


Impacts of Grazing on Vegetation and Soil Physicochemical Properties in Northern Yinshan Mountain Grasslands


Abstract
:1. Introduction
2. Material and Methods
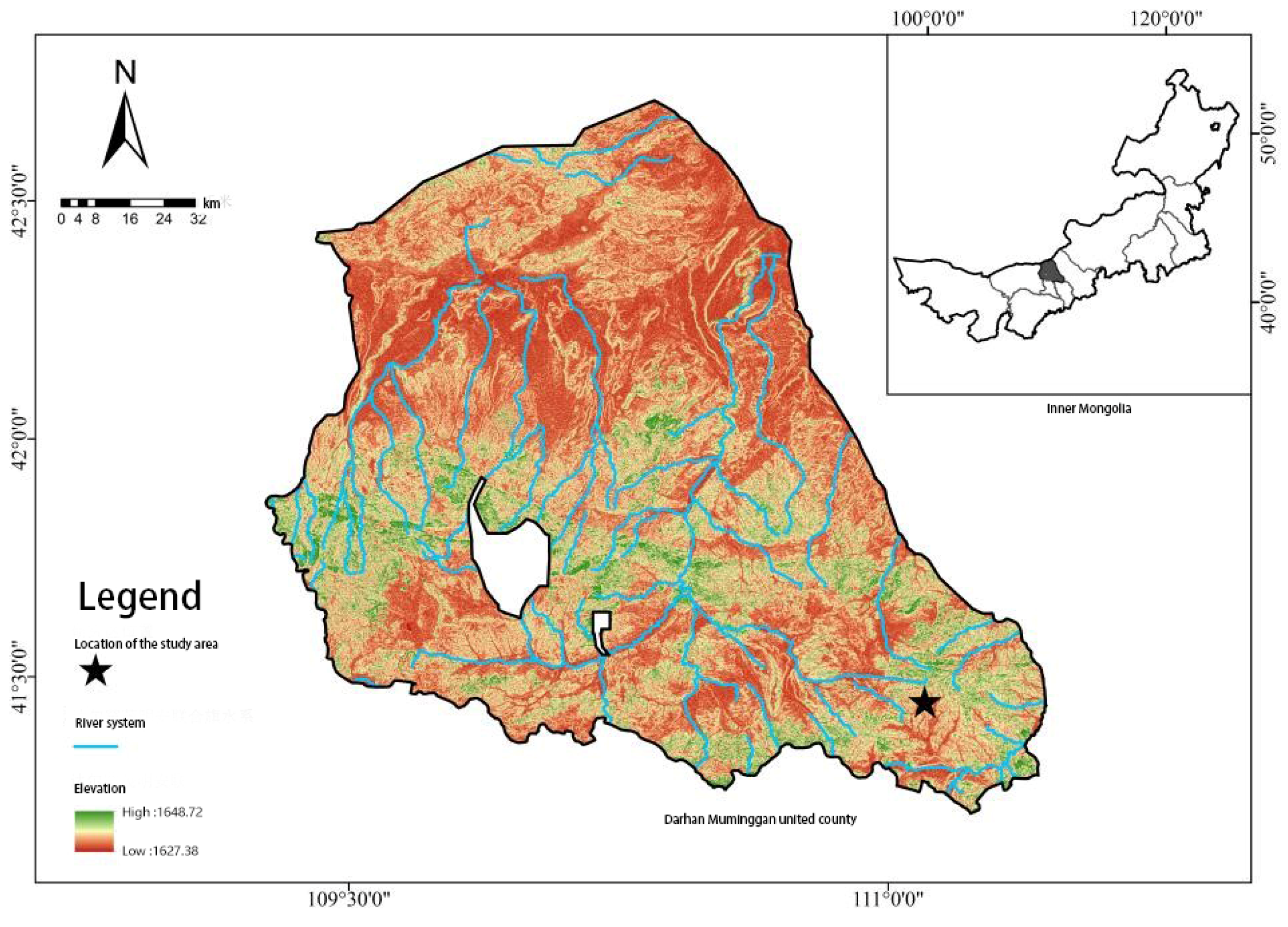
2.1. Research Area Overview
2.2. Experiment Design
2.2.1. Sample Plot Setup
2.2.2. Vegetation Survey and Methods
2.2.3. Soil Sampling and Determination Methods
2.3. Test Methods for Soil Chemical Properties
2.4. Data Processing and Statistical Analysis
3. Results
3.1. Effect of Grazing on the Composition of Vegetation Community and Importance Values
3.2. Effect of Grazing on the Vegetation Community Characteristics
3.3. Influence of Grazing on the Surface Soil Composition
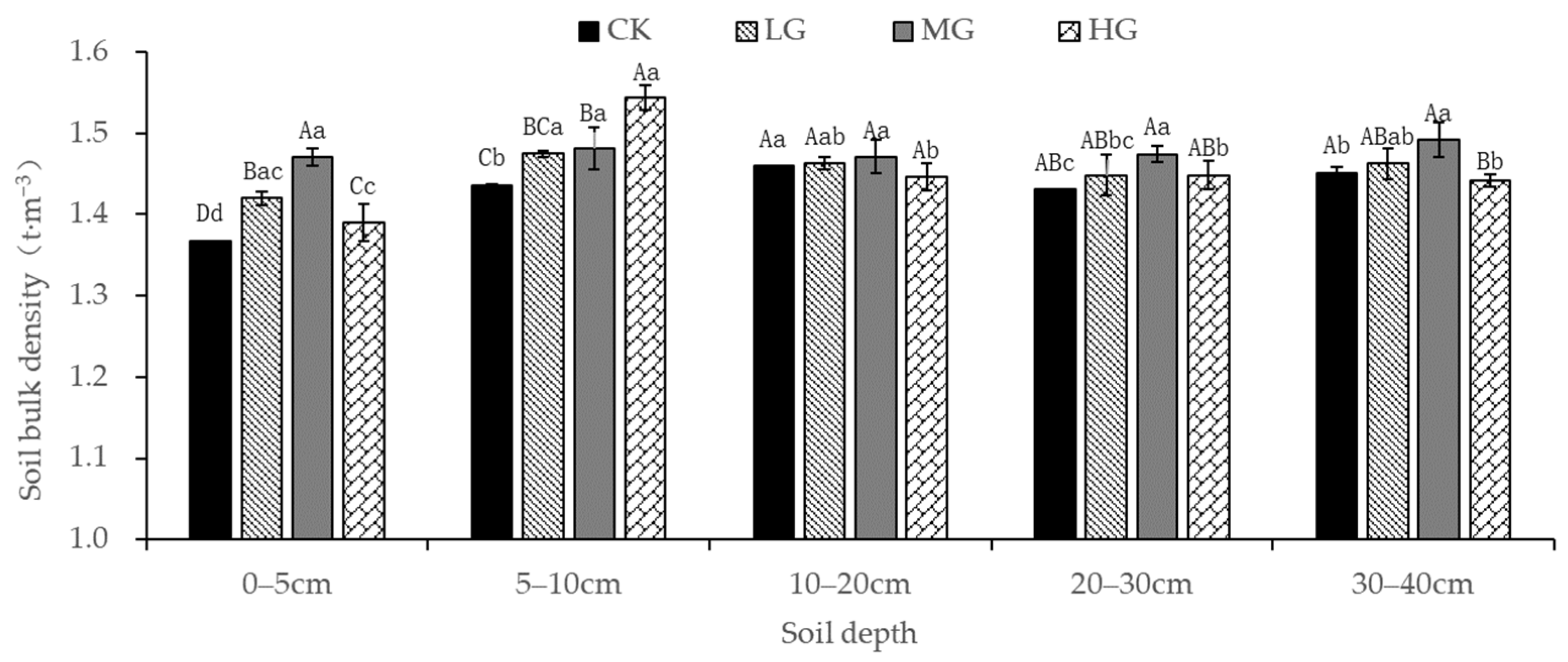
3.4. Influence of Grazing on the Soil Bulk Density
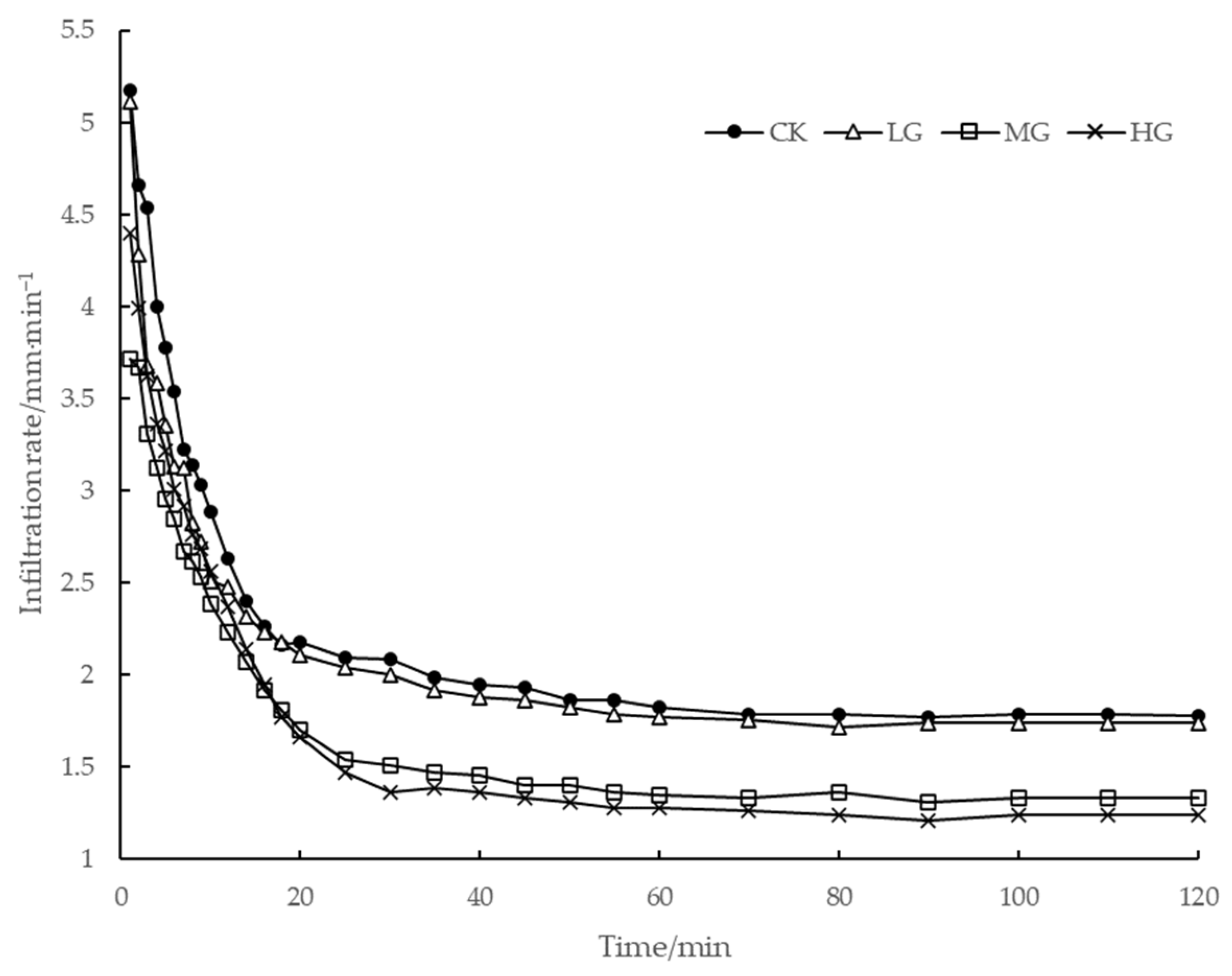
3.5. Influence of Grazing on the Soil Infiltration Performance
3.6. Effect of Grazing on the Soil Nutrients
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, Y.; Hou, L.; Zhang, Z.; Zhang, J.; Cheng, J.; Wei, G.; Lin, Y. Soil microbial diversity during 30 years of grassland restoration on the Loess Plateau, China: Tight linkages with plant diversity. Land Degrad. Dev. 2019, 30, 1172–1182. [Google Scholar] [CrossRef]
- Liu, M.; Dries, L.; Heijman, W.; Huang, J.; Zhu, X.; Hu, Y.; Chen, H. The Impact of Ecological Construction Programs on Grassland Conservation in Inner Mongolia, China. Land Degrad. Dev. 2018, 29, 326–336. [Google Scholar] [CrossRef]
- Guo, Q. Responses of grassland ecosystem productivity to altered precipitation regime: A review. Chin. J. Appl. Ecol. 2019, 30, 2201–2210. [Google Scholar]
- Wilcox, K.R.; Shi, Z.; Gherardi, L.A.; Lemoine, N.P.; Koerner, S.E.; Hoover, D.L.; Bork, E.; Byrne, K.M.; Cahill, J., Jr.; Collins, S.L.; et al. Asymmetric responses of primary productivity to precipitation extremes: A synthesis of grassland precipitation manipulation experiments. Glob. Chang. Biol. 2017, 23, 4376–4385. [Google Scholar] [CrossRef]
