
2.6. Effect of Trichoderma Species on Root Rot and Damping off Diseases under Greenhouse Conditions
2.7. Assessment of Defense-Related Enzymes
2.7.1. Polyphenol-Oxidase Activity
2.7.2. Peroxidase Activity
2.7.3. Catalase Activity
2.7.4. Chitinase Activity
2.8. Estimation of Total Phenol Content
2.9. Estimation of Total Chlorophyll Content
- Chl. a = 12.72A663 − 2.59A645
- Chl. b = 22.9A663 − 4.67A645
- Chl. t = 20.31A645 + 8.05A663
2.10. Statistical Analysis
3. Results
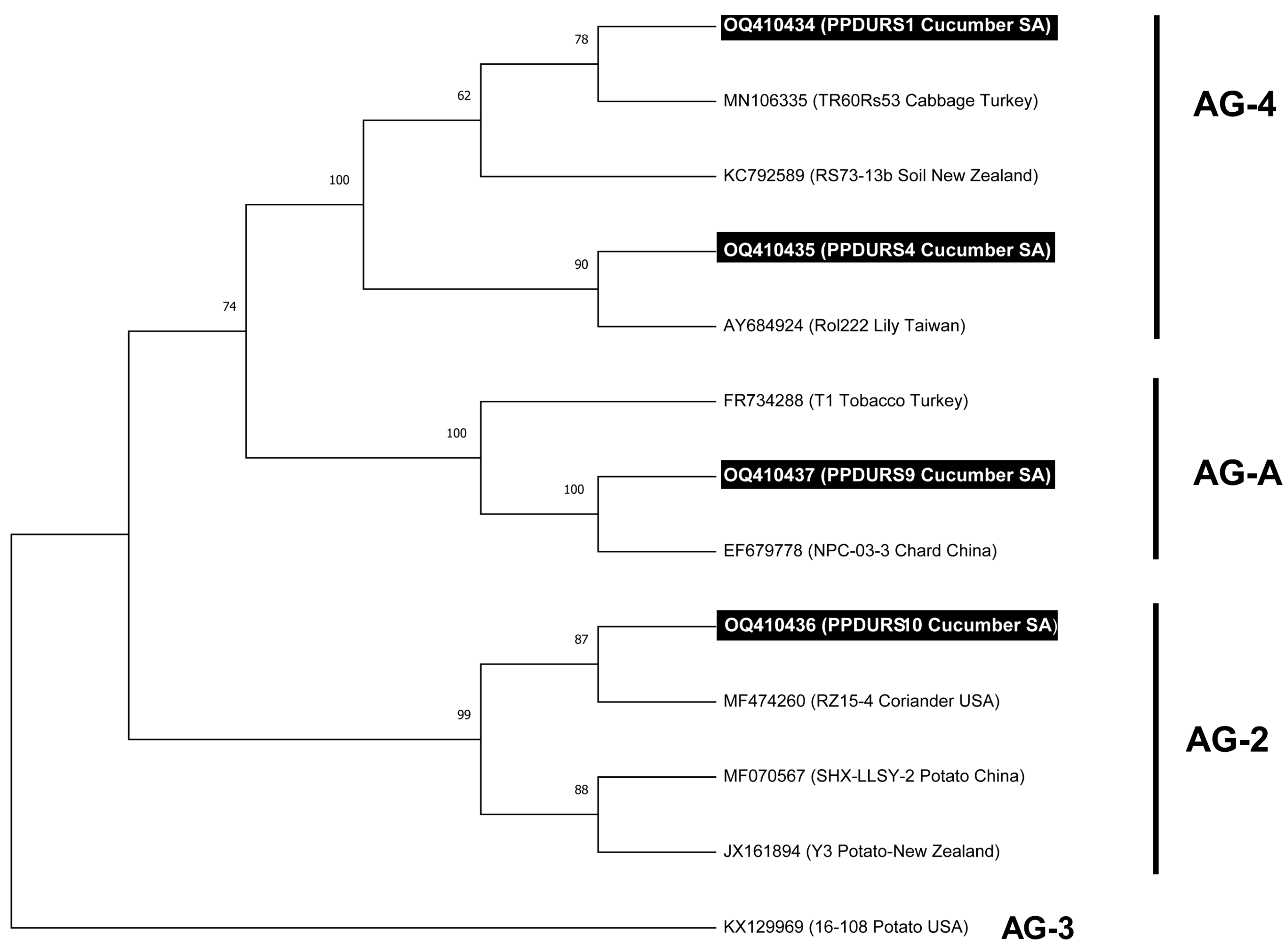
3.1. R. solani Isolation and Molecular Characterization
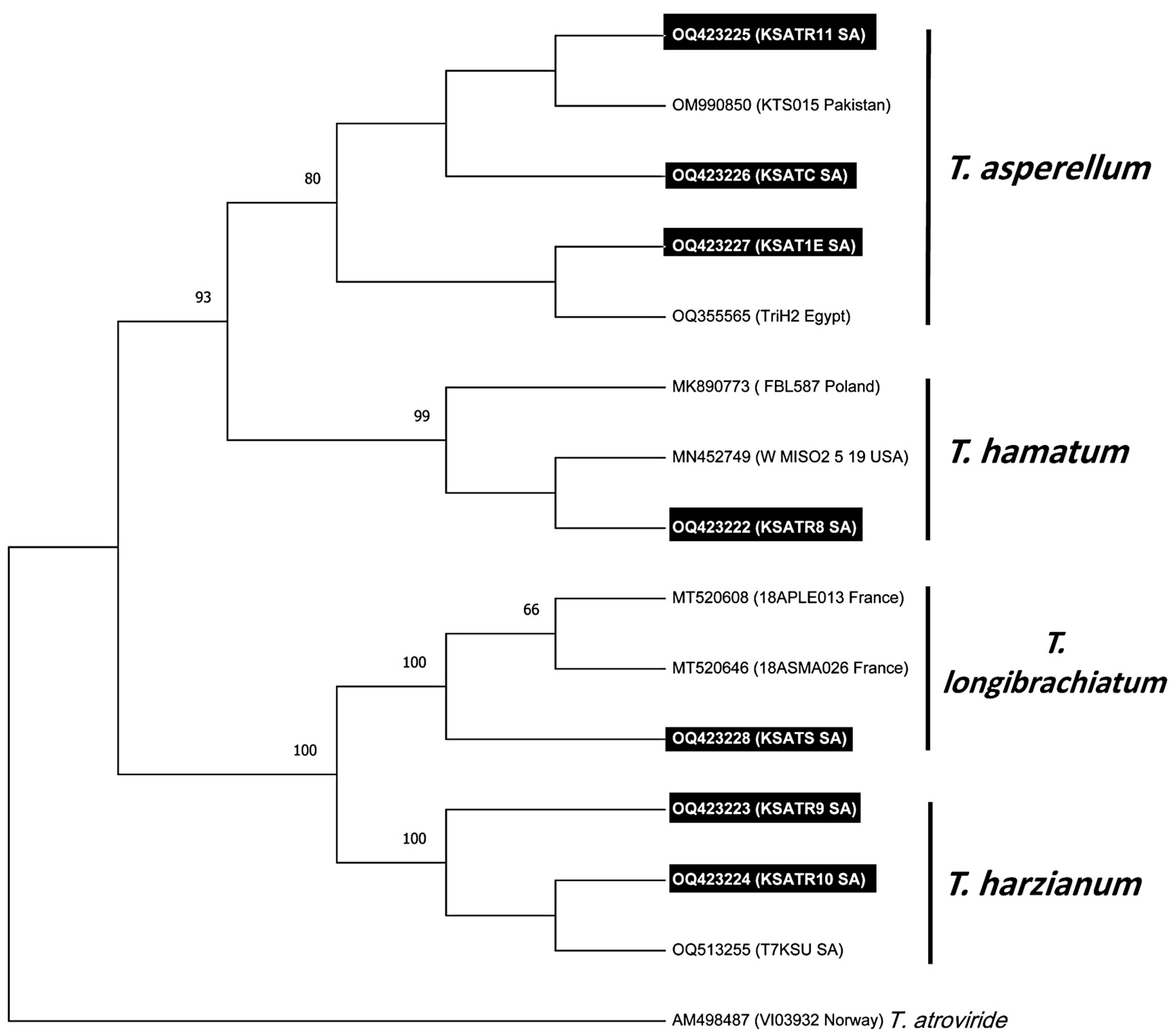
3.2. Trichoderma Species Isolation and Molecular Characterization
3.3. Pathogenicity Test
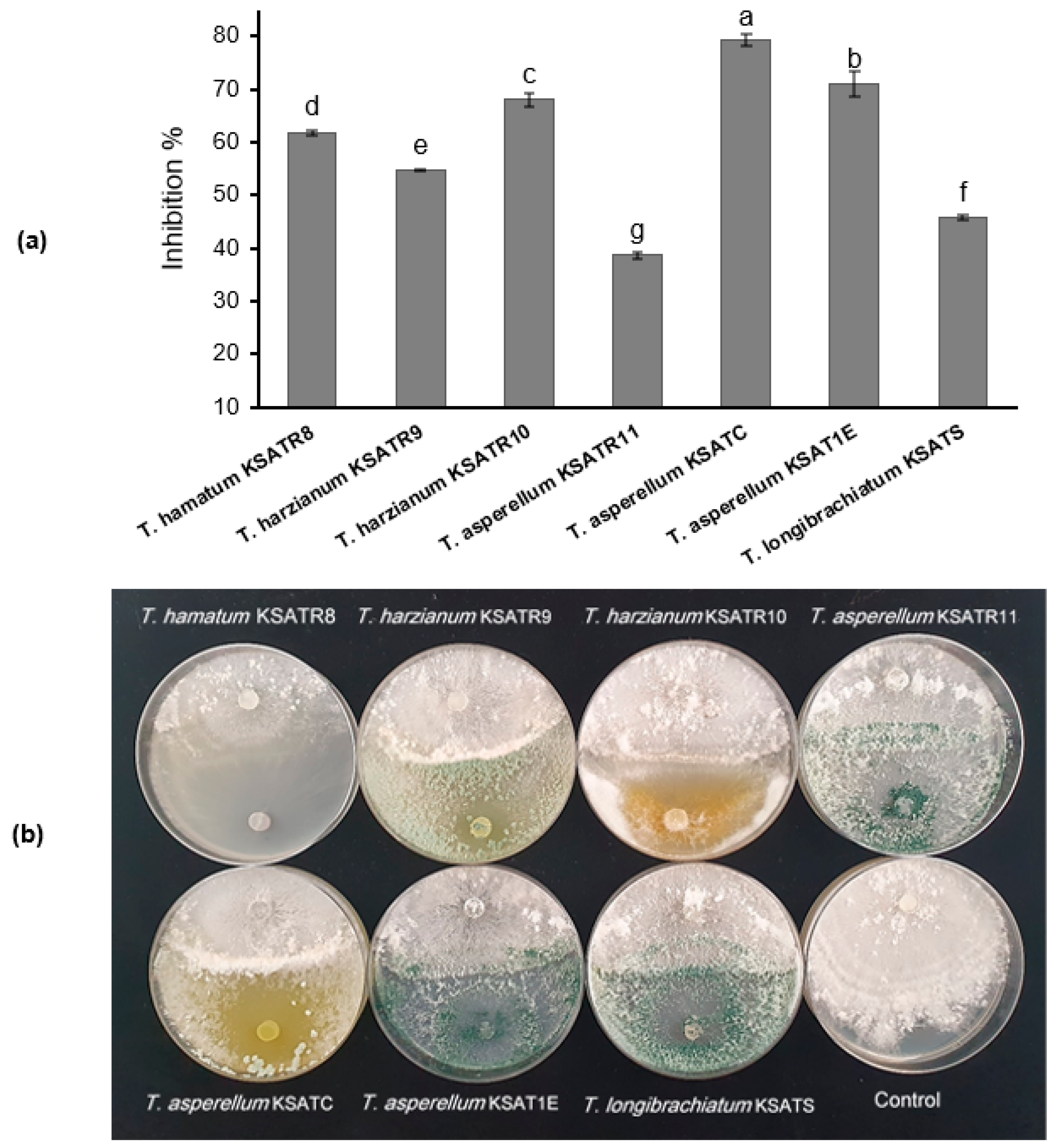
3.4. Antagonistic Activity of Trichoderma Species on the R. solani Mycelial Growth
3.5. Scanning Electron Microscopy (SEM) Examination
3.6. Effect of Trichoderma Species on Damping-Off and Root Rot Disease under Greenhouse Conditions
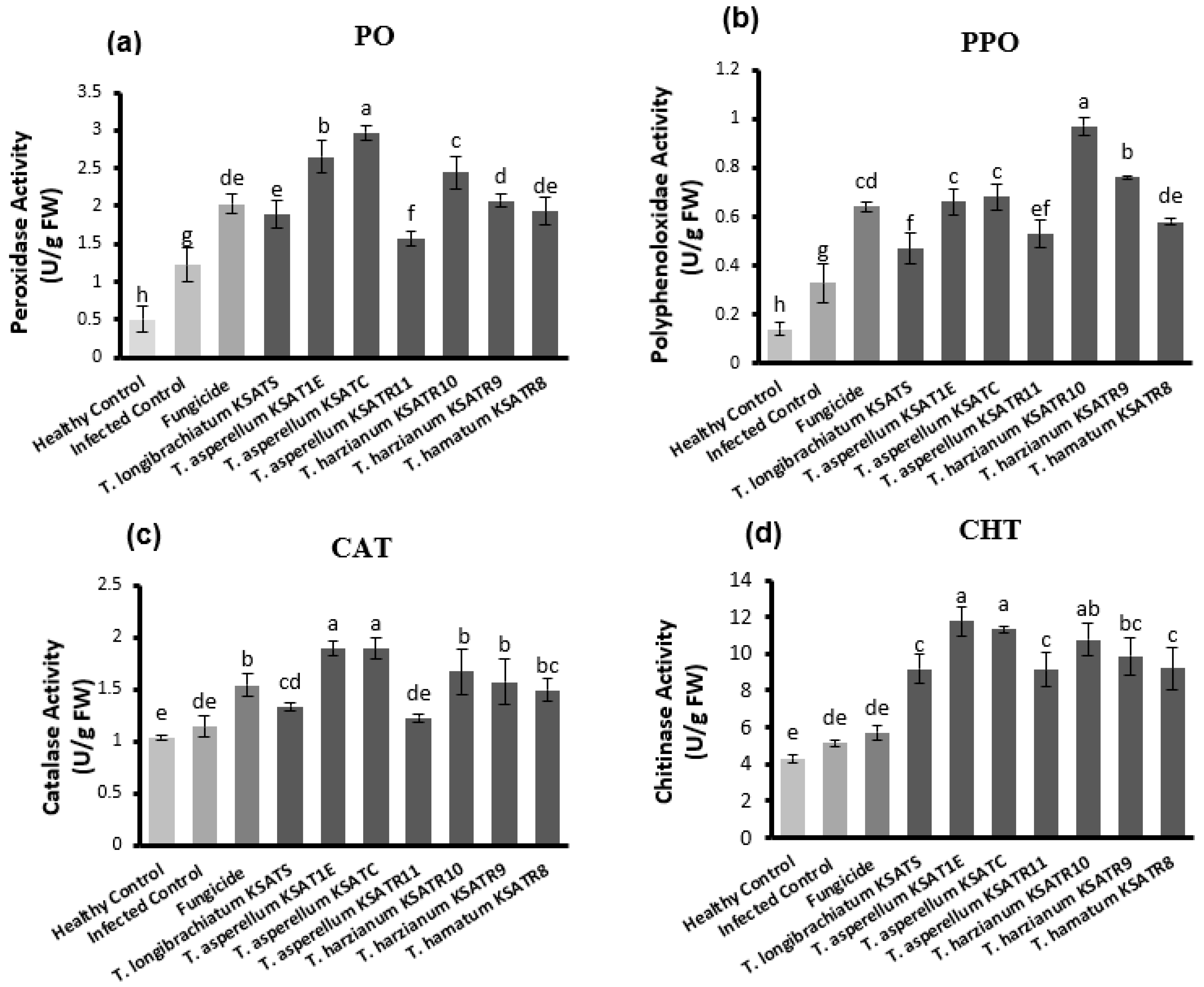
3.7. Assessment of Defense-Related Enzymes
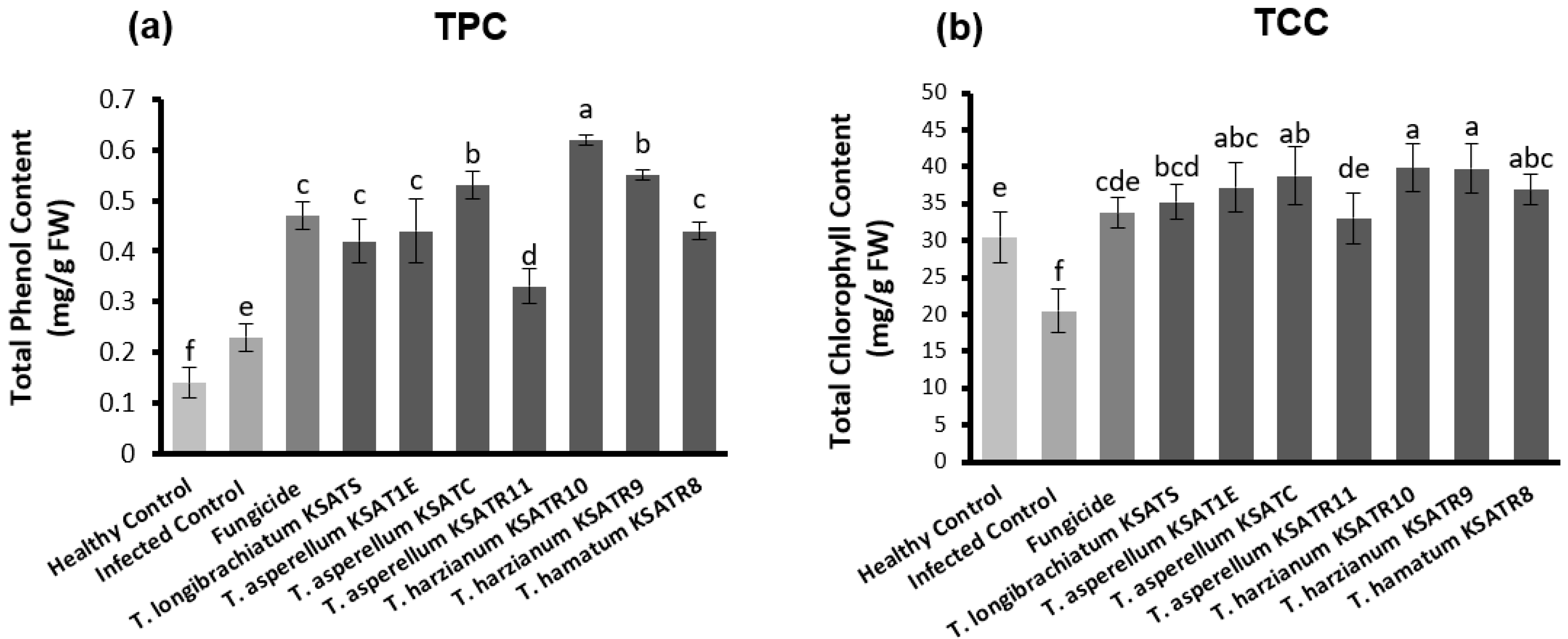
3.8. Estimation of TPC and TCC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crane, M.; Wehner, T.C.; Naegele, R.P. Cucumber Cultivars for Container Gardening and the Value of Field Trials for Predicting Cucumber Performance in Containers. HortScience 2018, 53, 16–22. [Google Scholar] [CrossRef]
- Mahmoud, A.F.; Abdalla, O.A. Biological Control of Fungi Associated with Damping-off and Root Rot Disease of Cucumber (Cucumis sativus L.). Arch. Phytopathol. Plant Prot. 2021, 54, 870–885. [Google Scholar] [CrossRef]
- El-Komy, M.H.; Al-Qahtani, R.M.; Widyawan, A.; Molan, Y.; Almasrahi, A. First Report of Fusarium Root and Stem Rot Caused by Fusarium oxysporum f. sp. radicis-cucumerinum on Greenhouse Cucumbers in Saudi Arabia. Plant Dis. 2021, 105, 3758. [Google Scholar] [CrossRef] [PubMed]
- Morsy, S.M.; Drgham, E.A.; Mohamed, G.M. Effect of Garlic and Onion Extracts or Their Intercropping on Suppressing Damping-off and Powdery Mildew Diseases and Growth Characteristics of Cucumber. Egypt. J. Phytopathol. 2009, 37, 35–46. [Google Scholar]
- Ristaino, J.B.; Johnston, S.A. Ecologically Based Approaches to Management of Phytophthora Blight on Bell Pepper. Plant Dis. 1999, 83, 1080–1089. [Google Scholar] [CrossRef]
- Simsek Ersahin, Y.; Haktanir, K.; Yanar, Y. Vermicompost suppresses Rhizoctonia solani Kühn in cucumber seedlings. J. Plant Dis. Prot. 2009, 116, 182–188. [Google Scholar] [CrossRef]
- Heflish, A.A.; Abdelkhalek, A.; Al-Askar, A.A.; Behiry, S.I. Protective and Curative Effects of Trichoderma asperelloides Ta41 on Tomato Root Rot Caused by Rhizoctonia solani Rs33. Agronomy 2021, 11, 1162. [Google Scholar] [CrossRef]
