

Valorization of Human Urine with Mixed Microalgae Examined through Population Dynamics, Nutrient Removal, and Biogas Content

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Urine Collection
2.2. Preparation of Microalgal Inoculum
2.3. Experimental Set Up
2.4. Biomethane Potential (BMP) Assay
2.5. Analytical Methods
2.6. Microbial Community Analysis
2.7. Principal Component Analysis (PCA)
3. Results and Discussion
3.1. Monitoring of Physical and Chemical Conditions during the Operation
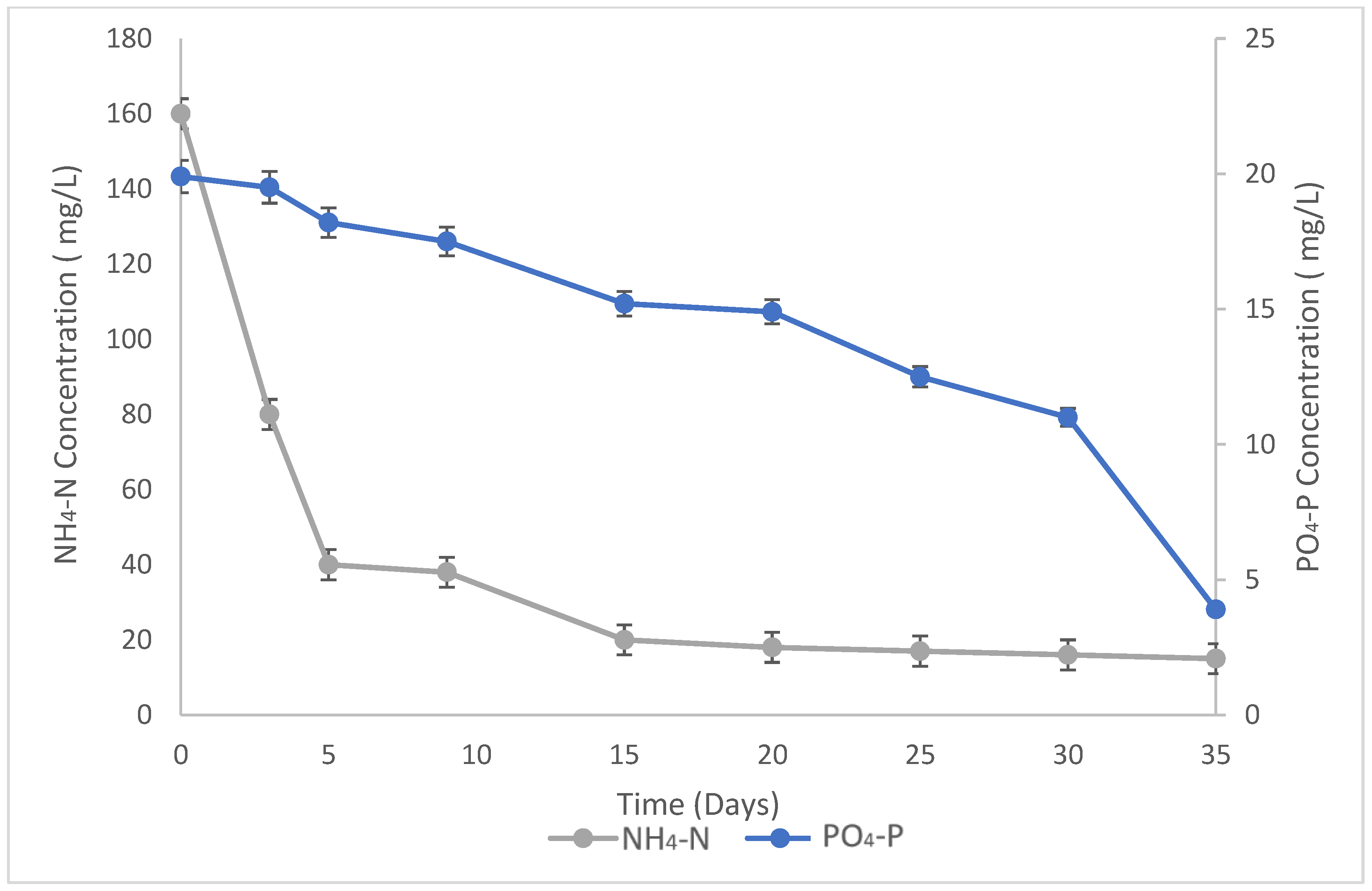
3.2. Growth and Nutrient Removal Results in Raceway Ponds
3.3. Biogas Results
3.4. Small-Scale vs. Large-Scale Genetic Comparison Results along with Synthetic Media
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baykal, B.B. Recycling/reusing grey water and yellow water (human urine): Motivations, perspectives and reflections into the future. Desalin. Water Treat. 2019, 172, 212–223. [Google Scholar] [CrossRef]
- Tuantet, K.; Janssen, M.; Temmink, H.; Zeeman, G.; Wijffels, R.H.; Buisman, C.J.N. Microalgae growth on concentrated human urine. J. Appl. Phycol. 2013, 26, 287–297. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Bui, X.T.; Ngo, H.H.; Nguyen, K.Q.; Nguyen, H.H.; Némery, J.; Varjani, S. Nutrient recovery and micro-algae biomass production from urine by membrane photobioreactor at low biomass retention times. Sci. Total Environ. 2021, 785, 147423. [Google Scholar] [CrossRef]
- Huesemann, M.; Chavis, A.; Edmundson, S.; Rye, D.; Hobbs, S.; Sun, N.; Wigmosta, M. Climate-simulated raceway pond culturing: Quantifying the maximum achievable annual biomass productivity of Chlorella sorokiniana in the contiguous USA. J. Appl. Phycol. 2017, 30, 287–298. [Google Scholar] [CrossRef]
- Aditya, L.; Mahlia, T.I.; Nguyen, L.N.; Vu, H.P.; Nghiem, L.D. Microalgae-bacteria consortium for wastewater treatment and biomass production. Sci. Total Environ. 2022, 838, 155871. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hu, Z.; Qi, Y.; Song, C.; Chen, G. The interactions of algae-activated sludge symbiotic system and its effects on wastewater treatment and lipid accumulation. Bioresour. Technol. 2019, 292, 122017. [Google Scholar] [CrossRef]
- Bankston, E.; Wang, Q.; Higgins, B.T. Algae support populations of heterotrophic, nitrifying, and phos-phate-accumulating bacteria in the treatment of poultry litter anaerobic digestate. Chem. Eng. J. 2020, 398, 125550. [Google Scholar] [CrossRef]
- Maza-Márquez, P.; González-Martínez, A.; Juárez-Jiménez, B.; Rodelas, B.; González-López, J. Microalgae-Bacteria Consortia for the removal of phenolic compounds from industrial wastewaters. In Approaches in Bioremediation: The New Era of Environmental Microbiology and Nanobiotechnology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 135–184. [Google Scholar] [CrossRef]