- Browning, D.M.; Maynard, J.J.; Karl, J.W.; Peters, D.C. Breaks in MODIS time series portend vegetation change: Verification using long-term data in an arid grassland ecosystem. Ecol. Appl. 2017, 27, 1677–1693. [Google Scholar] [CrossRef]
- Fay, P.A.; Kaufman, D.M.; Nippert, J.B.; Carlisle, J.D.; Harper, C.W. Changes in grassland ecosystem function due to extreme rainfall events: Implications for responses to climate change. Glob. Chang. Biol. 2010, 14, 1600–1608. [Google Scholar] [CrossRef]
- Wang, G.; Yang, C.; Wang, M. Measurement and Spatial Distribution of the Development Level of Animal Husbandry’s Modernization in China. J. Huazhong Agric. Univ. (Soc. Sci. Ed.) 2018, 6, 7–13+150–151. [Google Scholar]
- Shi, S.D. Output value and production situation of animal husbandry in China. China Anim. Husb. 2016, 14, 42–43. [Google Scholar]
- Ministry of Water Resources of the People’s Republic of China. Bulletin on soil and water conservation of the first national water conservancy census. Soil Water Conserv. China 2013, 11, 2–3. [Google Scholar]
- Ren, J. Pratacul Turalthesarus; Agricultural Press of China: Beijing, China, 2008; p. 851. [Google Scholar]
- Meng, Z.; Dang, X.; Gao, Y.; Ren, X.; Ding, Y.; Wang, M. Interactive effects of wind speed, vegetation coverage, and soil moisture in controlling wind erosion in a temperate desert steppe, Inner Mongolia of China. Arid Zone Sci. Engl. 2018, 10, 14. [Google Scholar] [CrossRef]
- Xu, A.; Xu, D.; Liu, J. Point pattern analysis of dominant populations based on null models in the desert steppe in Ningxia. Acta Ecol. Sin. 2020, 40, 4180–4187. [Google Scholar]
- Liao, H.; Tuvshintogtokh, I.; Guo, T.; Zhao, J. Effects of Grazing Exclusion on the Vegetation Community Composition and the Community Stability of Dry Steppe and Mountain Steppe Ecosystems in Mongolia. Acta Sci. Nat. Univ. Pekin. 2020, 56, 471–478. [Google Scholar]
