
Exploring Carbohydrate Concentration Fluctuations in Pistachio (Pistacia vera L. cv Uzun) for Deeper Insights into Alternate Bearing Patterns

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
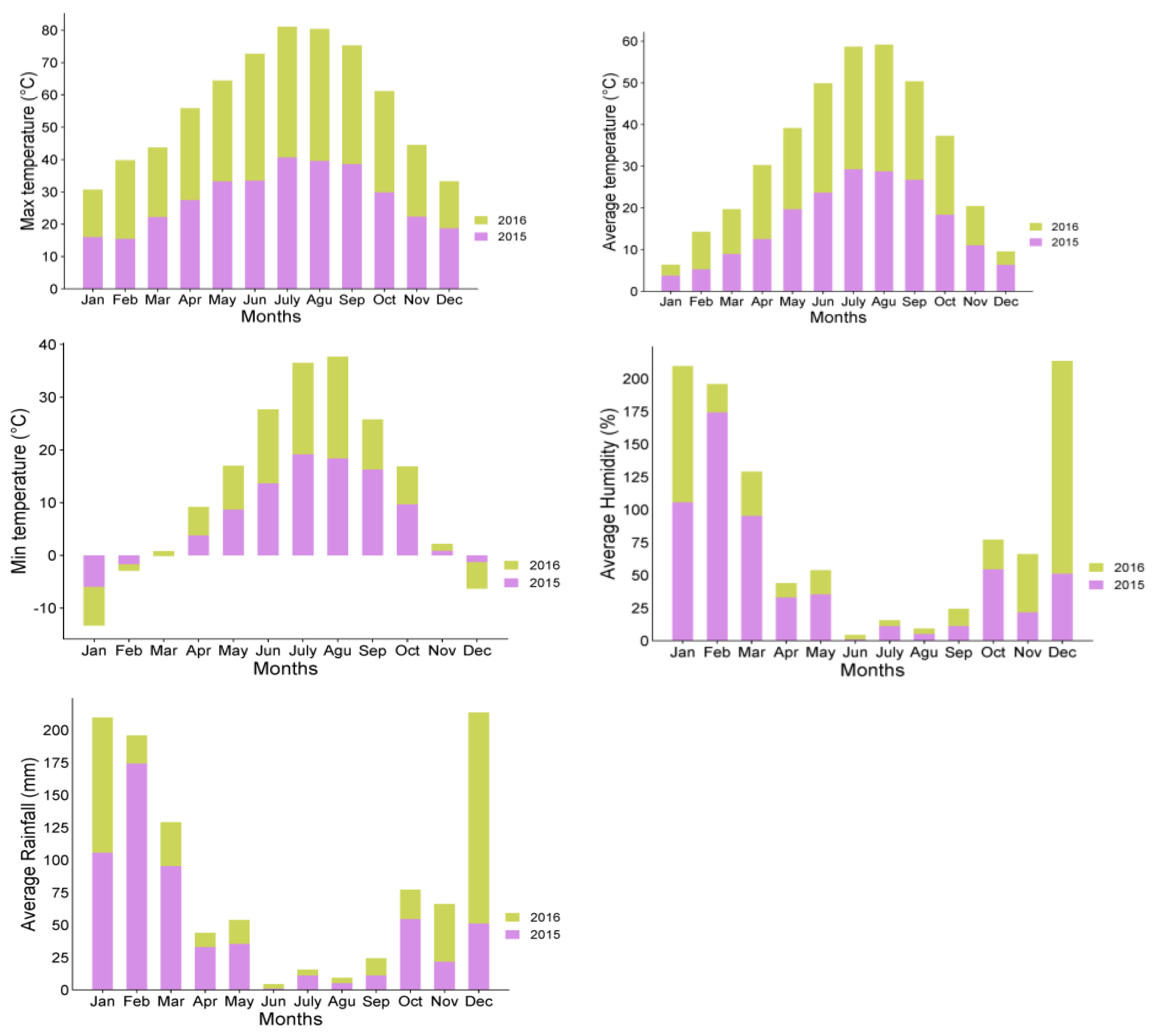
2.1. Plant Materials
2.2. Methods
2.2.1. Determination of Starch Analysis
2.2.2. Determination of Soluble Sugar Analysis
2.2.3. Statistical Analysis
3. Results
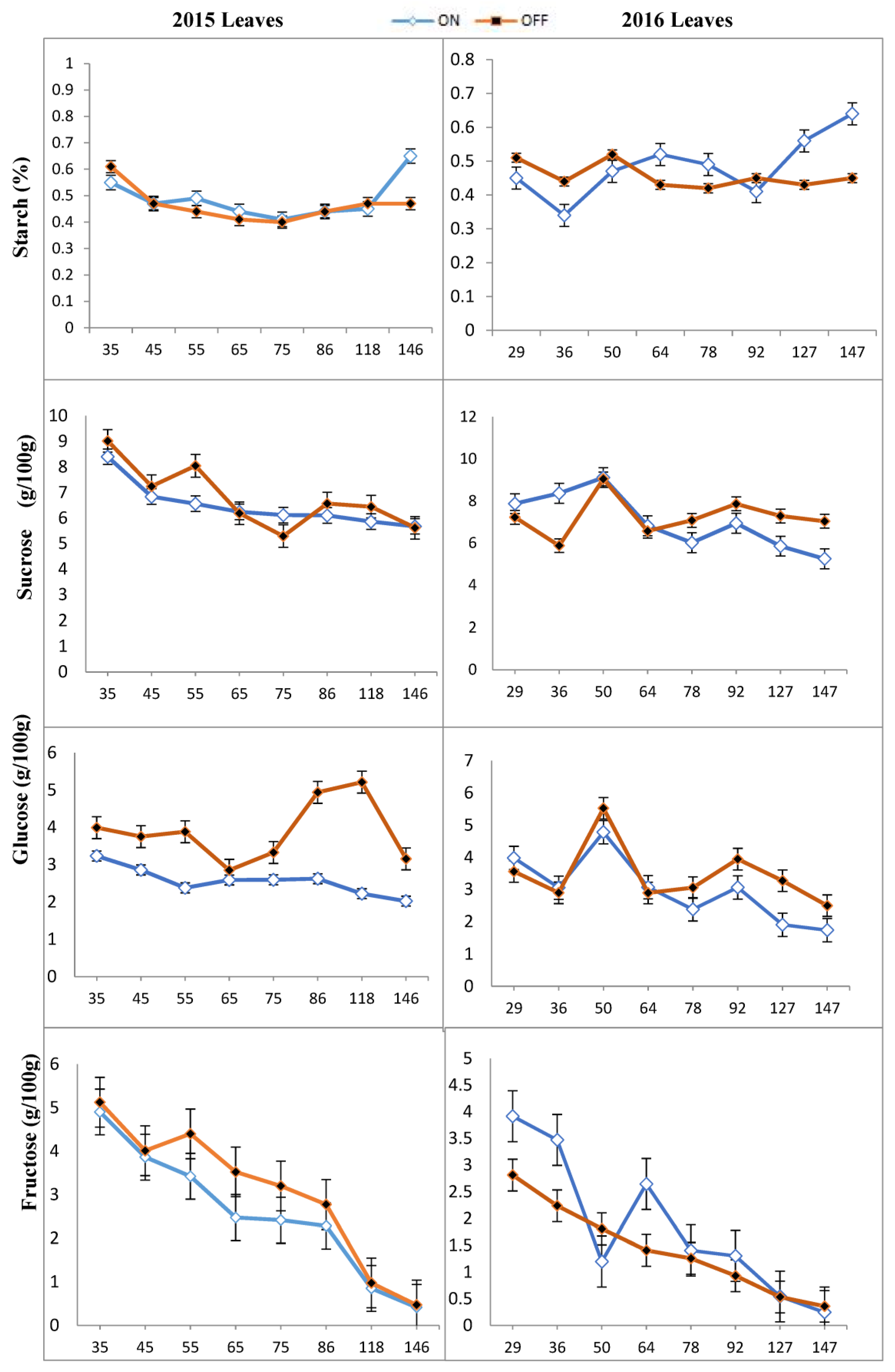
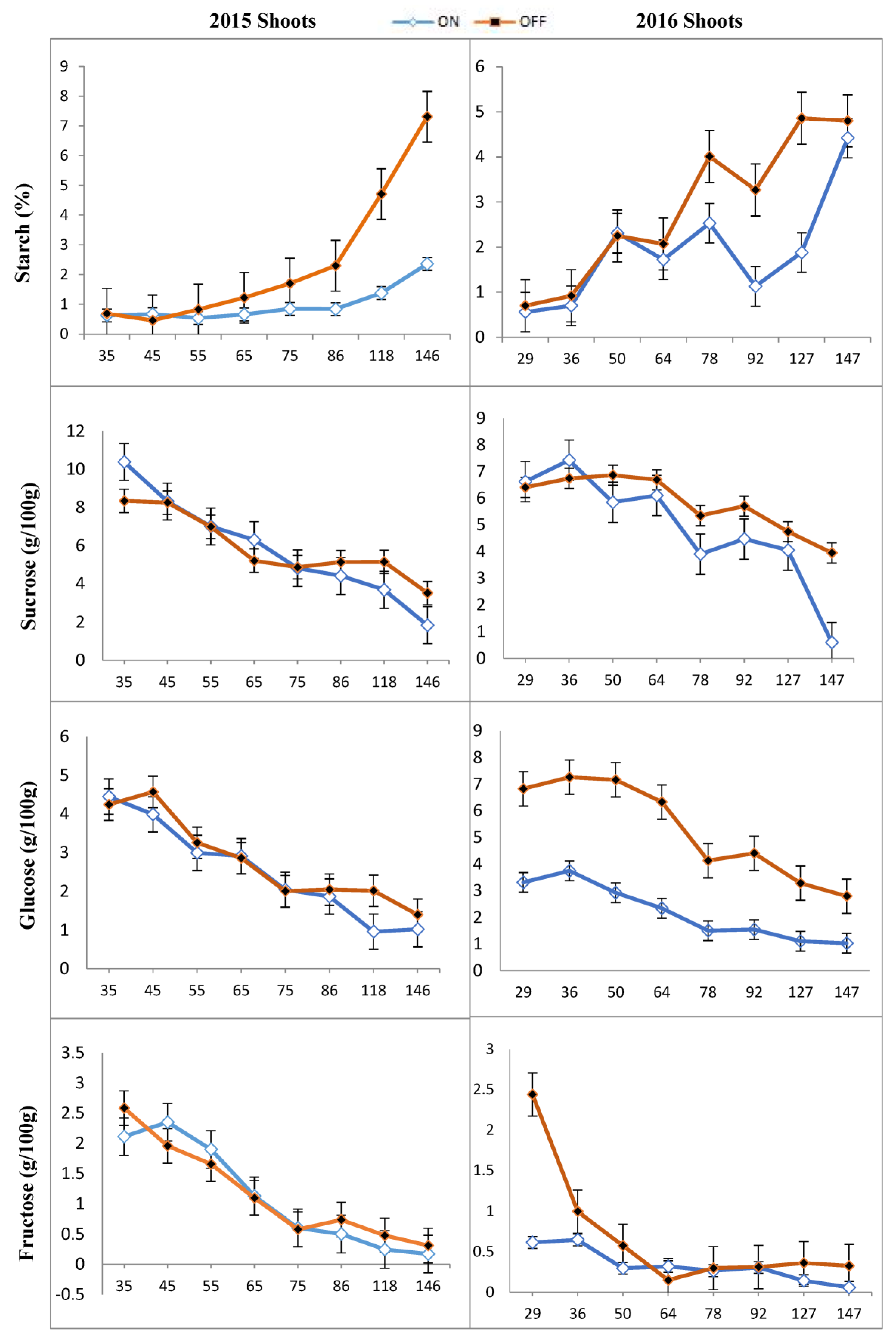
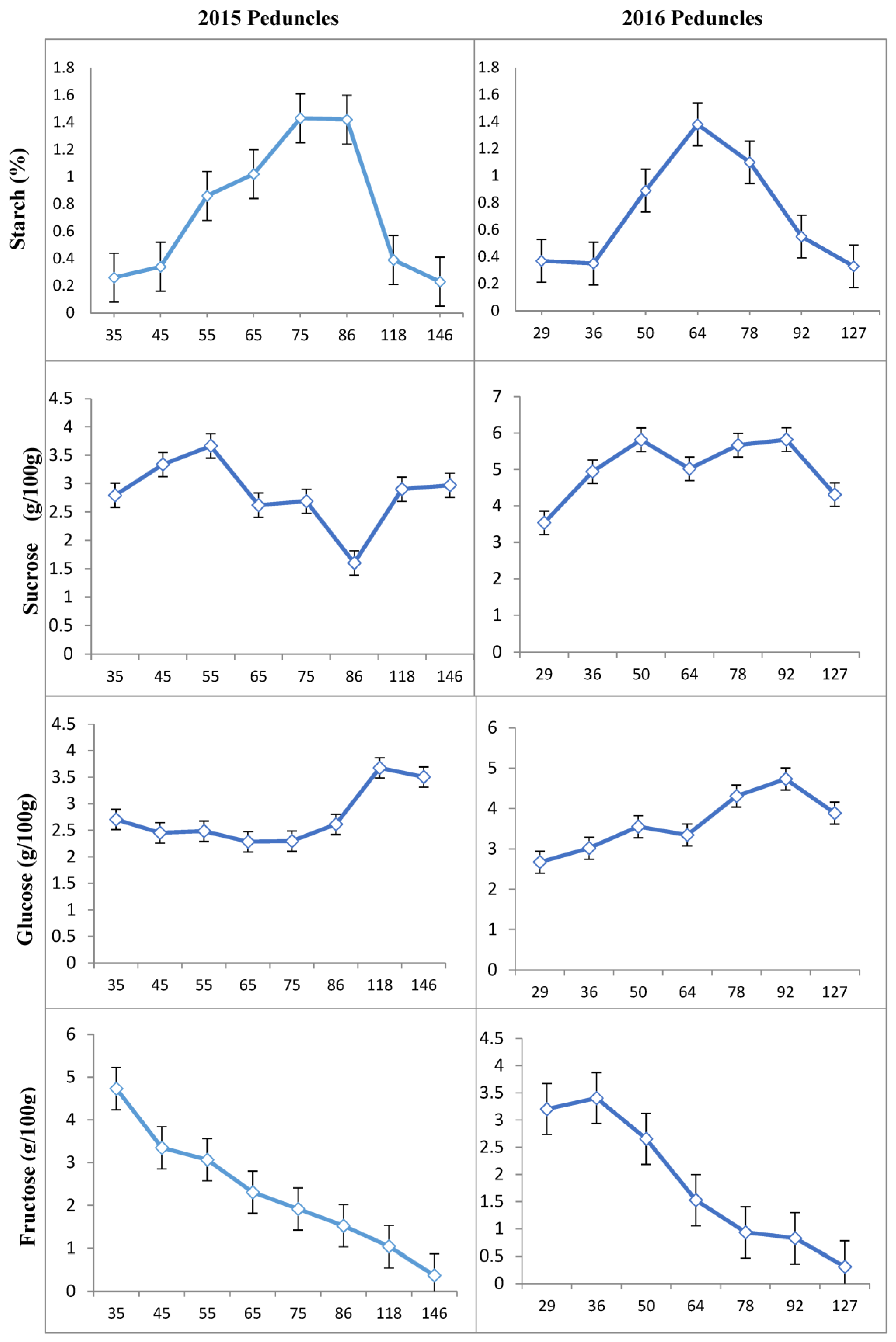
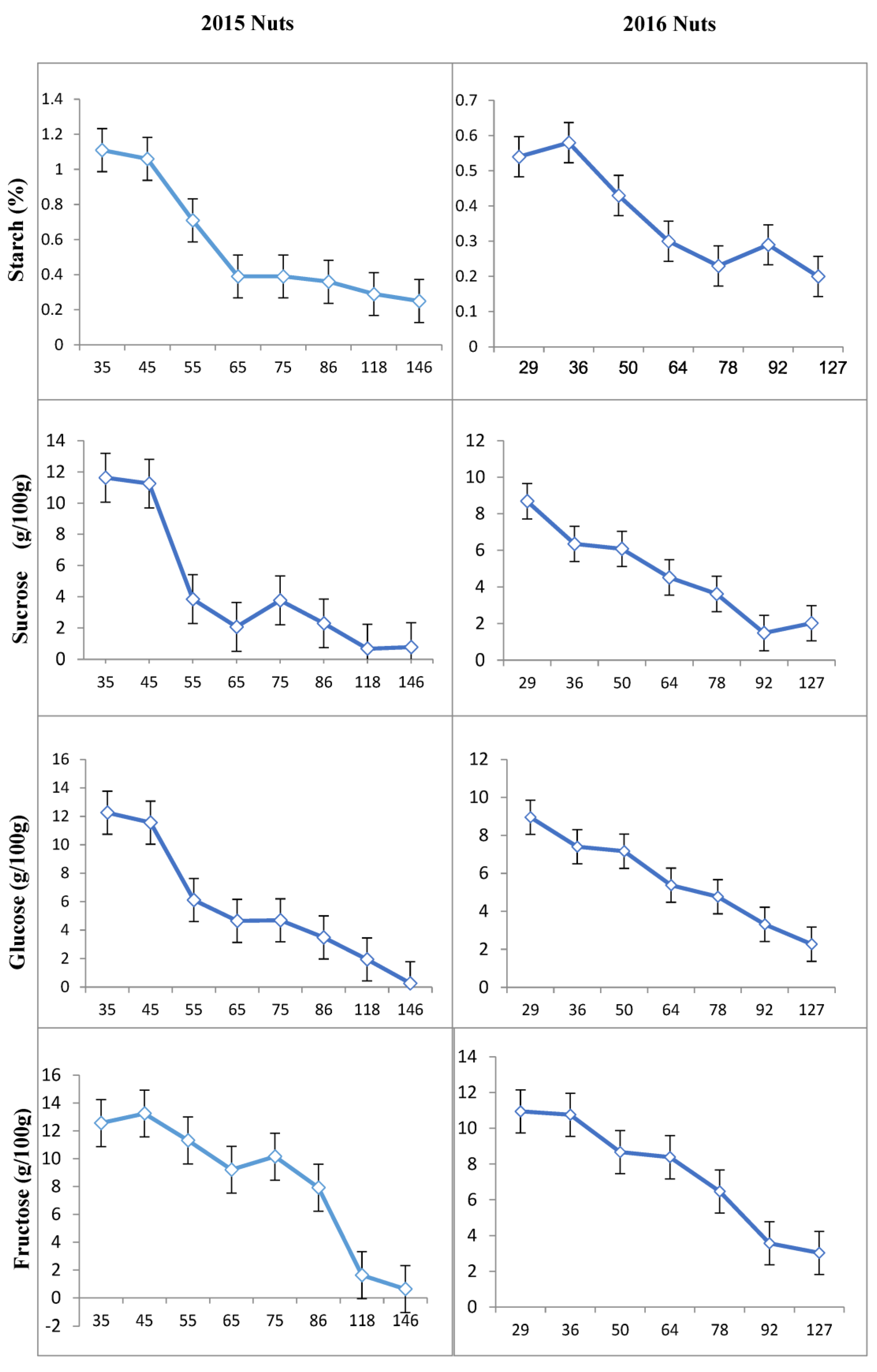
3.1. Concentration of Carbohydrates in Different Tissues of ‘Uzun’ Pistachio
3.2. Mann–Whitney U Test Results
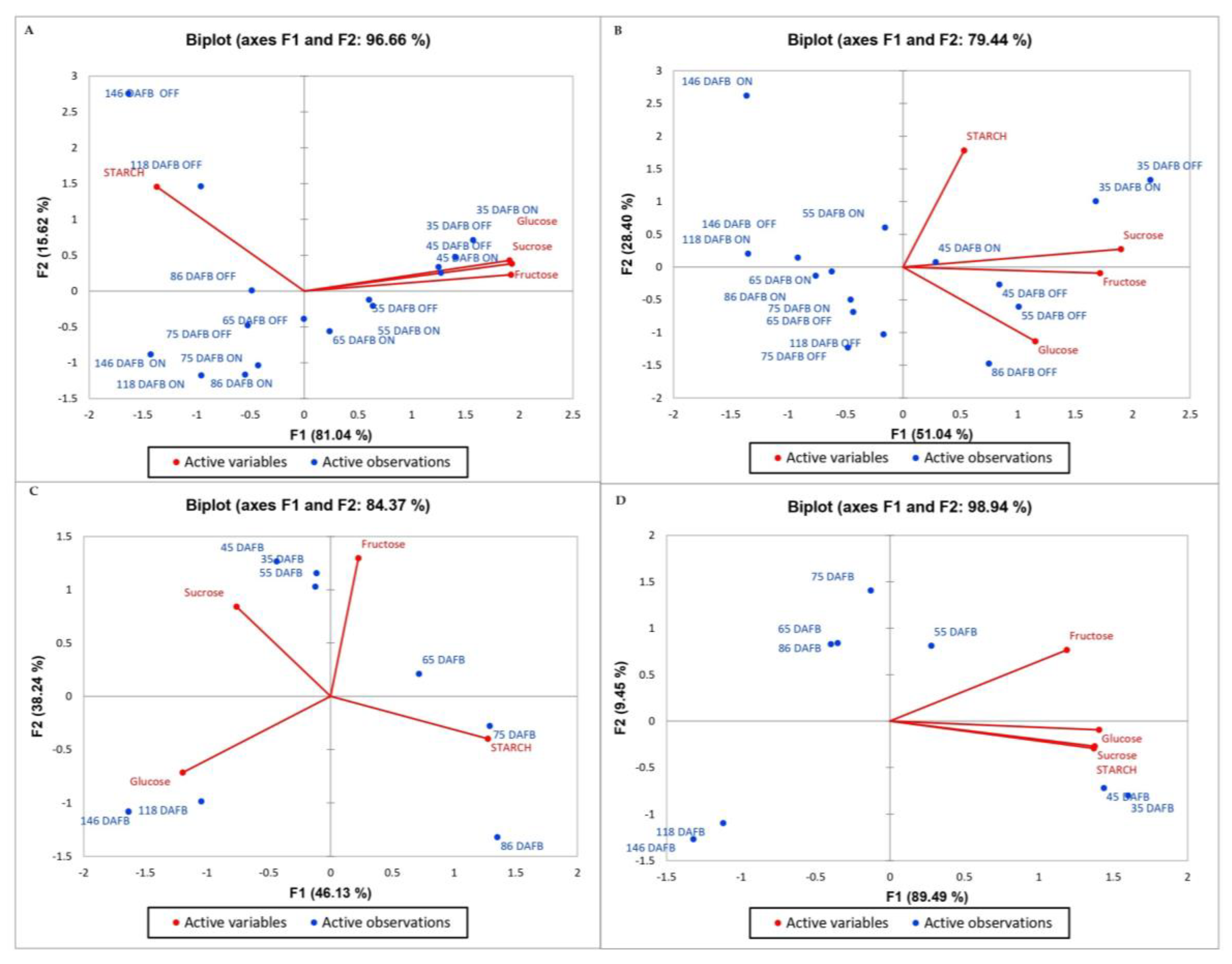
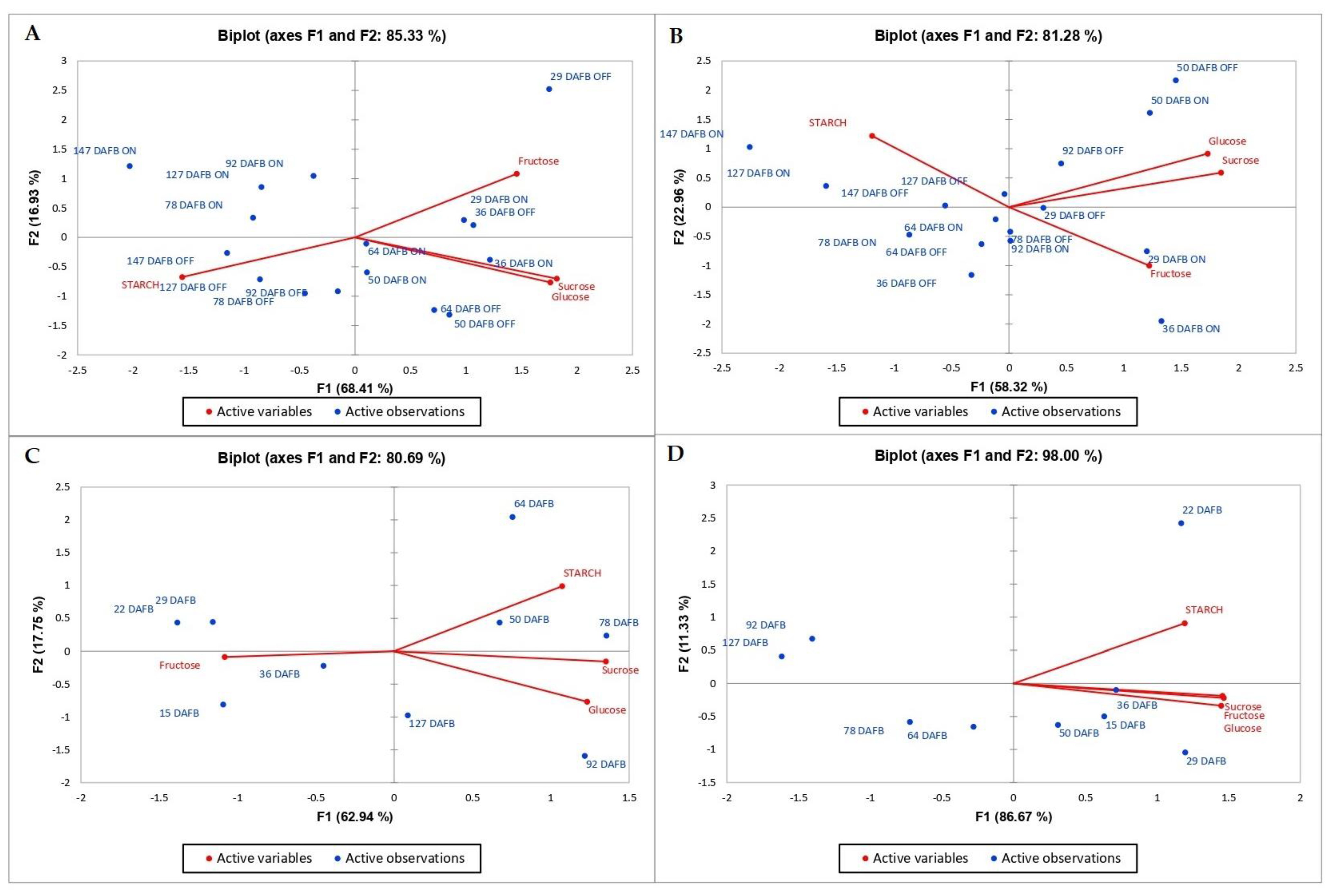
3.3. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gundesli, M.A.; Kafkas, S.; Zarifikhosroshahi, M.; Kafkas, N.E. Role of endogenous polyamines in the alternate bearing phenomenon in pistachio. Turk. J. Agric. For. 2019, 43, 265–274. [Google Scholar] [CrossRef]
- Jangid, R.; Kumar, A.; Masu, M.M.; Kanade, N.; Pant, D. Alternate Bearing in Fruit Crops: Causes and Control Measures. Asian J. Agric. Res. 2023, 10, 10–19. [Google Scholar] [CrossRef]
