
Olive Ridley Sea Turtle Incubation in Natural Conditions Is Possible on Guatemalan Beaches

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Natural Nests
2.2. Table of Development
- The CRL/SCL ratio was determined for both C. caretta and C. mydas.
- When only CRL information was available, the equivalent SCL for the stages of C. caretta and C. mydas development was estimated using the previous cross products. The equivalent SCL is the expected carapace size for an embryonic stage without a carapace, while taking into account the allometry between CRL and SCL.
- The SCL of L. olivacea embryos was then determined using the average of the cross product with the known size of SCL for this species and the equivalent SCL sizes of C. caretta and C. mydas embryos.
2.3. Thermal Reaction Norm of Embryo Growth
2.4. Modeling Embryo Growth
2.5. Hatching Success, Stage, and Age at Death
2.6. Dynamic Analysis of Hatching Success
3. Results
3.1. Table of Development for Lepidochelys olivacea
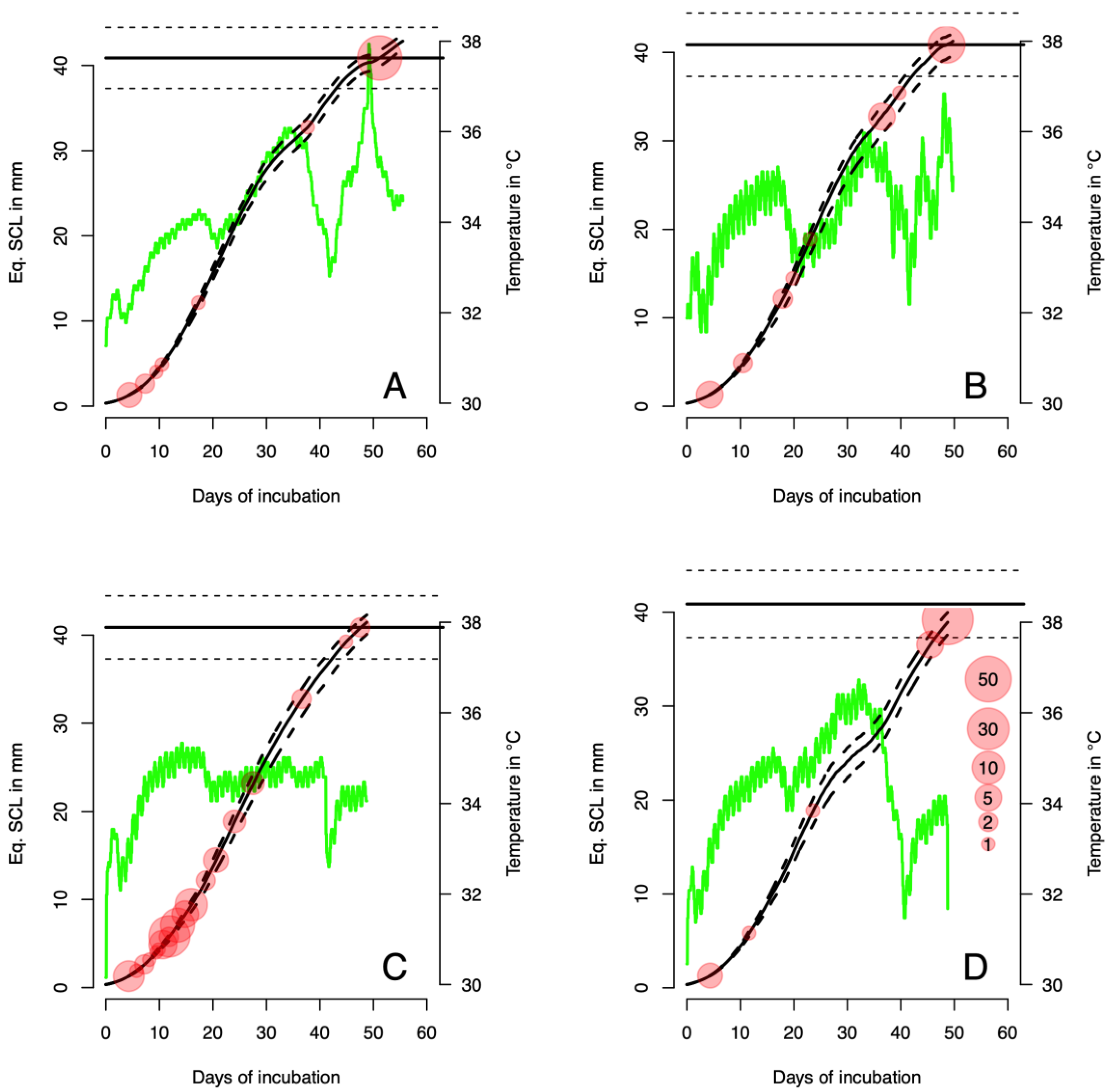
3.2. Temperatures in Natural Nests
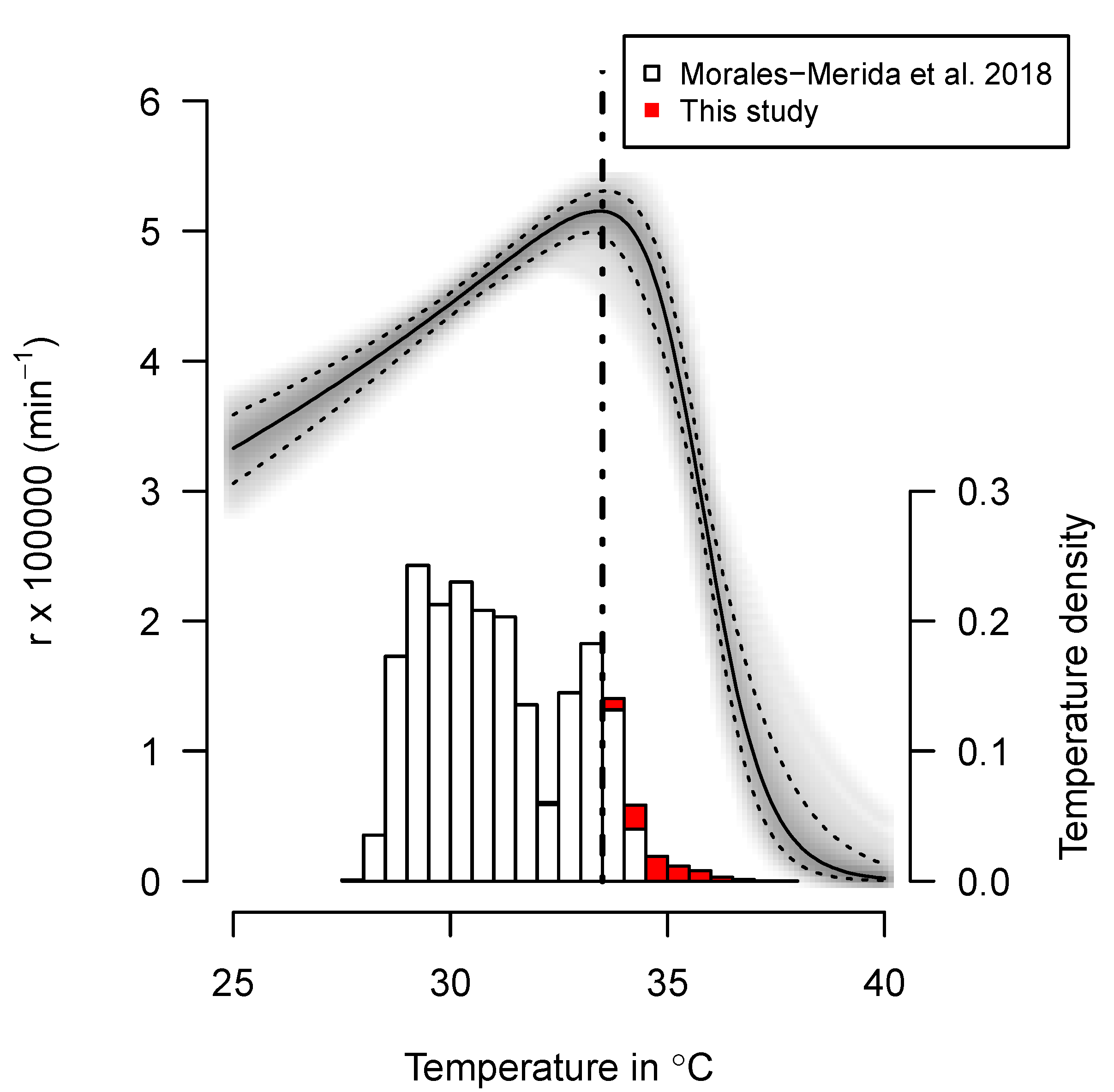
3.3. Thermal Reaction Norm of Embryo Growth
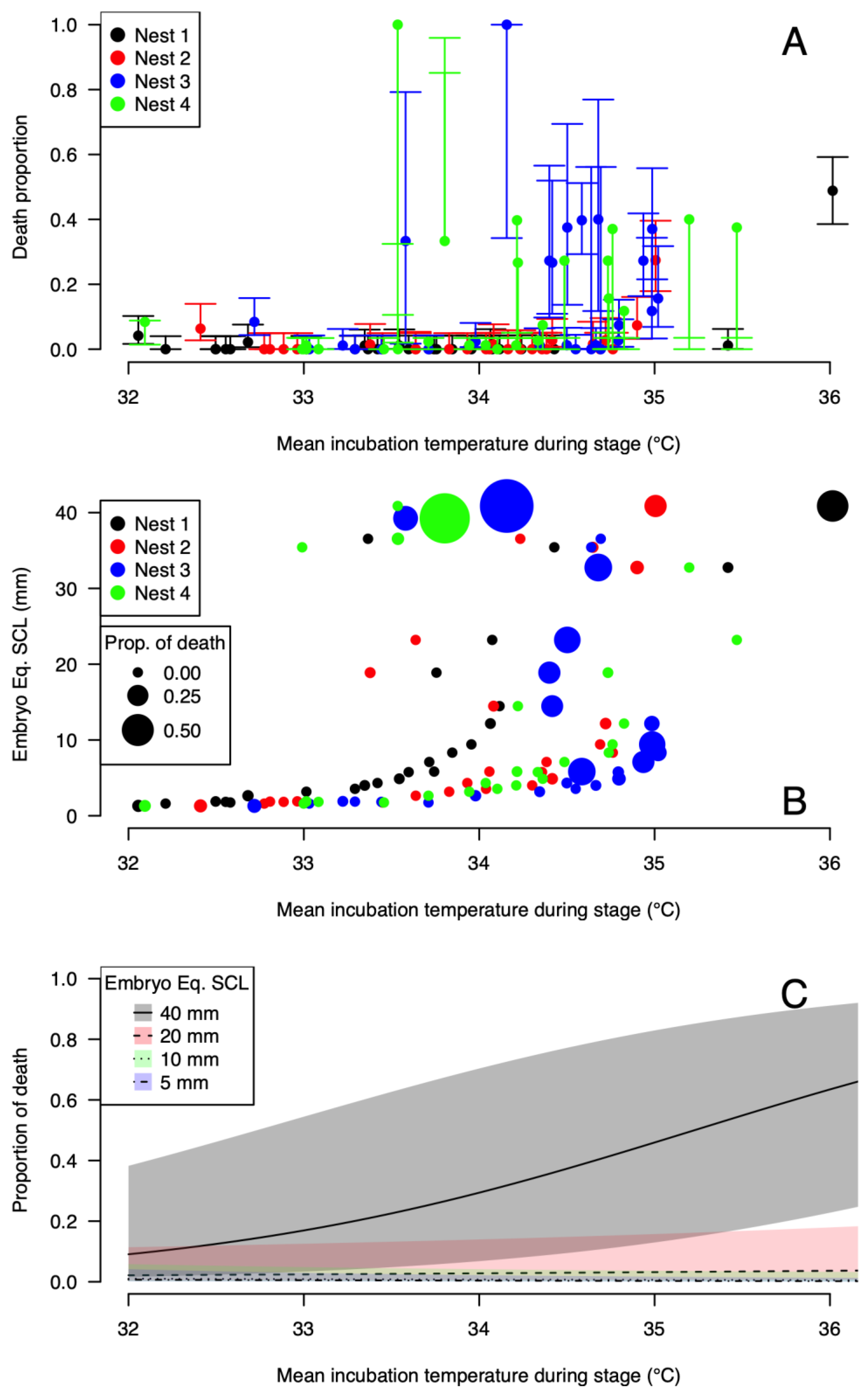
3.4. Stages of Embryo Death
3.5. Dynamic Analysis of Hatching Success
4. Discussion
4.1. Thermal Reaction Norm of Embryo Growth