- Donovan, M.; Monaghan, R. Impacts of grazing on ground cover, soil physical properties, and soil loss via surface erosion: A novel geospatial modeling approach. J. Environ. Manag. 2021, 287, 112206. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wu, L.; Chen, D.; Wu, Y.; Hu, S.; Li, L.; Bai, Y. Grazing simplifies soil micro-food webs and decouples their relationships with ecosystem functions in grasslands. Glob. Chang. Biol. 2020, 26, 960–970. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.U.; Yang, T.; Gao, Y.; Yao, G.; Dang, X.; Zhang, G. Dynamic distribution characteristics of soil microorganisms under different grazing intensities in Stipa klemenzii desert grassland. J. Arid Land Resour. Environ. 2016, 30, 180–185. [Google Scholar]
- Guo, J.; Dong, Z.; Li, J.; Wang, H.; Liu, T. Effects of Grazing intensity on Soil Physical Properties and Soil Erosion and Sediment Yield in Desert Steppe. Chin. J. Grassl. 2019, 41, 74–82. [Google Scholar]
- Zhu, L.; Qin, F.; Yang, C.; Ma, X. Drive-Mechanism of Land Erosion in the Cross Redion Between Farmland and Grassland in Vinshan Mountain. Res. Soil Water Conserv. 2008, 15, 34–37. [Google Scholar]
- Xu, M. A review of grassland carrying capacity: Perspective and dilemma for research in China on “forage-livestock balance”. Acta Prataculturae Sin. 2014, 23, 321–329. [Google Scholar]
- Wang, M.J.; Ma, C.S. A study on methods of estimating the carrying capacity of grassland. Grassl. China 1994, 5, 19–22. [Google Scholar]
- Wei, Z.; Han, G.; Yang, J.; Lv, X. The Response of Stipa breviflora Community to Stocking Rate. Chin. J. Grassl. 2000, 6, 1–5. [Google Scholar]
- Clancy, K.; Alba, V.M. Temperature and Time of Day Influence on Double-Ring Infiltrometer Steady-State Infiltration Rates. Soil Sci. Soc. Am. J. 2011, 75, 241–245. [Google Scholar] [CrossRef]
- Bremner, J.M.; Tabatabai, M.A. Use of an ammonia electrode for determination of ammonia in Kjedahl Analysis. Commun. Soil Sci. Plant Anal. 1972, 3, 159–165. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for determination of phosphate in natural waters. Anal. Chim Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Nelson, L.B.; Heidel, H. Soil Analysis Methods as Used in the Iowa State College Soil Testing Laboratory. Iowa State Coll. Agric. Bull. 1952, 57, 1–31. [Google Scholar]
