
The Future Climate under Different CO2 Emission Scenarios Significantly Influences the Potential Distribution of Achnatherum inebrians in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Occurrence Data
2.2. Environmental Variables
2.3. Ecological Niche Model
2.4. Data Analysis
3. Results
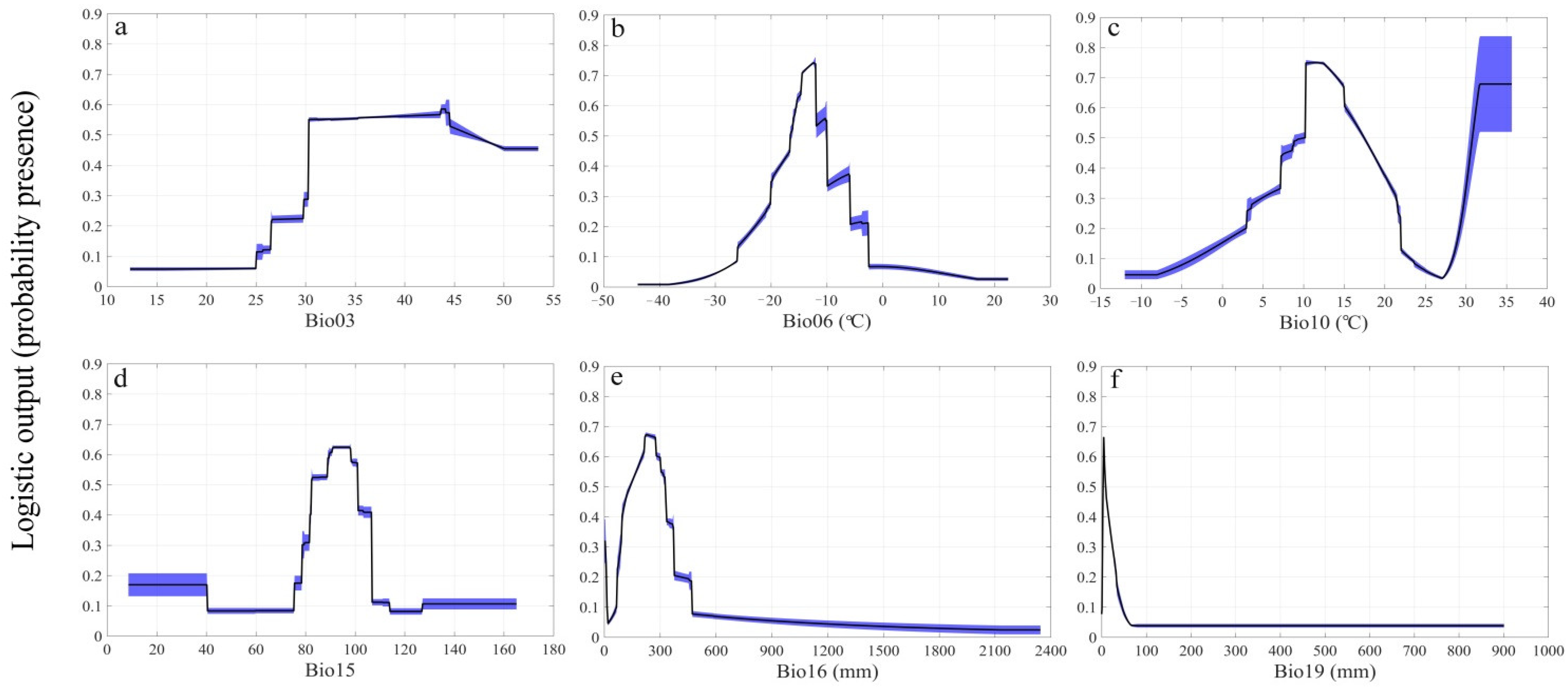
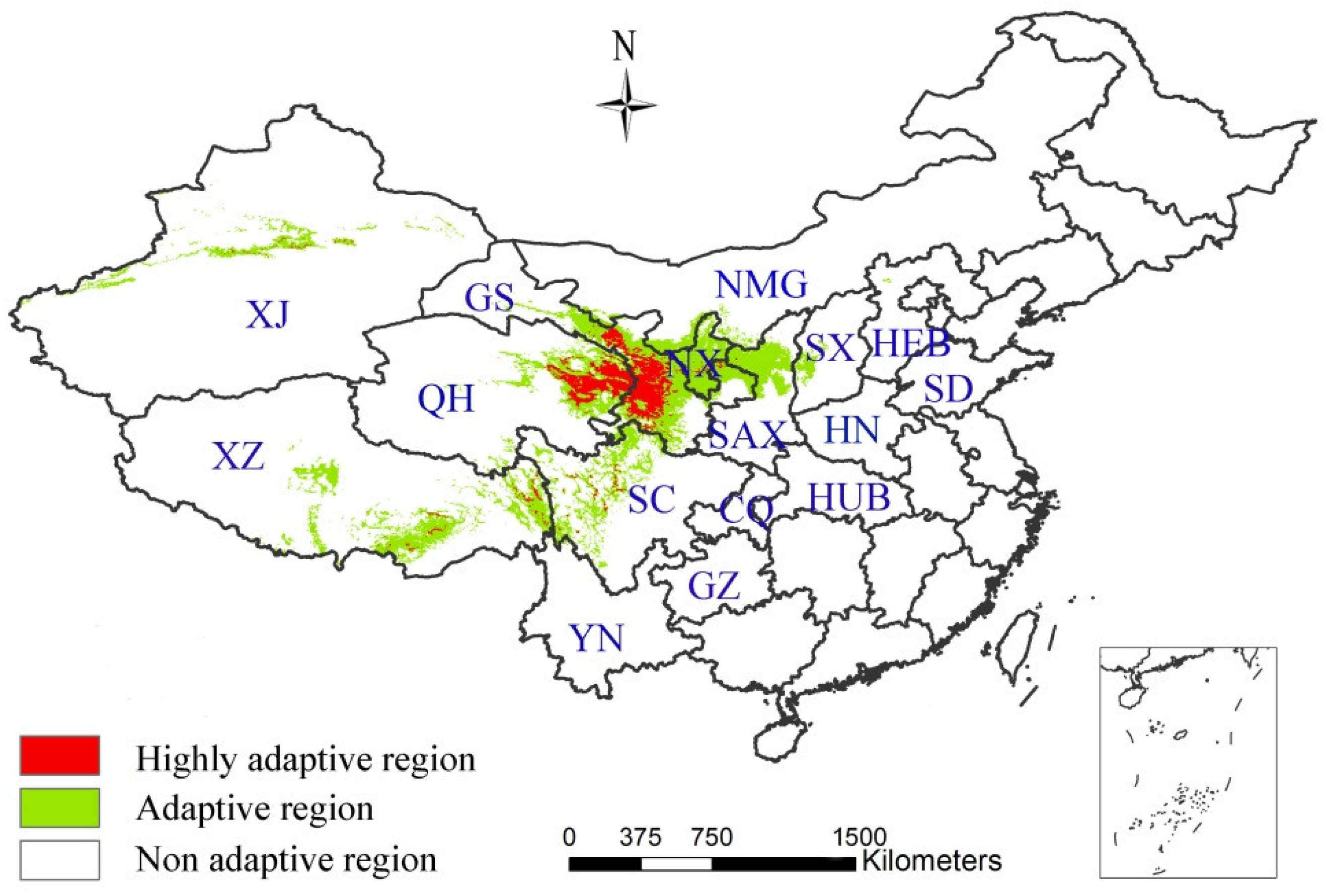