- Wallon, T.; Sauvageau, A.; Van der Heyden, H. Detection and Quantification of Rhizoctonia solani and Rhizoctonia solani AG1-IB Causing the Bottom Rot of Lettuce in Tissues and Soils by Multiplex QPCR. Plants 2021, 10, 57. [Google Scholar] [CrossRef]
- Williamson-Benavides, B.A.; Dhingra, A. Understanding Root Rot Disease in Agricultural Crops. Horticulturae 2021, 7, 33. [Google Scholar] [CrossRef]
- Erper, I.; Ozer, G.; Kalendar, R.; Avci, S.; Yildirim, E.; Alkan, M.; Turkkan, M. Genetic Diversity and Pathogenicity of Rhizoctonia spp. Isolates Associated with Red Cabbage in Samsun (Turkey). J. Fungi 2021, 7, 234. [Google Scholar] [CrossRef]
- Yang, G.; Li, C. General Description of Rhizoctonia Species Complex. In Plant pathology; Citeseer: Princeton, NJ, USA, 2012; ISBN 9535104896. [Google Scholar]
- Sneh, B.; Burpee, L.; Ogoshi, A. Identification of Rhizoctonia Species; APS Press: College Park, MD, USA, 1991; ISBN 089054123X. [Google Scholar]
- Yang, Y.; Zhao, C.; Guo, Z.; Wu, X. Anastomosis Groups and Pathogenicity of Binucleate Rhizoctonia Isolates Associated with Stem Canker of Potato in China. Eur. J. Plant Pathol. 2014, 139, 535–544. [Google Scholar] [CrossRef]
- Stodart, B.J.; Harvey, P.R.; Neate, S.M.; Melanson, D.L.; Scott, E.S. Genetic Variation and Pathogenicity of Anastomosis Group 2 Isolates of Rhizoctonia solani in Australia. Mycol. Res. 2007, 111, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Guleria, S.; Aggarwal, R.; Thind, T.S.; Sharma, T.R. Morphological and Pathological Variability in Rice Isolates of Rhizoctonia solani and Molecular Analysis of Their Genetic Variability. J. Phytopathol. 2007, 155, 654–661. [Google Scholar] [CrossRef]
- Gonzalez-Vera, A.D.; Bernardes-de-Assis, J.; Zala, M.; McDonald, B.A.; Correa-Victoria, F.; Graterol-Matute, E.J.; Ceresini, P.C. Divergence between Sympatric Rice-and Maize-Infecting Populations of Rhizoctonia solani AG-1 IA from Latin America. Phytopathology 2010, 100, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Ciampi, M.B.; Gale, L.R.; Lemos, E.G.; Ceresini, P.C. Distinctively Variable Sequence-Based Nuclear DNA Markers for Multilocus Phylogeography of the Soybean-and Rice-Infecting Fungal Pathogen Rhizoctonia solani AG-1 IA. Genet. Mol. Biol. 2009, 32, 840–846. [Google Scholar] [CrossRef]
- Carling, D.E.; Kuninaga, S.; Brainard, K.A. Hyphal Anastomosis Reactions, RDNA-Internal Transcribed Spacer Sequences, and Virulence Levels among Subsets of Rhizoctonia solani Anastomosis Group-2 (AG-2) and AG-BI. Phytopathology 2002, 92, 43–50. [Google Scholar] [CrossRef]
- Fenille, R.C.; Ciampi, M.B.; Kuramae, E.E.; Souza, N.L. Identification of Rhizoctonia solani Associated with Soybean in Brazil by RDNA-ITS Sequences. Fitopatol. Bras. 2003, 28, 413–419. [Google Scholar] [CrossRef]