- Keeney, D.R.; Nelson, D.W. Nitrogen-inorganic forms. In Methods of Soil Analysis, Part 2, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Agronomy Monograph; ASA and SSSA: Madison, WI, USA, 1982; Volume 9, pp. 643–698. [Google Scholar]
- Jiang, X.; Yang, Z.; Wang, G. The influence of disturbance on community structure and plant diversity of alpine meadow. Acta Bot. Boreali-Occident. Sin. 2003, 23, 1479–1485. [Google Scholar]
- Wang, M.J.; Zhao, M.L.; Cui, G.W.; Han, G.D. Effect of Grazing Intensities on Vegetation and Soil in Meadow Steppe. Acta Agrestia Sin. 2010, 18, 758–762. [Google Scholar]
- Wang, H.Y.; Dong, Z.; Guo, J.Y. Organic carbon storage properties in Stipa breviflora desert steppe vegetation soil systems under different grazing intensities. Acta Ecol. Sin. 2016, 36, 4617–4625. [Google Scholar]
- Curtin, C.G.; Sayre, N.F.; Lane, B.D. Transformations of the Chihuahuan Borderlands: Grazing, fragmentation, and biodiversity conservation in desert grasslands. Environ. Sci. Policy 2002, 5, 55–68. [Google Scholar] [CrossRef]
- Almeida, M.; Azeda, C.; Guiomar, N.; Pinto-Correia, T. The effects of grazing management in montado fragmentation and heterogeneity. Agrofor. Syst. 2016, 90, 69–85. [Google Scholar] [CrossRef]
- Das, D.; Jha, N.K.; Maikhuri, R.K. Fragmentation of Pastoral Grazing Landscape and Herd Migratory Routes: A Case Study from Indian Central Himalaya. Res. Lab. Agric. Biotechnol. Biochem. 2015, 9, 18–23. [Google Scholar] [CrossRef]
- Ding, H.-J.; Han, G.-D.; Wang, Z.-W.; Wang, C.-X.; Zhang, R.-Y.; Hu, J.-Y. Effect of Different Stocking Rates on Plant Community Characteristics in Stipa breviflora Desert Steppe. Chin. J. Grassl. 2014, 36, 55–60. [Google Scholar]
- Dong, Q.M.; Zhao, X.Q.; Yu-Shou, M.A.; Shi, J.J.; Wang, Y.L.; Li, S.-X.; Yang, S.H.; Shen, L. Effect of Grazing Intensity on Community Character and Aboveground Present Biomass of Alpine Mixed-sown Pasture. J. Grassl. 2012, 20, 10–16. [Google Scholar]
