
Modelling Distributions of Asian and African Rice Based on MaxEnt

Abstract
1. Introduction
2. Materials and Methods
2.1. Crop Landrace Study Areas
2.2. Occurrence Records
2.3. Environmental Predictors
2.4. Landrace Distribution Modelling
3. Results
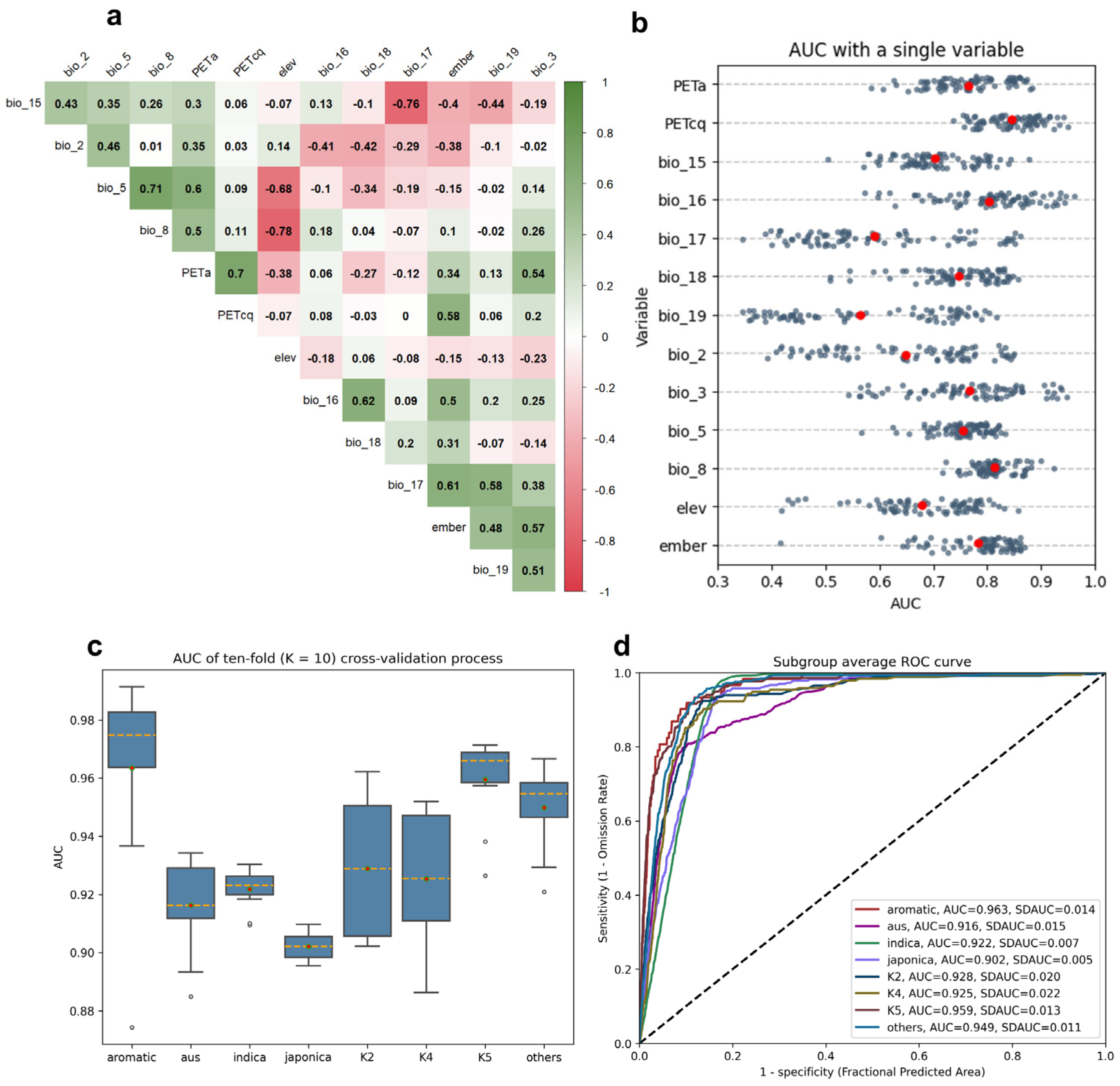
3.1. Modelling Validation
3.2. Predicted Distributions of Rice Landraces
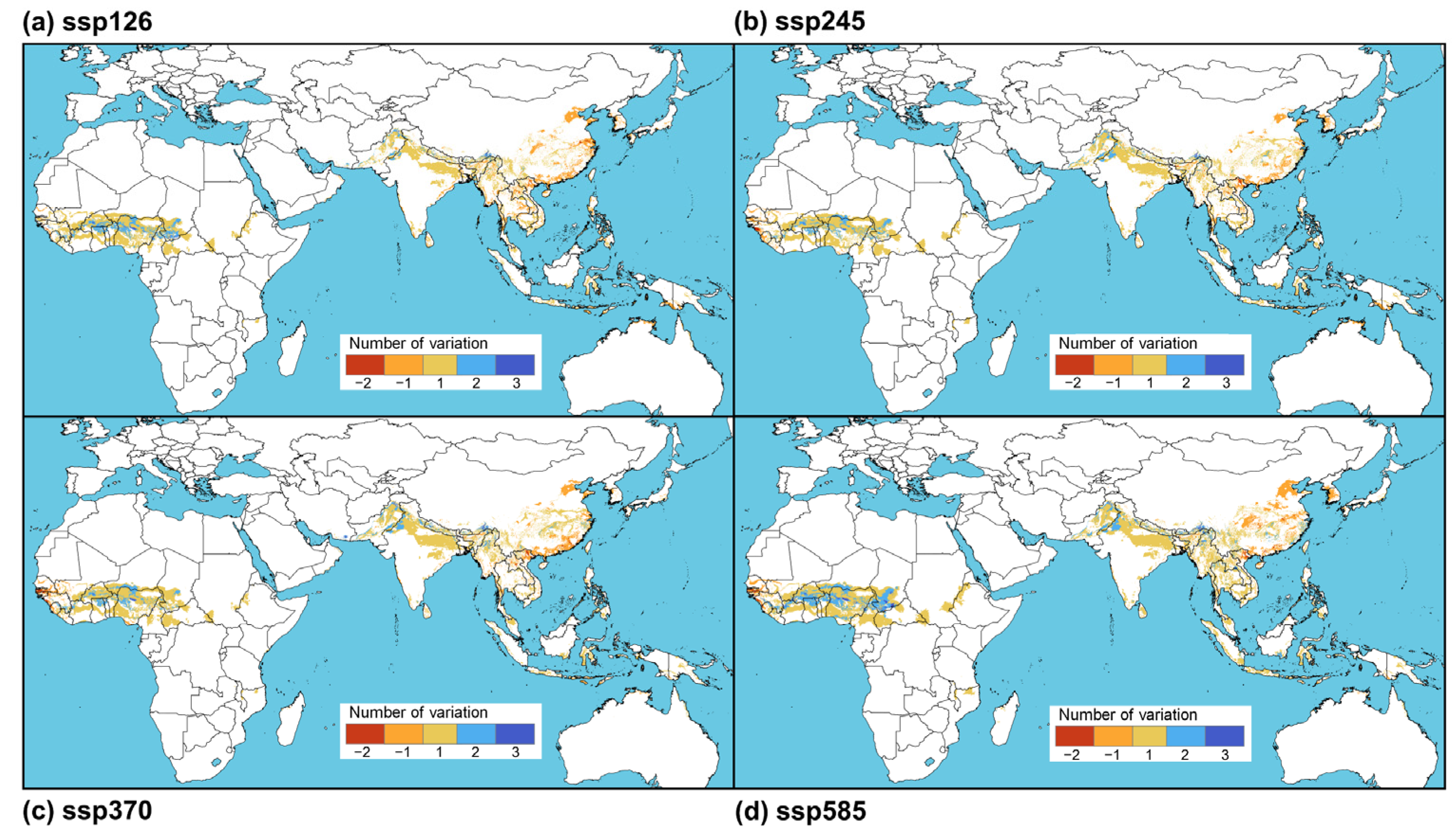
3.3. Changing Suitable Areas under Climate Change
3.4. Further Collection Assessments
3.5. Main Environmental Predictors Determining the Distributions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ramirez-Villegas, J.; Khoury, C.K.; Achicanoy, H.A.; Diaz, M.V.; Mendez, A.C.; Sosa, C.C.; Kehel, Z.; Guarino, L.; Abberton, M.; Aunario, J.; et al. State of ex situ conservation of landrace groups of 25 major crops. Nat. Plants 2022, 8, 491–499. [Google Scholar] [CrossRef]
- Villa, T.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef]
- Khoury, C.K.; Brush, S.; Costich, D.E.; Curry, H.A.; de Haan, S.; Engels, J.M.M.; Guarino, L.; Hoban, S.; Mercer, K.L.; Miller, A.J.; et al. Crop genetic erosion: Understanding and responding to loss of crop diversity. New Phytol. 2022, 233, 84–118. [Google Scholar] [CrossRef]
- Reem, H.; Toby, H. The use of wild relatives in crop improvement: A survey of developments over the last 20 years Euphytica. Neth. J. Plant Breed. 2007, 156, 1–13. [Google Scholar] [CrossRef]
- Tilman, D.; Clark, M. Global diets link environmental sustainability and human health. Nature 2014, 515, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Mercer, K.L.; Perales, H.R. Evolutionary response of landraces to climate change in centers of crop diversity. Evol. Appl. 2010, 3, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Toward an evolved concept of landrace. Front. Plant Sci. 2017, 8, 145. [Google Scholar] [CrossRef]
- Frison, E.A.; Cherfas, J.; Hodgkin, T. Agricultural Biodiversity Is Essential for a Sustainable Improvement in Food and Nutrition Security. Sustainability 2011, 3, 238–253. [Google Scholar] [CrossRef]
- Wambugu, P.W.; Ndjiondjop, M.N.; Henry, R. Genetics and Genomics of African Rice (Oryza glaberrima Steud) Domestication. Rice 2021, 14, 6. [Google Scholar] [CrossRef]
- Chen, C.; He, W.; Nassirou, T.Y.; Nsabiyumva, A.; Dong, X.; Adedze, Y.M.N.; Jin, D. Molecular characterization and genetic diversity of different genotypes of Oryza sativa and Oryza glaberrima. Electron. J. Biotechnol. 2017, 30, 48–57. [Google Scholar] [CrossRef]
