
Behavior and Biochemical Mechanism of High Iron Attapulgite Dosages Affecting Sewage Sludge Composting

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Composting Materials and Trials
2.2. Analysis of Physiochemical Properties
2.3. DNA Extraction, High-Throughput Sequencing, and Data Analysis
3. Results and Discussion
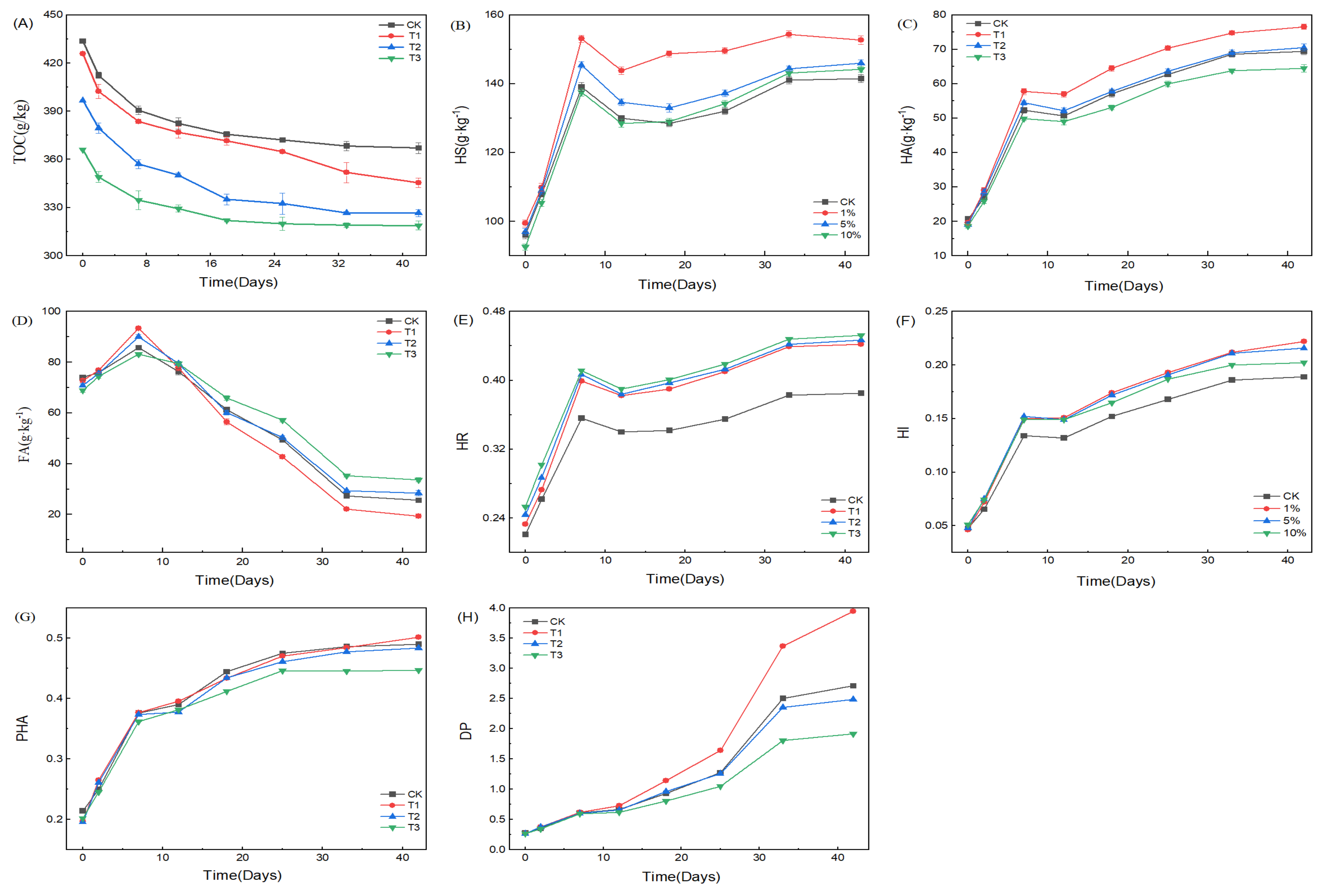
3.1. Effect of HIAP Dosage on SS Composting Humification
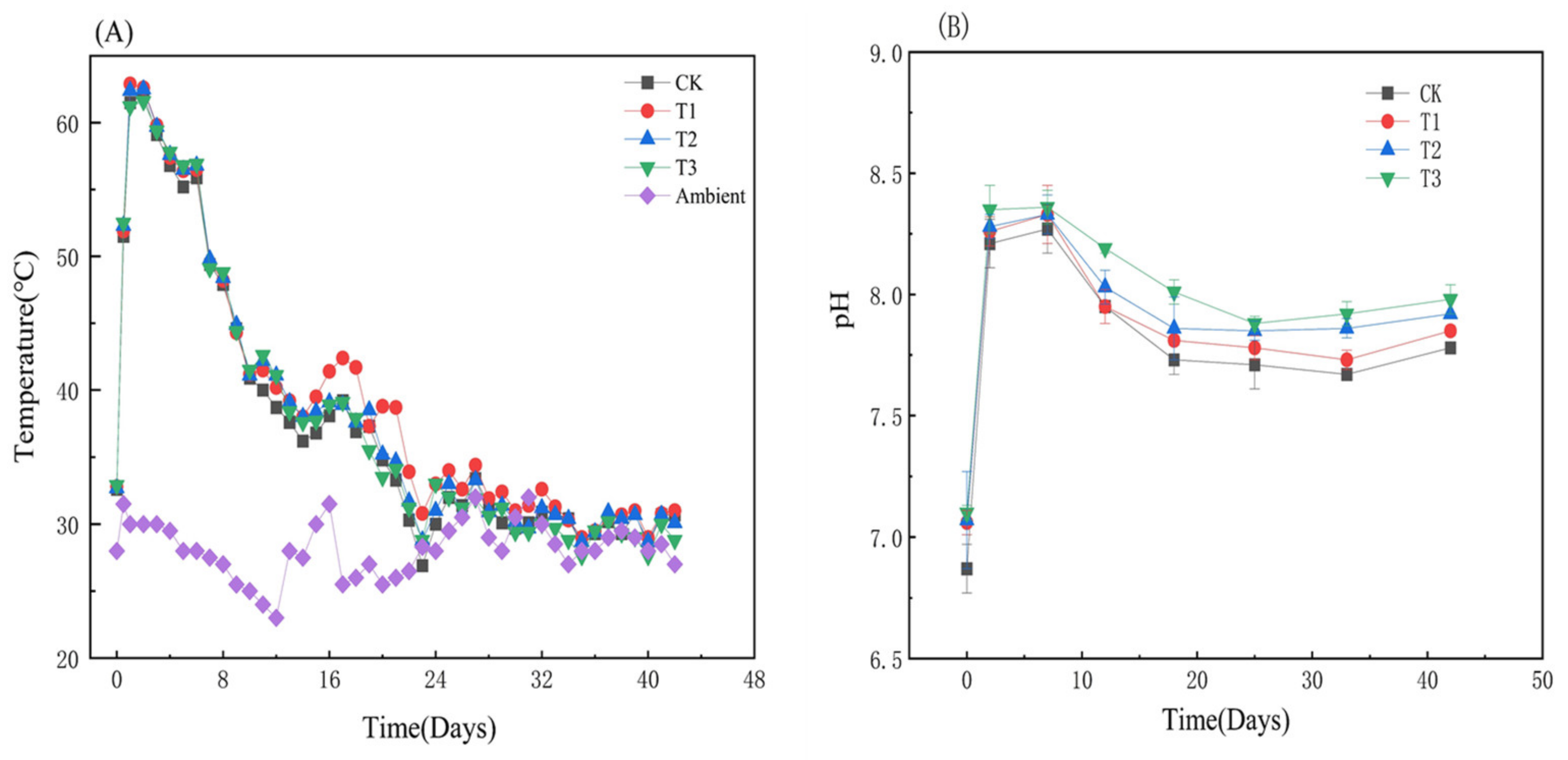
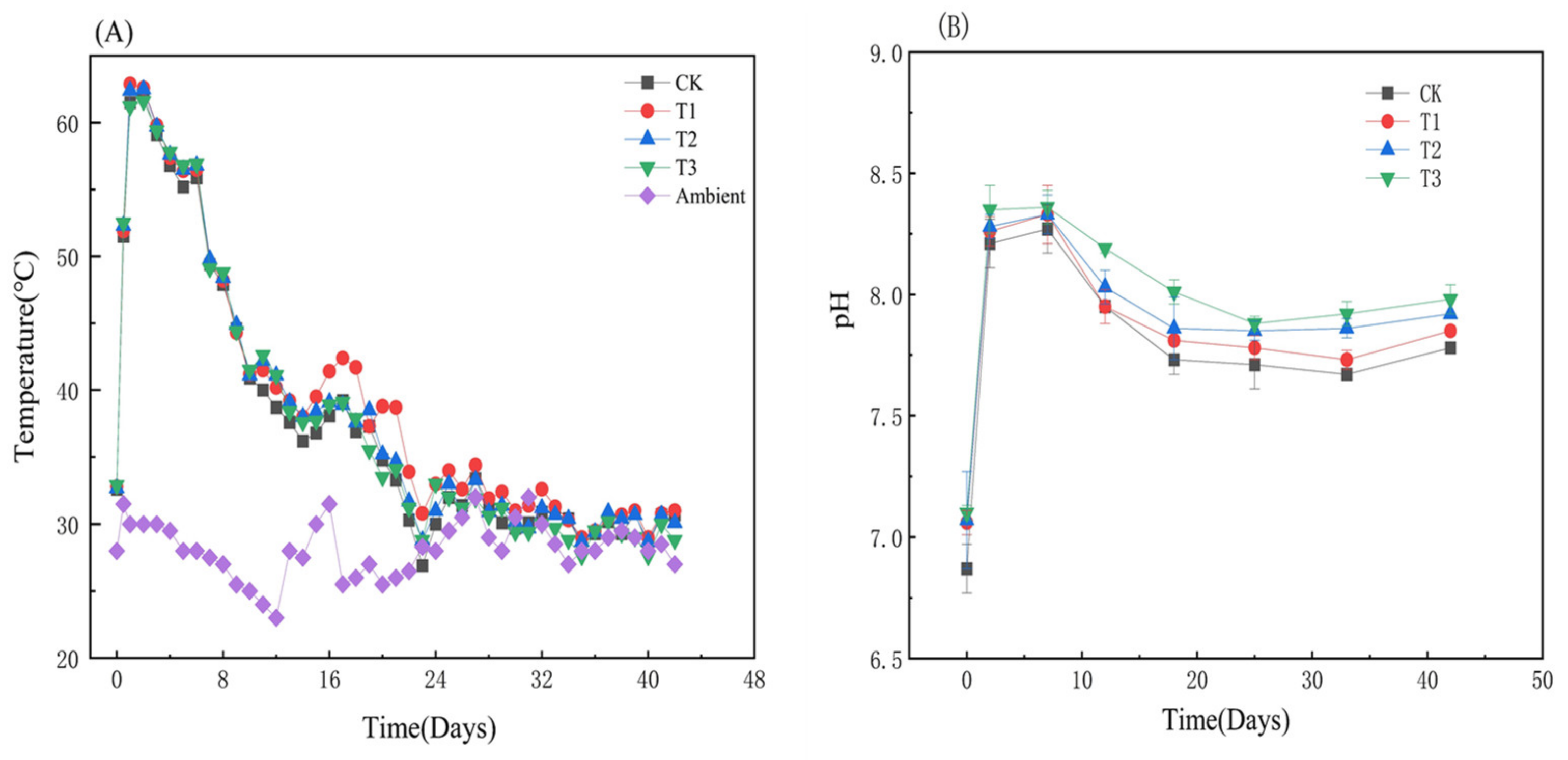
3.1.1. Variations of Temperature and pH during Composting
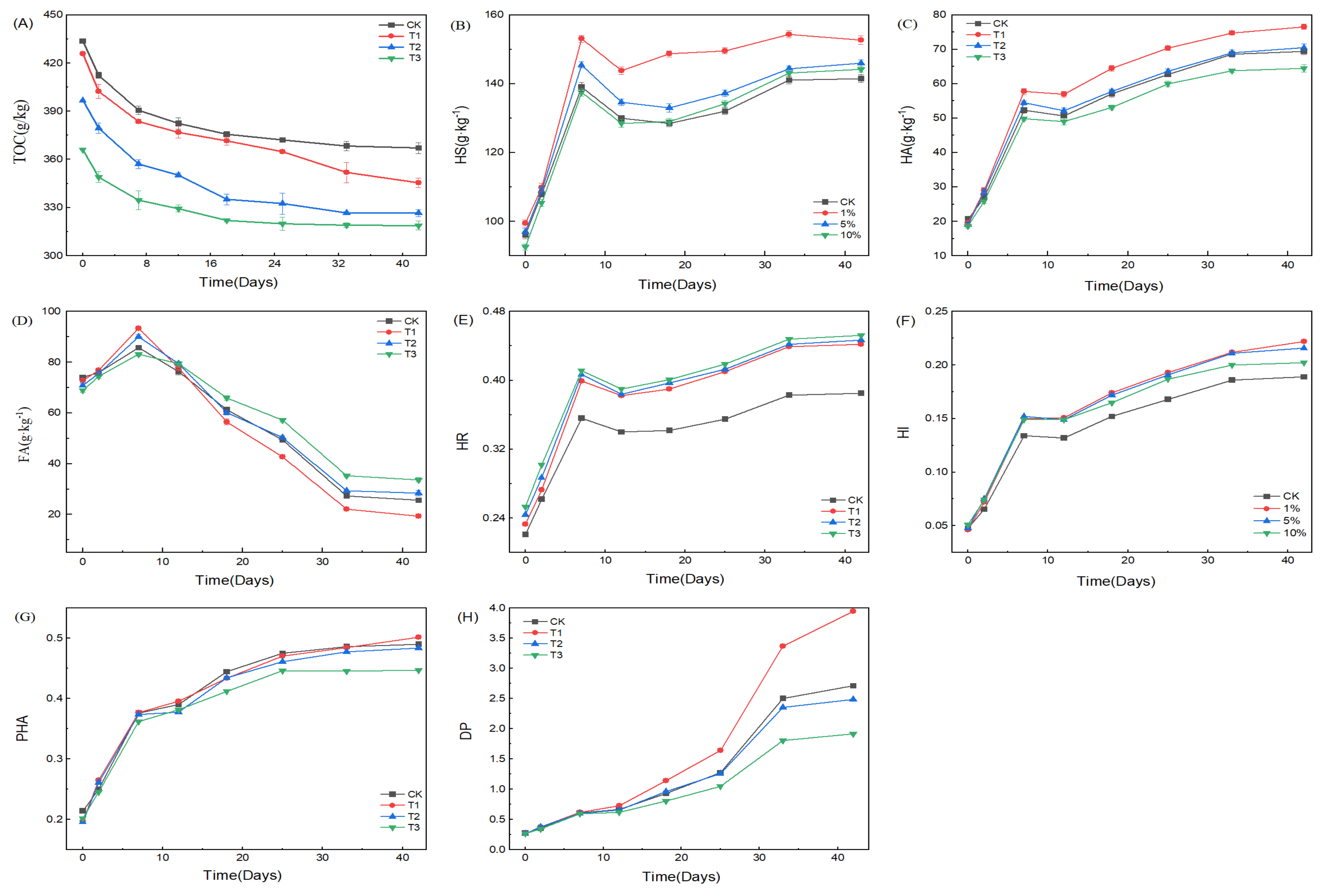
3.1.2. Variations of TOC, HS, HA, FA, and Humification Indexes with HIAP Dosage
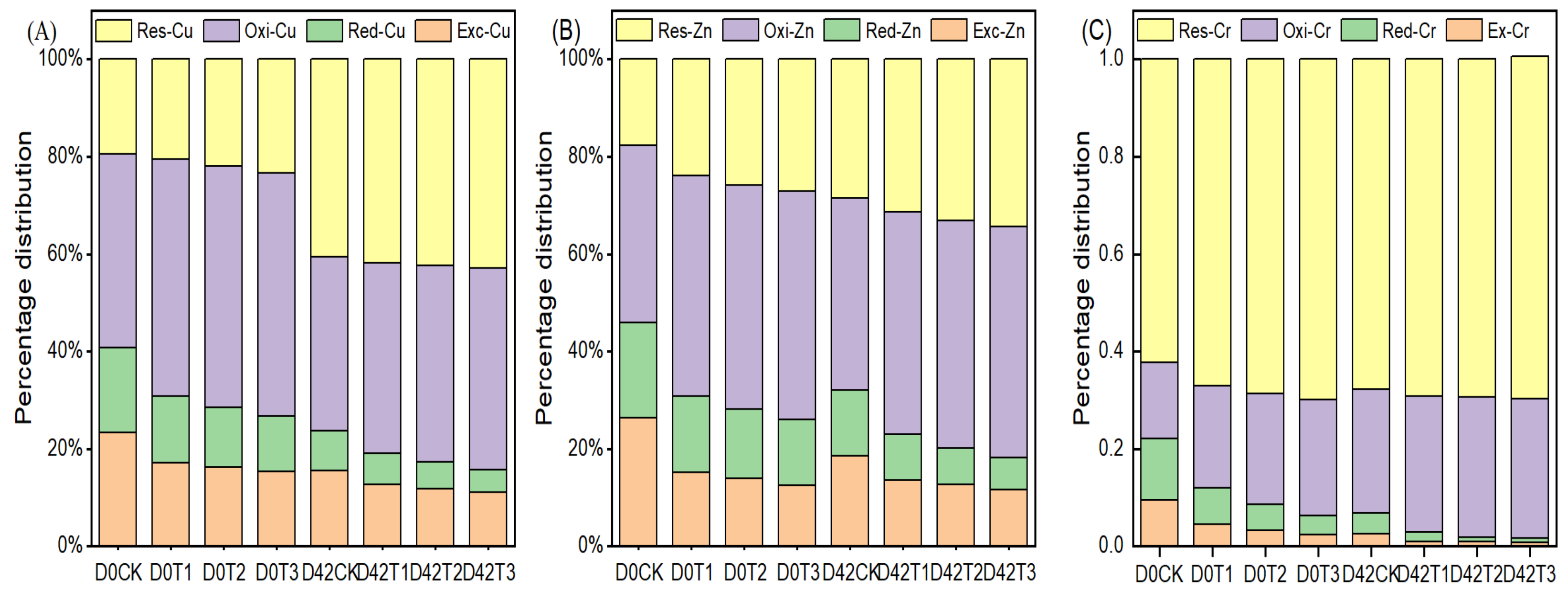
3.2. Effect of HIAP Dose on Passivating HMs during Composting Process
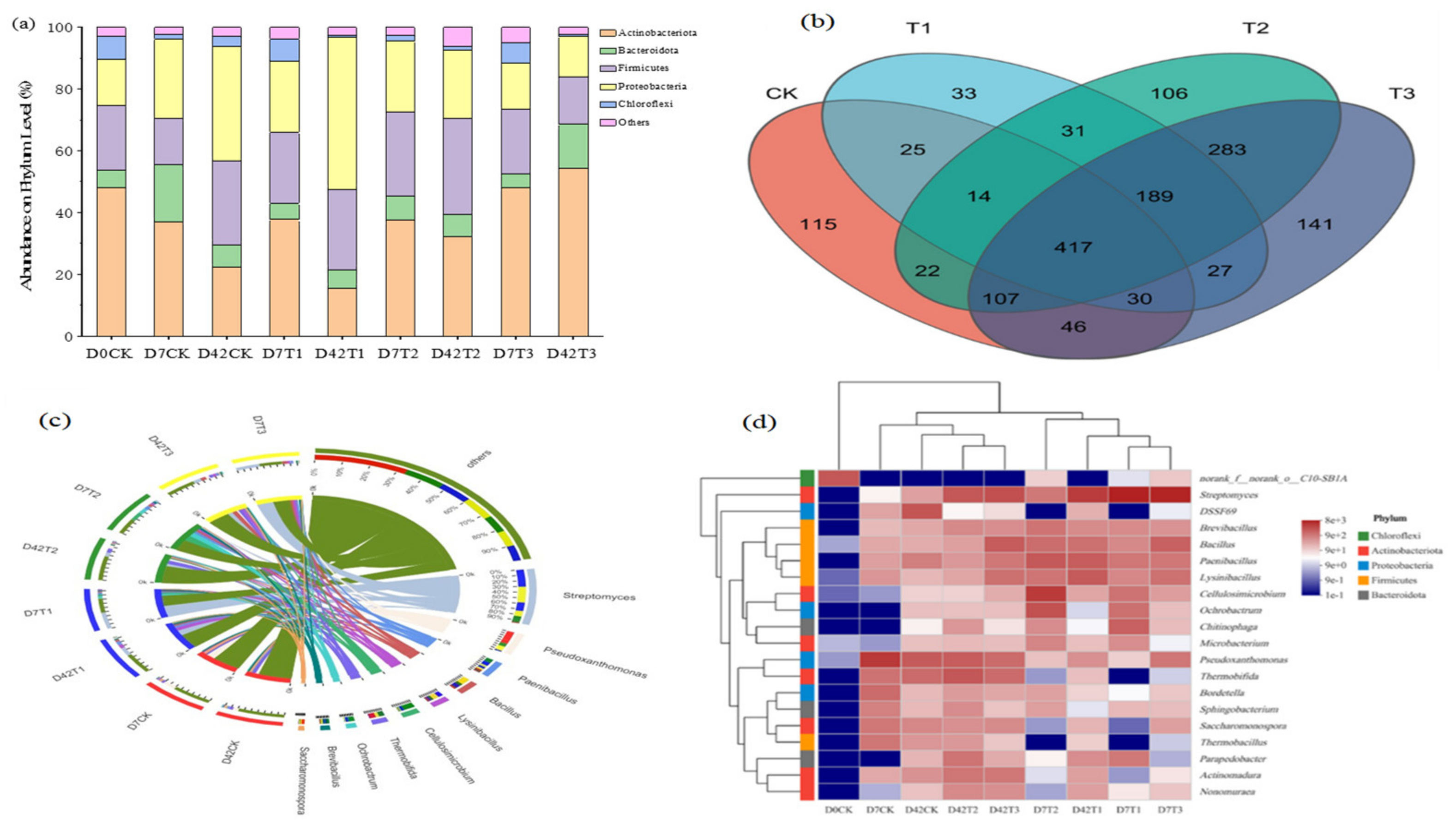
3.3. Succession of Microbial Community during Composting
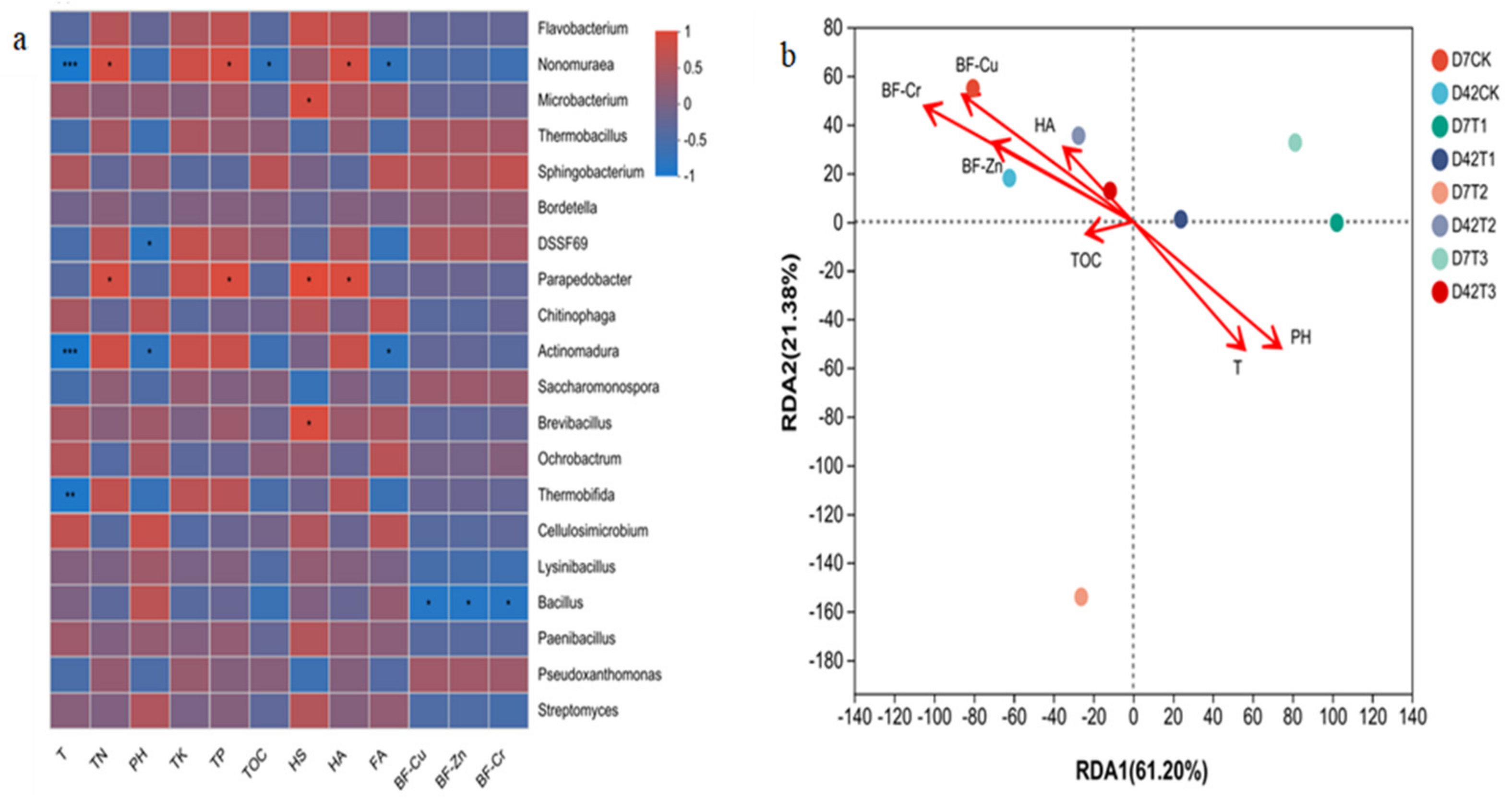
3.3.1. Effect of HIAP Dosage on Microbial Community
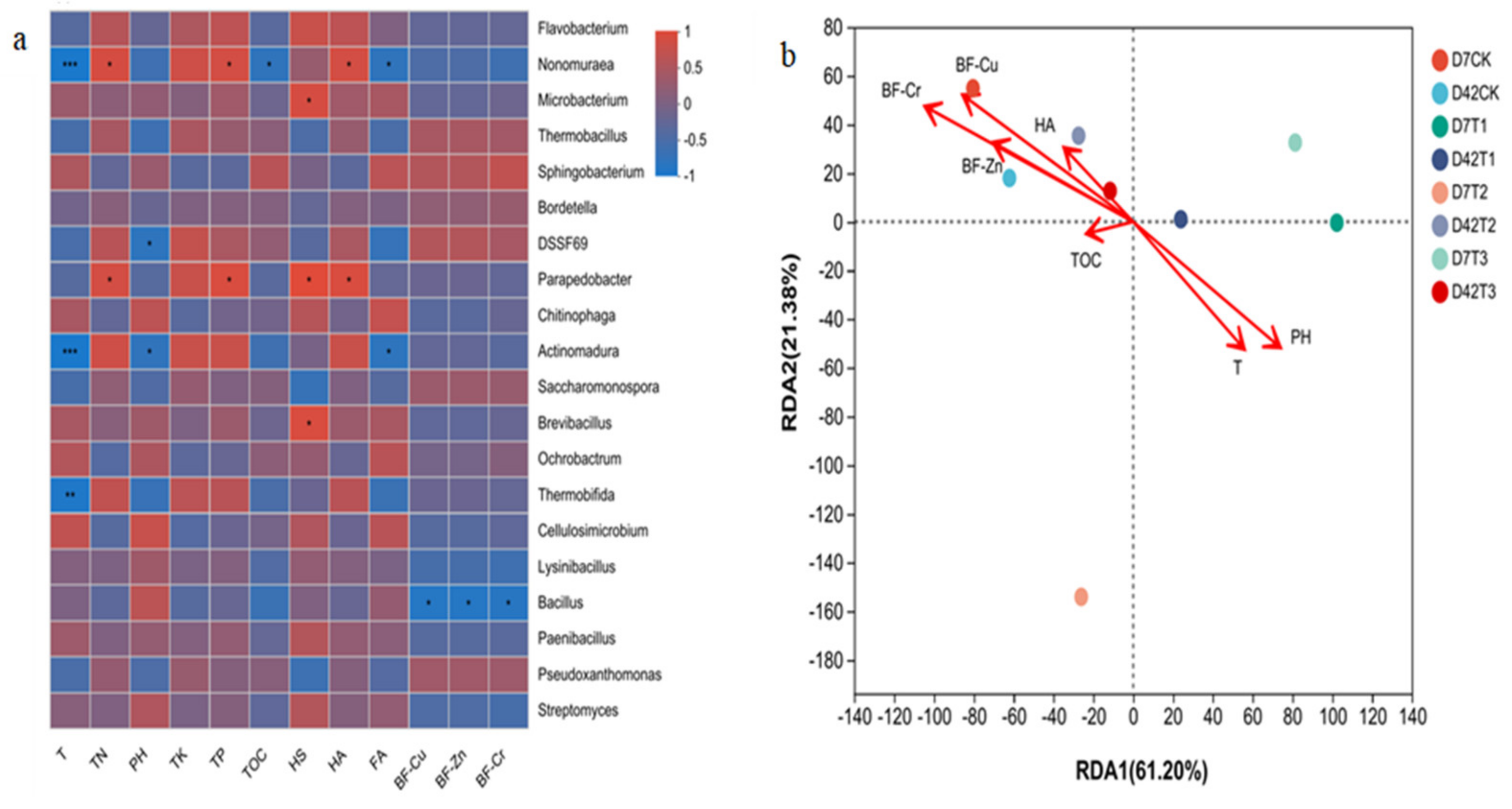
3.3.2. Relationship between Microbial Community and Environmental Factors
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Xiong, R.W.; Gao, X.F.; Tu, X.Y.; Mao, Y.L.; Jiang, L.; Zheng, L.; Du, Y.T. Heavy Metal Remediation in Sludge Compost: Recent Progress. J. Renew. Mater. 2022, 10, 469–486. [Google Scholar] [CrossRef]
- Li, H.; Zhang, T.; Shaheen, S.M.; Abdelrahman, H.; Ali, E.F.; Bolan, N.S.; Li, G.; Rinklebe, J. Microbial inoculants and struvite improved organic matter humification and stabilized phosphorus during swine manure composting: Multivariate and multiscale investigations. Bioresour. Technol. 2022, 351, 126976. [Google Scholar] [CrossRef] [PubMed]
- Borgulat, A.; Zgórska, A.; Głodniok, M. Comparison of different municipal sewage sludge products for potential ecotoxicity. Arch. Environ. Prot. 2022, 48, 92–99. [Google Scholar]