4.2. Thermal Reaction Norm for Hatching Success
4.3. Implications for the Conservation Strategy in Guatemala
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hart, C.E.; Maldonado-Gasca, A.; Ley-Quiñonez, C.P.; Flores-Peregrina, M.; de Jesús Romero-Villarruel, J.; Aranda-Mena, O.S.; Plata-Rosas, L.J.; Tena-Espinoza, M.; Llamas-González, I.; Zavala-Norzagaray, A.A.; et al. Status of olive ridley sea turtles (Lepidochelys olivacea) after 29 years of nesting rookery conservation in Nayarit and Bahía de Banderas, Mexico. Chelonian Conserv. Biol. 2018, 17, 27–36. [Google Scholar] [CrossRef]
- Abreu-Grobois, A.; Plotkin, P.T. Lepidochelys olivacea; IUCN SSC Marine Turtle Specialist Group: Gland, Switzerland, 2008. [Google Scholar]
- Rguez-Baron, J.M.; Kelez, S.; Liles, M.; Zavala-Norzagaray, A.; Torres-Suárez, O.L.; Amorocho, D.F.; Gaos, A.R. (Eds.) Sea Turtles in the East Pacific Ocean Region; IUCN-SSC Marine Turtle Specialist Group: Gland, Switzerland, 2019; p. 237. [Google Scholar]
- Morales-Mérida, B.A.; Muccio, C.; Girondot, M. Validating trends in olive ridley nesting track counts in Guatemala in the light of a national hatchery protection strategy. Oryx 2023, 57, 48–54. [Google Scholar] [CrossRef]
- CONAP. Estrategia Nacional de Manejo y Conservación de Tortugas Marinas de Guatemala; Consejo Nacional de Áreas Protegidas: Guatemala city, Guatamala, 2015; p. 58. [Google Scholar]
