
Retreat of Major European Tree Species Distribution under Climate Change—Minor Natives to the Rescue?


Abstract
:1. Introduction
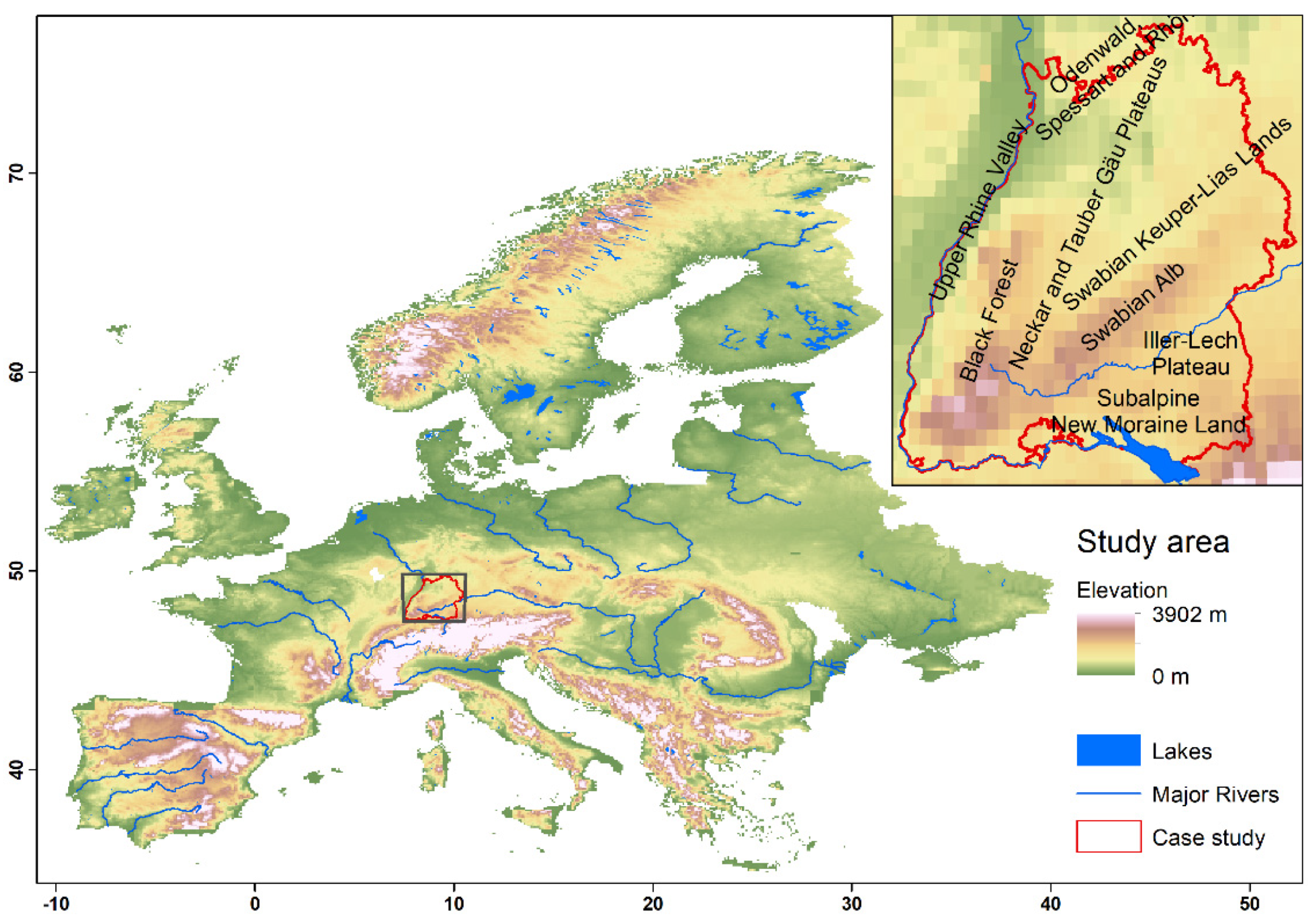
2. Materials and Methods
2.1. Analyzed Tree Species
2.2. Species Distribution Modelling
2.2.1. Species Data
2.2.2. Explanatory Variables
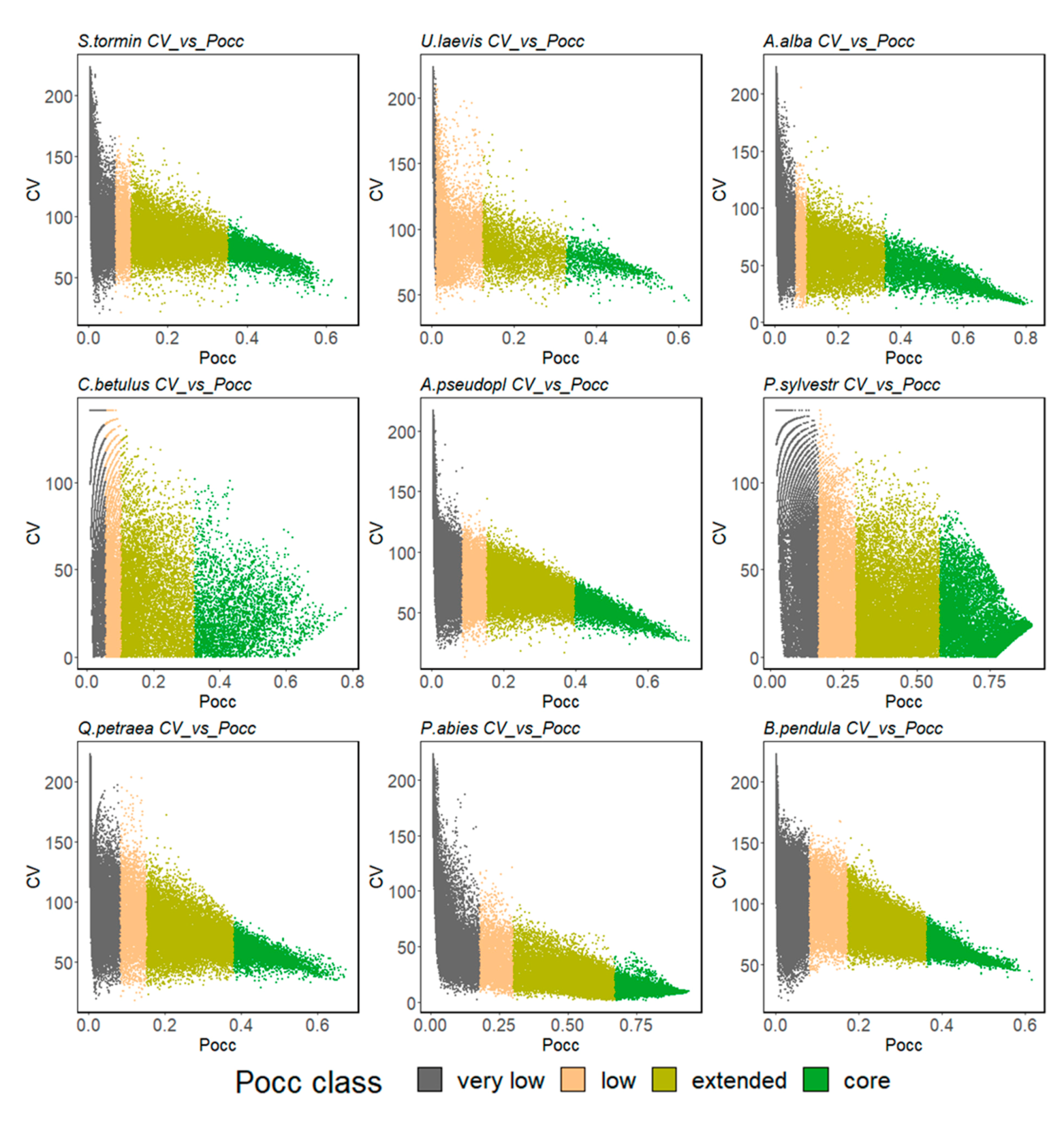
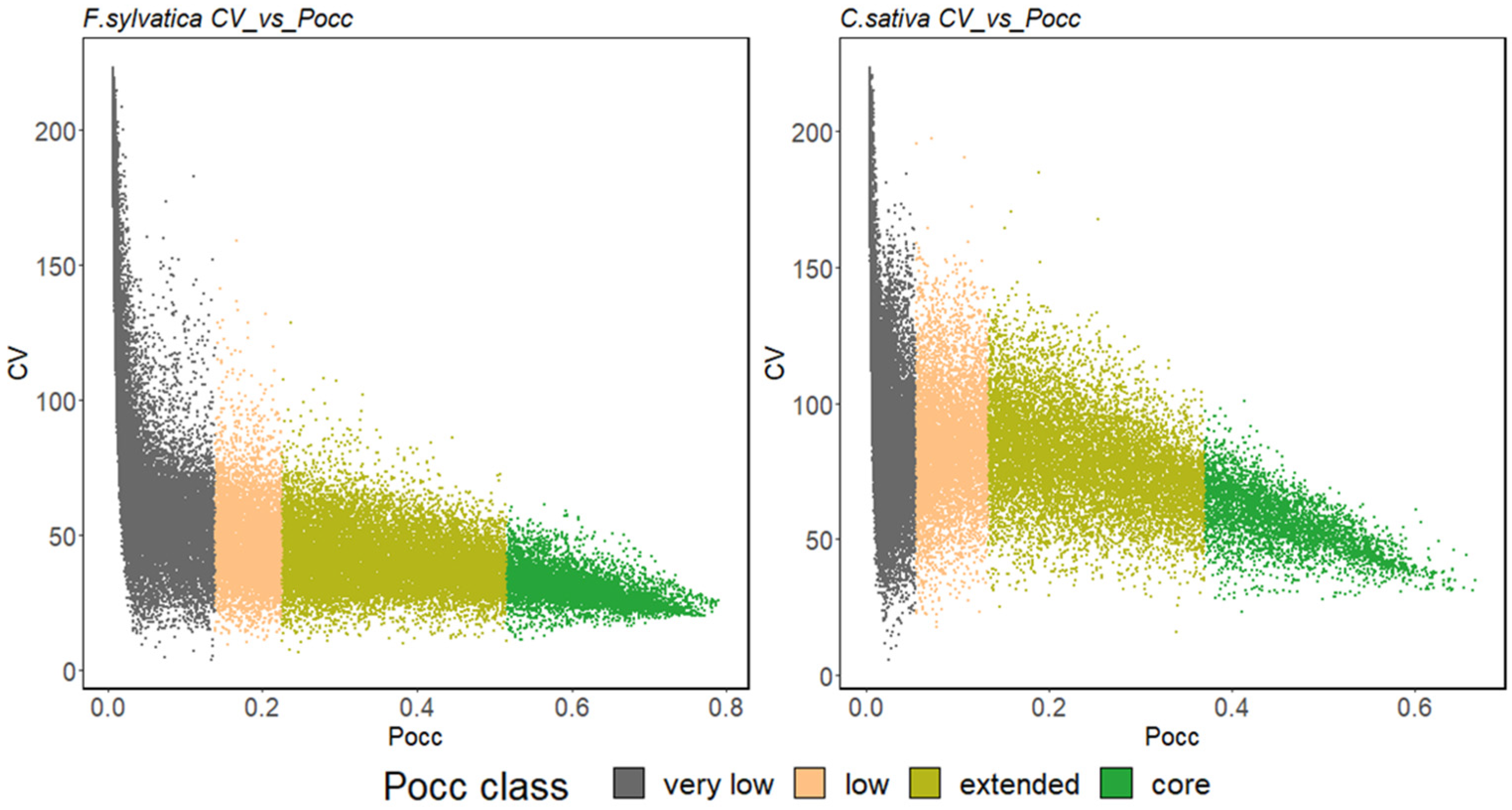
2.2.3. Ensemble Species Distribution Modelling
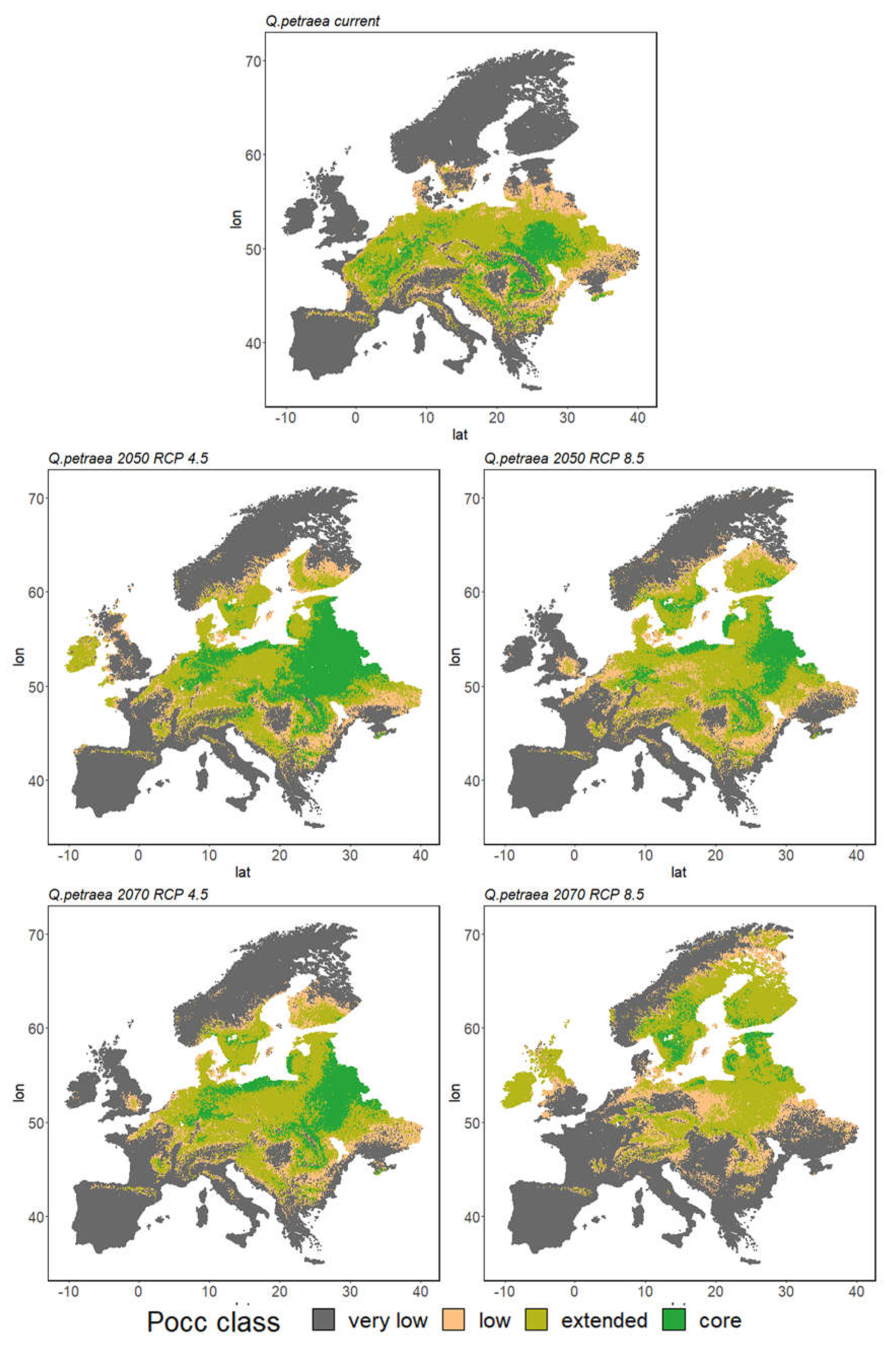
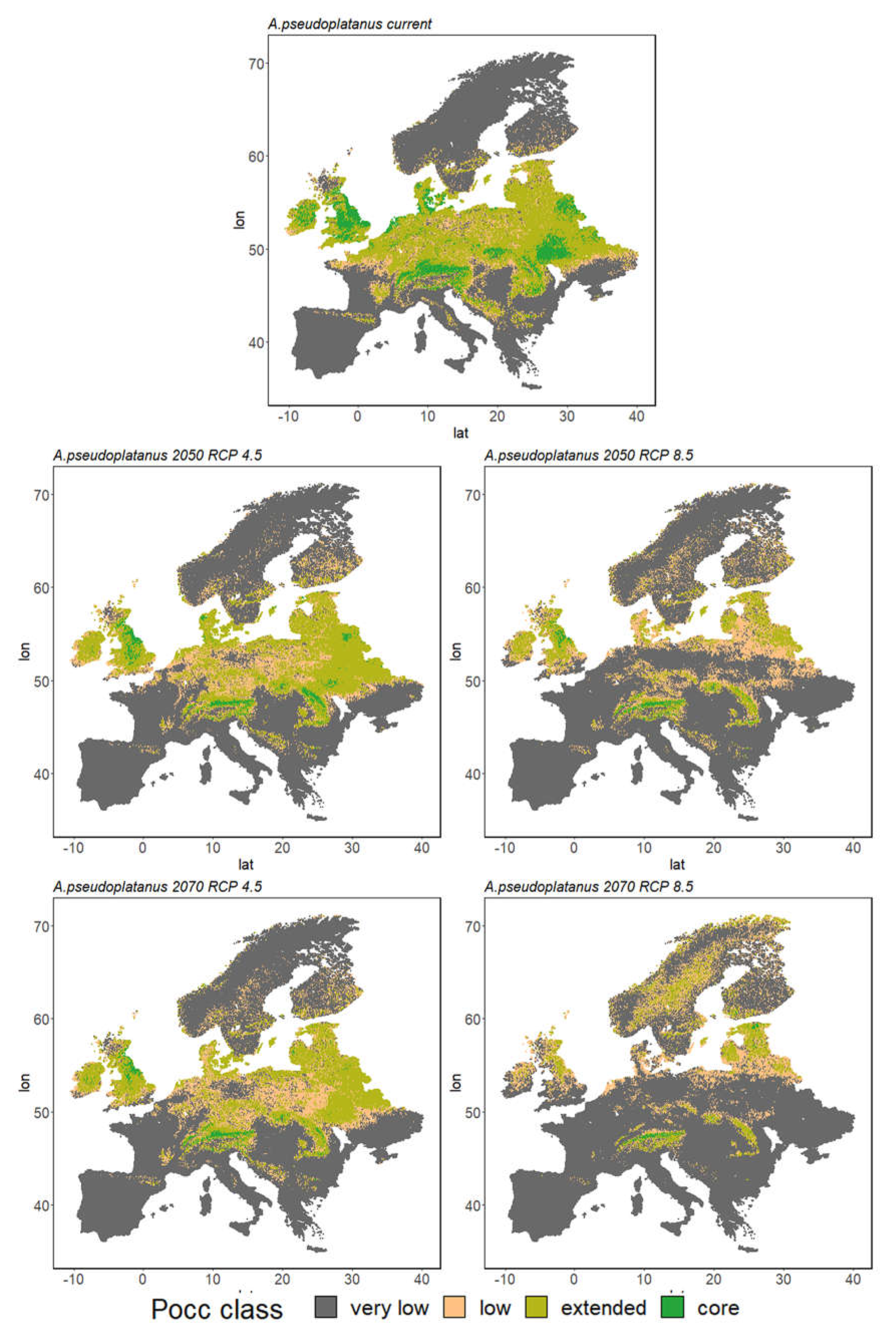
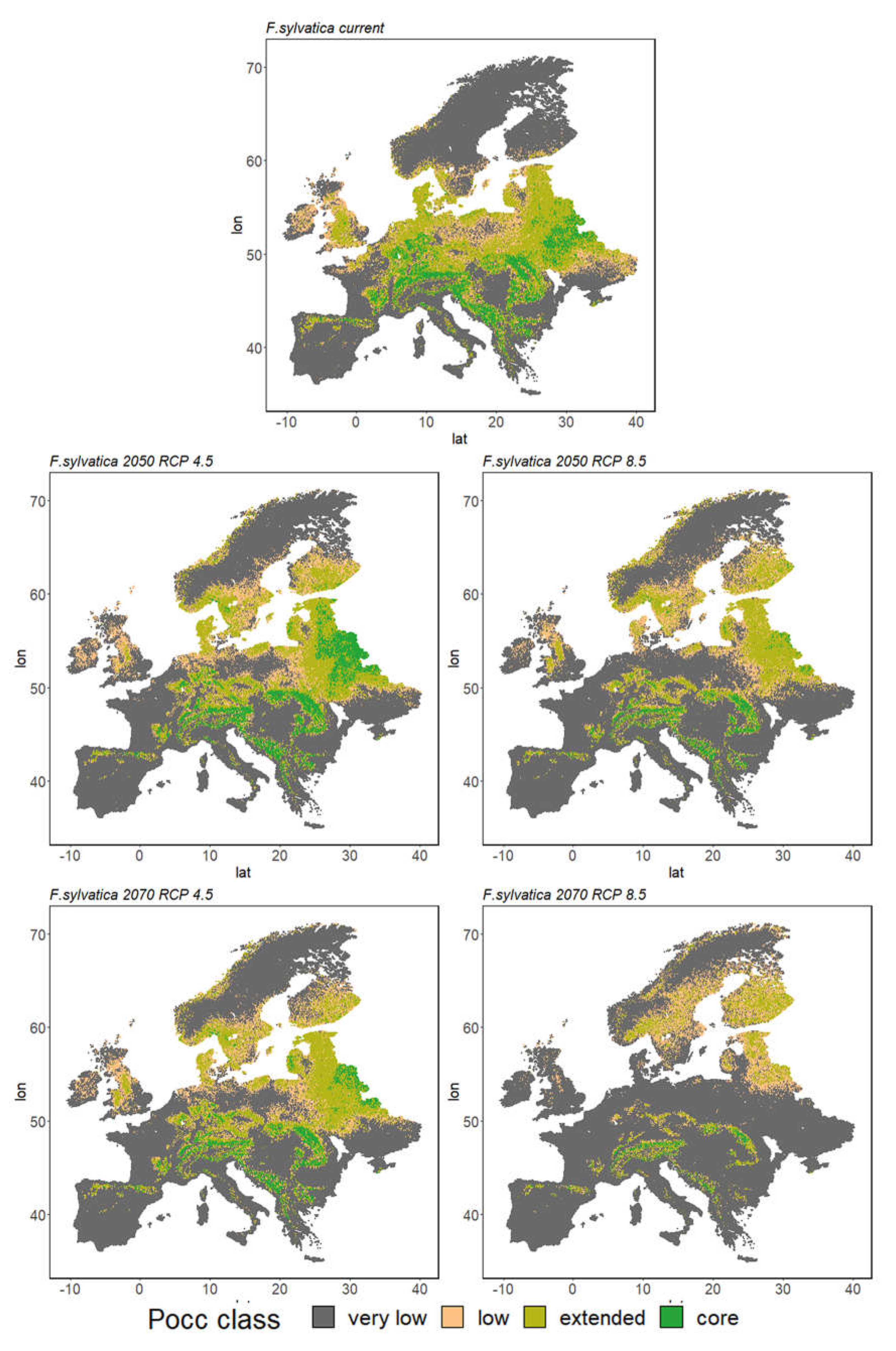
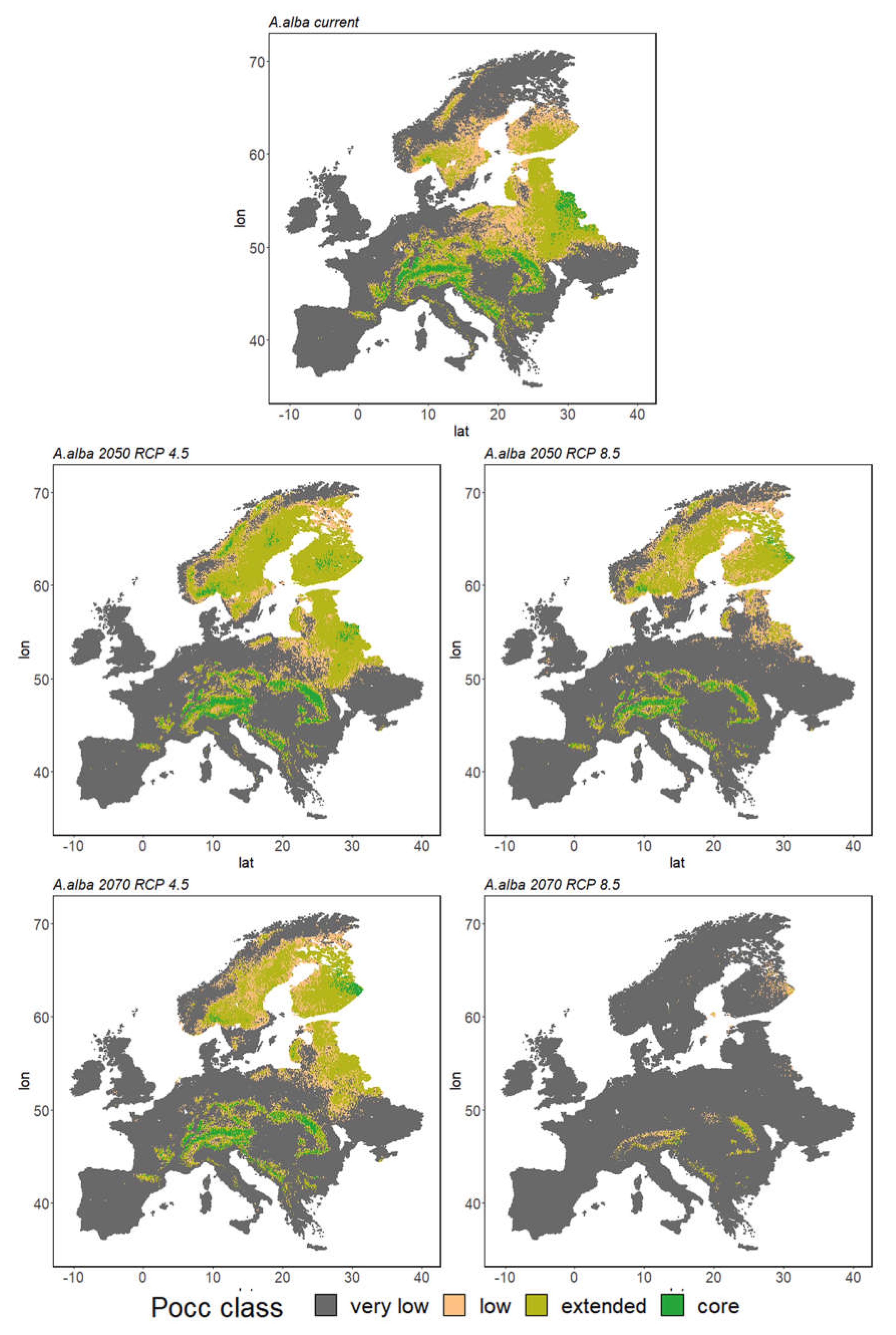
3. Results
3.1. Model Performance
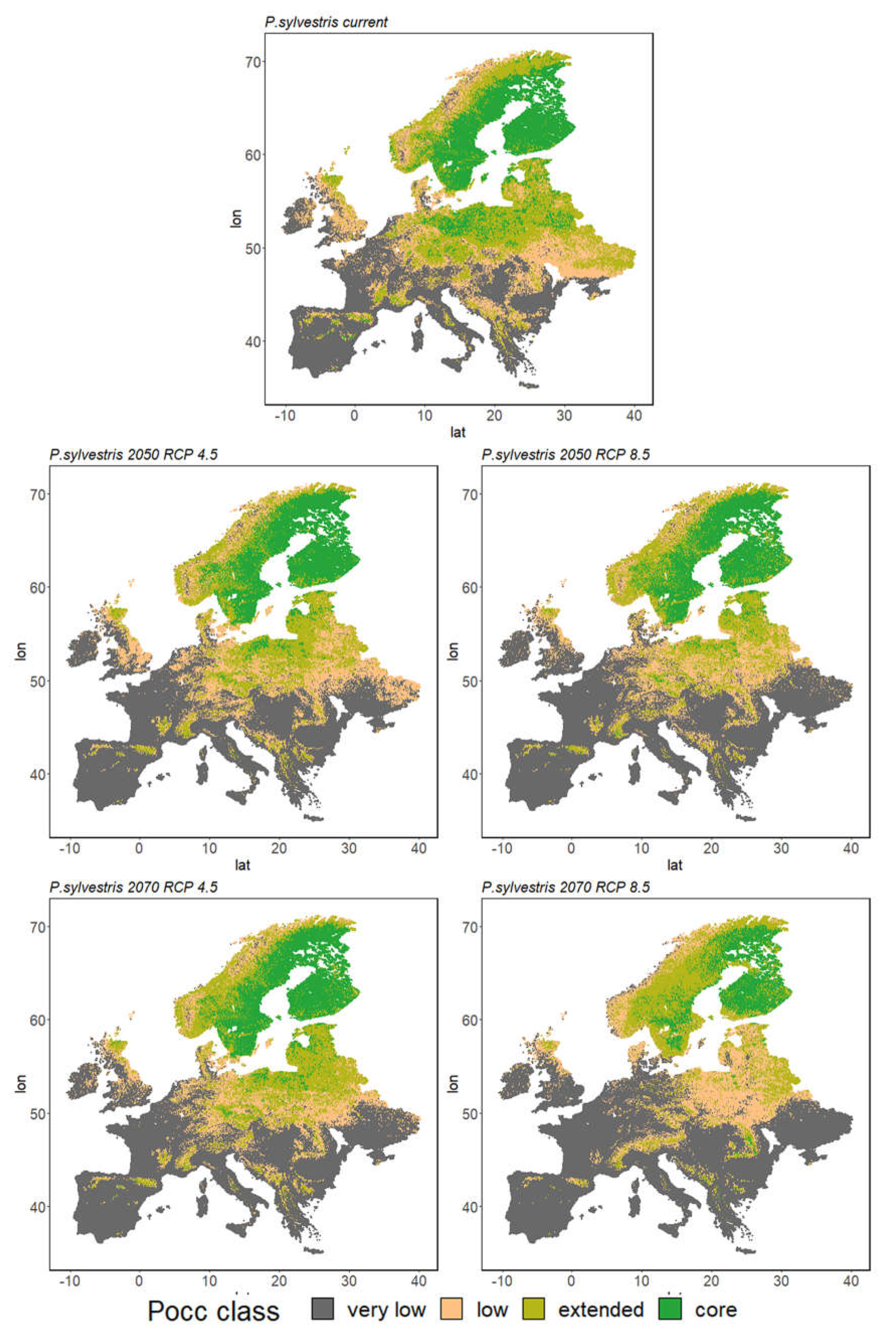
3.2. Ensemble Predictions
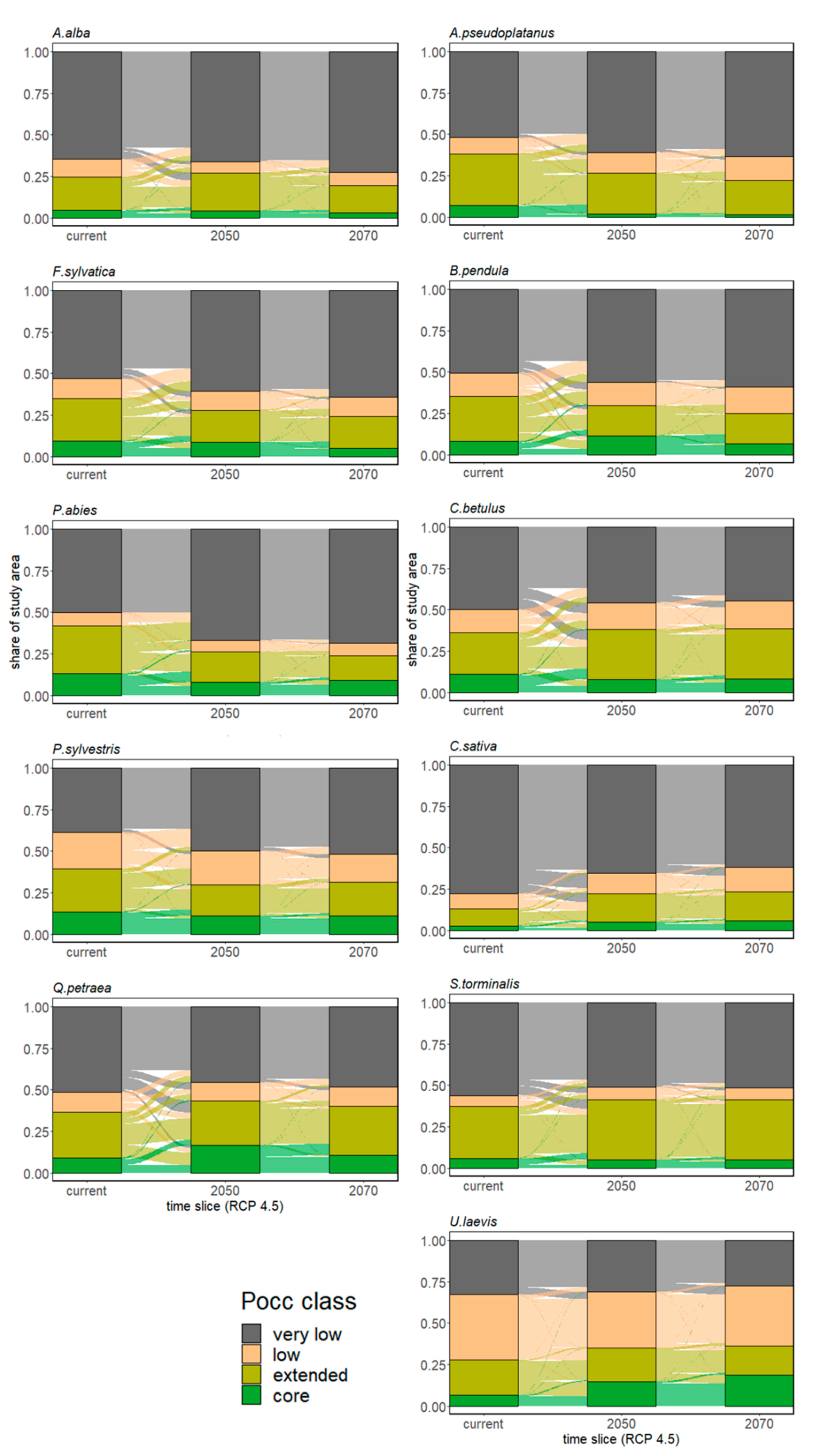
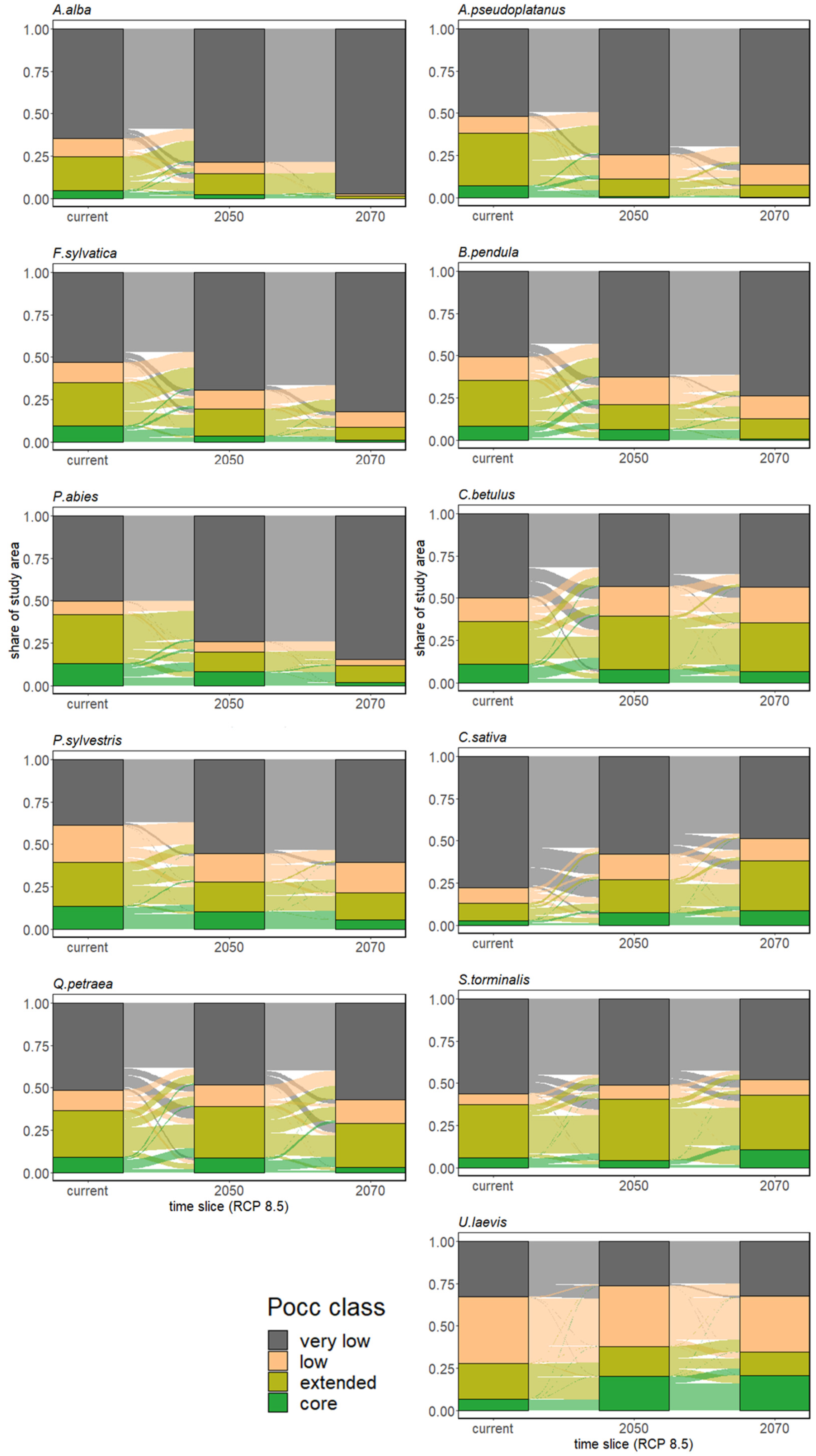
3.2.1. Range Size Dynamic
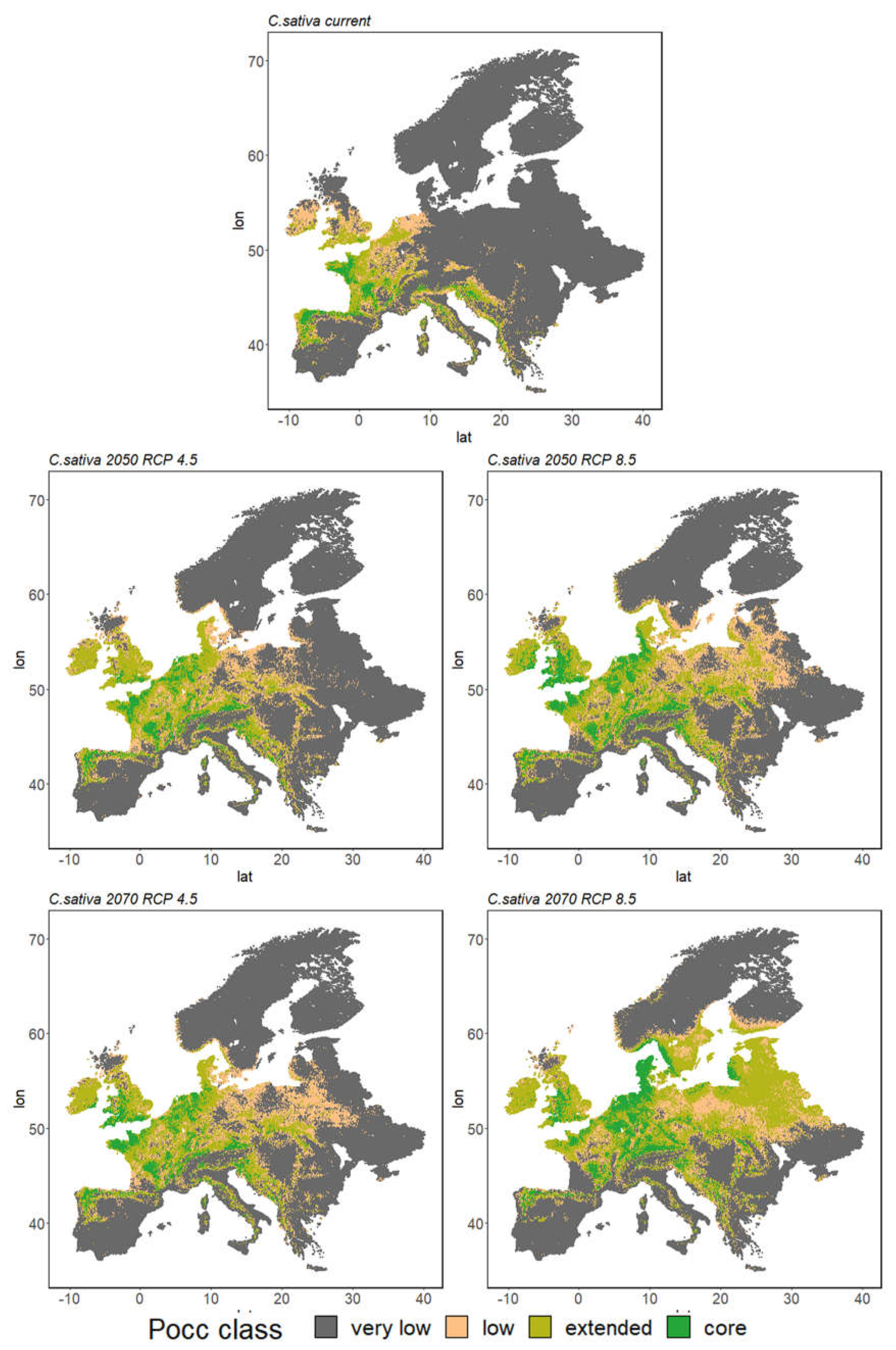
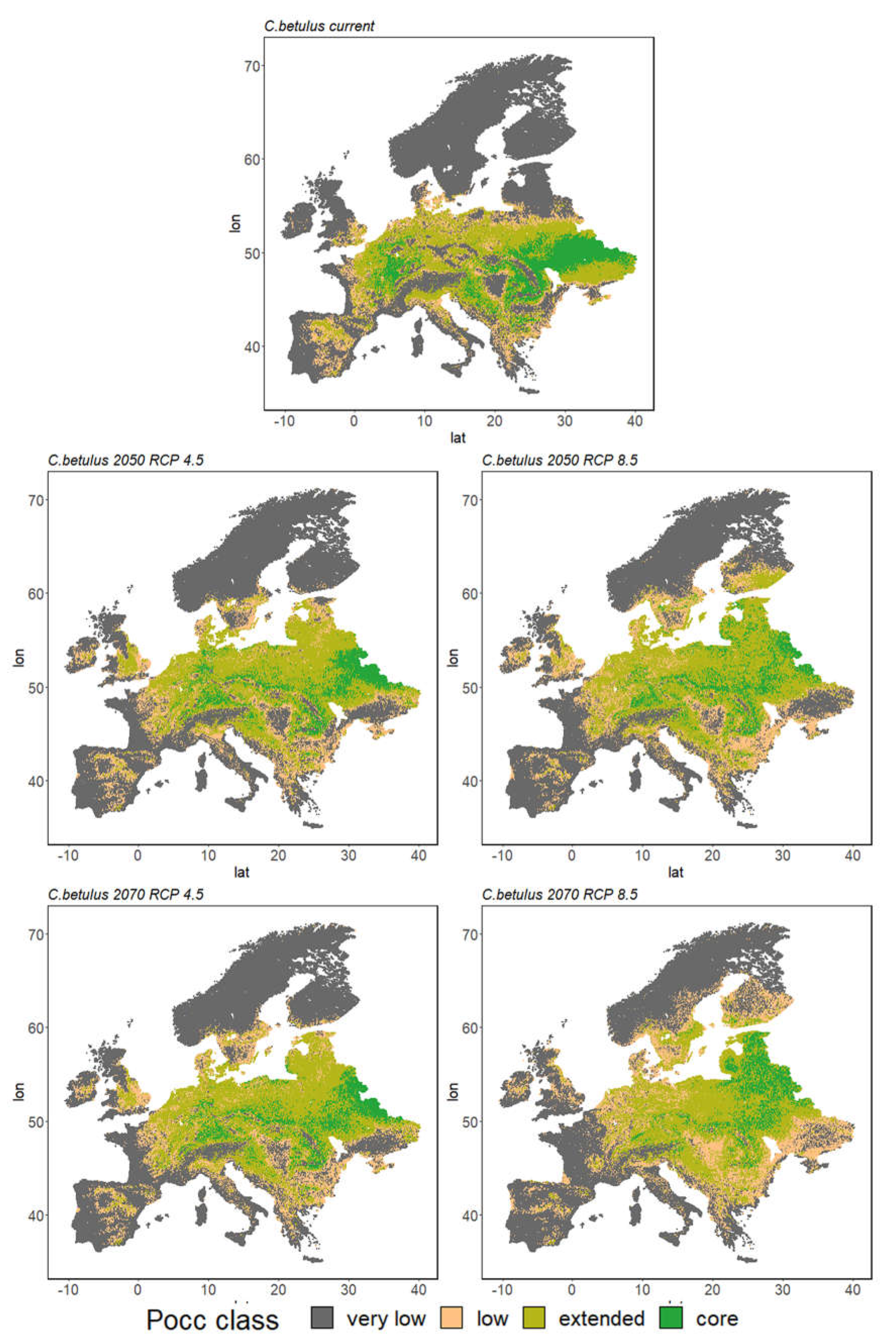
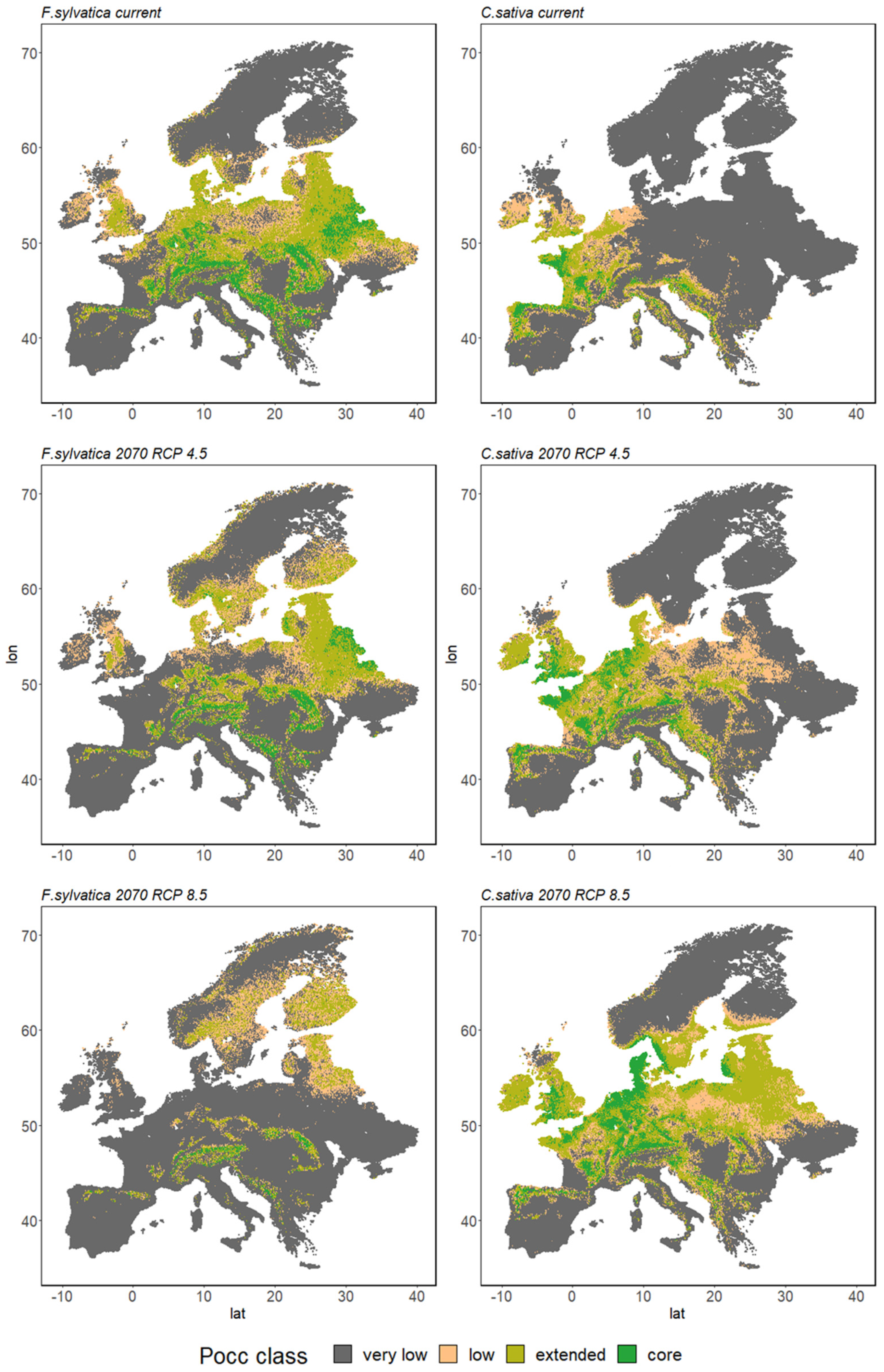
3.2.2. Species Range Shifts
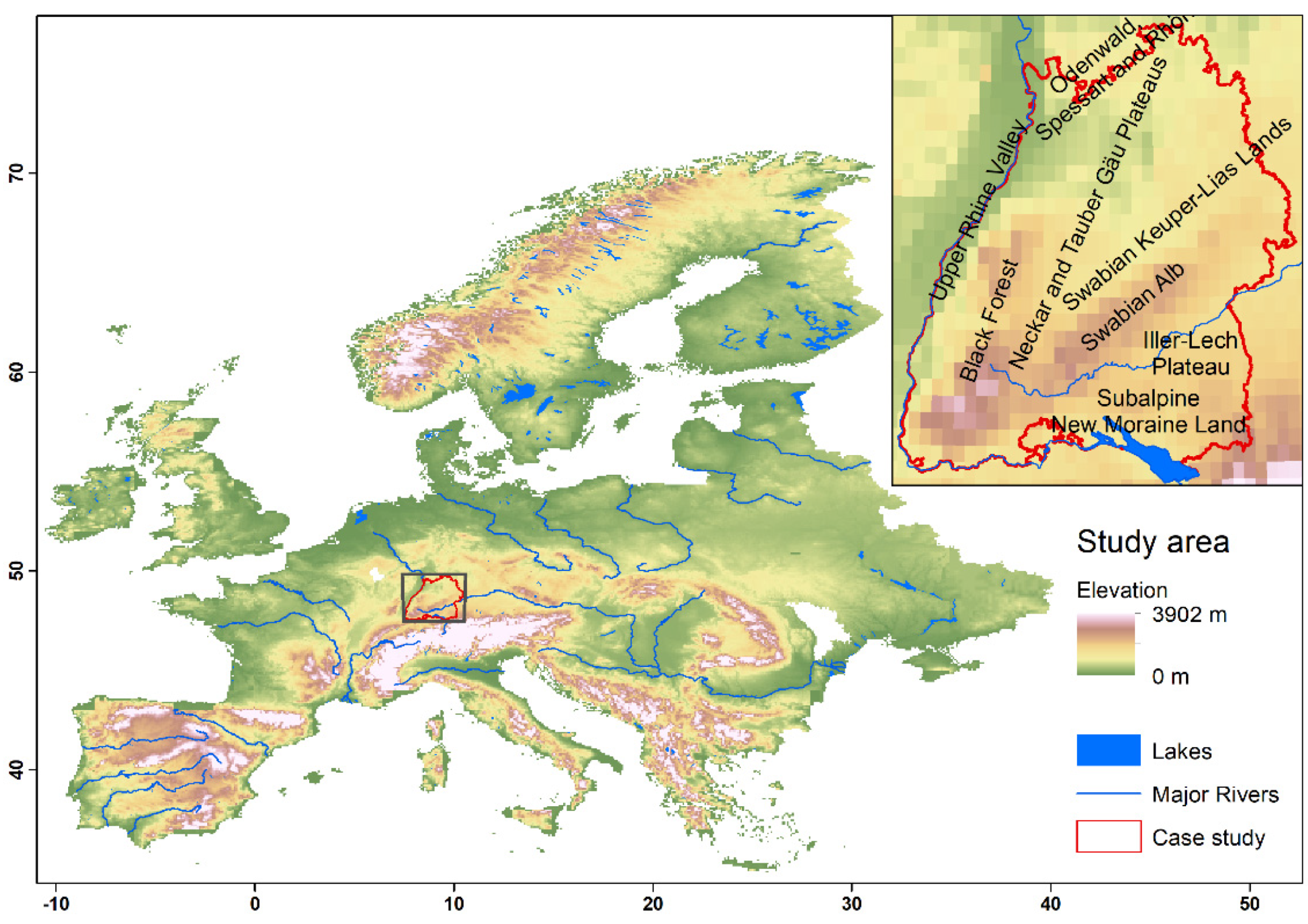
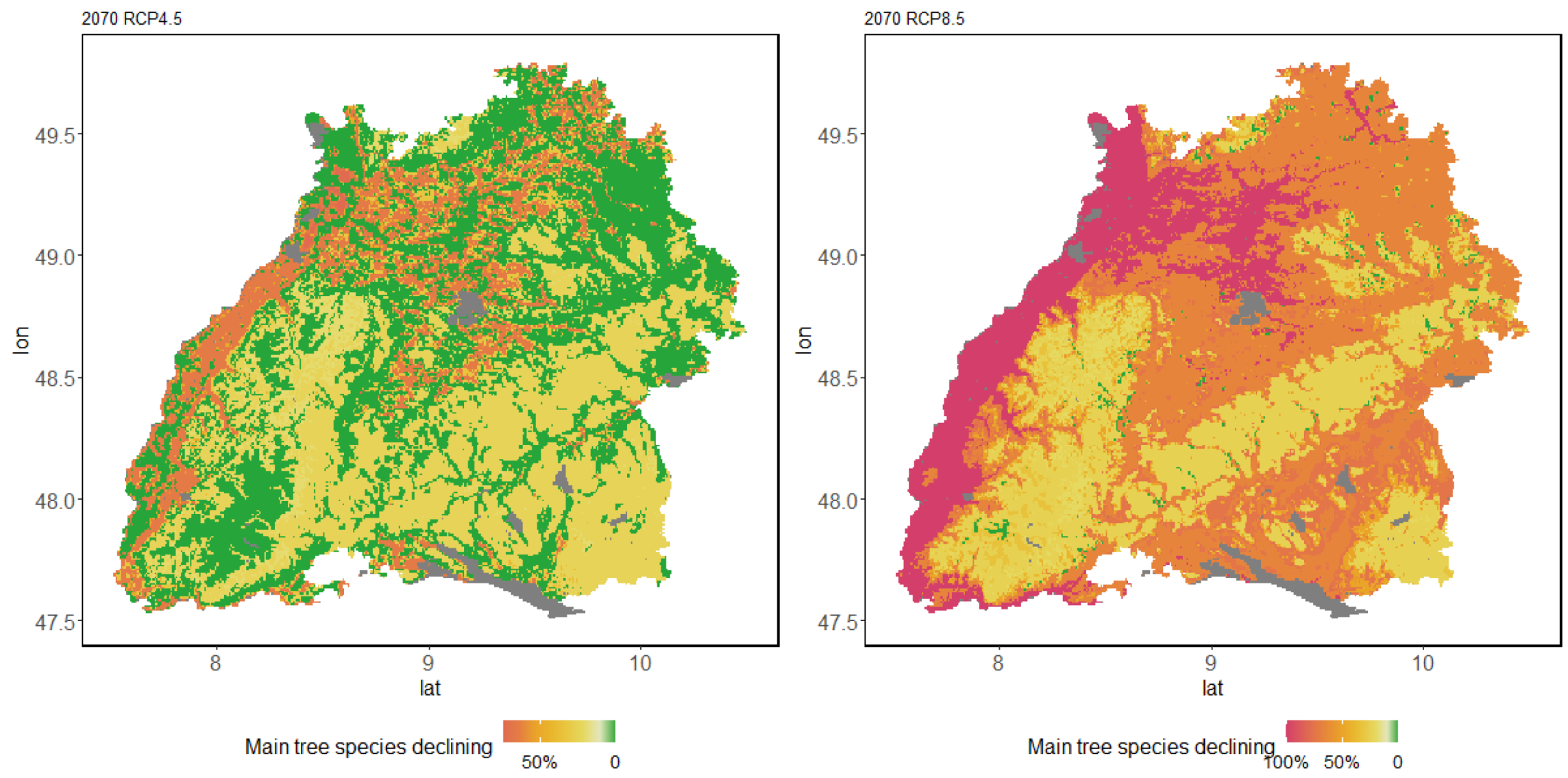
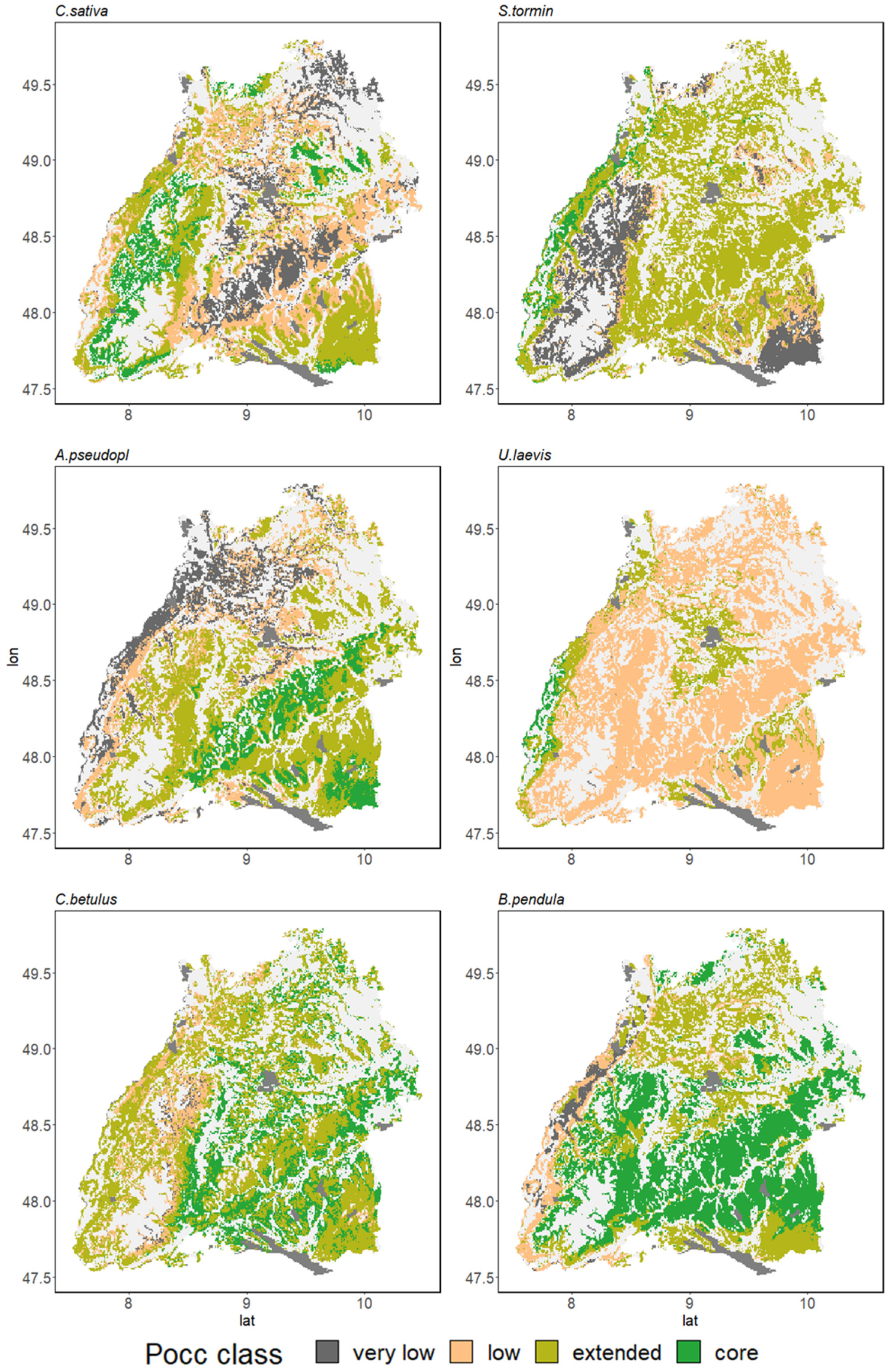
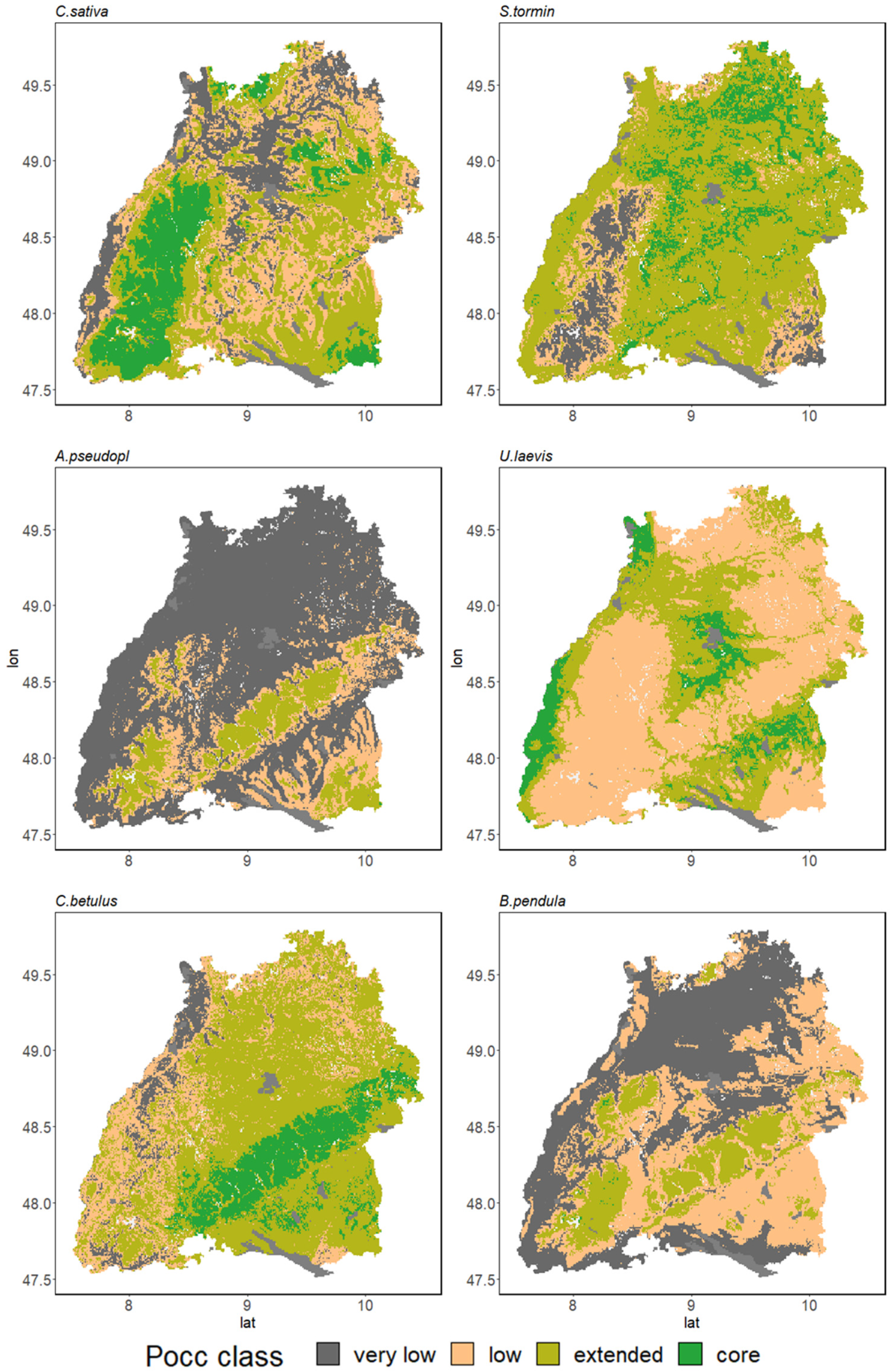
3.3. Range Dynamics in the Case Study of Baden-Wuerttemberg
3.3.1. Major Tree Species Decline
3.3.2. Conversion Potential of Alternative European Natives