- Bruni, C.; Akyol, Ç.; Cipolletta, G.; Eusebi, A.L.; Caniani, D.; Masi, S.; Colón, J.; Fatone, F. Decentralized community composting: Past, present and future aspects of Italy. Sustainability 2020, 12, 3319. [Google Scholar] [CrossRef]
- Shan, G.; Li, W.; Gao, Y.; Tan, W.; Xi, B. Additives for reducing nitrogen loss during composting: A Review. J. Clean. Prod. 2021, 307, 127308. [Google Scholar] [CrossRef]
- Bui, V.K.H.; Truong, H.B.; Hong, S.J.; Li, X.W.; Hur, J. Biotic and abiotic catalysts for enhanced humification in composting: A comprehensive review. J. Clean. Prod. 2023, 402, 136832. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, S.L.; Ning, X.; Yang, M.; Liu, M.B.; Zang, F.; Nan, Z.R. A promising amendment for the immobilization of heavy metal(loid)s in agricultural soil, northwest China. J. Soil. Sediment. 2021, 21, 2273–2286. [Google Scholar] [CrossRef]
- Ren, J.; Dai, L.; Tao, L. Stabilization of heavy metals in sewage sludge by attapulgite. J. Air Waste Manag. Assoc. 2021, 71, 392–399. [Google Scholar] [CrossRef]
- Liu, S.Y.; Liu, J.; Zhao, J.J.; Xia, D.S.; Pan, F.; Liu, C.; George, Z.K.; Fu, J. Palygorskite changes heavy metal bioavailability and microbial functional diversity in sewage sludge composting. Environ. Technol. 2015, 36, 2855–2862. [Google Scholar] [CrossRef]
- Lin, H.; Sun, W.C.; Yu, Y.J.; Ding, Y.Z.; Yang, Y.Y.; Zhang, Z.L.; Ma, J.W. Simultaneous reductions in antibiotics and heavy metal pollution during manure composting. Sci. Total Environ. 2021, 788, 147830. [Google Scholar] [CrossRef]
