
Nitrogen and Phosphorus Discriminate the Assembly Processes of Prokaryotic and Eukaryotic Algae in an Agricultural Drainage Receiving Lake


Abstract
1. Introduction
2. Materials and Methods
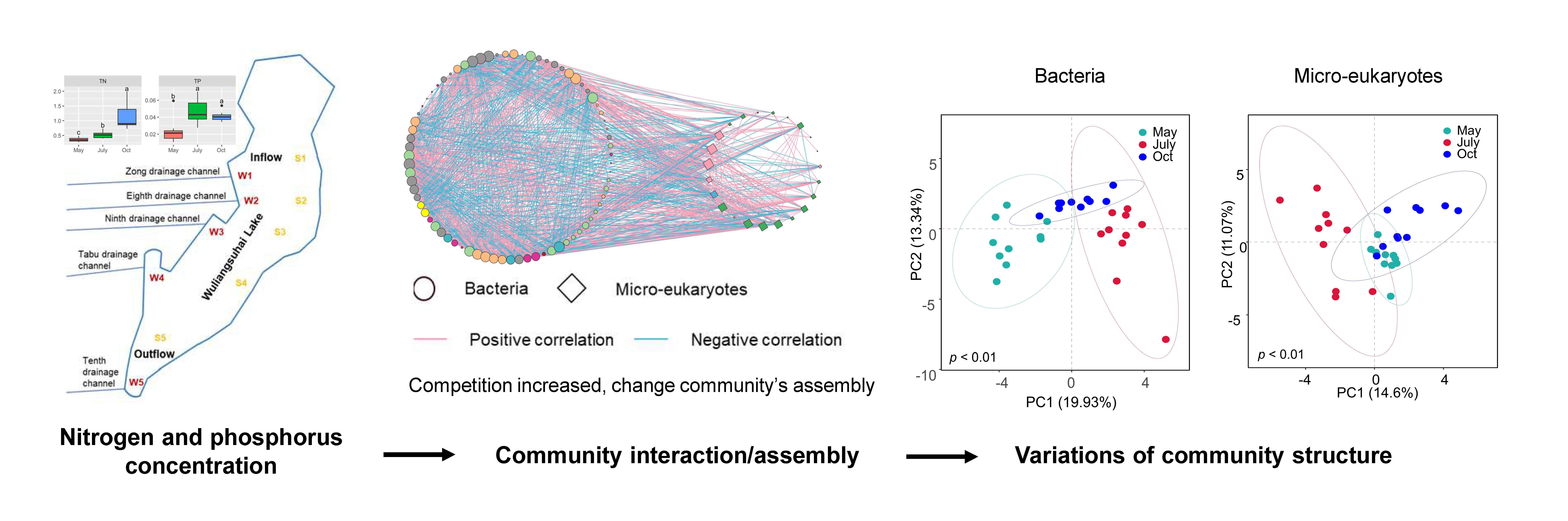
2.1. Study Sites and Sampling
2.2. Chemical Analysis
2.3. Biological Analysis
2.4. Statistical Analysis
3. Results
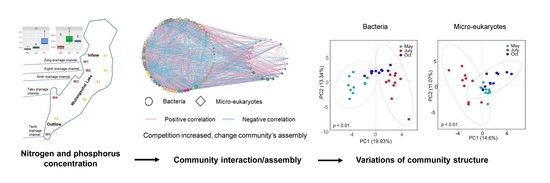
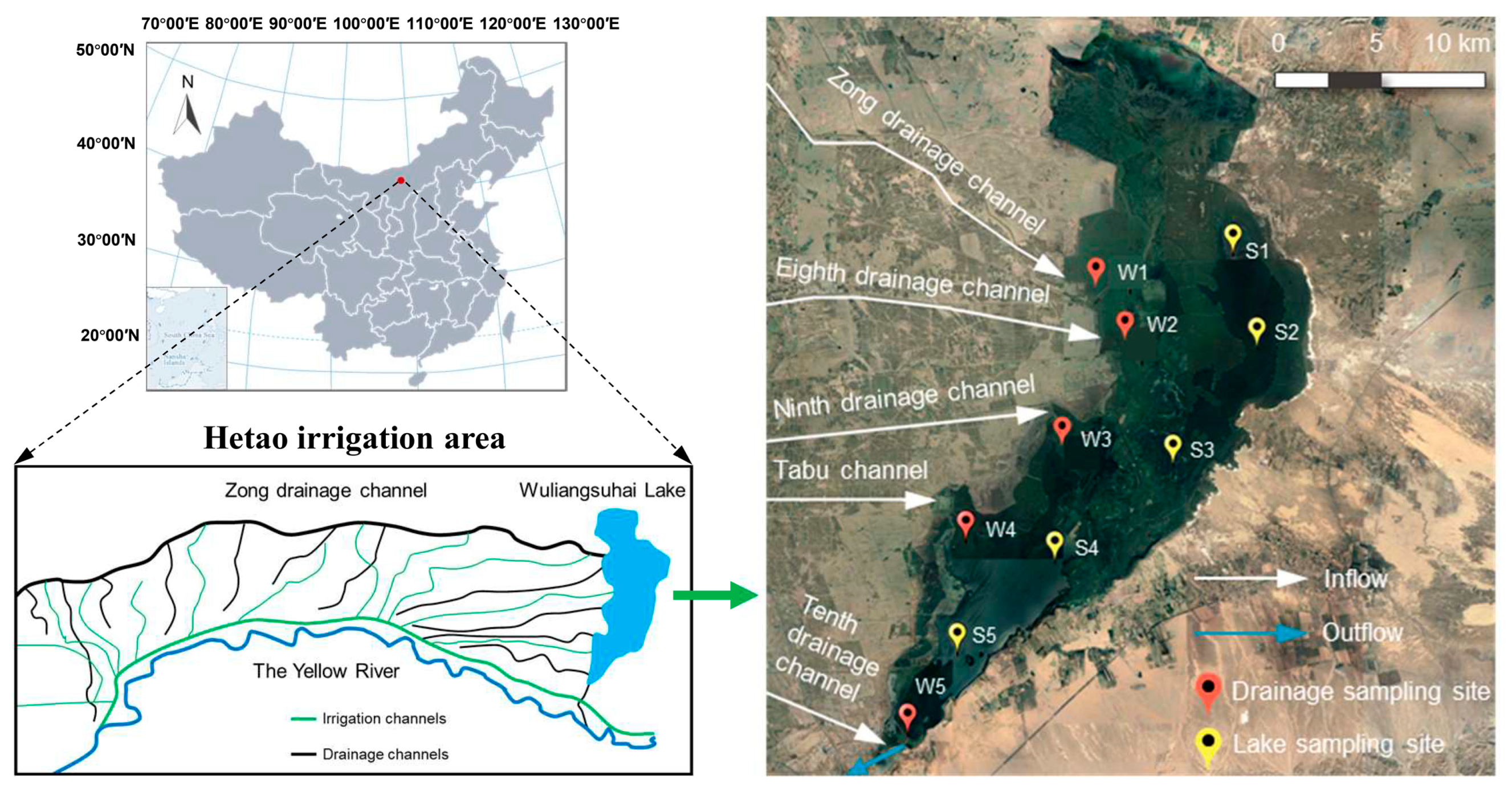
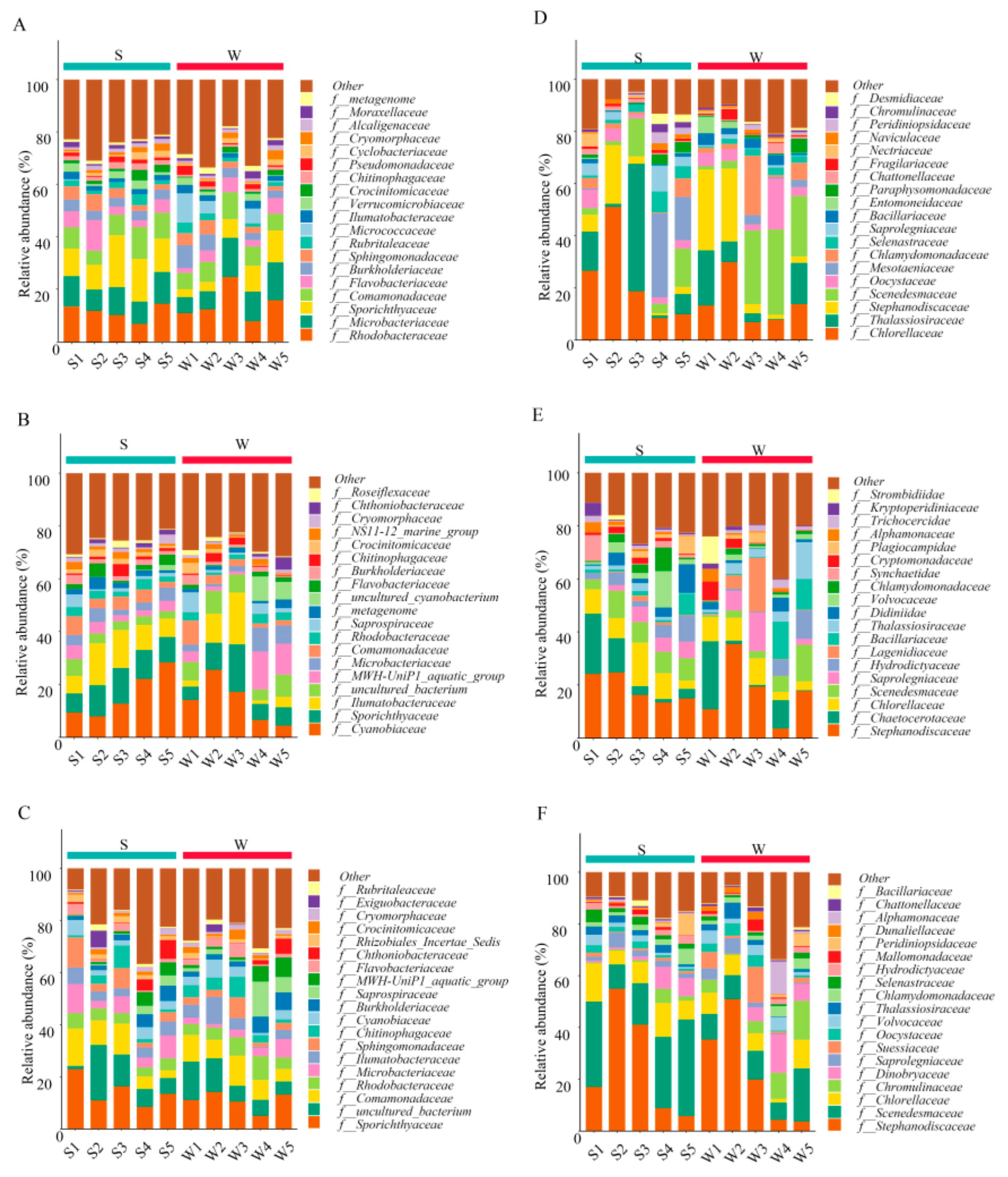
3.1. Variations in Bacterial Community Structure during the Seasons
3.2. Variations in Microeukaryotic Community Structure during the Seasons
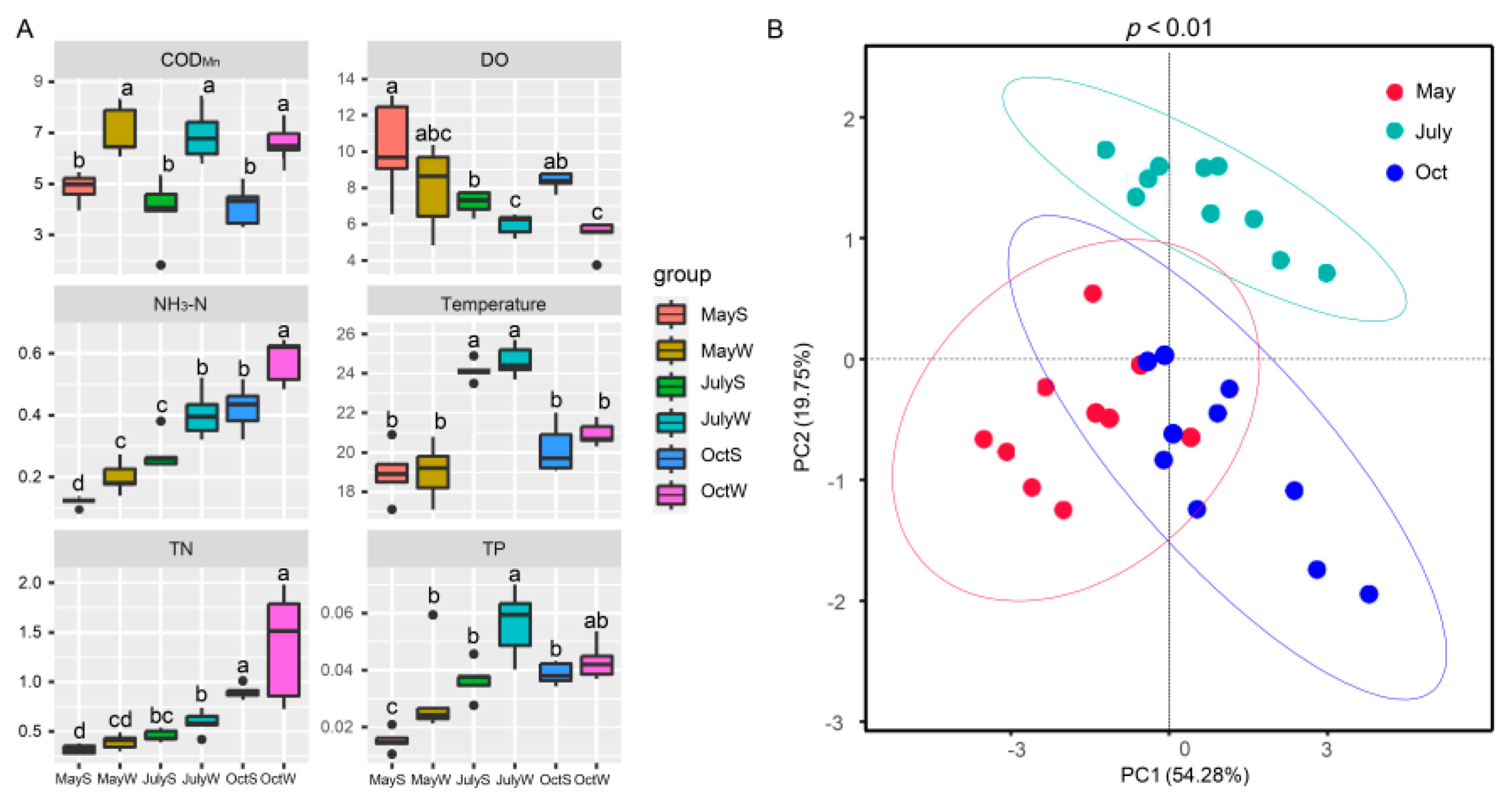
3.3. Seasonal Variations in Physicochemical Factors and Their Correlation with Microorganisms
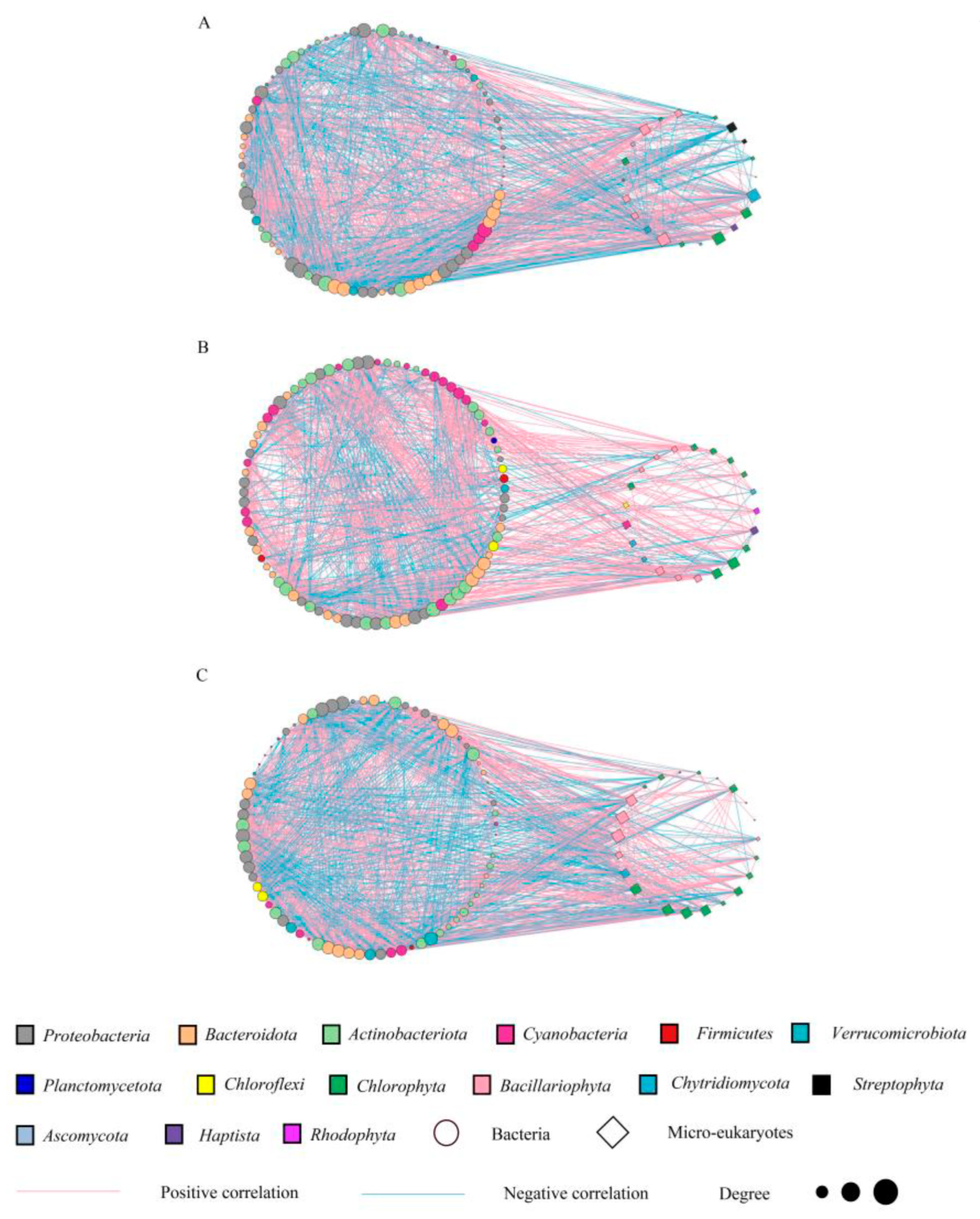
3.4. Interactions of the Bacterial and Microeukaryotic Communities
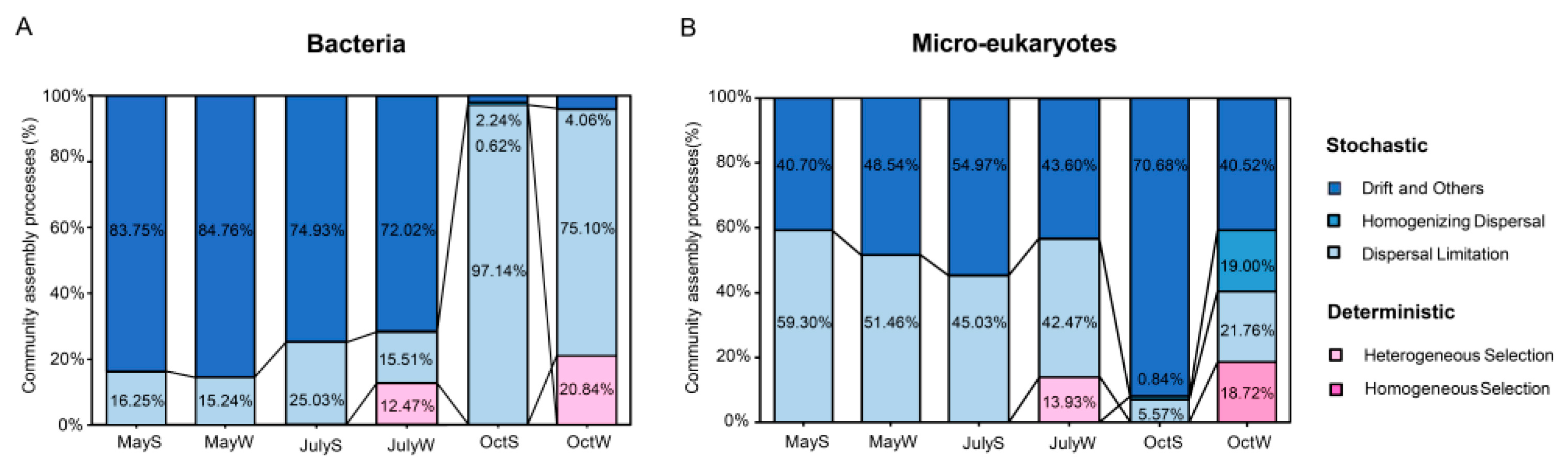
3.5. Community Assembly of Bacteria and Microeukaryotes
4. Discussion
4.1. N and P Inputs Significantly Affected the Seasonal Variations in Community Structure
4.2. Specialized Interaction Patterns between Phytoplankton and Bacteria with N and P Inputs
4.3. N and P Inputs Contributed Greatly to Community Assembly
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, X.; Wei, J.; Bai, N.; Cha, H.; Cao, C.; Zheng, K.; Liu, Y. The phosphorus fractions and adsorption-desorption characteristics in the Wuliangsuhai Lake, China. Environ. Sci. Pollut. Res. Int. 2018, 25, 20648–20661. [Google Scholar] [CrossRef]
- Ji, B.; Qin, H.; Guo, S.; Chen, W.; Zhang, X.; Liang, J. Bacterial communities of four adjacent fresh lakes at different trophic status. Ecotoxicol. Environ. Saf. 2018, 157, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; van der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [PubMed]
- Seymour, J.R.; Amin, S.A.; Raina, J.-B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton–bacteria relationships. Nat. Microbiol. 2017, 2, 17065. [Google Scholar] [CrossRef] [PubMed]
- Tanentzap, A.J.; Fitch, A.; Orland, C.; Emilson, E.J.S.; Yakimovich, K.M.; Osterholz, H.; Dittmar, T. Chemical and microbial diversity covary in fresh water to influence ecosystem functioning. Proc. Natl. Acad. Sci. USA 2019, 116, 24689–24695. [Google Scholar] [CrossRef]
- Foo, J.L.; Ling, H.; Lee, Y.S.; Chang, M.W. Microbiome engineering: Current applications and its future. Biotechnol. J. 2017, 12, 1600099. [Google Scholar] [CrossRef] [PubMed]
- Fallahi, A.; Rezvani, F.; Asgharnejad, H.; Khorshidi Nazloo, E.; Hajinajaf, N.; Higgins, B. Interactions of microalgae-bacteria consortia for nutrient removal from wastewater: A review. Chemosphere 2021, 272, 129878. [Google Scholar] [CrossRef]