- Sharma, N.; Singh, S.K.; Mahato, A.K.; Ravishankar, H.; Dubey, A.K.; Singh, N.K. Physiological and molecular basis of alternate bearing in perennial fruit crops. Sci. Hortic. 2019, 243, 214–225. [Google Scholar] [CrossRef]
- Haim, D.; Shalom, L.; Simhon, Y.; Shlizerman, L.; Kamara, I.; Morozov, M.; Sadka, A. Alternate bearing in fruit trees: Fruit presence induces polar auxin transport in citrus and olive stem and represses IAA release from the bud. J. Exp. Bot. 2021, 72, 2450–2462. [Google Scholar] [CrossRef]
- Ali, H.; Abbas, A.; Rehman, A.U. Alternate bearing in fruit plants. J. Agric. Biol. Sci. 2022, 1, 1–6. [Google Scholar] [CrossRef]
- Baninasab, B.; Rahemi, M. Possible role of non-structural carbohydrates in alternate bearing of pistachio. Eur. J. Hortic. Sci. 2006, 71, 277–282. [Google Scholar]
- Vemmos, S.N. Alternate bearing and the possible role of carbohydrates in bud abscission of pistachio (Pistacia vera L.). In Proceedings of the XIV GREMPA Meeting on Pistachios and Almonds, Zaragoza: CIHEAM/FAO/AUA/TEI Kalamatas/NAGREF, Athens, Greece, 30 March–5 April 2010; Volume 94, pp. 9–18. [Google Scholar]