- Zachow, C.; Grosch, R.; Berg, G. Impact of Biotic and A-Biotic Parameters on Structure and Function of Microbial Communities Living on Sclerotia of the Soil-Borne Pathogenic Fungus Rhizoctonia solani. Appl. Soil Ecol. 2011, 48, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.S.; Doni, F.; Mispan, M.S.; Saiman, M.Z.; Yusuf, Y.M.; Oke, M.A.; Suhaimi, N.S.M. Harnessing Trichoderma in Agriculture for Productivity and Sustainability. Agronomy 2021, 11, 2559. [Google Scholar] [CrossRef]
- Kumari, N.; Katoch, S. Wilt and Root Rot Complex of Important Pulse Crops: Their Detection and Integrated Management. In Management of Fungal Pathogens in Pulses: Current Status and Future Challenges; Springer: Cham, Switzerland, 2020; pp. 93–119. [Google Scholar]
- Gaulin, E.; Jacquet, C.; Bottin, A.; Dumas, B. Root Rot Disease of Legumes Caused by Aphanomyces euteiches. Mol. Plant Pathol. 2007, 8, 539–548. [Google Scholar] [CrossRef]
- Myresiotis, C.K.; Karaoglanidis, G.S.; Tzavella-Klonari, K. Resistance of Botrytis cinerea Isolates from Vegetable Crops to Anilinopyrimidine, Phenylpyrrole, Hydroxyanilide, Benzimidazole, and Dicarboximide Fungicides. Plant Dis. 2007, 91, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Saba, H.; Vibhash, D.; Manisha, M.; Prashant, K.S.; Farhan, H.; Tauseef, A. Trichoderma—A Promising Plant Growth Stimulator and Biocontrol Agent. Mycosphere 2012, 3, 524–531. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuang, W.-Y. Trichoderma brevicrassum Strain TC967 with Capacities of Diminishing Cucumber Disease Caused by Rhizoctonia solani and Promoting Plant Growth. Biol. Control 2020, 142, 104151. [Google Scholar] [CrossRef]
- Singh, S.; Balodi, R. Bio-Management of Soil Borne Pathogens Infesting Cucumber (Cucumis Sativus L.) under Protected Cultivation System. Biol. Control 2021, 157, 104569. [Google Scholar] [CrossRef]
- Sánchez-Montesinos, B.; Santos, M.; Moreno-Gavíra, A.; Marín-Rodulfo, T.; Gea, F.J.; Diánez, F. Biological Control of Fungal Diseases by Trichoderma aggressivum f. europaeum and Its Compatibility with Fungicides. J. Fungi 2021, 7, 598. [Google Scholar] [CrossRef]
- Rini, C.R.; Sulochana, K.K. Management of Seedling Rot of Chilli (Capsicum annuum L.) Using Trichoderma spp. and Fluorescent pseudomonads (Pseudomonas fluorescens). J. Trop. Agric. 2006, 44, 79–82. [Google Scholar]
- Gallou, A.; Cranenbrouck, S.; Declerck, S. Trichoderma harzianum Elicits Defence Response Genes in Roots of Potato Plantlets Challenged by Rhizoctonia solani. Eur. J. Plant Pathol. 2009, 124, 219–230. [Google Scholar] [CrossRef]
- Fernandes, T.; Lopes, F.A.C.; Steindorff, A.S.; Brandao, R.S.; Jesuino, R.S.A.; Ulhoa, C.J. Mycoparasitism Studies of Trichoderma Species against Three Phytopathogenic Fungi: Evaluation of Antagonism and Hydrolytic Enzyme Production. Biotechnol. Lett. 2013, 35, 1461–1468. [Google Scholar]