- Xu, R.; Zhang, K.; Liu, P.; Khan, A.; Xiong, J.; Tian, F.; Li, X. A critical review on the interaction of substrate nutrient balance and microbial community structure and function in anaerobic co-digestion. Bioresour. Technol. 2018, 247, 1119–1127. [Google Scholar] [CrossRef]
- Zheng, L.; Wang, X.; Ren, M.; Yuan, D.; Tan, Q.; Xing, Y.; Xia, X.; Xie, E.; Ding, A. Comparing with oxygen, nitrate simplifies microbial community assembly and improves function as an electron acceptor in wastewater treatment. Environ. Pollut. 2022, 314, 120243. [Google Scholar] [CrossRef]
- Niu, K.; Yi-ning, L.; Shen, Z.; Fang-liang, H.; Jing-yun, F. Community assembly: The relative importance of neutral theory and niche theory. Biodivers. Sci. 2009, 17, 579. [Google Scholar] [CrossRef]
- Chave, J. Neutral theory and community ecology. Ecol. Lett. 2004, 7, 241–253. [Google Scholar] [CrossRef]
- Zheng, L.; Wang, X.; Ding, A.; Yuan, D.; Tan, Q.; Xing, Y.; Xie, E. Ecological Insights Into Community Interactions, Assembly Processes and Function in the Denitrifying Phosphorus Removal Activated Sludge Driven by Phosphorus Sources. Front. Microbiol. 2021, 12, 779369. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ning, D. Stochastic Community Assembly: Does It Matter in Microbial Ecology? Microbiol. Mol. Biol. Rev. 2017, 81, e00002-17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ning, D.; Yang, Y.; Van Nostrand, J.D.; Zhou, J.; Wen, X. Biodegradability of wastewater determines microbial assembly mechanisms in full-scale wastewater treatment plants. Water Res. 2020, 169, 115276. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, B.; Ning, D.; Zhang, Y.; Dai, T.; Wu, L.; Li, T.; Liu, W.; Zhou, J.; Wen, X. Seasonal dynamics of the microbial community in two full-scale wastewater treatment plants: Diversity, composition, phylogenetic group based assembly and co-occurrence pattern. Water Res. 2021, 200, 117295. [Google Scholar] [CrossRef]
- Ning, D.; Yuan, M.; Wu, L.; Zhang, Y.; Guo, X.; Zhou, X.; Yang, Y.; Arkin, A.P.; Firestone, M.K.; Zhou, J. A quantitative framework reveals ecological drivers of grassland microbial community assembly in response to warming. Nat. Commun. 2020, 11, 4717. [Google Scholar] [CrossRef]
- Zhang, T.; Xu, S.; Yan, R.; Wang, R.; Gao, Y.; Kong, M.; Yi, Q.; Zhang, Y. Similar geographic patterns but distinct assembly processes of abundant and rare bacterioplankton communities in river networks of the Taihu Basin. Water Res. 2022, 211, 118057. [Google Scholar] [CrossRef]
- Xu, R.; Yu, Z.; Zhang, S.; Meng, F. Bacterial assembly in the bio-cake of membrane bioreactors: Stochastic vs. deterministic processes. Water Res. 2019, 157, 535–545. [Google Scholar] [CrossRef]