- Yang, D.; Han, G.; Hu, Y. Effects of grazing intensity on plant diversity and aboveground biomass of Stipa baicalensis grassland. Chin. J. Ecol. 2006, 25, 1470–1475. [Google Scholar]
- Huang, C.; Zhang, Y.; Zhao, M.L.; Han, G.D. Effects of different grazing intensities on vegetation characteristics of desert steppe. Pratacultural Sci. 2013, 30, 1814–1818. [Google Scholar]
- Zhang, F.; Yang, Y.; Qiao, J.R.; Jia, L.X.; Zhao, T.Q.; Zhao, M.L. Effects of Utilization Ways on Species Diversity, Functional Traits and Aboveground Biomass in Stipagrandis Steppe. Chin. J. Grassl. 2019, 41, 1–8. [Google Scholar]
- Schönbach, P.; Wan, H.; Gierus, M.; Bai, Y.; Müller, K.; Lin, L.; Susenbeth, A.; Taube, F. Grassland responses to grazing: Effects of grazing intensity and management system in an Inner Mongolian steppe ecosystem. Plant Soil 2011, 340, 103–115. [Google Scholar] [CrossRef]
- Duan, M.J.; Gao, Q.Z.; Wan, Y.F.; Li, Y.; Guo, Y.Q.; Danjiu, L.B.; Luosang, J.C. Effect of grazing on community characteristics and species diversity of Stipa purpurea alpine grassland in Northern Tibet. Acta Ecol. Sin. 2010, 30, 3892–3900. [Google Scholar]
- Yang, L.; Han, M.; Li, J. Plant diversity change in grassland communities along a grazing disturbance gradient in the northeast China transect. Chin. J. Plant Ecol. 2001, 25, 110–114. [Google Scholar]
- Zhao, L.; Zhong, H.; Zhao, M. Effect of Enclosure and Grazed Management on Aboveground Biomass and Species Diversity in Sandy Grasslands of Horain Sandy Land, Eastern Inner Mongolia, China. Ecol. Environ. Sci. 2018, 27, 1783–1790. [Google Scholar]
- Neff, J.C.; Reynolds, R.L.; Belnap, J.; Lamothe, P. Multi-decadal impacts of grazing on soil physical and biogeochemical properties in southeast Utah. Ecol. Appl. 2005, 15, 87–95. [Google Scholar] [CrossRef]
- Russell, J.R.; Betteridge, K.; Costall, D.A.; Mackay, A.D. Cattlet reading effects on sediment loss and water infiltration. J. Range Manag. 2001, 54, 184–190. [Google Scholar] [CrossRef]