- Mir, R.A.; Sharma, A.; Mahajan, R. Crop Landraces: Present Threats and Opportunities for Conservation. In Rediscovery of Genetic and Genomic Resources for Future Food Security; Salgotra, R., Zargar, S., Eds.; Springer: Singapore, 2020. [Google Scholar] [CrossRef]
- B. Groombridge. Global Biodiversity: Status of the Earth’s Living Resources. A Report Compiled by the World Conservation Monitoring Centre. Geol. Mag. 1992, 132, 124. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Cao, B.; Bai, C.; Zhang, M.; Lu, Y.; Gao, P.; Yang, J.; Xue, Y.; Li, G. Future landscape of renewable fuel resources: Current and future conservation and utilization of main biofuel crops in China. Sci. Total Environ. 2022, 806, 150946. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Cao, B.; Bai, C.; Wu, K.; Xue, Y.; Yang, J.; Gao, P.; Liang, H.; Li, G. Concentrated conservation and utilization: Four medicinal crops for diabetes treatment showed similar habitat distribution patterns in China. Ind. Crop. Prod. 2020, 152, 112478. [Google Scholar] [CrossRef]
- Cámara-Leret, R.; Raes, N.; Roehrdanz, P.; De Fretes, Y.; Heatubun, C.D.; Roeble, L.; Schuiteman, A.; Van Welzen, P.C.; Hannah, L. Climate change threatens New Guinea’s biocultural heritage. Sci. Adv. 2019, 5, eaaz1455. [Google Scholar] [CrossRef] [PubMed]
- Pironon, S.; Etherington, T.R.; Borrell, J.S.; Kühn, N.; Macias-Fauria, M.; Ondo, I.; Tovar, C.; Wilkin, P.; Willis, K.J. Potential adaptive strategies for 29 Sub-Saharan crops under future climate change. Nat. Clim. Chang. 2019, 9, 758–763. [Google Scholar] [CrossRef]
- Castañeda-Álvarez, N.P.; Khoury, C.K.; Achicanoy, H.A.; Bernau, V.; Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Harker, R.H.; Jarvis, A.; Maxted, N.; et al. Global conservation priorities for crop wild relatives. Nat. Plants 2016, 2, 16022. [Google Scholar] [CrossRef]
- Khoury, C.K.; Carver, D.; Greene, S.L.; Williams, K.A.; Achicanoy, H.A.; Schori, M.; León, B.; Wiersema, J.H.; Frances, A. Crop wild relatives of the United States require urgent conservation action. Proc. Natl. Acad. Sci. USA 2020, 117, 33351–33357. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Title, P.O.; Bemmels, J.B. ENVIREM: An expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 2018, 41, 291–307. [Google Scholar] [CrossRef]
- Ramirez-Villegas, J.; Khoury, C.K.; Achicanoy, H.A.; Mendez, A.C.; Diaz, M.V.; Sosa, C.C.; Debouck, D.G.; Kehel, Z.; Guarino, L. A gap analysis modelling framework to prioritize collecting for ex situ conservation of crop landraces. Divers Distrib. 2020, 26, 730–742. [Google Scholar] [CrossRef]