4. Discussion
4.1. Growing Urgency: Major Tree Species Decline across Europe
4.2. Adapting to Climate Change with Minor Natives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sp. | Current | RCP 4.5 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2050 | 2070 | |||||||||||
| core | ext. | low | v.low | core | ext. | low | v.low | core | ext. | low | v.low | |
| Aa | 5% | 20% | 11% | 65% | 4% | 23% | 7% | 66% | 3% | 16% | 8% | 72% |
| Ap | 7% | 31% | 10% | 52% | 2% | 25% | 12% | 61% | 1% | 21% | 14% | 64% |
| Bp | 8% | 27% | 14% | 51% | 12% | 18% | 14% | 56% | 7% | 18% | 16% | 59% |
| Cb | 11% | 25% | 14% | 50% | 8% | 30% | 16% | 46% | 8% | 30% | 17% | 45% |
| Cs | 3% | 10% | 9% | 78% | 5% | 17% | 12% | 66% | 6% | 18% | 15% | 62% |
| Fs | 9% | 26% | 12% | 53% | 9% | 19% | 11% | 61% | 5% | 19% | 11% | 64% |
| Pa | 13% | 29% | 8% | 50% | 8% | 19% | 7% | 67% | 9% | 15% | 7% | 69% |
| Ps | 14% | 26% | 22% | 39% | 11% | 19% | 20% | 50% | 11% | 20% | 17% | 52% |
| Qp | 9% | 27% | 12% | 51% | 17% | 27% | 11% | 46% | 11% | 29% | 12% | 48% |
| St | 6% | 31% | 7% | 56% | 5% | 36% | 8% | 51% | 5% | 36% | 7% | 51% |
| Ul | 7% | 21% | 39% | 33% | 15% | 20% | 34% | 31% | 19% | 18% | 36% | 27% |
| Sp. | Current | RCP 8.5 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2050 | 2070 | |||||||||||