- Liu, Y.; Li, C.; Anderson, B.; Zhang, S.; Shi, X.; Zhao, S. A modified QWASI model for fate and transport modeling of mercury between the water-ice-sediment in Lake Ulansuhai. Chemosphere 2017, 176, 117–124. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, L.; Zhao, X.; Lian, J.; Zhang, Z. Accumulation and potential sources of heavy metals in soils of the Hetao area, Inner Mongolia, China. Pedosphere 2020, 30, 244–252. [Google Scholar] [CrossRef]
- Ma, L.; Wu, J.; Abuduwaili, J. Geochemical evidence of the anthropogenic alteration of element composition in lacustrine sediments from Wuliangsu Lake, north China. Quat. Int. 2013, 306, 107–113. [Google Scholar] [CrossRef]
- Yue, W.; Meng, K.; Hou, K.; Zuo, R.; Zhang, B.-T.; Wang, G. Evaluating climate and irrigation effects on spatiotemporal variabilities of regional groundwater in an arid area using EOFs. Sci. Total Environ. 2020, 709, 136147. [Google Scholar] [CrossRef] [PubMed]
- Köbbing, J.F.; Patuzzi, F.; Baratieri, M.; Beckmann, V.; Thevs, N.; Zerbe, S. Economic evaluation of common reed potential for energy production: A case study in Wuliangsuhai Lake (Inner Mongolia, China). Biomass Bioenergy 2014, 70, 315–329. [Google Scholar] [CrossRef]
- Rojas-Jimenez, K.; Grossart, H.P.; Cordes, E.; Cortes, J. Fungal Communities in Sediments Along a Depth Gradient in the Eastern Tropical Pacific. Front. Microbiol. 2020, 11, 575207. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Barabasi, A.L.; Oltvai, Z.N. Network biology: Understanding the cell’s functional organization. Nat. Rev. Genet. 2004, 5, 101–113. [Google Scholar] [CrossRef]
- Newman, M.E.J. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577. [Google Scholar] [CrossRef]
- Ma, X.; Li, X.; Liu, J.; Cheng, Y.; Zou, J.; Zhai, F.; Sun, Z.; Han, L. Soil microbial community succession and interactions during combined plant/white-rot fungus remediation of polycyclic aromatic hydrocarbons. Sci. Total Environ. 2021, 752, 142224. [Google Scholar] [CrossRef]
- Ma, J.; Qin, B.; Paerl, H.W.; Brookes, J.D.; Wu, P.; Zhou, J.; Deng, J.; Guo, J.; Li, Z. Green algal over cyanobacterial dominance promoted with nitrogen and phosphorus additions in a mesocosm study at Lake Taihu, China. Environ. Sci. Pollut. Res. Int. 2015, 22, 5041–5049. [Google Scholar] [CrossRef]
- Xue, Y.; Chen, L.; Zhao, Y.; Feng, Q.; Li, C.; Wei, Y. Shift of soil fungal communities under afforestation in Nanliu River Basin, southwest China. J. Environ. Manag. 2022, 302, 114130. [Google Scholar] [CrossRef]