- Zhu, J.; Gou, F.; Rossouw, G.; Begum, F.; Henke, M.; Johnson, E.; Seleznyova, A. Simulating organ biomass variability and carbohydrate distribution in perennial fruit crops: A comparison between the common assimilate pool and phloem carbohydrate transport models. Silico Plants 2021, 3, diab024. [Google Scholar] [CrossRef]
- Goyal, R.K.; Bishnoi, C. Assimilate partitioning and distribution in fruit crops: A review. J. Pharmacogn. Phytochem. 2017, 6, 479–484. [Google Scholar]
- Kozlowski, T.T. Carbohydrate sources and sinks in woody plants. Bot. Rev. 1992, 58, 107–222. [Google Scholar] [CrossRef]
- Crane, J.C.; Catlin, P.B.; Alshalan, I. Carbohydrate levels in the pistachio as related to alternate bearing. J. Am. Soc. Hort. Sci. 1976, 101, 371–374. [Google Scholar] [CrossRef]
- Crane, J.C.; Al Shalan, I.M. Carbohydrate and nitrogen levels in pistachio as related to shoot extension and yield. J. Am. Soc. Hort. Sci. 1977, 102, 396–399. [Google Scholar] [CrossRef]
- Nzima, M.D.; Martin, G.C.; Nishijima, C. Leaf development, dry matter accumulation, and distribution within branches of alternate-bearing ‘kerman’ pistachio trees. J. Am. Soc. Hortic. Sci. 1997, 122, 31–37. [Google Scholar] [CrossRef]