- Juarez, R.; Muccio, C. Sea turtle conservation in Guatemala. Mar. Turt. Newsl. 1997, 77, 15–17. [Google Scholar]
- Muccio, C.; Ortiz, L.; Martinez, J. Manual para la Conservación de las Tortugas Marinas en Guatemala, con un Enfasis en el Manejo de Tortugarios; ARCAS and CNOAP: Guatemala city, Guatemala, 2009; p. 57. [Google Scholar]
- Higginson, J.; Orantes, R. Manejo de Tortugas Marinas; Universidad de San Carlos de Guatemala: Guatemala City, Guatemala, 1988. [Google Scholar]
- Milne Edwards, H.; Vaillant, L. Recherches Zoologiques Pour Servir à L’histoire de la Faune de l’Amérique Centrale et du Mexique; Imprimerie Nationale: Paris, France, 1870–1909; p. 1012. [Google Scholar]
- Taylor, E.N.; Diele-Viegas, L.M.; Gangloff, E.J.; Hall, J.M.; Halpern, B.; Massey, M.D.; Rödder, D.; Rollinson, N.; Spears, S.; Sun, B.j.; et al. The thermal ecology and physiology of reptiles and amphibians: A user’s guide. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2021, 335, 13–44. [Google Scholar] [CrossRef] [PubMed]
- Mueller, M.S.; Ruiz-García, N.A.; García-Gasca, A.; Abreu-Grobois, F.A. Best swimmers hatch from intermediate temperatures: Effect of incubation temperature on swimming performance of olive ridley sea turtle hatchlings. J. Exp. Mar. Biol. Ecol. 2019, 519, 151186. [Google Scholar] [CrossRef]