| core | ext. | low | v.low | core | ext. | low | v.low | core | ext. | low | v.low | |
| Aa | 5% | 20% | 11% | 65% | 2% | 12% | 7% | 79% | 0% | 1% | 1% | 97% |
| Ap | 7% | 31% | 10% | 52% | 1% | 10% | 14% | 75% | 0% | 7% | 12% | 80% |
| Bp | 8% | 27% | 14% | 51% | 6% | 14% | 16% | 63% | 1% | 12% | 13% | 74% |
| Cb | 11% | 25% | 14% | 50% | 8% | 31% | 18% | 43% | 7% | 29% | 21% | 43% |
| Cs | 3% | 10% | 9% | 78% | 7% | 20% | 15% | 58% | 9% | 30% | 13% | 49% |
| Fs | 9% | 26% | 12% | 53% | 4% | 16% | 11% | 69% | 1% | 7% | 9% | 82% |
| Pa | 13% | 29% | 8% | 50% | 8% | 12% | 6% | 74% | 2% | 10% | 4% | 85% |
| Ps | 14% | 26% | 22% | 39% | 10% | 17% | 17% | 56% | 6% | 16% | 18% | 61% |
| Qp | 9% | 27% | 12% | 51% | 9% | 30% | 13% | 48% | 3% | 26% | 14% | 57% |
| St | 6% | 31% | 7% | 56% | 5% | 36% | 8% | 51% | 11% | 33% | 9% | 48% |
| Ul | 7% | 21% | 39% | 33% | 20% | 18% | 36% | 26% | 21% | 14% | 33% | 32% |




References
- Ray, D.; Bathgate, S.; Moseley, D.; Taylor, P.; Nicoll, B.; Pizzirani, S.; Gardiner, B. Comparing the provision of ecosystem services in plantation forests under alternative climate change adaptation management options in Wales. Reg. Environ. Chang. 2014, 15, 1501–1513. [Google Scholar] [CrossRef] [Green Version]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl. Ecol. 2020, 45, 86–103. [Google Scholar] [CrossRef]