- Mahvelati, N.M.; Archer, L.F.; Marino, G.; Fichtner, E.; Ferguson, L. Pistachio inflorescence bud abscission dynamics as a function of embryo weight, crop load and vegetative growth. Acta Hortic. 2018, 1229, 349–354. [Google Scholar] [CrossRef]
- Spann, T.M.; Beede, R.H.; DeJong, T.M. Seasonal carbohydrate storage and mobilization in bearing and non-bearing pistachio (Pistacia vera) trees. Tree Physiol. 2008, 28, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Vittal, H.; Sharma, N.; Dubey, A.K.; Shivran, M.; Singh, S.K.; Meena, M.C.; Sharma, R.M. Rootstock-mediated carbohydrate metabolism, nutrient contents, and physiological modifications in regular and alternate mango (Mangifera indica L.) scion varieties. PLoS ONE 2023, 18, e0284910. [Google Scholar] [CrossRef]
- Vemmos, S.N. Effects of shoot girdling on bud abscission, carbohydrate and nutrient concentrations in pistachio (Pistacia vera L.). J. Hortic. Sci. Biotech. 2005, 80, 529–536. [Google Scholar] [CrossRef]
- Mccleary, B.V.; Gibson, T.S.; Mugford, D.C. Measurement of total starch in cereal products by amyloglucosidase—α-amylase method: Collaborative study. J. AOAC Int. 1997, 80, 571–579. [Google Scholar] [CrossRef]
- Kara, D. Evaluation of trace metal concentrations in some herbs and herbal teas by principal component analysis. Food Chem. 2009, 114, 347–354. [Google Scholar] [CrossRef]