- Maulany, R.I.; Booth, D.T.; Baxter, G.S. The effect of incubation temperature on hatchling quality in the olive ridley turtle, Lepidochelys olivacea, from Alas Purwo National Park, East Java, Indonesia: Implications for hatchery management. Mar. Biol. 2012, 159, 2651–2661. [Google Scholar] [CrossRef]
- Fuentes, M.M.P.B.; Limpus, C.; Hamann, M. Vulnerability of sea turtle nesting grounds to climate change. Glob. Change Biol. 2011, 17, 140–153. [Google Scholar] [CrossRef]
- Abreu-Grobois, F.A.; Morales-Mérida, B.A.; Hart, C.E.; Guillon, J.-M.; Godfrey, M.H.; Navarro, E.; Girondot, M. Recent advances on the estimation of the thermal reaction norm for sex ratios. PeerJ 2020, 8, e8451. [Google Scholar] [CrossRef]
- Howard, R.; Bell, I.; Pike, D.A. Thermal tolerances of sea turtle embryos: Current understanding and future directions. Endanger. Species Res. 2014, 26, 75–86. [Google Scholar] [CrossRef]
- Laloë, J.-O.; Cozens, J.; Renom, B.; Taxonera, A.; Hays, G.C. Effects of rising temperature on the viability of an important sea turtle rookery. Nat. Clim. Change 2014, 4, 513–518. [Google Scholar] [CrossRef]
- Short, N.M.; Blair, R.W., Jr. (Eds.) Geomorphology from Space: A Global Overview of Regional Landforms; NASA, Scientific and Technical Branch: Washington, DC, USA, 1986; p. 717.