- Howell, C.R. Understanding the Mechanisms Employed by Trichoderma virens to Effect Biological Control of Cotton Diseases. Phytopathology 2006, 96, 178–180. [Google Scholar] [CrossRef]
- Nawrocka, J.; Małolepsza, U. Diversity in Plant Systemic Resistance Induced by Trichoderma. Biol. Control 2013, 67, 149–156. [Google Scholar] [CrossRef]
- Nawrocka, J.; Gromek, A.; Małolepsza, U. Nitric Oxide as a Beneficial Signaling Molecule in Trichoderma atroviride TRS25-Induced Systemic Defense Responses of Cucumber Plants against Rhizoctonia solani. Front. Plant Sci. 2019, 10, 421. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, N.; Yong, X.; Yang, X.; Shen, Q. Biocontrol of Rhizoctonia solani Damping-off Disease in Cucumber with Bacillus pumilus SQR-N43. Microbiol. Res. 2012, 167, 135–143. [Google Scholar] [CrossRef]
- Verma, M.; Brar, S.K.; Tyagi, R.D.; Surampalli, R.Y.; Valero, J.R. Antagonistic Fungi, Trichoderma spp.: Panoply of Biological Control. Biochem. Eng. J. 2007, 37, 1–20. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational Research on Trichoderma: From’omics to the Field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, O.D.; Sinclair, J.B. Basic Plant Pathology Methods, 2nd ed.; Lewis Publishers: Boca Raton, FL, USA, 1995. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi; Burgess Publishing Company: Minneapolis, MN, USA, 1972. [Google Scholar]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A Plant DNA Minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.; Nnis, M.A.; Gelfand, D.H.; Sninsky, J.J. PCR Protocols-A Guide to Methods and Applications; The University of Michigan: Ann Arbor, MI, USA; Academic Press: Cambridge, UK, 1990. [Google Scholar]
- Misawa, T.; Kurose, D.; Shishido, K.; Toda, T.; Kuninaga, S. Characterization of a New Subgroup of Rhizoctonia solani Anastomosis Group 3 (AG-3 TM) Associated with Tomato Leaf Blight. J. Gen. Plant Pathol. 2020, 86, 457–467. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Hassan, N.; Elsharkawy, M.M.; Villajuan-Abgona, R.; Hyakumachi, M. A Nonpathogenic Species of Binucleate Rhizoctonia Inhibits the Formation of Infection Structures Caused by Rhizoctonia solani on Cucumber. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2015, 65, 208–214. [Google Scholar]
- Montealegre, J.; Valderrama, L.; Sánchez, S.; Herrera, R.; Besoain, X.; Pérez, L.M. Biological Control of Rhizoctonia solani in Tomatoes with Trichoderma harzianum Mutants. Electron. J. Biotechnol. 2010, 13, 1–2. [Google Scholar] [CrossRef]
- Ghosh, P.; Roy, A.; Hess, D.; Ghosh, A.; Das, S. Deciphering the Mode of Action of a Mutant Allium sativum Leaf Agglutinin (MASAL), a Potent Antifungal Protein on Rhizoctonia solani. BMC Microbiol. 2015, 15, 1–16. [Google Scholar] [CrossRef]