- Quero, G.M.; Perini, L.; Pesole, G.; Manzari, C.; Lionetti, C.; Bastianini, M.; Marini, M.; Luna, G.M. Seasonal rather than spatial variability drives planktonic and benthic bacterial diversity in a microtidal lagoon and the adjacent open sea. Mol. Ecol. 2017, 26, 5961–5973. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Hu, Y.; Zhang, S.; Wu, R.; Guo, X. Microplastics in the surface water of Wuliangsuhai Lake, northern China. Sci. Total Environ. 2020, 723, 137820. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, Y.; Ma, T.; Yang, H.; He, J. Arsenic associations in sediments from shallow aquifers of northwestern Hetao Basin, Inner Mongolia. Environ. Earth Sci. 2011, 64, 2001–2011. [Google Scholar] [CrossRef]
- Yuan, M.M.; Guo, X.; Wu, L.; Zhang, Y.; Xiao, N.; Ning, D.; Shi, Z.; Zhou, X.; Wu, L.; Yang, Y.; et al. Climate warming enhances microbial network complexity and stability. Nat. Clim. Chang. 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Zhang, L.; Bai, J.; Zhang, K.; Wang, Y.; Xiao, R.; Campos, M.; Acuña, J.; Jorquera, M.A. Occurrence, bioaccumulation and ecological risks of antibiotics in the water-plant-sediment systems in different functional areas of the largest shallow lake in North China: Impacts of river input and historical agricultural activities. Sci. Total Environ. 2023, 857, 159260. [Google Scholar] [CrossRef] [PubMed]
- Räike, A.; Pietiläinen, O.P.; Rekolainen, S.; Kauppila, P.; Pitkänen, H.; Niemi, J.; Raateland, A.; Vuorenmaa, J. Trends of phosphorus, nitrogen and chlorophyll a concentrations in Finnish rivers and lakes in 1975–2000. Sci. Total Environ. 2003, 310, 47–59. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Schmidt, S.K.; Fukami, T.; O’Neill, S.P.; Bilinski, T.M.; Stanish, L.F.; Knelman, J.E.; Darcy, J.L.; Lynch, R.C.; Wickey, P.; et al. Patterns and processes of microbial community assembly. Microbiol. Mol. Biol. Rev. 2013, 77, 342–356. [Google Scholar] [CrossRef]
- Shang, Y.; Wu, X.; Wang, X.; Wei, Q.; Ma, S.; Sun, G.; Zhang, H.; Wang, L.; Dou, H.; Zhang, H. Factors affecting seasonal variation of microbial community structure in Hulun Lake, China. Sci. Total Environ. 2022, 805, 150294. [Google Scholar] [CrossRef]
- Markou, G.; Vandamme, D.; Muylaert, K. Microalgal and cyanobacterial cultivation: The supply of nutrients. Water Res. 2014, 65, 186–202. [Google Scholar] [CrossRef] [PubMed]
- Gardner, W.S.; Newell, S.E.; McCarthy, M.J.; Hoffman, D.K.; Lu, K.; Lavrentyev, P.J.; Hellweger, F.L.; Wilhelm, S.W.; Liu, Z.; Bruesewitz, D.A.; et al. Community Biological Ammonium Demand: A Conceptual Model for Cyanobacteria Blooms in Eutrophic Lakes. Environ. Sci. Technol. 2017, 51, 7785–7793. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, M.H.; Hamilton, D.P.; Etemad-Shahidi, A.; Helfer, F. Individual-based modelling of cyanobacteria blooms: Physical and physiological processes. Sci. Total Environ. 2021, 792, 148418. [Google Scholar] [CrossRef] [PubMed]