- Xia, A.; Cheng, J.; Murphy, J.D. Innovation in biological production and upgrading of methane and hydrogen for use as gaseous transport biofuel. Biotechnol. Adv. 2016, 34, 451–472. [Google Scholar] [CrossRef]
- Rawat, I.; Kumar, R.R.; Mutanda, T.; Bux, F. Dual role of microalgae: Phycoremediation of domestic wastewater and biomass production for sustainable biofuels production. Appl. Energy 2011, 88, 3411–3424. [Google Scholar] [CrossRef]
- Mishra, S.; Mohanty, K. Co-HTL of domestic sewage sludge and wastewater treatment derived microalgal biomass—An integrated biorefinery approach for sustainable biocrude production. Energy Convers. Manag. 2020, 204, 112312. [Google Scholar] [CrossRef]
- Muñoz, R.; Gonzalez-Fernandez, C. (Eds.) Microalgae-Based Biofuels and Bioproducts: From Feedstock Cultivation to End-Products; Woodhead Publishing: Sawston, UK, 2017. [Google Scholar]
- Ermis, H.; Güven-Gülhan, Ü.; Çakır, T.; Altınbaş, M. Microalgae growth and diversity in anaerobic digestate compared to synthetic media. Biofuel Res. J. 2022, 9, 1551–1561. [Google Scholar] [CrossRef]
- Barsanti, L.; Gualteri, P. Algae-Anatomy, Biochemistry and Biotechnology; Taylor &Francis: Oxfordshire, UK, 2006. [Google Scholar]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae-Identification and Use as Bioindicators, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010. [Google Scholar]
- APHA (American Public Health Association). Standard Methods for the Examination of Water and Wastewater, 21st ed.; APHA: Washington, DC, USA, 2005. [Google Scholar]
- Ebeling, J.M.; Timmons, M.B.; Bisogni, J.J. Engineering analysis of the stoichiometry of photoautotrophic, auto-trophic, and heterotrophic removal of ammonia–nitrogen in aquaculture systems. Aquaculture 2006, 257, 346–358. [Google Scholar] [CrossRef]
- Chatterjee, P.; Granatier, M.; Ramasamy, P.; Kokko, M.; Lakaniemi, A.-M.; Rintala, J. Microalgae grow on source separated human urine in Nordic climate: Outdoor pilot-scale cultivation. J. Environ. Manag. 2019, 237, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Zou, G.; Liu, Y.; Zhang, Q.; Zhou, T.; Xiang, S.; Gu, Z.; Huang, Q.; Yan, H.; Zheng, H.; Wu, X.; et al. Cultivation of Chlorella vulgaris in a Light-Receiving-Plate (LRP)-Enhanced Raceway Pond for Ammonium and Phosphorus Removal from Pretreated Pig Urine. Energies 2020, 13, 1644. [Google Scholar] [CrossRef]
- Saarnio, S. Nutrient Recovery From Source-Separated Human Urine by Microalgae in Continuously Fed Raceway Pond. Master’s Thesis, Tampere University, Tampere, Finland, 2019. [Google Scholar]
- Cai, T.; Park, S.Y.; Li, Y. Nutrient recovery from wastewater streams by microalgae: Status and prospects. Renew. Sustain. Energy Rev. 2013, 19, 360–369. [Google Scholar] [CrossRef]
- Gentili, F.G. Microalgal biomass and lipid production in mixed municipal, dairy, pulp and paper wastewater together with added flue gases. Bioresour. Technol. 2014, 169, 27–32. [Google Scholar] [CrossRef]