- Zhang, M.; Guo, J.; Dong, Z.; Hongli, L.I.; Jinrong, L.I.; Zhou, X.; Han, X.; Chen, P. Simulation of Runoff and Sediment Yield in Desert Steppe with Different Grazing lntensities. J. Soil Water Conserv. 2018, 32, 41–46. [Google Scholar]
- Wang, Q.; Meng, Z.; Wang, J.; Dang, X.; Zhang, X.; Wang, Z.; Zhao, Q.; Zhang, J. Response of the vegetation soil under almost-natural restoration in the Xilamuren grassland. Acta Ecol. Sin. 2017, 37, 1159–1167. [Google Scholar]
- Gao, Y.; Han, X.; Wang, S. The effects of grazing on grassland soils. Acta Ecol. Sin. 2004, 24, 790–797. [Google Scholar]
- Negrón, M.; López, I.; Drner, J. Consequences of intensive grazing by dairy cows of contrasting live weights on volcanic ash topsoil structure and pasture dynamics. Soil Tillage Res. 2019, 189, 88–97. [Google Scholar] [CrossRef]
- Reszkowska, A.; Krümmelbein, J.; Gan, L.; Peth, S.; Horn, R. Influence of grazing on soil water and gas fluxes of two Inner Mongolian steppe ecosystems. Soil Tillage Res. 2011, 111, 180–189. [Google Scholar] [CrossRef]
- Hughes, J.D.; Packer, I.J.; Michalk, D.L.; Dowling, P.M.; King, W.M.; Brisbane, S.; Millar, G.D.; Priest, S.M.; Kemp, D.R.; Koen, T.B. Sustainable grazing systems for the Central Tablelands of New South Wales. 4. Soil water dynamics and runoff events for differently-managed pasture types. Aust. J. Exp. Agric. 2006, 46, 483–494. [Google Scholar] [CrossRef]
- Mapfumo, E.; Chanasyk, D.S.; Willms, W.D. Simulating daily soil water under foothills fescue grazing with the soil and water assessment tool model (Alberta, Canada). Hydrol. Process. 2010, 18, 2787–2800. [Google Scholar] [CrossRef]
- Li, J.; Yao, N.; Zhao, Y.; Fan, T.; Zhang, J.; Lan, Z.; Yi, J.; Si, B. Characteristics of soil water distribution and evaluation of recharge rate under different grazing history in the Xilin Gol Steppe. Chin. J. Plant Ecol. 2018, 42, 1033–1042. [Google Scholar] [CrossRef]
- Zhang, Y.; Hang, J.; Li, Z. A Study of the Effects of Different Grazing Intensities on Soil Physical Properties. Acta Agrestia Sin. 2002, 10, 74–78. [Google Scholar]