- Elad, Y.; Chet, I.; Katan, J. Trichoderma harzianum: A Biocontrol Agent Effective against Sclerotium rolfsii and Rhizoctonia solani. Phytopathology 1980, 70, 119–121. [Google Scholar] [CrossRef]
- Ketta, H.A.; Elkhateeb, N.M.; Saleh, M.M.; Kamel, S.M. Efficiency Assessment of Combinations between Rhizobium leguminosarum and Trichoderma spp. for Controlling of Pea (Pisum sativum L.) Damping-off Disease. Egypt. J. Phytopathol. 2021, 49, 1–14. [Google Scholar] [CrossRef]
- Ismail, A.M. Efficacy of Copper Oxide and Magnesium Oxide Nanoparticles on Controlling Black Scurf Disease on Potato. Egypt. J. Phytopathol. 2021, 49, 116–130. [Google Scholar] [CrossRef]
- Ismail, A.M.; El-Gawad, A.; Mona, E. Antifungal Activity of MgO and ZnO Nanoparticles against Powdery Mildew of Pepper under Greenhouse Conditions. Egypt. J. Agric. Res. 2021, 99, 421–434. [Google Scholar]
- Matta, A.; Dimond, A.E. Symptoms of Fusarium Wilt in Relation to Quantity of Fungus and Enzyme Activity in Tomato Stems. Phytopathology 1963, 53, 574. [Google Scholar]
- Allam, A.I.; Hollis, J.P. Sulfide Inhibition of Oxidases in Rice Roots. Phytopathology 1972, 62, 634–639. [Google Scholar] [CrossRef]
- Aebi, H.E. Catalase. In Methods of Enzymatic Analysis; Academic Press: Cambridge, MA, USA, 1983; pp. 273–286. [Google Scholar]
- Wirth, S.J.; Wolf, G.A. Dye-Labelled Substrates for the Assay and Detection of Chitinase and Lysozyme Activity. J. Microbiol. Methods 1990, 12, 197–205. [Google Scholar] [CrossRef]
- Snell, F.D.; Snell, C.T. Colorimetric Methods; D. Van Nostrand Co. Inc.: Toronto, ON, Canada; New York, NY, USA; London, UK, 1953; Volume III, 606p. [Google Scholar]
- Nagata, M.; Yamashita, I. Simple Method for Simultaneous Determination of Chlorophyll and Carotenoids in Tomato Fruit. Nippon. Shokuhin Kogyo Gakkaishi 1992, 39, 925–928. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Erper, I.; Hatat Karaca, G.; Özkoc, I. Characterization of Rhizoctonia species Causing Root-Rot of Cucumber Plants Grown in Greenhouses in Samsun, Turkey. In II Balkan Symposium on Vegetables and Potatoes; ISHS: Thessaloniki, Greece, 2000; pp. 531–534. [Google Scholar]
- Eken, C.; Tuncer, S. Rhizoctonia Species and Anastomosis Groups Isolated from Tomato and Cucumber in Erzincan, Turkey. Int. J. Res. Agric. For. 2019, 6, 26–31. [Google Scholar]
- Ersahin, Y.S.; Haktanir, K.; Yanar, Y. Vermicompost Suppresses Rhizoctonia solani Kühn in Cucumber Seedlings/Wurmkompost Unterdrückt Die Entwicklung von Rhizoctonia solani Kühn an Gurkenkeimlingen. J. Plant Dis. Prot. 2009, 116, 182–188. [Google Scholar] [CrossRef]
- Yıldırım, E.; Erper, I. Characterization and Pathogenicity of Rhizoctonia spp. Isolated from Vegetable Crops Grown in Greenhouses in Samsun Province, Turkey. Biosci. J. 2017, 33, 257–267. [Google Scholar]