- Choi, J.Y.; Purugganan, M. Multiple Origin but Single Domestication Led to Oryza sativa. G3 Genes|Genomes|Genet. 2018, 8, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Zaidem, M.; Gutaker, R.; Dorph, K.; Singh, R.K.; Purugganan, M.D. The complex geography of domestication of the African rice Oryza glaberrima. PLoS Genet. 2019, 15, e1007414. [Google Scholar] [CrossRef]
- Garris, A.; Tai, T.; Coburn, J.; Kresovich, S.; McCouch, S. Genetic Structure and Diversity in Oryza sativa L. Genetics 2005, 169, 1631–1638. [Google Scholar] [CrossRef]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef]
- Ho, T. A Data Complexity Analysis of Comparative Advantages of Decision Forest Constructors. Pattern Anal. Appl. 2002, 5, 102–112. [Google Scholar] [CrossRef]
- Reshef, D.; Reshef, Y.; Finucane, H.; Grossman, S.; McVean, G.; Turnbaugh, P.; Lander, E.; Mitzenmacher, M.; Sabeti, P. Detecting novel associations in large data sets. Science 2011, 334, 1518–1524. [Google Scholar] [CrossRef]
- Lavery, M.R.; Acharya, P.; Sivo, S.A.; Xu, L. Number of predictors and multicollinearity: What are their effects on error and bias in regression? Commun. Stat. -Simul. Comput. 2019, 48, 27–38. [Google Scholar] [CrossRef]
- Stanton, J.C.; Pearson, R.G.; Horning, N.; Ersts, P.; Reşit Akçakaya, H. Combining static and dynamic variables in species distribution models under climate change. Methods Ecol. Evol. 2012, 3, 349–357. [Google Scholar] [CrossRef]
- Vavilov, N.I. The centres of origin of cultivated plants. Works Appl. Bot. Plant Breed. 1926, 16, 1–248. [Google Scholar]
- McCouch, S.R.; Wright, M.H.; Tung, C.-W.; Maron, L.G.; McNally, K.L.; Fitzgerald, M.; Singh, N.; DeClerck, G.; Agosto-Perez, F.; Korniliev, P.; et al. Open access resources for genome-wide association mapping in rice. Nat. Commun. 2016, 7, 10532. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Sun, X.; Caiji, Z.; Yang, J.; Di Cui, D.; Cao, G.; Ma, X.; Han, B.; Xue, D.; et al. Influence of ethnic traditional cultures on genetic diversity of rice landraces under on-farm conservation in southwest China. J. Ethnobiol. Ethnomed. 2016, 12, 51. [Google Scholar] [CrossRef]
- Burke, M.B.; Lobell, D.B.; Guarino, L. Shifts in African crop climates by 2050, and the implications for crop improvement and genetic resources conservation. Glob. Environ. Chang. 2009, 19, 317–325. [Google Scholar] [CrossRef]