- Gundesli, M.A. Determination of Sugar, Total Phenol contents-and Antioxidant Activity of various parts ‘Uzun’ pistachio cultivar (Pistacia vera L.). Int. J. Agric. Environ. Food Sci. 2020, 4, 62–69. [Google Scholar]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.D.; Ogé, L.; Hamama, L.; Atanassova, R. The sugar-signaling hub: Overview of regulators and interaction with the hormonal and metabolic network. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef]
- Takeda, F.; Crane, J.C. Abscisic acid in pistachio as related to inflorescence bud abscission [and consequent alternate bearing]. J. Am. Soc. Hortic. Sci. 1980, 105, 573–576. [Google Scholar] [CrossRef]
- Gómez-González, S.; Ruiz-Jiménez, J.; Priego-Capote, F.; Luque de Castro, M.D. Qualitative and quantitative sugar profiling in olive fruits, leaves, and stems by gas chromatography-tandem mass spectrometry (GC-MS/MS) after ultrasound-assisted leaching. J. Agric. Food Chem. 2010, 58, 12292–12299. [Google Scholar] [CrossRef]
- Nzima, M.D.; Martin, G.C.; Nishijima, C. Seasonal changes in total nonstructural carbohydrates within branches and roots of naturally “OFF” and “ON” Kerman’ pistachio trees. J. Am. Soc. Hortic. Sci. 1997, 122, 856–862. [Google Scholar] [CrossRef]