- Xie, E.; Zhao, X.; Li, K.; Zhang, P.; Zhou, X.; Zhao, X. Microbial community structure in the river sediments from upstream of Guanting Reservoir: Potential impacts of reclaimed water recharge. Sci. Total Environ. 2021, 766, 142609. [Google Scholar] [CrossRef]
- Yu, M.; Liu, S.; Li, G.; Zhang, H.; Xi, B.; Tian, Z.; Zhang, Y.; He, X. Municipal wastewater effluent influences dissolved organic matter quality and microbial community composition in an urbanized stream. Sci. Total Environ. 2020, 705, 135952. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Zheng, L.; Tan, Q.; Wang, X.; Xing, Y.; Wang, H.; Wang, S.; Zhu, G. Comammox activity dominates nitrification process in the sediments of plateau wetland. Water Res. 2021, 206, 117774. [Google Scholar] [CrossRef]
- Tian, L.; Yan, Z.; Wang, C.; Xu, S.; Jiang, H. Habitat heterogeneity induces regional differences in sediment nitrogen fixation in eutrophic freshwater lake. Sci. Total Environ. 2021, 772, 145594. [Google Scholar] [CrossRef]
- Shen, C.; He, J.Z.; Ge, Y. Seasonal dynamics of soil microbial diversity and functions along elevations across the treeline. Sci. Total Environ. 2021, 794, 148644. [Google Scholar] [CrossRef]
- Zhu, C.; Zhang, J.; Nawaz, M.Z.; Mahboob, S.; Al-Ghanim, K.A.; Khan, I.A.; Lu, Z.; Chen, T. Seasonal succession and spatial distribution of bacterial community structure in a eutrophic freshwater Lake, Lake Taihu. Sci. Total Environ. 2019, 669, 29–40. [Google Scholar] [CrossRef]
- Hongxia, M.; Jingfeng, F.; Jiwen, L.; Su, j.; Zhiyi, W.; Yantao, W.; Dongwei, L.; Mengfei, L.; Tingting, S.; Yuan, J.; et al. Full-length 16S rRNA gene sequencing reveals spatiotemporal dynamics of bacterial community in a heavily polluted estuary, China. Environ. Pollut. 2021, 275, 116567. [Google Scholar] [CrossRef]
- Pringault, O.; Bouvy, M.; Carre, C.; Mejri, K.; Bancon-Montigny, C.; Gonzalez, C.; Leboulanger, C.; Hlaili, A.S.; Goni-Urriza, M. Chemical contamination alters the interactions between bacteria and phytoplankton. Chemosphere 2021, 278, 130457. [Google Scholar] [CrossRef]
- Jia, R.; Qu, Z.; You, P.; Qu, D. Effect of biochar on photosynthetic microorganism growth and iron cycling in paddy soil under different phosphate levels. Sci. Total Environ. 2018, 612, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Azam, F.; Fenchel, T.M.; Field, J.G.; Gray, J.S.; Meyer-Reil, L.-A.; Thingstad, F.J.M.E.P.S. The Ecological Role of Water-Column Microbes in the Sea*. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Santi, I.; Tsiola, A.; Dimitriou, P.D.; Fodelianakis, S.; Kasapidis, P.; Papageorgiou, N.; Daffonchio, D.; Pitta, P.; Karakassis, I. Prokaryotic and eukaryotic microbial community responses to N and P nutrient addition in oligotrophic Mediterranean coastal waters: Novel insights from DNA metabarcoding and network analysis. Mar. Environ. Res. 2019, 150, 104752. [Google Scholar] [CrossRef] [PubMed]
- Meyer, N.; Bigalke, A.; Kaulfuß, A.; Pohnert, G. Strategies and ecological roles of algicidal bacteria. FEMS Microbiol. Rev. 2017, 41, 880–899. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ren, K.; Isabwe, A.; Chen, H.; Liu, M.; Yang, J. Stochastic processes shape microeukaryotic community assembly in a subtropical river across wet and dry seasons. Microbiome 2019, 7, 138. [Google Scholar] [CrossRef]
- Mo, Y.; Zhang, W.; Yang, J.; Lin, Y.; Yu, Z.; Lin, S. Biogeographic patterns of abundant and rare bacterioplankton in three subtropical bays resulting from selective and neutral processes. ISME J. 2018, 12, 2198–2210. [Google Scholar] [CrossRef]
- Lin, Q.; De Vrieze, J.; Li, C.; Li, J.; Li, J.; Yao, M.; Hedenec, P.; Li, H.; Li, T.; Rui, J.; et al. Temperature regulates deterministic processes and the succession of microbial interactions in anaerobic digestion process. Water Res. 2017, 123, 134–143. [Google Scholar] [CrossRef]
- Xun, W.; Li, W.; Xiong, W.; Ren, Y.; Liu, Y.; Miao, Y.; Xu, Z.; Zhang, N.; Shen, Q.; Zhang, R. Diversity-triggered deterministic bacterial assembly constrains community functions. Nat. Commun. 2019, 10, 3833. [Google Scholar] [CrossRef]
- Wang, K.; Yan, H.; Peng, X.; Hu, H.; Zhang, H.; Hou, D.; Chen, W.; Qian, P.; Liu, J.; Cai, J.; et al. Community assembly of bacteria and archaea in coastal waters governed by contrasting mechanisms: A seasonal perspective. Mol. Ecol. 2020, 29, 3762–3776. [Google Scholar] [CrossRef]
- Li, H.; Song, C.; Yang, L.; Qin, H.; Cao, X.; Zhou, Y. Nutrients regeneration pathway, release potential, transformation pattern and algal utilization strategies jointly drove cyanobacterial growth and their succession. J. Environ. Sci. 2021, 103, 255–267. [Google Scholar] [CrossRef]
- Logares, R.; Tesson, S.V.M.; Canback, B.; Pontarp, M.; Hedlund, K.; Rengefors, K. Contrasting prevalence of selection and drift in the community structuring of bacteria and microbial eukaryotes. Environ. Microbiol. 2018, 20, 2231–2240. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Lu, H.P.; Sastri, A.; Yeh, Y.C.; Gong, G.C.; Chou, W.C.; Hsieh, C.H. Contrasting the relative importance of species sorting and dispersal limitation in shaping marine bacterial versus protist communities. ISME J. 2018, 12, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Sexton, J.P.; Montiel, J.; Shay, J.E.; Stephens, M.R.; Slatyer, R.A. Evolution of Ecological Niche Breadth. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 183–206. [Google Scholar] [CrossRef]