- Mirmajlessi, S.M.; Safaie, N.; Mostafavi, H.A.; Mansouripour, S.M.; Mahmoudy, S.B. Genetic Diversity among Crown and Root Rot Isolates of Rhizoctonia solani Isolated from Cucurbits Using PCR Based Techniques. Afr. J. Agric. Res. 2012, 7, 583–590. [Google Scholar]
- Desvani, S.D.; Lestari, I.B.; Wibowo, H.R.; Supyani; Poromarto, S.H. Hadiwiyono Morphological Characteristics and Virulence of Rhizoctonia solani Isolates Collected from Some Rice Production Areas in Some Districts of Central Java. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2018; Volume 2014, p. 20068. [Google Scholar]
- Mukhopadhyay, A. Biological control of soil borne plant pathogens by Trichoderma spp. Indian J. Mycol. Plant Pathol. 1987, 17, 1–10. [Google Scholar]
- Ryu, J.-Y.; Jin, R.-D.; Kim, Y.-W.; Lee, H.-B.; Kim, K.-Y. Biocontrol of Damping-off (Rhizoctonia solani) in Cucumber by Trichoderma asperellum T-5. Korean J. Soil Sci. Fertil. 2006, 39, 185–194. [Google Scholar]
- Hassan, M.; Ahmed, H.; Kamel, S.; El-Hamed, A.; Yousef, H. Biological Control of Damping-off and Root Rot Disease Caused by Rhizoctonia solani on Cucumber Plants. Fayoum J. Agric. Res. Dev. 2021, 35, 525–541. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, B.; Zhang, J.; Gan, Y. Identification of the Antifungal Activity of Trichoderma longibrachiatum T6 and Assessment of Bioactive Substances in Controlling Phytopathgens. Pestic. Biochem. Physiol. 2018, 147, 59–66. [Google Scholar] [CrossRef]
- Trillas, M.I.; Casanova, E.; Cotxarrera, L.; Ordovás, J.; Borrero, C.; Avilés, M. Composts from Agricultural Waste and the Trichoderma asperellum Strain T-34 Suppress Rhizoctonia solani in Cucumber Seedlings. Biol. Control 2006, 39, 32–38. [Google Scholar] [CrossRef]
- Huang, X.; Chen, L.; Ran, W.; Shen, Q.; Yang, X. Trichoderma harzianum Strain SQR-T37 and Its Bio-Organic Fertilizer Could Control Rhizoctonia solani Damping-off Disease in Cucumber Seedlings Mainly by the Mycoparasitism. Appl. Microbiol. Biotechnol. 2011, 91, 741–755. [Google Scholar] [CrossRef] [PubMed]
- Inbar, J.; Menendez, A.N.A.; Chet, I. Hyphal Interaction between Trichoderma harzianum and Sclerotinia sclerotiorum and Its Role in Biological Control. Soil Biol. Biochem. 1996, 28, 757–763. [Google Scholar] [CrossRef]
- De Melo, I.S.; Faull, J.L. Parasitism of Rhizoctonia solani by Strains of Trichoderma spp. Sci. Agric. 2000, 57, 55–59. [Google Scholar] [CrossRef]
- El-Kazzaz, M.K.; Ghoneim, K.E.; Agha, M.K.M.; Helmy, A.; Behiry, S.I.; Abdelkhalek, A.; Saleem, M.H.; Al-Askar, A.A.; Arishi, A.A.; Elsharkawy, M.M. Suppression of Pepper Root Rot and Wilt Diseases Caused by Rhizoctonia solani and Fusarium oxysporum. Life 2022, 12, 587. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Ashraf, S. Role of Trichoderma spp. as a Biocontrol Agent of Fungal Plant Pathogens. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 497–506. ISBN 978-981-10-3473-2. [Google Scholar]