- Ho, L.C.; Baker, D.A. Regulation of loading and unloading in long distance transport systems. Phys. Plant 1982, 56, 225–230. [Google Scholar] [CrossRef]
- Schaffer, A.A.; Goldschmidt, E.E.; Goren, R.; Galili, D. Fruit set and carbohydrate status in alternate and nonalternate bearing Citrus cultivars. J. Am. Soc. Hort. Sci. 1985, 110, 574–578. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year 2015 | Year 2016 | ||
|---|---|---|---|
| Date | * DAFB | Date | DAFB |
| - | - | 22 April | 15 |
| - | - | 29 April | 22 |
| 15 May | 35 | 6 May | 29 |
| 25 May | 45 | 13 May | 36 |
| 4 June | 55 | 27 May | 50 |
| 14 June | 65 | 10 June | 64 |
| 24 June | 75 | 24 June | 78 |
| 5 July | 86 | 8 July | 92 |
| 6 August | 118 | 12 August | 127 |
| 3 September | 146 | 1 September | 147 |
| Sucrose | Glucose | Fructose | Starch | ||
|---|---|---|---|---|---|
| shoot | Mann–Whitney U | 28.500 | 27.000 | 31.000 | 21.000 |
| Wilcoxon W | 64.500 | 63.000 | 67.000 | 57.000 | |
| Z | −0.368 | −0.525 | −0.105 | −1.155 | |
| Asymp. Sig. (2-tailed) | 0.713 | 0.600 | 0.916 | 0.248 | |
| leaf | Mann–Whitney U | 22.000 | 3.000 | 25.000 | 25.000 |