- Chiu, S.-Y.; Kao, C.-Y.; Chen, T.-Y.; Chang, Y.-B.; Kuo, C.-M.; Lin, C.-S. Cultivation of microalgal Chlorella for biomass and lipid production using wastewater as nutrient resource. Bioresour. Technol. 2015, 184, 179–189. [Google Scholar] [CrossRef]
- Purba, L.D.; Abdullah, N.; Yuzir, A.; Zamyadi, A.; Shimizu, K.; Hermana, J. Rapid development of microal-gae-bacteria granular sludge using low-strength domestic wastewater. J. Water Environ. Technol. 2021, 19, 96–107. [Google Scholar] [CrossRef]
- Barreiro-Vescovo, S.; González-Fernández, C.; de Godos, I. Characterization of communities in a microalgae-bacteria system treating domestic wastewater reveals dominance of phototrophic and pigmented bacteria. Algal Res. 2021, 59, 102447. [Google Scholar] [CrossRef]
- Fan, H.; Wang, K.; Wang, C.; Yu, F.; He, X.; Ma, J.; Li, X. A comparative study on growth characters and nutrients removal from wastewater by two microalgae under optimized light regimes. Environ. Technol. Innov. 2020, 19, 100849. [Google Scholar] [CrossRef]
- Biswas, T.; Bhushan, S.; Prajapati, S.K.; Chaudhuri, S.R. An eco-friendly strategy for dairy wastewater remediation with high lipid microalgae-bacterial biomass production. J. Environ. Manag. 2021, 286, 112196. [Google Scholar] [CrossRef] [PubMed]
- Makut, B.B.; Das, D.; Goswami, G. Production of microbial biomass feedstock via co-cultivation of microal-gae-bacteria consortium coupled with effective wastewater treatment: A sustainable approach. Algal Res. 2019, 37, 228–239. [Google Scholar] [CrossRef]
- Mohd Udaiyappan, A.F.; Hasan, H.A.; Takriff, M.S.; Abdullah, S.R.S.; Maeda, T.; Mustapha, N.A.; Mohd Yasin, N.; Nazashida Mohd Hakimi, N.I. Microalgae-bacteria interaction in palm oil mill effluent treatment. J. Water Process Eng. 2020, 35, 101203. [Google Scholar] [CrossRef]
- Passos, F.; Cordeiro, P.H.M.; Baeta, B.; de Aquino, S.F.; Perez-Elvira, S.I. Anaerobic co-digestion of coffee husks and microalgal biomass after thermal hydrolysis. Bioresour. Technol. 2018, 253, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Posadas, E.; Bochon, S.; Coca, M.; García-González, M.; García-Encina, P.; Muñoz, R. Microalgae-based agro-industrial wastewater treatment: A preliminary screening of biodegradability. J. Appl. Phycol. 2014, 26, 2335–2345. [Google Scholar] [CrossRef]
- Hemalatha, M.; Sravan, J.S.; Min, B.; Mohan, S.V. Microalgae-biorefinery with cascading resource recovery design associated to dairy wastewater treatment. Bioresour. Technol. 2019, 284, 424–429. [Google Scholar] [CrossRef]
- Gramegna, G.; Scortica, A.; Scafati, V.; Ferella, F.; Gurrieri, L.; Giovannoni, M.; Bassi, R.; Sparla, F.; Mattei, B.; Benedetti, M. Exploring the potential of microalgae in the recycling of dairy wastes. Bioresour. Technol. Rep. 2020, 12, 100604. [Google Scholar] [CrossRef]