- Franzaring, J.; Holz, I.; Kauf, Z.; Fangmeier, A. Responses of the novel bioenergy plant species Sida hermaphrodita (L.) rusby and Silphium perfoliatum L. to CO2 fertilization at different temperatures and water supply. Biomass Bioenerg. 2015, 81, 574–583. [Google Scholar] [CrossRef]
- García, R.M.; Parra-Quijano, M.; Iriondo, J.M. A Multispecies Collecting Strategy for Crop Wild Relatives Based on Complementary Areas with a High Density of Ecogeographical Gaps. Crop Sci. 2017, 57, 1059–1069. [Google Scholar] [CrossRef]
- Yu, X.; Tao, X.; Liao, J.; Liu, S.; Xu, L.; Yuan, S.; Zhang, Z.; Wang, F.; Deng, N.; Huang, J.; et al. Predicting potential cultivation region and paddy area for ratoon rice production in China using Maxent model. Field Crops Res. 2022, 275, 108372. [Google Scholar] [CrossRef]
- Frieler, K.; Schauberger, B.; Arneth, A.; Balkovič, J.; Chryssanthacopoulos, J.; Deryng, D.; Elliott, J.; Folberth, C.; Khabarov, N.; Müller, C.; et al. Understanding the weather signal in national crop-yield variability. Earths Future 2017, 5, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Wang, H.; Ma, X.; Peng, H.; Shan, J. Modeling the current land suitability and future dynamics of global soybean cultivation under climate change scenarios. Field Crop. Res. 2021, 263, 108069. [Google Scholar] [CrossRef]
- Hanson, J.O.; Rhodes, J.R.; Riginos, C.; Fuller, R.A. Environmental and geographic variables are effective surrogates for genetic variation in conservation planning. Proc. Natl Acad. Sci. USA 2017, 114, 12755–12760. [Google Scholar] [CrossRef] [PubMed]
- Khoury, C.K.; Amariles, D.; Soto, J.S.; Diaz, M.V.; Sotelo, S.; Sosa, C.C.; Ramírez-Villegas, J.; Achicanoy, H.A.; Velásquez-Tibatá, J.; Guarino, L.; et al. Comprehensiveness of conservation of useful wild plants: An operational indicator for biodiversity and sustainable development targets. Ecol. Indic. 2019, 98, 420–429. [Google Scholar] [CrossRef]
- Winkler, K.; Fuchs, R.; Rounsevell, M.; Herold, M. Global land use changes are four times greater than previously estimated. Nat. Commun. 2021, 12, 2501. [Google Scholar] [CrossRef] [PubMed]
- Braunisch, V.; Coppes, J.; Arlettaz, R.; Suchant, R.; Schmid, H.; Bollmann, K. Selecting from correlated climate variables: A major source of uncertainty for predicting species distributions under climate change. Ecography 2013, 36, 971–983. [Google Scholar] [CrossRef]
- Grenouillet, G.; Buisson, L.; Casajus, N.; Lek, S. Ensemble modelling of species distribution: The effects of geographical and environmental ranges. Ecography 2011, 34, 9–17. [Google Scholar] [CrossRef]