- Chen, Y.N.; Tang, P.; Li, Y.P.; Chen, L.; Jiang, H.J.; Liu, Y.H.; Luo, X.L. Effect of attapulgite on heavy metals passivation and microbial community during co-composting of river sediment with agricultural wastes. Chemosphere 2022, 299, 134347. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.Z.; Jia, X.S.; Xu, P.Z.; Huang, X.; Gu, W.J.; Zhang, F.B.; Yang, S.H.; Tang, S.H. The addition of modified attapulgite reduces the emission of nitrous oxide and ammonia from aerobically composted chicken manure. J. Air Waste Manag. 2012, 62, 1174–1181. [Google Scholar] [CrossRef]
- Pan, J.T.; Li, R.H.; Zhai, L.M.; Zhang, Z.Q.; Ma, J.Y.; Liu, H.B. Influence of palygorskite addition on biosolids composting process enhancement. J. Clean. Prod. 2019, 217, 371–379. [Google Scholar] [CrossRef]
- Zhang, W.M.; Yu, C.X.; Wang, X.J.; Yin, S.Q.; Chang, X.Y. Additives improved saprotrophic fungi for the formation of humic acids in chicken manure and corn stover mix composting. Bioresour. Technol. 2022, 346, 126626. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.G.; Yang, W.J.; Lu, T.Y.; Ru, X.; Dai, Z.B.; Meshack, A.O.; Hou, J.X. Heterogeneous Fenton for Removal Rhodamine B by Iron bearing Attapulgite Granular Catalysts. Water Air Soil Pollut. 2023, 234, 61. [Google Scholar] [CrossRef]
- Sun, H.Y.; Xing, R.Z.; Ye, X.Y.; Yin, K.; Zhang, Y.; Chen, Z.; Zhou, S. Reactive oxygen species accelerate humification process during iron mineral-amended sludge composting. Bioresour. Technol. 2023, 370, 128544. [Google Scholar] [CrossRef]
- Nemati, K.; Bakar, N.K.A.; Abas, M.R.; Sobhanzadeh, E. Speciation of heavy metals by modified BCR sequential extraction procedure in different depths of sediments from Sungai Buloh, Selangor, Malaysia. J. Hazard Mater. 2011, 192, 402–410. [Google Scholar] [CrossRef]
- Tmecc, T.W.; Leege, P.; Millner, P.; Watson, M.E. Test Methods for the Examination of Composts and Composting; The US Composting Council: Raleigh, NC, USA; US Government Printing Office: Washington, DC, USA, 2003. [Google Scholar]