- Marshall, J.S. The geomorphology and physiographic provinces of Central America. In Central America: Geology, Hazards, & Resources; Bundschuh, J., Alvarado, G.E., Eds.; Taylor-Francis: London, UK, 2007; Volume 1, pp. 75–121. [Google Scholar] [CrossRef]
- Ariano-Sánchez, D.; Nesthus, A.; Rosell, F.; Reinhardt, S. Developed black beaches—Too hot to emerge? Factors affecting sand temperatures at nesting grounds of olive ridley sea turtles (Lepidochelys olivacea). Clim. Change Ecol. 2023, 5, 100074. [Google Scholar] [CrossRef]
- Morales-Merida, B.A.; Bustamante, D.M.; Monsinjon, J.; Girondot, M. Reaction norm of embryo growth rate dependent on incubation temperature in the Olive Ridley sea turtle, Lepidochelys olivacea, from Pacific Central America. J. Embryol. 2018, 1, 12–24. [Google Scholar]
- Crastz, F. Embryological stages of the marine turtle Lepidochelys olivacea (Eschscholtz). Rev. De Biol. Trop. 1982, 30, 113–120. [Google Scholar]
- Kaska, Y.; Downie, R. Embryological development of sea turtles (Chelonia mydas, Caretta caretta) in the Mediterranean. Zool. Middle East 1999, 19, 55–69. [Google Scholar] [CrossRef]
- Kaska, Y. Studies on the Embryology, Ecology and Evolution of Sea Turtles in the Eastern Mediterranean. Ph.D. Thesis, University of Glasgow, Glasgow, Walles, 1998. [Google Scholar]
- Miller, J.D. Embryology of marine turtles. In Biology of the Reptilia; Gans, C., Billet, F., Maderson, P.F., Eds.; Wiley-Liss: New-York, NY, USA, 1985; pp. 270–328. [Google Scholar]
- Miller, J.D.; Mortimer, J.A.; Limpus, C.J. A field key to the developmental stages of marine turtles (Cheloniidae) with notes on the development of Dermochelys. Chelonian Conserv. Biol. 2017, 16, 111–122. [Google Scholar] [CrossRef]
- Girondot, M.; Kaska, Y. A model to predict the thermal reaction norm for the embryo growth rate from field data. J. Therm. Biol. 2014, 45, 96–102. [Google Scholar] [CrossRef]
- Sharpe, P.J.H.; DeMichele, D.W. Reaction kinetics of poikilotherm development. J. Theor. Biol. 1977, 64, 649–670. [Google Scholar] [CrossRef] [PubMed]
- Schoolfield, R.M.; Sharpe, P.J.; Magnuson, C.E. Non-linear regression of biological temperature-dependent rate models based on absolute reaction-rate theory. J. Theor. Biol. 1981, 88, 719–731. [Google Scholar] [CrossRef]
- Laird, A.K. Dynamics of tumor growth. Br. J. Cancer 1964, 18, 490–502. [Google Scholar] [CrossRef]
- Morales Mérida, A. Relationship between Incubation Length and the Sex Ratio of the Sea Turtles Lepidochelys olivacea in the Natural Reserve of Multiple Uses, in the Pacific Coast of Guatemala; Universidad de San Carlos de Guatemala: Guatemala City, Guatemala, 2012. [Google Scholar]
- Metropolis, N.; Rosenbluth, A.W.; Rosenbluth, M.N.; Teller, A.H.; Teller, E. Equations of state calculations by fast computing machines. J. Chem. Phys. 1953, 21, 1087–1092. [Google Scholar] [CrossRef]
- Hastings, W.K. Monte Carlo sampling methods using Markov chains and their applications. Biometrika 1970, 57, 97–109. [Google Scholar] [CrossRef]