- La Porta, N.; Capretti, P.; Thomsen, I.M.; Kasanen, R.; Hietala, A.M.; Von Weissenberg, K. Forest pathogens with higher damage potential due to climate change in Europe. Can. J. Plant Pathol. 2008, 30, 177–195. [Google Scholar] [CrossRef]
- Beckage, B.; Gross, L.J.; Lacasse, K.; Carr, E.; Metcalf, S.S.; Winter, J.M.; Howe, P.D.; Fefferman, N.; Franck, T.; Zia, A.; et al. Linking models of human behaviour and climate alters projected climate change. Nat. Clim. Chang. 2018, 8, 79–84. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Kriegler, E.; Ebi, K.L.; Kemp-Benedict, E.; Riahi, K.; Rothman, D.S.; van Ruijven, B.J.; van Vuuren, D.P.; Birkmann, J.; Kok, K.; et al. The roads ahead: Narratives for shared socioeconomic pathways describing world futures in the 21st century. Glob. Environ. Chang. 2017, 42, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, T.G. Atmospheric circulation as a source of uncertainty in climate change projections. Nat. Geosci. 2014, 7, 703–708. [Google Scholar] [CrossRef]
- Yousefpour, R.; Hanewinkel, M. Climate Change and Decision-Making Under Uncertainty. Curr. For. Rep. 2016, 2, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef]