- Liu, L.; Wang, S.; Chen, J. Anthropogenic activities change the relationship between microbial community taxonomic composition and functional attributes. Environ. Microbiol. 2021, 23, 6663–6675. [Google Scholar] [CrossRef]
- Legendre, P.; Mi, X.; Ren, H.; Ma, K.; Yu, M.; Sun, I.-F.; He, F. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef]
- Pandit, S.N.; Kolasa, J.; Cottenie, K. Contrasts between habitat generalists and specialists: An empirical extension to the basic metacommunity framework. Ecology 2009, 90, 2253–2262. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Stegen, J.C.; van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef]
- Ofiteru, I.D.; Lunn, M.; Curtis, T.P.; Wells, G.F.; Criddle, C.S.; Francis, C.A.; Sloan, W.T. Combined niche and neutral effects in a microbial wastewater treatment community. Proc. Natl. Acad. Sci. USA 2010, 107, 15345–15350. [Google Scholar] [CrossRef]
- Feng, K.; Wang, S.; Wei, Z.; Wang, Z.; Zhang, Z.; Wu, Y.; Zhang, Y.; Deng, Y. Niche width of above- and below-ground organisms varied in predicting biodiversity profiling along a latitudinal gradient. Mol. Ecol. 2020, 29, 1890–1902. [Google Scholar] [CrossRef]
- Sánchez-Clemente, R.; Guijo, M.I.; Nogales, J.; Blasco, R. Carbon Source Influence on Extracellular pH Changes along Bacterial Cell-Growth. Genes 2020, 11, 1292. [Google Scholar] [CrossRef]
- Barnett Samuel, E.; Youngblut Nicholas, D.; Koechli Chantal, N.; Buckley Daniel, H. Multisubstrate DNA stable isotope probing reveals guild structure of bacteria that mediate soil carbon cycling. Proc. Natl. Acad. Sci. USA 2021, 118, e2115292118. [Google Scholar] [CrossRef] [PubMed]
- Nehls, U.; Plassard, C. Nitrogen and phosphate metabolism in ectomycorrhizas. New Phytol. 2018, 220, 1047–1058. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Sampling Time | S1 | S2 | S3 | S4 | S5 | W1 | W2 | W3 | W4 | W5 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| May | 0.12 | 0.13 | 0.12 | 0.10 | 0.14 | 0.27 | 0.23 | 0.18 | 0.14 | 0.18 | |
| NH3-N (mg/L) | July | 0.26 | 0.38 | 0.24 | 0.24 | 0.26 | 0.52 | 0.43 | 0.39 | 0.35 | 0.32 |
| Oct | 0.46 | 0.52 | 0.44 | 0.38 | 0.32 | 0.64 | 0.63 | 0.62 | 0.52 | 0.48 | |
| May | 0.28 | 0.27 | 0.37 | 0.32 | 0.36 | 0.49 | 0.41 | 0.43 | 0.31 | 0.34 | |
| TN (mg/L) | July | 0.42 | 0.53 | 0.50 | 0.40 | 0.43 | 0.74 | 0.65 | 0.58 | 0.42 | 0.56 |
| Oct | 0.83 | 1.01 | 0.91 | 0.87 | 0.88 | 1.98 | 1.79 | 1.51 | 0.73 | 0.86 | |