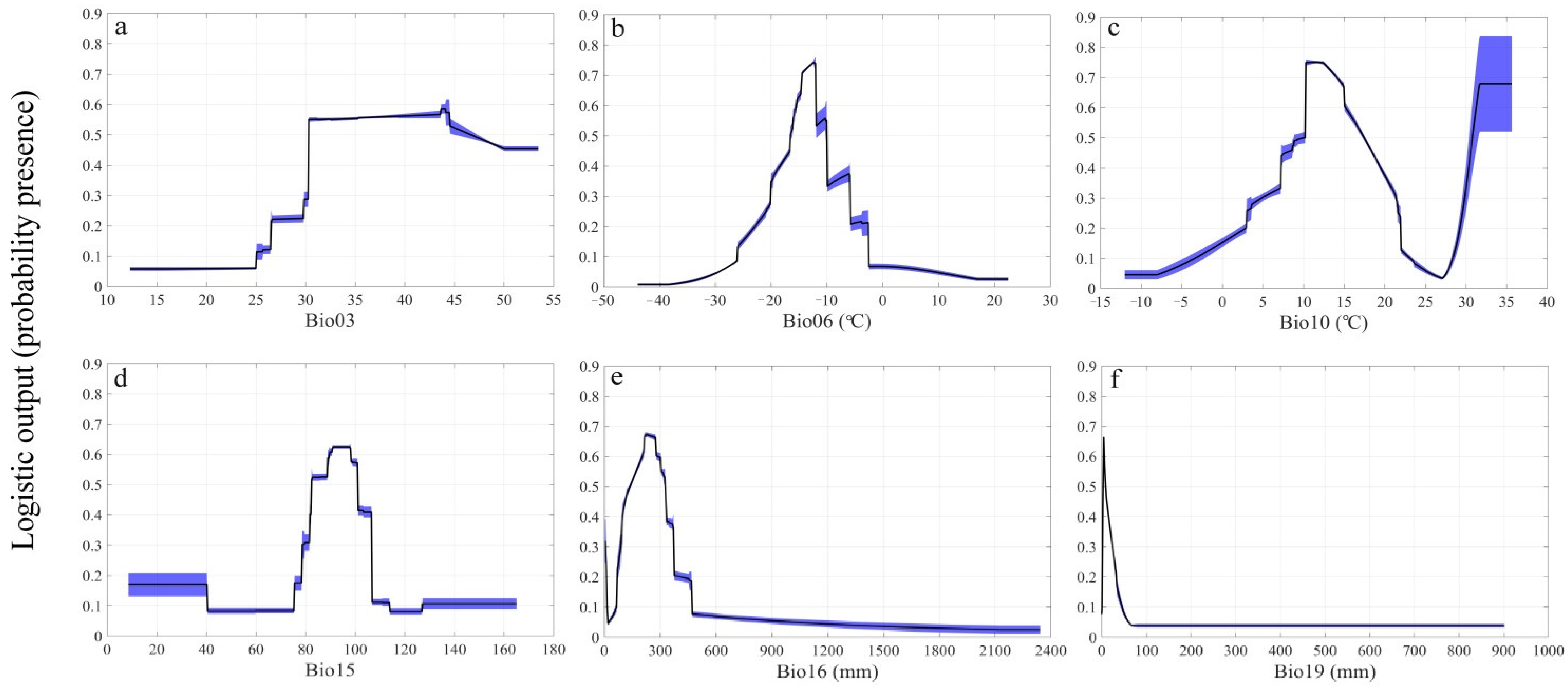
3.1. Model Performance and Importance of Predictor Variables