- Seidl, R.; Lexer, M.J. Forest management under climatic and social uncertainty: Trade-offs between reducing climate change impacts and fostering adaptive capacity. J. Environ. Manag. 2013, 114, 461–469. [Google Scholar] [CrossRef]
- Hörl, J.; Keller, K.; Yousefpour, R. Reviewing the performance of adaptive forest management strategies with robustness analysis. For. Policy Econ. 2020, 119, 102289. [Google Scholar] [CrossRef]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Mori, A.S.; Lertzman, K.P.; Gustafsson, L.; Cadotte, M. Biodiversity and ecosystem services in forest ecosystems: A research agenda for applied forest ecology. J. Appl. Ecol. 2017, 54, 12–27. [Google Scholar] [CrossRef]
- Thurm, E.A.; Hernandez, L.; Baltensweiler, A.; Ayan, S.; Rasztovits, E.; Bielak, K.; Zlatanov, T.M.; Hladnik, D.; Balic, B.; Freudenschuss, A.; et al. Alternative tree species under climate warming in managed European forests. For. Ecol. Manag. 2018, 430, 485–497. [Google Scholar] [CrossRef]
- Buras, A.; Menzel, A. Projecting Tree Species Composition Changes of European Forests for 2061–2090 under RCP 4.5 and RCP 8.5 Scenarios. Front. Plant Sci. 2018, 9, 1986. [Google Scholar] [CrossRef] [Green Version]