| May | 0.02 | 0.02 | 0.02 | 0.01 | 0.01 | 0.06 | 0.03 | 0.02 | 0.02 | 0.02 | |
| TP (mg/L) | July | 0.03 | 0.05 | 0.04 | 0.04 | 0.03 | 0.07 | 0.06 | 0.06 | 0.05 | 0.04 |
| Oct | 0.04 | 0.04 | 0.04 | 0.03 | 0.04 | 0.05 | 0.05 | 0.04 | 0.04 | 0.04 | |
| May | 5.23 | 5.46 | 4.60 | 3.97 | 4.98 | 8.35 | 7.89 | 6.45 | 6.07 | 6.45 | |
| CODMn (mg/L) | July | 1.82 | 5.36 | 4.60 | 4.06 | 3.94 | 8.46 | 7.43 | 6.77 | 6.18 | 5.80 |
| Oct | 3.47 | 5.22 | 4.50 | 4.33 | 3.31 | 7.69 | 6.98 | 6.49 | 6.33 | 5.52 | |
| May | 6.56 | 13.09 | 9.08 | 12.47 | 9.68 | 8.65 | 10.38 | 4.85 | 6.43 | 9.70 | |
| DO (mg/L) | July | 7.80 | 6.30 | 7.32 | 7.73 | 6.81 | 5.19 | 5.58 | 6.27 | 6.38 | 6.50 |
| Oct | 7.63 | 8.79 | 8.77 | 8.37 | 8.25 | 3.75 | 5.58 | 5.97 | 5.57 | 5.98 | |
| May | 20.90 | 18.90 | 19.40 | 18.50 | 17.10 | 19.20 | 20.80 | 19.80 | 18.20 | 17.10 | |
| T (℃) | July | 24.20 | 24.10 | 24.90 | 23.50 | 24.00 | 24.20 | 23.70 | 24.40 | 25.20 | 25.70 |
| Oct | 19.10 | 19.20 | 19.70 | 22.00 | 20.90 | 20.30 | 20.70 | 21.80 | 21.30 | 20.60 |
| Item | Nodes | Edges | Average | Network | Network | Modularity | Average Clustering | Average | Positive |
|---|---|---|---|---|---|---|---|---|---|
| Degree | Diameter | Density | (No. Modules) | Coefficient (avgCC) | Path Length | Correlation | |||
| May | 108 | 1499 | 27.76 | 5 | 0.26 | 1.75 (4) | 0.65 | 2.06 | 60.84% |
| July | 104 | 1162 | 22.35 | 5 | 0.22 | 1.39 (7) | 0.65 | 2.22 | 68.33% |
| Oct | 99 | 1397 | 28.22 | 5 | 0.29 | 2.93 (5) | 0.69 | 2.04 | 58.48% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, D.; Zheng, H.; Cheng, J.; Wu, G.; Zheng, L.; Xie, E. Nitrogen and Phosphorus Discriminate the Assembly Processes of Prokaryotic and Eukaryotic Algae in an Agricultural Drainage Receiving Lake. Sustainability 2023, 15, 2584. https://doi.org/10.3390/su15032584
Huang D, Zheng H, Cheng J, Wu G, Zheng L, Xie E. Nitrogen and Phosphorus Discriminate the Assembly Processes of Prokaryotic and Eukaryotic Algae in an Agricultural Drainage Receiving Lake. Sustainability. 2023; 15(3):2584. https://doi.org/10.3390/su15032584
Chicago/Turabian StyleHuang, Dongnan, Han Zheng, Jing Cheng, Guanxiong Wu, Lei Zheng, and En Xie. 2023. "Nitrogen and Phosphorus Discriminate the Assembly Processes of Prokaryotic and Eukaryotic Algae in an Agricultural Drainage Receiving Lake" Sustainability 15, no. 3: 2584. https://doi.org/10.3390/su15032584
APA StyleHuang, D., Zheng, H., Cheng, J., Wu, G., Zheng, L., & Xie, E. (2023). Nitrogen and Phosphorus Discriminate the Assembly Processes of Prokaryotic and Eukaryotic Algae in an Agricultural Drainage Receiving Lake. Sustainability, 15(3), 2584. https://doi.org/10.3390/su15032584