- Pan, C.N.; Zhao, Y.; Zhao, L.; Wu, J.Q.; Zhang, X.; Xie, X.Y.; Kang, K.J.; Jia, L.M. Modified montmorillonite and illite adjusted the preference of biotic and abiotic pathways of humus formation during chicken manure composting. Bioresour. Technol. 2021, 319, 124121. [Google Scholar] [CrossRef]
- Huang, Y.; Dan, Y.L.; Shah, G.M.; Chen, W.; Wang, W.; Xu, Y.D.; Huang, H.Y. Hyperthermophilic pretreatment composting significantly accelerates humic substance formation by regulating precursor production and microbial communities. Waste Manag. 2019, 92, 89–96. [Google Scholar] [CrossRef]
- Wu, X.Y.; Wang, J.S.; Yu, Z.J.; Charles, A.; Shen, L.; Wu, X.L.; Li, J.K.; Yu, R.L.; Liu, Y.D.; Zeng, W.M. Impact of bamboo sphere amendment on composting performance and microbial community succession in food waste composting. J. Environ. Manag. 2022, 303, 114144. [Google Scholar] [CrossRef]
- Yang, P.P.; Yin, H.; Peng, H.; Tang, S.Y.; Lu, M.; Liu, H. Effects of exogenous microorganism inoculation on efficiency and bacterial community structure of sludge composting. HuanjingKexue 2017, 38, 3536–3543. [Google Scholar]
- Liu, H.; Yin, H.; Tang, S.Y.; Wei, K.; Peng, H.; Lu, G.N.; Dang, Z. Effects of benzo [a] pyrene (BaP) on the composting and microbial community of sewage sludge. Chemosphere 2019, 222, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Bernal, M.P.; Alburquerque, J.A.; Moral, R. Composting of animal manures and chemical criteria for compost maturity assessment- A review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Zhai, W.; Du, Y.; Zhao, Y.; Wei, Z.; Wu, J.; Xie, X.; Yang, H.; Wu, D.; Guo, T. Core bacterial community driven the conversion of fulvic acid components during composting with adding manganese dioxide. Bioresour. Technol. 2021, 337, 125495. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Z.; Jiang, Y.; Li, R.; Ren, X.N.; Zhao, J.; Shen, F.; Wang, M.; Zhang, Z. Evaluation of medical stone amendment for the reduction of nitrogen loss and bioavailability of heavy metals during pig manure composting. Bioresour. Technol. 2016, 220, 297–304. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, X.Y. Addition of seaweed and bentonite accelerates the two-stage composting of green waste. Bioresour. Technol. 2017, 243, 154–162. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Awasthi, S.K.; Wang, Q.; Awasthi, M.K.; Zhao, J.; Chen, H.; Ren, X.; Wang, M.; Zhang, Z. Role of Ca-bentonite to improve the humification, enzymatic activities, nutrient transformation and end product quality during sewage sludge composting. Bioresour. Technol. 2018, 262, 80–89. [Google Scholar] [CrossRef]
- Liu, L.; Ye, Q.; Wu, Q.; Liu, T.; Peng, S. Effect of biochar addition on sludge aerobic composting and greenbelt utilization. Environ. Technol. Innov. 2021, 21, 101279. [Google Scholar] [CrossRef]
- Akyol, C.; Ince, O.; Ince, B. Crop-based composting of lignocellulosic digesates: Focus on bacterial and fungal diversity. Bioresour. Technol. 2019, 288, 121549. [Google Scholar] [CrossRef]
- Qi, H.; Wei, Z.; Zhang, J.; Zhao, Y.; Wu, J.; Gao, X.; Liu, Z.; Li, Y. Effect of MnO2 on biotic and abiotic pathways of humic-like substance formation during composting of different raw materials. Waste Manag. 2019, 87, 326–334. [Google Scholar] [CrossRef]
- Ren, X.; Wang, Q.; Li, R.; Chang, C.C.; Pan, J.; Zhang, Z. Effect of clay on greenhouse gas emissions and humification during pig manure composting as supported by spectroscopic evidence. Sci. Total Environ. 2020, 737, 139712. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Zhang, Z.H.; Zhang, D.W.; Tian, Y.; Nan, J.; Feng, Y.J. Hydrothermal pretreatment and compound microbial agents promoting high-quality kitchen waste compost: Superior humification degree and reduction of odor. Sci. Total Environ. 