- Zhang, X.; Zhang, W.; Sai, X.; Chun, F.; Li, X.; Lu, X.; Wang, H. Grazing altered soil aggregates, nutrients, and enzyme activities in a Stipa kirschnii steppe of Inner Mongolia. Soil Tillage Res. 2022, 219, 105327. [Google Scholar] [CrossRef]
- Roberts, A.J.; Johnson, N.C. Effects of Mob-Grazing on Soil and Range Quality Vary with Plant Species and Season in a Semiarid Grassland. Rangel. Ecol. Manag. 2021, 79, 139–149. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Plants | Importance Values (%) | |||
|---|---|---|---|---|
| CK | LG | MG | HG | |
| Heteropappus altaicus | 24.9 | 18.34 | 17.55 | 9.88 |
| Carex duriuscula | 14.75 | - | 10.94 | 15.94 |
| Stipa krylovii | 14.28 | 19.25 | 15.21 | 18.45 |
| Cleistogenes squarrosa | 12.15 | 22.46 | 16.89 | 18.08 |
| Artimisia gmelinii | 12.15 | 6.19 | 4.93 | - |
| Agropyron cristatum | 11.74 | 13.83 | 16.23 | 16.33 |
| Convolvulus ammannii | 10.65 | 8.47 | 10.2 | 12.34 |
| Leymus chinensis | 9.84 | 9.04 | 9.88 | 9.42 |
| Kochia prostrata | 9.33 | 5.72 | 6.93 | 4.11 |
| Allium tenuissimum | 7.92 | 5.76 | 4.08 | 8.32 |
| Androsace incana | 7.12 | - | - | 4.39 |
| Arenaria juncea | 6.74 | - | - | - |
| Melilotoides ruthenica | 6.49 | 4.8 | 3.21 | 4.84 |
| Iris tenuifolin | 5.54 | 8.15 | 6.04 | - |
| Haplophyllum dauricum | 4.99 | - | - | - |
| Linum perenne | 4.99 | 4.97 | 2.29 | 4.07 |
| Gentiana dahurica | 3.6 | - | - | 2.64 |
| Hedysarum brachypterum | 3.59 | 3.25 | 4.04 | 4.09 |
| Ptilotrichum canescens | 3.56 | - | - | - |
| Astragalus.discolor | 3.33 | 4.03 | 3.43 | 3.88 |
| Artimisia frigida | 3.04 | 5.75 | 2.91 | 4.98 |
| Iris lactea | 2.77 | - | - | - |
| Community Characteristics | CK | LG | MG | HG | F | p |
|---|---|---|---|---|---|---|
| Cover (%) | 43 ± 3 a | 41 ± 3 a | 37 ± 3 b | 30 ± 3 c | 35.508 | 0.000 |
| Height (cm) | 10.35 ± 6.22 a | 5.93 ± 3.89 b | 5.82 ± 4.25 b | 4.46 ± 3.37 c | 33.235 | 0.000 |
| Above-ground biomass (g) | 86.97 ± 9.53 a | 49.89 ± 6.74 b | 38.19 ± 5.60 c | 42.15 ± 11.83 b | 32.371 | 0.000 |
| Patrick species richness index | 8.91 ± 2.23 b | 8.28 ± 1.21 c | 8.82 ± 2.12 b | 9.64 ± 1.37 a | 0.978 | 0.413 |