- Raftery, A.E.; Lewis, S.M. The number of iterations, convergence diagnostics and generic Metropolis algorithms. In Practical Markov Chain Monte Carlo; Gilks, W.R., Spiegelhalter, D.J., Richardson, S., Eds.; Chapman and Hall: London, UK, 1995. [Google Scholar]
- Rosenthal, J.S. Optimal proposal distributions and adaptive MCMC. In MCMC Handbook; Brooks, S., Gelman, A., Jones, G., Meng, X.-L., Eds.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2011; pp. 93–112. [Google Scholar]
- Phillott, A.D.; Godfrey, M.H. Assessing the evidence of « infertile » sea turtle eggs. Endanger. Species Res. 2020, 41, 329–338. [Google Scholar] [CrossRef]
- Phillott, A.D.; Godfrey, M.H.; Avens, L.I. Distinguishing between fertile and infertile sea turtle eggs. Mar. Turt. Newsl. 2021, 162, 18–21. [Google Scholar]
- Wilson, E.B. Probable inference, the law of succession, and statistical inference. J. Am. Stat. Assoc. 1927, 22, 209–212. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Sherlock, C. Optimal scaling of the random walk Metropolis: General criteria for the 0.234 acceptance rule. J. Appl. Probab. 2013, 50, 1–15. [Google Scholar] [CrossRef]
- Monsinjon, J.; Jribi, I.; Hamza, A.; Ouerghi, A.; Kaska, Y.; Girondot, M. Embryonic growth rate thermal reaction norm of Mediterranean Caretta caretta embryos from two different thermal habitats, Turkey and Libya. Chelonian Conserv. Biol. 2017, 16, 172–179. [Google Scholar] [CrossRef]
- Wallace, B.P.; DiMatteo, A.D.; Hurley, B.J.; Finkbeiner, E.M.; Bolten, A.B.; Chaloupka, M.Y.; Hutchinson, B.J.; Abreu-Grobois, F.A.; Amorocho, D.; Bjorndal, K.A.; et al. Regional management units for marine turtles: A novel framework for prioritizing conservation and research across multiple scales. PLoS ONE 2010, 5, e15465. [Google Scholar] [CrossRef]
- Miller, J.D. Reproduction in sea turtles. In The Biology of Sea Turtles; Lutz, P.L., Musick, J.A., Eds.; CRC Press: New York, NY, USA, 1997; pp. 51–81. [Google Scholar]
- Ackerman, R.A. The nest environment and the embryonic development of sea turtles. In The Biology of Sea Turtles; Lutz, P.L., Musick, J.A., Eds.; CRC Press: New York, NY, USA, 1997; pp. 83–106. [Google Scholar]
- Valverde, R.A.; Wingard, S.; Gómez, F.; Tordoir, M.T.; Orrego, C.M. Field lethal incubation temperature of olive ridley sea turtle Lepidochelys olivacea embryos at a mass nesting rookery. Endanger. Species Res. 2010, 12, 77–86. [Google Scholar] [CrossRef]
- Howard, R.; Bell, I.; Pike, D.A. Tropical flatback turtle (Natator depressus) embryos are resilient to the heat of climate change. J. Exp. Biol. 2015, 218, 3330–3335. [Google Scholar] [CrossRef]
- Sönmez, B. Relationship between metabolic heating and nest parameters in green turtles (Chelonia mydas, L. 1758) on Samandağ Beach, Turkey. Zool. Sci. 2018, 35, 243–248. [Google Scholar] [CrossRef]
- Zbinden, J.A.; Margaritoulis, D.; Arlettaz, R. Metabolic heating in Mediterranean loggerhead sea turtle clutches. J. Exp. Mar. Biol. Ecol. 2006, 334, 151–157. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Nest 1 | Nest 2 | Nest 3 | Nest 4 |