- Talapatra, N.; Gautam, R.; Mittal, V.; Ghosh, U.K. A comparative study of the growth of microalgae-bacteria symbiotic consortium with the axenic culture of microalgae in dairy wastewater through extraction and quantification of chlorophyll. Mater. Today Proc. 2021, in press. [CrossRef]
- de la Lama-Calvente, D.; Cubero, J.; Fernández-Rodríguez, M.J.; Jiménez-Rodríguez, A.; Borja, R. Anaerobic co-digestion of microalgae and industrial wastes: A critical and bibliometric review. In Progress in Microalgae Research—A Path for Shaping Sustainable Futures; Intech Open: London, UK, 2022. [Google Scholar] [CrossRef]
- Böjti, T.; Kovács, K.L.; Kakuk, B.; Wirth, R.; Rákhely, G.; Bagi, Z. Pretreatment of poultry manure for efficient biogas production as monosubstrate or co-fermentation with maize silage and corn stover. Anaerobe 2017, 46, 138–145. [Google Scholar] [CrossRef]
- Béji, O.; Adouani, N.; Poncin, S.; Li, H.-Z. Growth of Microalgae-Bacteria Flocs for Nutrient Recycling from Digestate and Liquid Slurry and Methane Production by Anaerobic Digestion. Appl. Sci. 2022, 12, 7634. [Google Scholar] [CrossRef]
- Mahdy, A.; Fotidis, I.A.; Mancini, E.; Ballesteros, M.; González-Fernández, C.; Angelidaki, I. Ammonia tolerant inocula provide a good base for anaerobic digestion of microalgae in third generation biogas process. Bioresour. Technol. 2017, 225, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Arashiro, L.T.; Ferrer, I.; Pániker, C.C.; Gómez-Pinchetti, J.L.; Rousseau, D.P.L.; Van Hulle, S.W.H.; Garfí, M. Natural pigments and biogas recovery from Microalgae Grown in Wastewater. ACS Sustain. Chem. Eng. 2020, 8, 10691–10701. [Google Scholar] [CrossRef] [PubMed]
- Solé-Bundó, M.; Salvadó, H.; Passos, F.; Garfí, M.; Ferrer, I. Strategies to optimize microalgae conversion to biogas: Co-Digestion, pretreatment and Hydraulic Retention Time. Molecules 2018, 23, 2096. [Google Scholar] [CrossRef]
- Vieira, H.H.; Bagatini, I.; Guinart, C.M.; Vieira, A.A.H. tufA gene as molecular marker for freshwater Chlorophyceae. Algae 2016, 31, 155–165. [Google Scholar] [CrossRef]
- Malavasi, V.; Costelli, C.; Orsini, M.; Cusano, R.; Angius, A.; Cao, G. Deep genomic analysis of the Chlorella sorokiniana SAG 211-8k chloroplast. Eur. J. Phycol. 2017, 52, 320–329. [Google Scholar] [CrossRef]
- Baytut, Ö.; Gürkanli, C.T.; Gönülol, A.; Özkoç, I. Molecular phylogeny of Chlorella-related chlorophytes (Chlorophyta) from Anatolian freshwaters of Turkey. Turk. J. Bot. 2014, 38, 600–607. [Google Scholar] [CrossRef]
- Burja, M.A.; Tamagnini, P.; Bustard, M.T.; Wright, P.C. Identification of the green alga, Chlorella vulgaris (SDC1) using cya-nobacteria derived 16S rDNA primers: Targeting the chloroplast. FEMS Microbiol. Lett. 2001, 202, 195–203. [Google Scholar] [CrossRef]