- Walentowski, H.; Falk, W.; Mette, T.; Kunz, J.; Bräuning, A.; Meinardus, C.; Zang, C.; Sutcliffe, L.M.E.; Leuschner, C. Assessing future suitability of tree species under climate change by multiple methods: A case study in southern Germany. Ann. For. Res. 2017, 60, 101–126. [Google Scholar] [CrossRef] [Green Version]
- Sousa-Silva, R.; Verbist, B.; Lomba, Â.; Valent, P.; Suškevičs, M.; Picard, O.; Hoogstra-Klein, M.A.; Cosofret, V.-C.; Bouriaud, L.; Ponette, Q.; et al. Adapting forest management to climate change in Europe: Linking perceptions to adaptive responses. For. Policy Econ. 2018, 90, 22–30. [Google Scholar] [CrossRef]
- Vinceti, B.; Manica, M.; Lauridsen, N.; Verkerk, P.J.; Lindner, M.; Fady, B. Managing forest genetic resources as a strategy to adapt forests to climate change: Perceptions of European forest owners and managers. Eur. J. For. Res. 2020, 139, 1107–1119. [Google Scholar] [CrossRef]
- Pyttel, P.; Kunz, J.; Bauhus, J. Growth, regeneration and shade tolerance of the Wild Service Tree (Sorbus torminalis (L.) Crantz) in aged oak coppice forests. Trees 2013, 27, 1609–1619. [Google Scholar] [CrossRef]
- Thurm, E.A.; Falk, W. Standortsansprüche seltener Baumarten. AFZ-DerWald. 2019, pp. 32–35. Available online: https://www.researchgate.net/publication/335033693_Standortsanspruche_seltener_Baumarten (accessed on 22 April 2022).