2023, 862, 160657. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Li, R.; Wu, S.; He, Q.; Ling, Z.; Liu, T.; Wang, Q.; Zhang, Z.Q.; Quan, F.S. Cattle manure compost humification process by inoculation ammonia-oxidizing bacteria. Bioresour. Technol. 2022, 344, 126314. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Yue, D.; Fukushima, M.; Fukuchi, S.; Nishimoto, R.; Nie, Y. Enhanced humification by carbonated basic oxygen furnace steel slag–II. Process characterization and the role of inorganic components in the formation of humic-like substances. Bioresour. Technol. 2012, 114, 637–643. [Google Scholar] [CrossRef]
- Wang, Q.H.; Hao, K.H.; Chelsea, B.; Kou, Y.; An, Z.X.; Mohamed, G.E.D.; Chen, C.M. The role and potential of attapulgite in catalytic pyrolysis of refinery waste activated sludge. Petrol. Sci. 2022, 19, 354–362. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, Y.; Zhao, W.; Yang, T.; Zhang, X.; Xie, X.; Cui, H.; Wei, Z. Effect of precursors combined with bacteria communities on the formation of humic substances during different materials composting. Bioresour. Technol. 2017, 226, 191–199. [Google Scholar] [CrossRef]
- Miura, A.; Okabe, R.; Izumo, K.; Fukushima, M. Influence of the physicochemical properties of clay minerals on the degree of darkening via polycondensation reactions between catechol and glycine. Appl. Clay Sci. 2009, 46, 277–282. [Google Scholar] [CrossRef]
- Chen, H.F.; Koopal, L.K.; Xiong, J.; Avena, M.; Tan, W.F. Mechanisms of soil humic acid adsorption onto montmorillonite and kaolinite. J. Colloid Interface Sci. 2017, 504, 457–467. [Google Scholar] [CrossRef]
- Zhang, L.C.; Luo, L.; Zhang, S.Z. Integrated investigations on the adsorption mechanisms of fulvic and humic acids on three clay minerals. Colloid Surf. A 2012, 406, 84–90. [Google Scholar] [CrossRef]