- Wang, F.; Men, X.; Zhang, G.; Liang, K.; Xin, Y.; Wang, J.; Li, A.; Zhang, H.; Liu, H.; Wu, L. Assessment of 16S rRNA gene primers for studying bacterial community structure and function of aging flue-cured tobaccos. AMB Express 2018, 8, 182. [Google Scholar] [CrossRef]
- Hanshew, A.S.; Mason, C.J.; Raffa, K.F.; Currie, C.R. Minimization of chloroplast contamination in 16S rRNA gene pyrosequencing of insect herbivore bacterial communities. J. Microbiol. Methods 2013, 95, 149–155. [Google Scholar] [CrossRef]
- Lage, S.; Gentili, F.G. Chemical composition and species identification of microalgal biomass grown at pilot-scale with mu-nicipal wastewater and CO2 from flue gases. Chemosphere 2023, 313, 137344. [Google Scholar] [CrossRef]
- Carney, L.T.; Lane, T.W. Parasites in algae mass culture. Front. Microbiol. 2014, 5, 278. [Google Scholar] [CrossRef] [PubMed]
- Vallet, M.; Baumeister, T.U.H.; Kaftan, F.; Grabe, V.; Buaya, A.; Thines, M.; Svatoš, A.; Pohnert, G. The oomycete Lagenisma coscinodisci hijacks host alkaloid synthesis during infection of a marine diatom. Nat. Commun. 2019, 10, 4938. [Google Scholar] [CrossRef] [PubMed]
- Sauvage, T.; Schmidt, W.E.; Suda, S.; Fredericq, S. A metabarcoding framework for facilitated survey of endolithic phototrophs with tufa. BMC Ecol. 2016, 16, 1–21. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 10% Urine | |
|---|---|
| pH | 9.5 |
| PO4-P (mg/L) | 19.9 |
| NH4-N (mg/L) | 160 |
| Mixed liquor suspended solids (mg/L) | 400 |
| BMP 1 | BMP 2 | BLANK | |
|---|---|---|---|
| Microalgae Biomass (g) | 4 | 6 | - |
| Cow Manure (g) | 12 | 12 | 12 |
| Biogas Yield (L/kg VS) | 254.96 | 196.38 | 35.71 |
| PO4 Removal (%) | NH4 Removal (%) | Biomass Production (g/L) | |
|---|---|---|---|
| Urine treatment by mixed microalgae | 80.4 | 90.6 | 1.84 |
| Parameters | 10% Urine |
|---|---|
| Experimental observed biomass (mg/L) | 1840 |
| Experimental PO4-P removal (mg/L) | 16 |
| Experimental NH4-N removal (mg/L) | 145 |
| Theoretical PO4-P removal according to experimentally observed biomass (mg PO4-P L-1) | 26 |
| Theoretical NH4-N removal according to experimentally observed biomass (mg NH4-N L-1) | 136 |
| % Abundance | |||||
|---|---|---|---|---|---|
| Marker Genes | Phylum | Family/Genus/Species | Large-Scale Urine Sample | Lab-Scale Synthetic Media (BBM) Sample | Lab-Scale Urine Sample |
| 16S rRNA | Chlorophyta | Chlorella sorokiniana chloroplast | 27.94 | 0.39 | 4.71 |
| Chlorophyta | Chlorella sorokiniana mitochondrion | 16.44 | 0.41 | 4.55 | |
| Cyanobacteria | Synechocystis PCC-6803 | 0.15 | 23.62 | 0.18 | |
| Cyanobacteria | Cyanobium PCC-6307 | 0.04 | 12.31 | 0.04 | |
| Gemmatimonadota | Gemmatimonas | 0.14 | 11.55 | 0.25 | |
| Bacteroidota | Aurantisolimonas | 0.03 | 4.63 | 0.03 | |
| Proteobacteria | Reyranella | 0.03 | 3.67 | 0.04 | |
| Cyanobacteria | Leptolyngbya PCC-6306 | 0.02 | 2.23 | 0.02 | |
| Chlorophyta | Desmodesmus sp. | 0.12 | 0.06 | 23.79 | |