- de Rigo, D.; Bosco, C.; San-Miguel-Ayanz, J.; Houston Durrant, T.; Barredo, J.I.; Strona, G.; Caudullo, G.; Di Leo, M.; Boca, R. Forest resources in Europe: An integrated perspective on ecosystem services, disturbances and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Orsi, F.; Ciolli, M.; Primmer, E.; Varumo, L.; Geneletti, D. Mapping hotspots and bundles of forest ecosystem services across the European Union. Land Use Policy 2020, 99, 104840. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; Jactel, H.; Scherer-Lorenzen, M.; Garcia-Valdes, R.; Bugmann, H. Long-term response of forest productivity to climate change is mostly driven by change in tree species composition. Sci. Rep. 2018, 8, 5627. [Google Scholar] [CrossRef] [Green Version]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2012, 3, 203–207. [Google Scholar] [CrossRef]
- Eggers, J.; Holmgren, S.; Nordström, E.-M.; Lämås, T.; Lind, T.; Öhman, K. Balancing different forest values: Evaluation of forest management scenarios in a multi-criteria decision analysis framework. For. Policy Econ. 2019, 103, 55–69. [Google Scholar] [CrossRef]
- Balana, B.B.; Mathijs, E.; Muys, B. Assessing the sustainability of forest management: An application of multi-criteria decision analysis to community forests in northern Ethiopia. J. Environ. Manag. 2010, 91, 1294–1304. [Google Scholar] [CrossRef]
- Schwenk, W.S.; Donovan, T.M.; Keeton, W.S.; Nunery, J.S. Carbon storage, timber production, and biodiversity: Comparing ecosystem services with multi-criteria decision analysis. Ecol. Appl. 2012, 22, 1612–1627. [Google Scholar] [CrossRef] [PubMed]
- de Avila, A.L.; Häring, B.; Rheinbay, B.; Brüchert, F.; Hirsch, M.; Albrecht, A. Artensteckbriefe 2.0 Alternative Baumarten im Klimawandel—Eine Stoffsammlung; Forstliche Versuchs- und Forschungsanstalt Baden-Württemberg (FVA): Freiburg, Germany, 2021. [Google Scholar]
- Booth, T.H. Species distribution modelling tools and databases to assist managing forests under climate change. For. Ecol. Manag. 2018, 430, 196–203. [Google Scholar] [CrossRef]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species distribution modelling to support forest management. A literature review. Ecol. Model. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Zurell, D.; Jeltsch, F.; Dormann, C.F.; Schröder, B. Static species distribution models in dynamically changing systems: How good can predictions really be? Ecography 2009, 32, 733–744. [Google Scholar] [CrossRef]
- Thuiller, W. Patterns and uncertainties of species’ range shifts under climate change. Glob. Chang. Biol. 2004, 10, 2020–2027. [Google Scholar] [CrossRef]
- Mauri, A.; Strona, G.; San-Miguel-Ayanz, J. EU-Forest, a high-resolution tree occurrence dataset for Europe. Sci. Data 2017, 4, 160123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Gardner, A.S.; Maclean, I.M.D.; Gaston, K.J.; Serra-Diaz, J. Climatic predictors of species distributions neglect biophysiologically meaningful variables. Divers. Distrib. 2019, 25, 1318–1333. [Google Scholar] [CrossRef] [Green Version]
- Karger, D.N.; Conrad, O.; Bohner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hengl, T.; Mendes de Jesus, J.; Heuvelink, G.B.; Ruiperez Gonzalez, M.; Kilibarda, M.; Blagotic, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, E.; Mellert, K.H.; Göttlein, A. Nährstoffstatus naturnaher Böden in Europa. Wald. Landsch. Nat. 2019, 18, 5–13. [Google Scholar]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Brun, P.; Thuiller, W.; Chauvier, Y.; Pellissier, L.; Wüest, R.O.; Wang, Z.; Zimmermann, N.E. Model complexity affects species distribution projections under climate change. J. Biogeogr. 2019, 47, 130–142. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paz, S.; Frelich, L.E.; Jagodzinski, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Goberville, E.; Beaugrand, G.; Hautekeete, N.C.; Piquot, Y.; Luczak, C. Uncertainties in the projection of species distributions related to general circulation models. Ecol. Evol. 2015, 5, 1100–1116. [Google Scholar] [CrossRef]
- Porfirio, L.L.; Harris, R.M.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.L.; Mackey, B. Improving the use of species distribution models in conservation planning and management under climate change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [Green Version]
- Dubrovsky, M.; Trnka, M.; Holman, I.P.; Svobodova, E.; Harrison, P.A. Developing a reduced-form ensemble of climate change scenarios for Europe and its application to selected impact indicators. Clim. Chang. 2014, 128, 169–186. [Google Scholar] [CrossRef]
- Kaini, S.; Nepal, S.; Pradhananga, S.; Gardner, T.; Sharma, A.K. Representative general circulation models selection and downscaling of climate data for the transboundary Koshi river basin in China and Nepal. Int. J. Climatol. 2020, 40, 4131–4149. [Google Scholar] [CrossRef] [Green Version]
- Fajardo, J.; Corcoran, D.; Roehrdanz, P.R.; Hannah, L.; Marquet, P.A.; Kriticos, D. GCMcompareR: A web application to assess differences and assist in the selection of general circulation models for climate change research. Methods Ecol. Evol. 2020, 11, 656–663. [Google Scholar] [CrossRef] [Green Version]
- Moss, R.H.; Edmonds, J.A.; Hibbard, K.A.; Manning, M.R.; Rose, S.K.; van Vuuren, D.P.; Carter, T.R.; Emori, S.; Kainuma, M.; Kram, T.; et al. The next generation of scenarios for climate change research and assessment. Nature 2010, 463, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Rogelj, J.; Meinshausen, M.; Knutti, R. Global warming under old and new scenarios using IPCC climate sensitivity range estimates. Nat. Clim. Chang. 2012, 2, 248–253. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Ferro, C.A.T.; Stephenson, D.B. Extremal Dependence Indices: Improved Verification Measures for Deterministic Forecasts of Rare Binary Events. Weather Forecast. 2011, 26, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F.; Georges, M.D.; Thuiller, C.W. Package ‘Biomod2’; Species Distribution Modeling within an Ensemble Forecasting Framework. CRAN. 2021. Available online: https://cran.r-project.org/web/packages/biomod2/biomod2.pdf (accessed on 23 April 2022).