- Jindo, K.; Sonoki, T.; Matsumoto, K.; Canellas, L.; Roig, A.; Sanchez-Monedero, M.A. Influence of biochar addition on the humic substances of composting manures. Waste Manag. 2016, 49, 545–552. [Google Scholar] [CrossRef]
- Cui, H.; Ou, Y.; Wang, L.X.; Yan, B.X.; Li, Y.X.; Bao, M. Critical passivation mechanisms on heavy metals during aerobic composting with different grain-size zeolites. J. Hazard Mater. 2021, 406, 124313. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, L.; Zhan, J.; Guo, X.S. Variation and factors on heavy metal speciation during co-composting of rural sewage sludge and typical rural organic solid waste. J. Environ. Manag. 2022, 306, 114418. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, G.; Castellano-Hinojosa, A.; Correa-Galeote, D.; Bedmar, E.J. Evolution of bacterial diversity during two-phase olive mill waste ("alperujo") composting by 16S rRNA gene pyrosequencing. Bioresour. Technol. 2017, 224, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.Z.; Li, X.X.; Zeng, Y.; Wang, S.P.; Sun, Z.Y.; Tang, Y.Q. Dynamic change of bacterial community during dairy manure composting process revealed by high-throughput sequencing and advanced bioinformatics tools. Bioresour. Technol. 2020, 306, 123091. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.P.; Chu, S.; Wang, B.; Zhang, D.; Zhou, P. Dynamic bacterial assembly driven by Streptomyces griseorubens JSD-1 inoculants correspond to composting performance in swine manure and rice straw co-composting. Bioresour. Technol. 2020, 313, 123692. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.P.; Li, M.X.; Song, L.Y.; Wang, C.; Yang, S.; Yan, Z.Y.; Wang, Y.Q. Study on a nitrogen-retaining microbial agent to reduce nitrogen loss during chicken manure composting and nitrogen transformation mechanism. J. Clean. Prod. 2021, 285, 124813. [Google Scholar] [CrossRef]