| Wilcoxon W | 58.000 | 39.000 | 61.000 | 61.000 | |
| Z | −1.050 | −3.046 | −0.736 | −0.747 | |
| Asymp. Sig. (2-tailed) | 0.294 | 0.002 | 0.462 | 0.455 | |
| Sucrose | Glucose | Fructose | Starch | ||
|---|---|---|---|---|---|
| shoot | Mann–Whitney U | 23.000 | 16.000 | 23.500 | 20.500 |
| Wilcoxon W | 59.000 | 52.000 | 59.500 | 56.500 | |
| Z | −0.945 | −1.680 | −0.893 | −1.209 | |
| Asymp. Sig. (2-tailed) | 0.345 | 0.093 | 0.372 | 0.227 | |
| leaf | Mann–Whitney U | 27.000 | 25.500 | 28.000 | 23.500 |
| Wilcoxon W | 63.000 | 61.500 | 64.000 | 59.500 | |
| Z | −0.525 | −0.684 | −0.421 | −0.897 | |
| Asymp. Sig. (2-tailed) | 0.600 | 0.494 | 0.674 | 0.370 |
| Regression Estimated for 2015 | Regression Estimated for 2016 | |
|---|---|---|
| sucrose | −0.737 *** | −1.337 * |
| (0.072) | (0.022) | |
| glucose | −0.261 | −0.343 |
| (0.570) | (0.580) | |
| fructose | 0.580 | 0.664 |
| (0.113) | (0.130) | |
| starch | −0.894 | −1.354 * |
| (0.128) | (0.027) | |
| _cons | 6.406 *** | 9.824 * |
| (0.060) | (0.023) | |
| N | 48 | 48 |
| LR Statistics | 18.55 *** (0.0696) | 24.24 ** (0.0117) |
| Pseudo R2 | 0.304 | 0.3967 |