- Nawrocka, J.; Małolepsza, U.; Szymczak, K.; Szczech, M. Involvement of Metabolic Components, Volatile Compounds, PR Proteins, and Mechanical Strengthening in Multilayer Protection of Cucumber Plants against Rhizoctonia solani Activated by Trichoderma atroviride TRS25. Protoplasma 2018, 255, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Konappa, N.; Krishnamurthy, S.; Siddaiah, C.N.; Ramachandrappa, N.S.; Chowdappa, S. Evaluation of Biological Efficacy of Trichoderma asperellum against Tomato Bacterial Wilt Caused by Ralstonia solanacearum. Egypt. J. Biol. Pest Control. 2018, 28, 63. [Google Scholar] [CrossRef]
- Yedidia, I.; Shoresh, M.; Kerem, Z.; Benhamou, N.; Kapulnik, Y.; Chet, I. Concomitant Induction of Systemic Resistance to Pseudomonas syringae Pv. lachrymans in Cucumber by Trichoderma asperellum (T-203) and Accumulation of Phytoalexins. Appl. Environ. Microbiol. 2003, 69, 7343–7353. [Google Scholar] [CrossRef]
- Taranto, F.; Pasqualone, A.; Mangini, G.; Tripodi, P.; Miazzi, M.M.; Pavan, S.; Montemurro, C. Polyphenol Oxidases in Crops: Biochemical, Physiological and Genetic Aspects. Int. J. Mol. Sci. 2017, 18, 377. [Google Scholar] [CrossRef]
- Behiry, S.; Soliman, S.A.; Massoud, M.A.; Abdelbary, M.; Kordy, A.M.; Abdelkhalek, A.; Heflish, A. Trichoderma pubescens Elicit Induced Systemic Resistance in Tomato Challenged by Rhizoctonia solani. J. Fungi 2023, 9, 167. [Google Scholar] [CrossRef]
- Singh, B.N.; Singh, A.; Singh, S.P.; Singh, H.B. Trichoderma harzianum-Mediated Reprogramming of Oxidative Stress Response in Root Apoplast of Sunflower Enhances Defence against Rhizoctonia solani. Eur. J. Plant Pathol. 2011, 131, 121–134. [Google Scholar] [CrossRef]
- Han, H.S.; Lee, K.D. Plant Growth Promoting Rhizobacteria Effect on Antioxidant Status, Photosynthesis, Mineral Uptake and Growth of Lettuce under Soil Salinity. Res. J. Agric. Biol. Sci. 2005, 1, 210–215. [Google Scholar]
- Akladious, S.A.; Abbas, S.M. Application of Trichoderma harziunum T22 as a Biofertilizer Supporting Maize Growth. Afr. J. Biotechnol. 2012, 11, 8672–8683. [Google Scholar]
- Abbas, A.; Mubeen, M.; Zheng, H.; Sohail, M.A.; Shakeel, Q.; Solanki, M.K.; Iftikhar, Y.; Sharma, S.; Kashyap, B.K.; Hussain, S. Trichoderma spp. Genes Involved in the Biocontrol Activity against Rhizoctonia solani. Front. Microbiol. 2022, 13, 884469. [Google Scholar] [CrossRef] [PubMed]
- Doni, F.; Fathurrahman, F.; Mispan, M.S.; Suhaimi, N.S.M.; Yusoff, W.M.W.; Uphoff, N. Transcriptomic Profiling of Rice Seedlings Inoculated with the Symbiotic Fungus Trichoderma asperellum SL2. J. Plant Growth Regul. 2019, 38, 1507–1515. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | AGs | Pre-Damping off % | Post-Damping off % | Survival % | DI % |