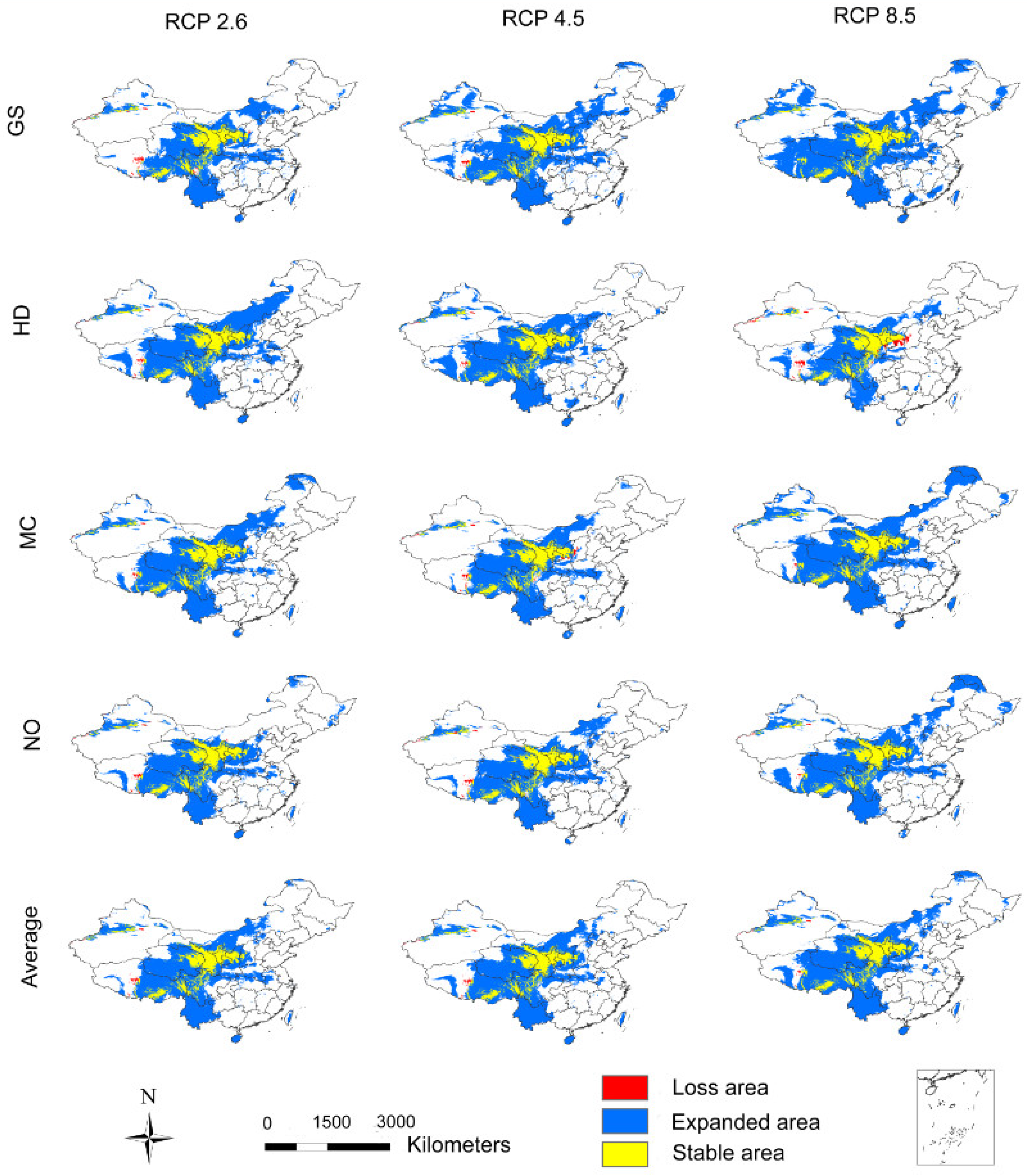
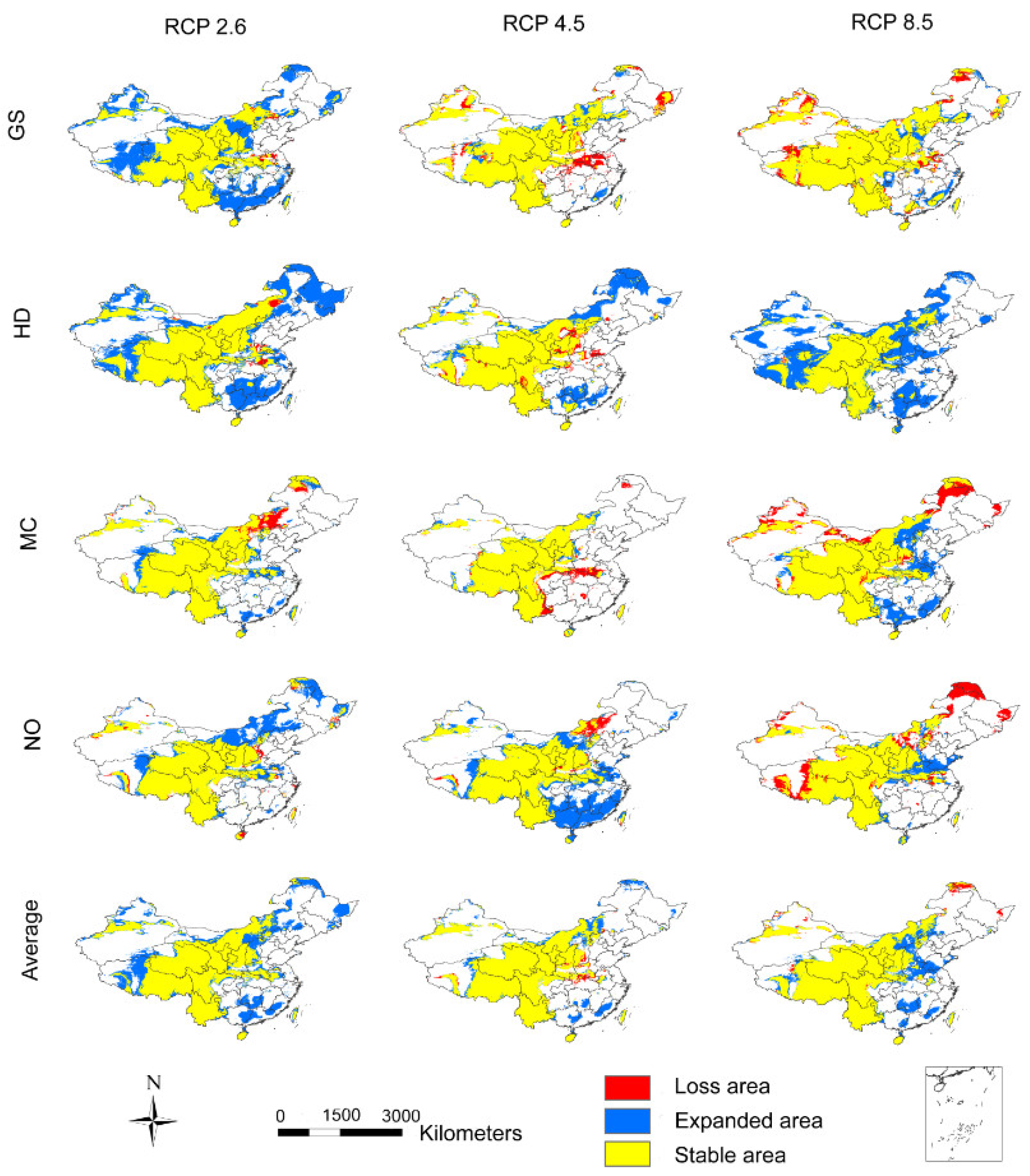
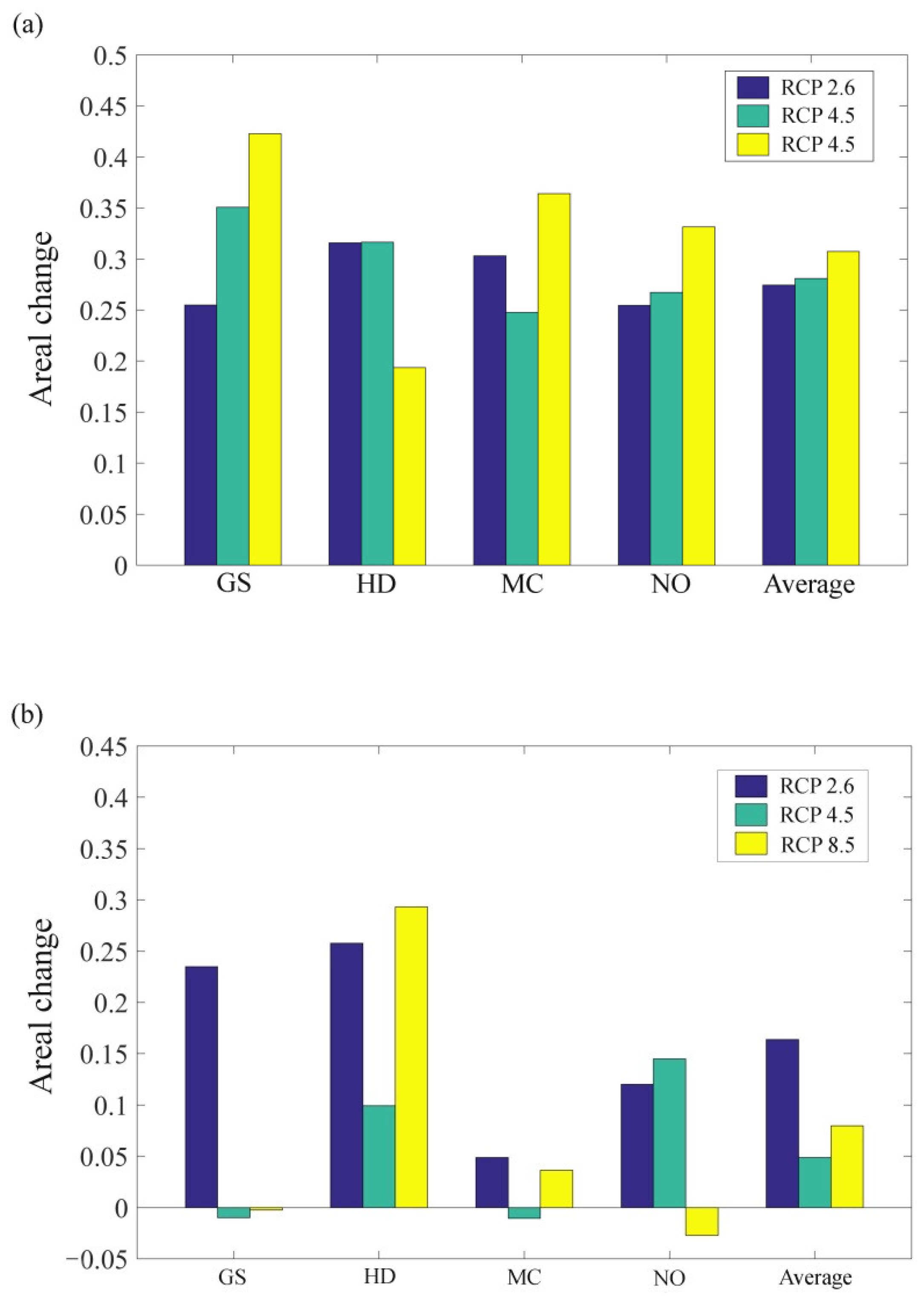