| Hosmer–Lemeshow Test Chi-Square | 37.35 (0.4067) | 36.69 (0.4366) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guney, M.; Gundesli, M.A.; Guney, M.; Kafkas, S.; Kafkas, N.E. Exploring Carbohydrate Concentration Fluctuations in Pistachio (Pistacia vera L. cv Uzun) for Deeper Insights into Alternate Bearing Patterns. Sustainability 2023, 15, 15300. https://doi.org/10.3390/su152115300
Guney M, Gundesli MA, Guney M, Kafkas S, Kafkas NE. Exploring Carbohydrate Concentration Fluctuations in Pistachio (Pistacia vera L. cv Uzun) for Deeper Insights into Alternate Bearing Patterns. Sustainability. 2023; 15(21):15300. https://doi.org/10.3390/su152115300
Chicago/Turabian StyleGuney, Mujgan, Muhammet Ali Gundesli, Murat Guney, Salih Kafkas, and Nesibe Ebru Kafkas. 2023. "Exploring Carbohydrate Concentration Fluctuations in Pistachio (Pistacia vera L. cv Uzun) for Deeper Insights into Alternate Bearing Patterns" Sustainability 15, no. 21: 15300. https://doi.org/10.3390/su152115300
APA StyleGuney, M., Gundesli, M. A., Guney, M., Kafkas, S., & Kafkas, N. E. (2023). Exploring Carbohydrate Concentration Fluctuations in Pistachio (Pistacia vera L. cv Uzun) for Deeper Insights into Alternate Bearing Patterns. Sustainability, 15(21), 15300. https://doi.org/10.3390/su152115300