- Song, Y.J.; Li, R.Y.; Wang, Y.X.; Hou, Y.; Chen, G.Y.; Yan, B.B.; Cheng, Z.J.; Mu, L. Co-composting of cattle manure and wheat straw covered with a semipermeable membrane: Organic matter humification and bacterial community succession. Environ. Sci. Pollut. Res. 2023, 30, 32776–32789. [Google Scholar] [CrossRef]
- Rastogi, G.; Bhalla, A.; Adhikari, A.; Bischoff, K.M.; Hughes, S.R.; Christopher, L.P.; Sani, R.K. Characterization of thermostable cellulases produced by Bacillus and Geobacillus strains. Bioresour. Technol. 2010, 101, 8798–8806. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Wang, B.; Jiang, X.; Zeng, W.; Yu, R.; Wu, X.; Shen, L.; Wu, X.; Li, J.; Liu, Y. Behavior and Biochemical Mechanism of High Iron Attapulgite Dosages Affecting Sewage Sludge Composting. Sustainability 2023, 15, 12527. https://doi.org/10.3390/su151612527
Yu Z, Wang B, Jiang X, Zeng W, Yu R, Wu X, Shen L, Wu X, Li J, Liu Y. Behavior and Biochemical Mechanism of High Iron Attapulgite Dosages Affecting Sewage Sludge Composting. Sustainability. 2023; 15(16):12527. https://doi.org/10.3390/su151612527
Chicago/Turabian StyleYu, Zhaojing, Bin Wang, Xiaoya Jiang, Weimin Zeng, Runlan Yu, Xiaoyan Wu, Li Shen, Xueling Wu, Jiaokun Li, and Yuandong Liu. 2023. "Behavior and Biochemical Mechanism of High Iron Attapulgite Dosages Affecting Sewage Sludge Composting" Sustainability 15, no. 16: 12527. https://doi.org/10.3390/su151612527
APA StyleYu, Z., Wang, B., Jiang, X., Zeng, W., Yu, R., Wu, X., Shen, L., Wu, X., Li, J., & Liu, Y. (2023). Behavior and Biochemical Mechanism of High Iron Attapulgite Dosages Affecting Sewage Sludge Composting. Sustainability, 15(16), 12527. https://doi.org/10.3390/su151612527