|---|---|---|---|---|---|
| PPDURS1 | AG-4 | 80.00 a * | 16.67 cd | 3.33 ef | 33.33 bcd |
| PPDURS2 | 76.67 ab | 23.33 bcd | 0.00 f | 0.00 d | |
| PPDURS3 | 66.67 ab | 23.33 bcd | 10.00 def | 66.67 abc | |
| PPDURS4 | 70.00 ab | 16.67 cd | 13.33 cde | 16.67 cd | |
| PPDURS5 | 63.33 bc | 13.33 de | 23.33 c | 61.11 abc | |
| PPDURS6 | 50.00 c | 30.00 b | 20.00 cd | 100.00 a | |
| PPDURS7 | 33.33 d | 26.67 bc | 40.00 b | 70.00 ab | |
| PPDURS8 | 26.67 d | 50.00 a | 23.33 c | 100.00 a | |
| PPDURS9 | AG-A | 3.33 e | 3.33 ef | 93.34 a | 0.00 d |
| PPDU RS10 | AG-2-1 | 0.00 e | 0.00 f | 100.00 a | 0.00 d |
| Healthy control | 0.00 e | 0.00 f | 100.00 a | 0.00 d | |
| L.S.D ≥ 5 | 14.97 | 11.176 | 11.749 | 52.79 | |
| Treatments | Pre Damping off % | Post Damping off % | Efficacy % | Survival % | DI % |
|---|---|---|---|---|---|
| T. hamatum KSATR8 | 40 b * | 26.67 ab | 33.33 | 33.33 e | 22.22 abc |
| T. harzianum KSATR9 | 36.67 b | 30.00 ab | 33.33 | 33.33 e | 30.55 ab |
| T. harzianum KSATR10 | 16.67 c | 26.67 ab | 56.67 | 56.67 d | 17.78 bc |
| T. asperellum KSATR11 | 50.00 b | 40.00 a | 10 | 13.33 g | 16.67 bc |
| T. asperellum KSATC | 10.00 cd | 13.33 bcd | 80 | 76.67 c | 4.76 c |
| T. asperellum KSAT1E | 13.33 cd | 23.33 abc | 63.34 | 63.34 d | 9.52 bc |
| T. longibrachiatum KSATS | 36.67 a | 40.00 a | 23.33 | 23.33 f | 44.44 a |
| Rizolex-T 50% | 6.67 cd | 6.67 cd | 86.67 | 86.67 b | 3.70 c |
| Infected control | 86.667 a | 13.33 bcd | 0 | 0.00 f | 0.00 c |
| Healthy control | 0 d | 0.00 d | 0 | 100.00 a | 0.00 c |
| L.S.D ≥ 5 | 13.97 | 18.36 | - | 9.95 | 24.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almaghasla, M.I.; El-Ganainy, S.M.; Ismail, A.M. Biological Activity of Four Trichoderma Species Confers Protection against Rhizoctonia solani, the Causal Agent of Cucumber Damping-Off and Root Rot Diseases. Sustainability 2023, 15, 7250. https://doi.org/10.3390/su15097250
Almaghasla MI, El-Ganainy SM, Ismail AM. Biological Activity of Four Trichoderma Species Confers Protection against Rhizoctonia solani, the Causal Agent of Cucumber Damping-Off and Root Rot Diseases. Sustainability. 2023; 15(9):7250. https://doi.org/10.3390/su15097250
Chicago/Turabian StyleAlmaghasla, Mustafa I., Sherif Mohamed El-Ganainy, and Ahmed Mahmoud Ismail. 2023. "Biological Activity of Four Trichoderma Species Confers Protection against Rhizoctonia solani, the Causal Agent of Cucumber Damping-Off and Root Rot Diseases" Sustainability 15, no. 9: 7250. https://doi.org/10.3390/su15097250
APA StyleAlmaghasla, M. I., El-Ganainy, S. M., & Ismail, A. M. (2023). Biological Activity of Four Trichoderma Species Confers Protection against Rhizoctonia solani, the Causal Agent of Cucumber Damping-Off and Root Rot Diseases. Sustainability, 15(9), 7250. https://doi.org/10.3390/su15097250