| Chlorophyta | Coelastrella sp. | 0.21 | 0.09 | 15.20 | |
| Proteobacteria | Porphyrobacter | 0.81 | 1.64 | 8.33 | |
| Chlorophyta | Chloroplast | 0.42 | 1.08 | 6.87 | |
| Proteobacteria | Sphingomonas piscinae | 0.02 | 0.03 | 3.89 | |
| Proteobacteria | Roseomonas stagni | 0 | 0.02 | 2.70 | |
| Proteobacteria | Ahniella | 0 | 1.01 | 2.46 | |
| Planctomycetota | Blastopirellula | 0.02 | 0.54 | 2.16 | |
| 18S rRNA | Chlorophyta | Trebouxiophyceae | 47.92 | 18.38 | 2.87 |
| Blastocladiomycota | Paraphysoderma sedebokerense | 17.87 | 12.62 | 0.26 | |
| Amoebozoa | Dactylopodida | 2.70 | 0.02 | 0.02 | |
| Chlorophyta | Chlorophyceae | 2.32 | 8.12 | 62.29 | |
| Ochrophyta | Spumella-like flagellate | 0.06 | 13.37 | 0.04 | |
| Cryptomycota | Paramicrosporidium | 0.40 | 10.16 | 0.53 | |
| Collodictyonidae | Diphylleia rotans | 0.47 | 5.01 | 0.40 | |
| Ciliophora | Cyclidium | 0 | 0.08 | 9.94 | |
| Ciliophora | Telotrochidium | 0.03 | 0.41 | 2.66 | |
| Chlorophyta | Chlamydopodium starrii | 0.10 | 0.20 | 2.39 | |
| Ciliophora | Opisthonecta | 0.09 | 0.15 | 2.07 | |
| 23S rRNA | Chlorophyta | Chlorella sorokiniana chloroplast | 75.45 | 0.62 | 9.72 |
| Chlorophyta | Chloroplast | 2.88 | 1.47 | 53.21 | |
| Proteobacteria | Acetobacteraceae | 10.63 | 1.24 | 5.64 | |
| Cyanobacteria | Synechocystis PCC-6803 | 0.81 | 49.94 | 0.46 | |
| Cyanobacteria | Cyanobium gracile | 0.19 | 37.30 | 0.17 | |
| Cyanobacteria | Leptolyngbya boryana | 0.12 | 3.93 | 0.10 | |
| Proteobacteria | Roseomonas stagni | 0.39 | 0.11 | 12.29 | |
| Bacteroidota | Mariniradius saccharolyticus | 0.18 | 0.07 | 10.83 | |
| Proteobacteria | Brevundimonas | 0.96 | 0.09 | 2.13 | |
| tufA | Chlorophyta | Chlorella sorokiniana | 93.94 | 26.94 | 13.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ermis, H.; Guven Gulhan, U.; Akca, M.S.; Cakir, T.; Altinbas, M. Valorization of Human Urine with Mixed Microalgae Examined through Population Dynamics, Nutrient Removal, and Biogas Content. Sustainability 2023, 15, 6922. https://doi.org/10.3390/su15086922
Ermis H, Guven Gulhan U, Akca MS, Cakir T, Altinbas M. Valorization of Human Urine with Mixed Microalgae Examined through Population Dynamics, Nutrient Removal, and Biogas Content. Sustainability. 2023; 15(8):6922. https://doi.org/10.3390/su15086922
Chicago/Turabian StyleErmis, Hande, Unzile Guven Gulhan, Mehmet Sadik Akca, Tunahan Cakir, and Mahmut Altinbas. 2023. "Valorization of Human Urine with Mixed Microalgae Examined through Population Dynamics, Nutrient Removal, and Biogas Content" Sustainability 15, no. 8: 6922. https://doi.org/10.3390/su15086922
APA StyleErmis, H., Guven Gulhan, U., Akca, M. S., Cakir, T., & Altinbas, M. (2023). Valorization of Human Urine with Mixed Microalgae Examined through Population Dynamics, Nutrient Removal, and Biogas Content. Sustainability, 15(8), 6922. https://doi.org/10.3390/su15086922