- Jiménez-Valverde, A. Prevalence affects the evaluation of discrimination capacity in presence-absence species distribution models. Biodivers. Conserv. 2021, 30, 1331–1340. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Michiels, H.G.; Kandler, G. Converting probabilistic tree species range shift projections into meaningful classes for management. J. Environ. Manag. 2014, 134, 153–165. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.0.5; Programming Language; The R Project for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Chakraborty, D.; Móricz, N.; Rasztovits, E.; Dobor, L.; Schueler, S. Provisioning forest and conservation science with high-resolution maps of potential distribution of major European tree species under climate change. Ann. For. Sci. 2021, 78, 26. [Google Scholar] [CrossRef]
- Mauri, A.; De Rigo, D.; Caudullo, G. Abies alba in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016; pp. 48–49. [Google Scholar]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.D.; Steinacher, M.; Untenecker, J.; Vescovi, E.; Allen, J.R.M.; Carraro, G.; Conedera, M.; et al. The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef] [Green Version]
- Hanewinkel, M.; Hummel, S.; Cullmann, D.A. Modelling and economic evaluation of forest biome shifts under climate change in Southwest Germany. For. Ecol. Manag. 2010, 259, 710–719. [Google Scholar] [CrossRef]
- Liang, Y.; Duveneck, M.J.; Gustafson, E.J.; Serra-Diaz, J.M.; Thompson, J.R. How disturbance, competition, and dispersal interact to prevent tree range boundaries from keeping pace with climate change. Glob. Chang. Biol. 2018, 24, e335–e351. [Google Scholar] [CrossRef] [PubMed]
- Sittaro, F.; Paquette, A.; Messier, C.; Nock, C.A. Tree range expansion in eastern North America fails to keep pace with climate warming at northern range limits. Glob. Chang. Biol. 2017, 23, 3292–3301. [Google Scholar] [CrossRef] [PubMed]
- Heiland, L.; Kunstler, G.; Ruiz-Benito, P.; Buras, A.; Dahlgren, J.; Hülsmann, L. Divergent occurrences of juvenile and adult trees are explained by both environmental change and ontogenetic effects. Ecography 2022, 2022, e06042. [Google Scholar] [CrossRef]
- Iverson, L.R.; McKenzie, D. Tree-species range shifts in a changing climate: Detecting, modeling, assisting. Landsc. Ecol. 2013, 28, 879–889. [Google Scholar] [CrossRef]
- Mishra, S.; Hättenschwiler, S.; Yang, X. The plant microbiome: A missing link for the understanding of community dynamics and multifunctionality in forest ecosystems. Appl. Soil Ecol. 2020, 145, 103345. [Google Scholar] [CrossRef]
- Plomion, C.; Bastien, C.; Bogeat-Triboulot, M.-B.; Bouffier, L.; Déjardin, A.; Duplessis, S.; Fady, B.; Heuertz, M.; Le Gac, A.-L.; Le Provost, G.; et al. Forest tree genomics: 10 achievements from the past 10 years and future prospects. Ann. For. Sci. 2016, 73, 77–103. [Google Scholar] [CrossRef] [Green Version]
- Bettenfeld, P.; Fontaine, F.; Trouvelot, S.; Fernandez, O.; Courty, P.-E. Woody Plant Declines. What’s Wrong with the Microbiome? Trends Plant Sci. 2020, 25, 381–394. [Google Scholar] [CrossRef]
- Doonan, J.M.; Broberg, M.; Denman, S.; McDonald, J.E. Host-microbiota-insect interactions drive emergent virulence in a complex tree disease. Proc. Biol. Sci. 2020, 287, 20200956. [Google Scholar] [CrossRef]
- Ware, I.M.; Van Nuland, M.E.; Yang, Z.K.; Schadt, C.W.; Schweitzer, J.A.; Bailey, J.K. Climate-driven divergence in plant-microbiome interactions generates range-wide variation in bud break phenology. Commun. Biol. 2021, 4, 748. [Google Scholar] [CrossRef]
- Van Nuland, M.E.; Ware, I.M.; Schadt, C.W.; Yang, Z.; Bailey, J.K.; Schweitzer, J.A. Natural soil microbiome variation affects spring foliar phenology with consequences for plant productivity and climate-driven range shifts. New Phytol. 2021, 232, 762–775. [Google Scholar] [CrossRef]
- Kunstler, G.; Guyennon, A.; Ratcliffe, S.; Rüger, N.; Ruiz-Benito, P.; Childs, D.Z.; Dahlgren, J.; Lehtonen, A.; Thuiller, W.; Wirth, C.; et al. Demographic performance of European tree species at their hot and cold climatic edges. J. Ecol. 2020, 109, 1041–1054. [Google Scholar] [CrossRef]
- Maringer, J.; Stelzer, A.-S.; Paul, C.; Albrecht, A.T. Ninety-five years of observed disturbance-based tree mortality modeled with climate-sensitive accelerated failure time models. Eur. J. For. Res. 2020, 140, 255–272. [Google Scholar] [CrossRef]
- Lorenz, R.; Stalhandske, Z.; Fischer, E.M. Detection of a Climate Change Signal in Extreme Heat, Heat Stress, and Cold in Europe From Observations. Geophys. Res. Lett. 2019, 46, 8363–8374. [Google Scholar] [CrossRef] [Green Version]
- Vitali, V.; Buntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Vitasse, Y.; Bottero, A.; Cailleret, M.; Bigler, C.; Fonti, P.; Gessler, A.; Levesque, M.; Rohner, B.; Weber, P.; Rigling, A.; et al. Contrasting resistance and resilience to extreme drought and late spring frost in five major European tree species. Glob. Chang. Biol. 2019, 25, 3781–3792. [Google Scholar] [CrossRef]
- Mette, T.; Brandl, S.; Kölling, C. Climate Analogues for Temperate European Forests to Raise Silvicultural Evidence Using Twin Regions. Sustainability 2021, 13, 6522. [Google Scholar] [CrossRef]
- Broadmeadow, M.S.J.; Ray, D.; Samuel, C.J.A. Climate change and the future for broadleaved tree species in Britain. For. Int. J. For. Res. 2005, 78, 145–161. [Google Scholar] [CrossRef] [Green Version]
- Morecroft, M.D.; Stokes, V.J.; Taylor, M.E.; Morison, J.I.L. Effects of climate and management history on the distribution and growth of sycamore (Acer pseudoplatanus L.) in a southern British woodland in comparison to native competitors. Forestry 2008, 81, 59–74. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Feifel, M.; Karimi, Z.; Schuldt, B.; Choat, B.; Jansen, S. Leaf gas exchange performance and the lethal water potential of five European species during drought. Tree Physiol. 2016, 36, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Conedera, M.; Tinner, W.; Krebs, P.; de Rigo, D.; Caudullo, G. Castanea sativa in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; p. e0125e. [Google Scholar]
- Gossner, M.M.; Wende, B.; Levick, S.; Schall, P.; Floren, A.; Linsenmair, K.E.; Steffan-Dewenter, I.; Schulze, E.-D.; Weisser, W.W. Deadwood enrichment in European forests—Which tree species should be used to promote saproxylic beetle diversity? Biol. Conserv. 2016, 201, 92–102. [Google Scholar] [CrossRef]
- Venturas, M.; Fuentes-Utrilla, P.; López, R.; Perea, R.; Fernández, V.; Gascó, A.; Guzmán, P.; Li, M.; Rodríguez-Calcerrada, J.; Miranda, E.; et al. Ulmus laevis in the Iberian Peninsula: A review of its ecology and conservation. Iforest-Biogeosci. For. 2015, 8, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Vakkari, P.; Rusanen, M.; Kärkkäinen, K. High Genetic Differentiation in Marginal Populations of European White Elm (Ulmus laevis). Silva Fenn. 2009, 43, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; López, R.; Venturas, M.; Pita, P.; Gordaliza, G.G.; Gil, L.; Rodríguez-Calcerrada, J. Greater resistance to flooding of seedlings of Ulmus laevis than Ulmus minor is related to the maintenance of a more positive carbon balance. Trees 2015, 29, 835–848. [Google Scholar] [CrossRef]
- Danescu, A.; Albrecht, A.T.; Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 2016, 182, 319–333. [Google Scholar] [CrossRef]
- Pretzsch, H.; del Río, M.; Ammer, C.; Avdagic, A.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; Dirnberger, G.; Drössler, L.; et al. Growth and yield of mixed versus pure stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2015, 134, 927–947. [Google Scholar] [CrossRef] [Green Version]
- Albrich, K.; Rammer, W.; Thom, D.; Seidl, R. Trade-offs between temporal stability and level of forest ecosystem services provisioning under climate change. Ecol. Appl. 2018, 28, 1884–1896. [Google Scholar] [CrossRef] [Green Version]
- Vacek, Z.; Vacek, S.; Slanař, J.; Bílek, L.; Bulušek, D.; Štefančík, I.; Králíček, I.; Vančura, K. Adaption of Norway spruce and European beech forests under climate change: From resistance to close-to-nature silviculture. Cent. Eur. For. J. 2019, 65, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Neuner, S.; Albrecht, A.; Cullmann, D.; Engels, F.; Griess, V.C.; Hahn, W.A.; Hanewinkel, M.; Hartl, F.; Kolling, C.; Staupendahl, K.; et al. Survival of Norway spruce remains higher in mixed stands under a dryer and warmer climate. Glob. Chang. Biol. 2015, 21, 935–946. [Google Scholar] [CrossRef] [Green Version]
- Schelhaas, M.-J.; Nabuurs, G.-J.; Hengeveld, G.; Reyer, C.; Hanewinkel, M.; Zimmermann, N.E.; Cullmann, D. Alternative forest management strategies to account for climate change-induced productivity and species suitability changes in Europe. Reg. Environ. Chang. 2015, 15, 1581–1594. [Google Scholar] [CrossRef] [Green Version]
- Priebe, J.; Reimerson, E.; Hallberg-Sramek, I.; Sténs, A.; Sandström, C.; Mårald, E. Transformative change in context—stakeholders’ understandings of leverage at the forest–climate nexus. Sustain. Sci. 2022. [Google Scholar] [CrossRef]
- Schmidt, M.; Mölder, A.; Schönfelder, E.; Engel, F.; Schmiedel, I.; Culmsee, H. Determining ancient woodland indicator plants for practical use: A new approach developed in northwest Germany. For. Ecol. Manag. 2014, 330, 228–239. [Google Scholar] [CrossRef]
- Hernández-Morcillo, M.; Torralba, M.; Baiges, T.; Bernasconi, A.; Bottaro, G.; Brogaard, S.; Bussola, F.; Díaz-Varela, E.; Geneletti, D.; Grossmann, C.M.; et al. Scanning the solutions for the sustainable supply of forest ecosystem services in Europe. Sustain. Sci. 2022. [Google Scholar] [CrossRef]
- Zurell, D.; Thuiller, W.; Pagel, J.; Cabral, J.S.; Munkemuller, T.; Gravel, D.; Dullinger, S.; Normand, S.; Schiffers, K.H.; Moore, K.A.; et al. Benchmarking novel approaches for modelling species range dynamics. Glob. Chang. Biol. 2016, 22, 2651–2664. [Google Scholar] [CrossRef] [PubMed]
- Lee-Yaw, J.A.; McCune, J.L.; Pironon, S.; Sheth, S.N. Species distribution models rarely predict the biology of real populations. Ecography 2021. [Google Scholar] [CrossRef]
- Diaz-Balteiro, L.; Romero, C. Making forestry decisions with multiple criteria: A review and an assessment. For. Ecol. Manag. 2008, 255, 3222–3241. [Google Scholar] [CrossRef]
- Peterson, M.L.; Doak, D.F.; Morris, W.F. Incorporating local adaptation into forecasts of species’ distribution and abundance under climate change. Glob. Chang. Biol. 2019, 25, 775–793. [Google Scholar] [CrossRef]

| Expl. Variables | Source | Cs | St | Cb | Qp | Ul | Ap | Pa | Fs | Aa | Ps | Bp |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Climate | ||||||||||||
| Bio 1 | [35] | √ | √ | √ | √ | |||||||
| Bio 5 | [35] | √ | √ | √ | √ | |||||||
| Bio 6 | [35] | √ | √ | √ | √ | √ | ||||||
| Bio 12 | [35] | √ | √ | √ | √ | √ | √ | √ | ||||
| Bio 18 | [35] | √ | √ | √ | √ | √ | √ | √ | √ | √ | ||
| Bio 19 | [35] | √ | √ | √ | √ | |||||||
| CCI | own computation | √ | √ | √ | √ | √ | √ | √ | √ | √ | ||
| GDD5 | own computation | √ | √ | √ | ||||||||
| Soil | ||||||||||||
| soil pH | [36] | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | |
| available water content | [36] | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| soil nutrient status | [37] | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| Species | SE 1 | SP 1 | TSS | SEDI | AUC |
|---|---|---|---|---|---|
| Abies alba | 0.918 | 0.811 | 0.730 | 0.871 | 0.932 |
| Acer pseudoplatanus | 0.865 | 0.713 | 0.578 | 0.749 | 0.866 |
| Betula pendula | 0.832 | 0.752 | 0.584 | 0.757 | 0.867 |
| Carpinus betulus | 0.857 | 0.741 | 0.598 | 0.759 | 0.880 |
| Castanea sativa | 0.879 | 0.770 | 0.649 | 0.803 | 0.892 |
| Fagus sylvatica | 0.862 | 0.741 | 0.603 | 0.774 | 0.883 |
| Picea abies | 0.910 | 0.818 | 0.728 | 0.866 | 0.928 |
| Pinus sylvestris | 0.850 | 0.783 | 0.633 | 0.787 | 0.897 |
| Quercus petraea | 0.884 | 0.740 | 0.624 | 0.785 | 0.887 |
| Sorbus torminalis | 0.899 | 0.740 | 0.639 | 0.841 | 0.903 |
| Ulmus laevis | 0.829 | 0.793 | 0.622 | 0.783 | 0.886 |
| Mean | 0.871 | 0.764 | 0.635 | 0.798 | 0.893 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koch, O.; de Avila, A.L.; Heinen, H.; Albrecht, A.T. Retreat of Major European Tree Species Distribution under Climate Change—Minor Natives to the Rescue? Sustainability 2022, 14, 5213. https://doi.org/10.3390/su14095213
Koch O, de Avila AL, Heinen H, Albrecht AT. Retreat of Major European Tree Species Distribution under Climate Change—Minor Natives to the Rescue? Sustainability. 2022; 14(9):5213. https://doi.org/10.3390/su14095213
Chicago/Turabian StyleKoch, Olef, Angela Luciana de Avila, Henry Heinen, and Axel Tim Albrecht. 2022. "Retreat of Major European Tree Species Distribution under Climate Change—Minor Natives to the Rescue?" Sustainability 14, no. 9: 5213. https://doi.org/10.3390/su14095213
APA StyleKoch, O., de Avila, A. L., Heinen, H., & Albrecht, A. T. (2022). Retreat of Major European Tree Species Distribution under Climate Change—Minor Natives to the Rescue? Sustainability, 14(9), 5213. https://doi.org/10.3390/su14095213