| Simpson dominance index | 0.79 ± 0.08 a | 0.75 ± 0.11 b | 0.78 ± 0.07 a | 0.78 ± 0.02 a | 0.515 | 0.674 |
| Shannon–Wiener diversity index | 1.69 ± 0.27 a | 1.41 ± 0.21 b | 1.62 ± 0.19 b | 1.68 ± 0.12 a | 4.850 | 0.006 |
| Pielou evenness index | 0.79 ± 0.09 a | 0.67 ± 0.07 b | 0.76 ± 0.07 a | 0.77 ± 0.03 a | 5.602 | 0.003 |
| Grazing Intensities | Soil Depth | Sand | Fines | Clay Particles | ||||
|---|---|---|---|---|---|---|---|---|
| Very Coarse Sand | Coarse Sand | Medium Sand | Fine Sand | Very Fine Sand | ||||
| CK | 0–5 | 1.22 ± 0.19 b | 6.86 ± 0.79 c | 9.93 ± 1.81 b | 16.17 ± 1.64 a | 17.41 ± 1.56 a | 45.61 ± 5.06 a | 2.80 ± 0.49 a |
| LG | 0–5 | 2.13 ± 0.23 b | 10.52 ± 1.64 b | 13.26 ± 3.07 a | 16.65 ± 1.45 a | 16.10 ± 1.94 a | 39.10 ± 6.33 b | 2.23 ± 0.41 b |
| MG | 0–5 | 2.93 ± 0.67 b | 15.10 ± 1.96 a | 14.63 ± 3.54 a | 14.10 ± 3.70 c | 13.50 ± 0.41 b | 37.53 ± 4.72 c | 2.20 ± 0.38 b |
| HG | 0–5 | 11.80 ± 1.37 a | 9.70 ± 1.60 b | 9.57 ± 1.08 b | 15.61 ± 0.82 b | 15.13 ± 1.76 b | 36.33 ± 1.47 c | 1.85 ± 0.13 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Miao, P.; Zheng, Y.; Guo, J. Impacts of Grazing on Vegetation and Soil Physicochemical Properties in Northern Yinshan Mountain Grasslands. Sustainability 2023, 15, 16028. https://doi.org/10.3390/su152216028
Yang Z, Miao P, Zheng Y, Guo J. Impacts of Grazing on Vegetation and Soil Physicochemical Properties in Northern Yinshan Mountain Grasslands. Sustainability. 2023; 15(22):16028. https://doi.org/10.3390/su152216028
Chicago/Turabian StyleYang, Zhenqi, Ping Miao, Yufeng Zheng, and Jianying Guo. 2023. "Impacts of Grazing on Vegetation and Soil Physicochemical Properties in Northern Yinshan Mountain Grasslands" Sustainability 15, no. 22: 16028. https://doi.org/10.3390/su152216028
APA StyleYang, Z., Miao, P., Zheng, Y., & Guo, J. (2023). Impacts of Grazing on Vegetation and Soil Physicochemical Properties in Northern Yinshan Mountain Grasslands. Sustainability, 15(22), 16028. https://doi.org/10.3390/su152216028