|---|---|---|---|---|
| <6 | 4 | 5 | 8 | 4 |
| 6 | 1 | |||
| 11 | 1 | |||
| 12 | 2 | 2 | ||
| 13 | 1 | |||
| 15 | 1 | 1 | ||
| 16 | 1 | |||
| 17 | 1 | 2 | 6 | |
| 18 | 2 | |||
| 19 | 29 | 1 | ||
| 20 | 12 | |||
| 21 | 5 | |||
| 22 | 10 | |||
| 23 | 1 | 2 | 2 | |
| 24 | 1 | 4 | ||
| 25 | 1 | 3 | 1 | |
| 26 | 3 | |||
| 27 | 1 | 5 | 2 | |
| 28 | 1 | |||
| 29 | 5 | |||
| 30 | 1 | 93 | ||
| 31 | 42 | 17 | 2 | |
| Eggs | 96 | 79 | 95 | 112 |
| Hatchlings | 44 | 44 | 0 | 8 |
| Hatching success | 0.458 | 0.557 | 0.000 | 0.071 |
| Stage | Chelonia mydas | Caretta caretta | Lepidochelys olivacea |
|---|---|---|---|
| 6 | 1.370 | 1.317 | 1.305 |
| 7 | 1.599 | 1.704 | 1.609 |
| 8 | 1.827 | 1.781 | 1.753 |
| 9 | 1.827 | 1.936 | 1.833 |
| 10 | 1.903 | 1.936 | 1.868 |
| 11 | 1.979 | 1.936 | 1.903 |
| 12 | 2.817 | 2.633 | 2.644 |
| 13 | 3.273 | 3.253 | 3.173 |
| 14 | 3.578 | 3.718 | 3.552 |
| 15 | 3.958 | 4.260 | 4.006 |
| 16 | 4.035 | 4.802 | 4.321 |
| 17 | 5.176 | 4.879 | 4.880 |
| 18 | 5.785 | 6.041 | 5.759 |
| 19 | 5.938 | 6.041 | 5.828 |
| 20 | 7.232 | 7.358 | 7.098 |
| 21 | 8.750 (17) | 8.400 (21) | 8.329 |
| 22 | 9.970 (27) | 9.430 (49) | 9.417 |
| 23 | 11.260 (37) | 13.610 (49) | 12.167 |
| 24 | 16.090 (75) | 13.810 (100) | 14.468 |
| 25 | 20.010 (75) | 18.900 (100) | 18.886 |
| 26 | 24.320 (100) | 23.450 (125) | 23.202 |
| 27 | 35.330 (175) | 32.230 (200) | 32.755 |
| 28 | 37.840 (150) | 35.180 (170) | 35.424 |
| 29 | 39.990 (145) | 35.460 (160) | 36.547 |
| 30 | 42.510 (125) | 38.460 (144) | 39.246 |
| Model | n Parameters | AICc | ∆AICc | Akaike Weight |
|---|---|---|---|---|
| 1 | 2 | 1492.43 | 627.00 | 0.00 |
| meanT | 3 | 1377.92 | 512.48 | 0.00 |
| SCL | 3 | 901.82 | 36.39 | 0.00 |
| SCL + meanT | 4 | 890.37 | 24.93 | 0.00 |
| SCL + meanT + SCL:meanT | 5 | 865.43 | 0.00 | 0.93 |
| SCL + SCL:meanT | 4 | 870.74 | 5.31 | 0.07 |
| maxT + SCL:meanT | 4 | 883.73 | 18.29 | 0.00 |
| SCL:meanT | 3 | 887.37 | 21.94 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Mérida, B.A.; Morales-Cabrera, A.; Chúa, C.; Girondot, M. Olive Ridley Sea Turtle Incubation in Natural Conditions Is Possible on Guatemalan Beaches. Sustainability 2023, 15, 14196. https://doi.org/10.3390/su151914196
Morales-Mérida BA, Morales-Cabrera A, Chúa C, Girondot M. Olive Ridley Sea Turtle Incubation in Natural Conditions Is Possible on Guatemalan Beaches. Sustainability. 2023; 15(19):14196. https://doi.org/10.3390/su151914196
Chicago/Turabian StyleMorales-Mérida, B. Alejandra, Alejandra Morales-Cabrera, Carlos Chúa, and Marc Girondot. 2023. "Olive Ridley Sea Turtle Incubation in Natural Conditions Is Possible on Guatemalan Beaches" Sustainability 15, no. 19: 14196. https://doi.org/10.3390/su151914196
APA StyleMorales-Mérida, B. A., Morales-Cabrera, A., Chúa, C., & Girondot, M. (2023). Olive Ridley Sea Turtle Incubation in Natural Conditions Is Possible on Guatemalan Beaches. Sustainability, 15(19), 14196. https://doi.org/10.3390/su151914196