- Franceschini, S.; Gandola, E.; Martinoli, M.; Tancioni, L.; Scardi, M. Cascaded neural networks improving fish species prediction accuracy: The role of the biotic information. Sci. Rep. 2018, 8, 4581. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, J.-E.; DiTommaso, A.; Wang, R.-L.; Wu, R.-S. Predicting invasions of Wedelia trilobata (L.) Hitchc. with Maxent and GARP models. J. Plant Res. 2015, 128, 763–775. [Google Scholar] [CrossRef]
- Luyssaert, S.; Jammet, M.; Stoy, P.C.; Estel, S.; Pongratz, J.; Ceschia, E.; Churkina, G.; Don, A.; Erb, K.; Ferlicoq, M.; et al. Land management and land-cover change have impacts of similar magnitude on surface temperature. Nat. Clim. Chang. 2014, 4, 389–393. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Crop Landrace | Subgroup | Predicted Distribution Area |
|---|---|---|
| Asian rice (Oryza sativa L.) | aromatic | Central Pakistan, Nepal, Northeastern India, North Burma |
| aus | Central Pakistan, Northern and Northeastern India, Bangladesh, North Burma | |
| indica | Northeastern India, Bangladesh, Laos, Cambodia, Southern Vietnam, Southern China | |
| japonica | Southeastern India, Southern Thailand, Southern Vietnam, Philippines, Southern and Central China, Korea | |
| African rice (Oryza glaberrima Steud.) | K2 | Northern Ghana, Northern Togo, Northern Benin, Central Nigeria |
| K4 | Gambia, Senegal, Guinea Peso, Southern Mali, Central Burkina Faso, Northern Nigeria | |
| K5 | Guinea Peso, Western Guinea, Sierra Leone | |
| others | Southern Senegal, Guinea Peso, Sierra Leone, Northern Liberia, Eastern Nigeria |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.; Wang, H.; Chen, Y.; Tan, J.; Hong, J.; Yan, S.; Cao, Y.; Fang, W. Modelling Distributions of Asian and African Rice Based on MaxEnt. Sustainability 2023, 15, 2765. https://doi.org/10.3390/su15032765
Lin Y, Wang H, Chen Y, Tan J, Hong J, Yan S, Cao Y, Fang W. Modelling Distributions of Asian and African Rice Based on MaxEnt. Sustainability. 2023; 15(3):2765. https://doi.org/10.3390/su15032765
Chicago/Turabian StyleLin, Yunan, Hao Wang, Yanqing Chen, Jiarui Tan, Jingpeng Hong, Shen Yan, Yongsheng Cao, and Wei Fang. 2023. "Modelling Distributions of Asian and African Rice Based on MaxEnt" Sustainability 15, no. 3: 2765. https://doi.org/10.3390/su15032765
APA StyleLin, Y., Wang, H., Chen, Y., Tan, J., Hong, J., Yan, S., Cao, Y., & Fang, W. (2023). Modelling Distributions of Asian and African Rice Based on MaxEnt. Sustainability, 15(3), 2765. https://doi.org/10.3390/su15032765