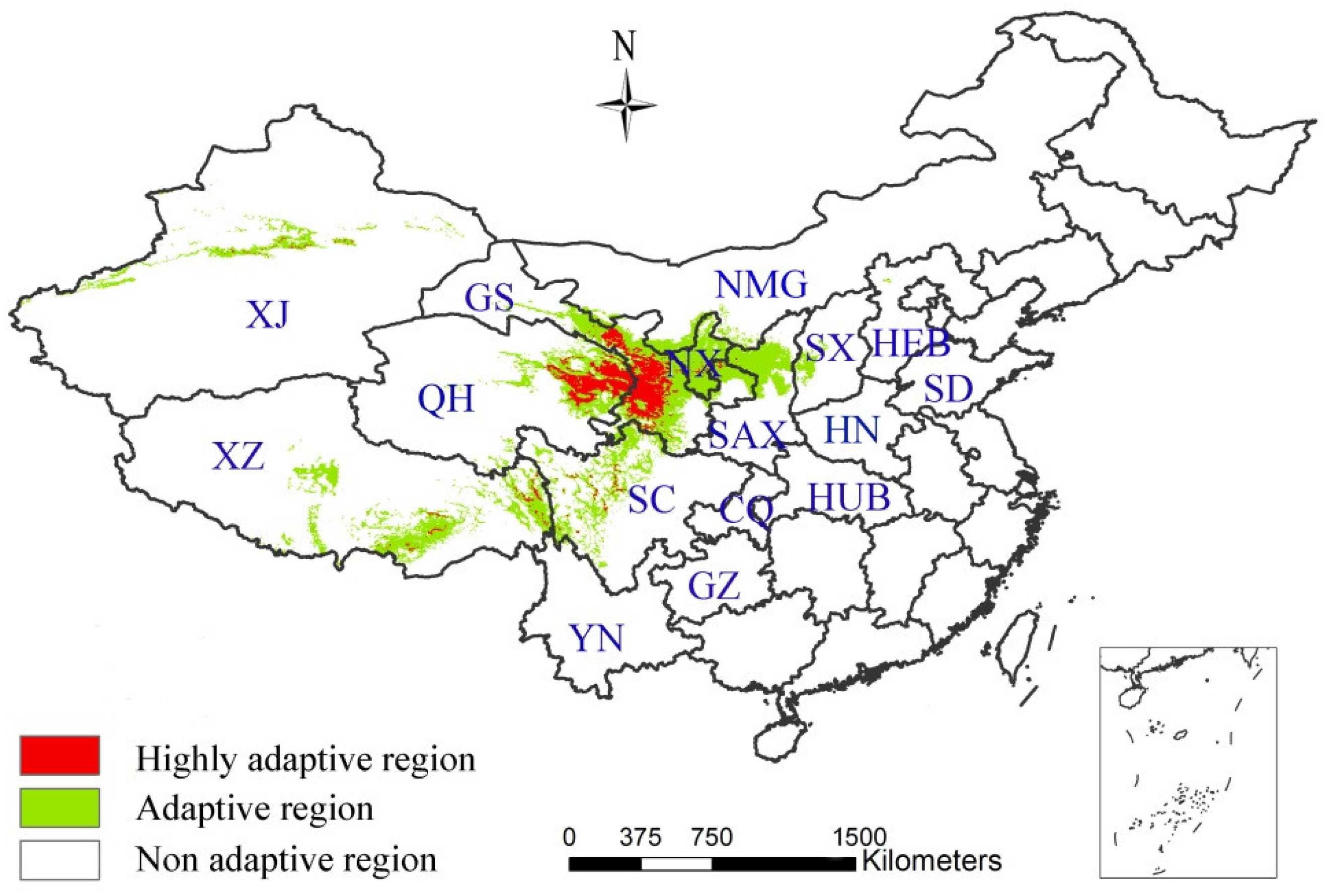
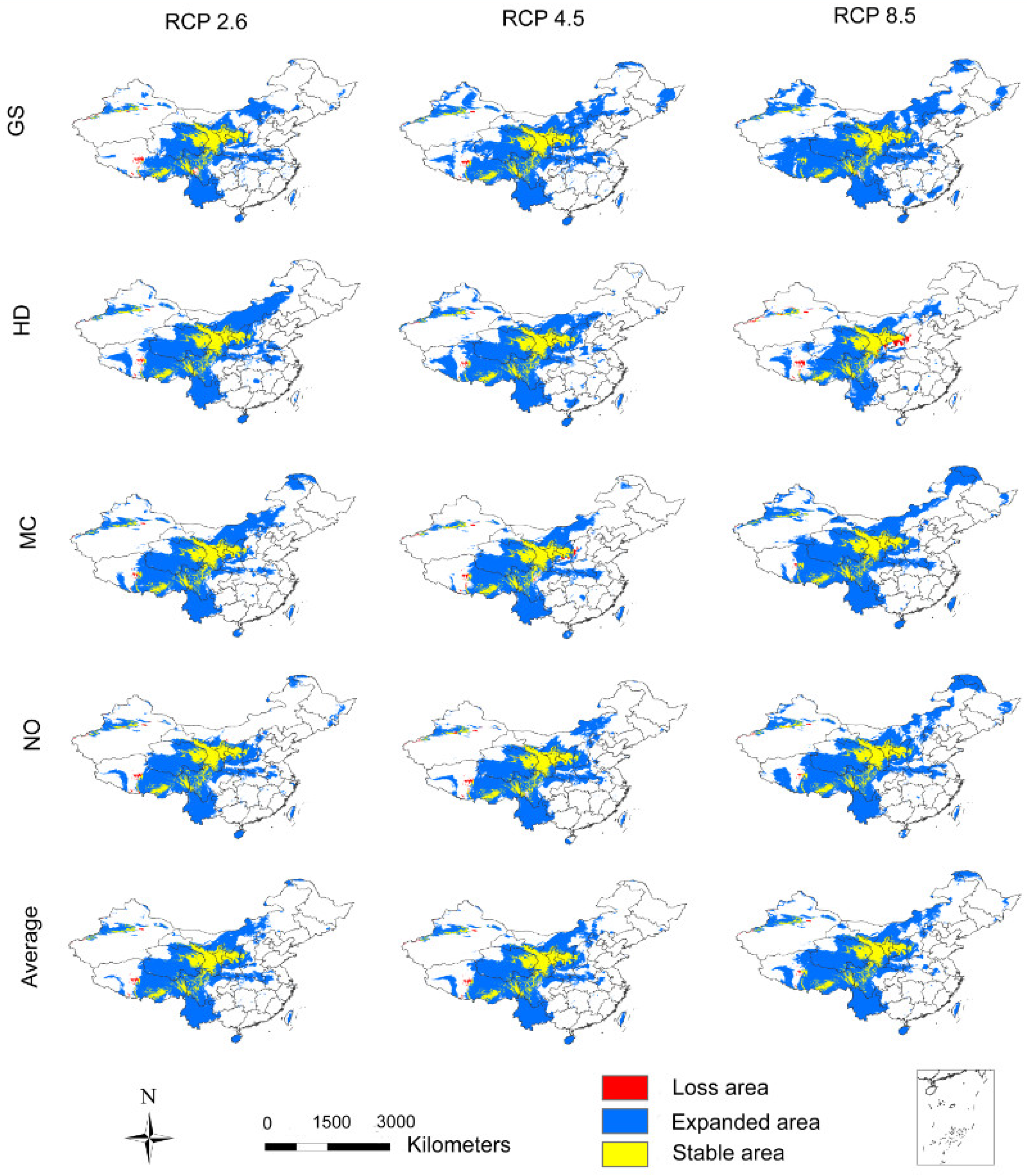
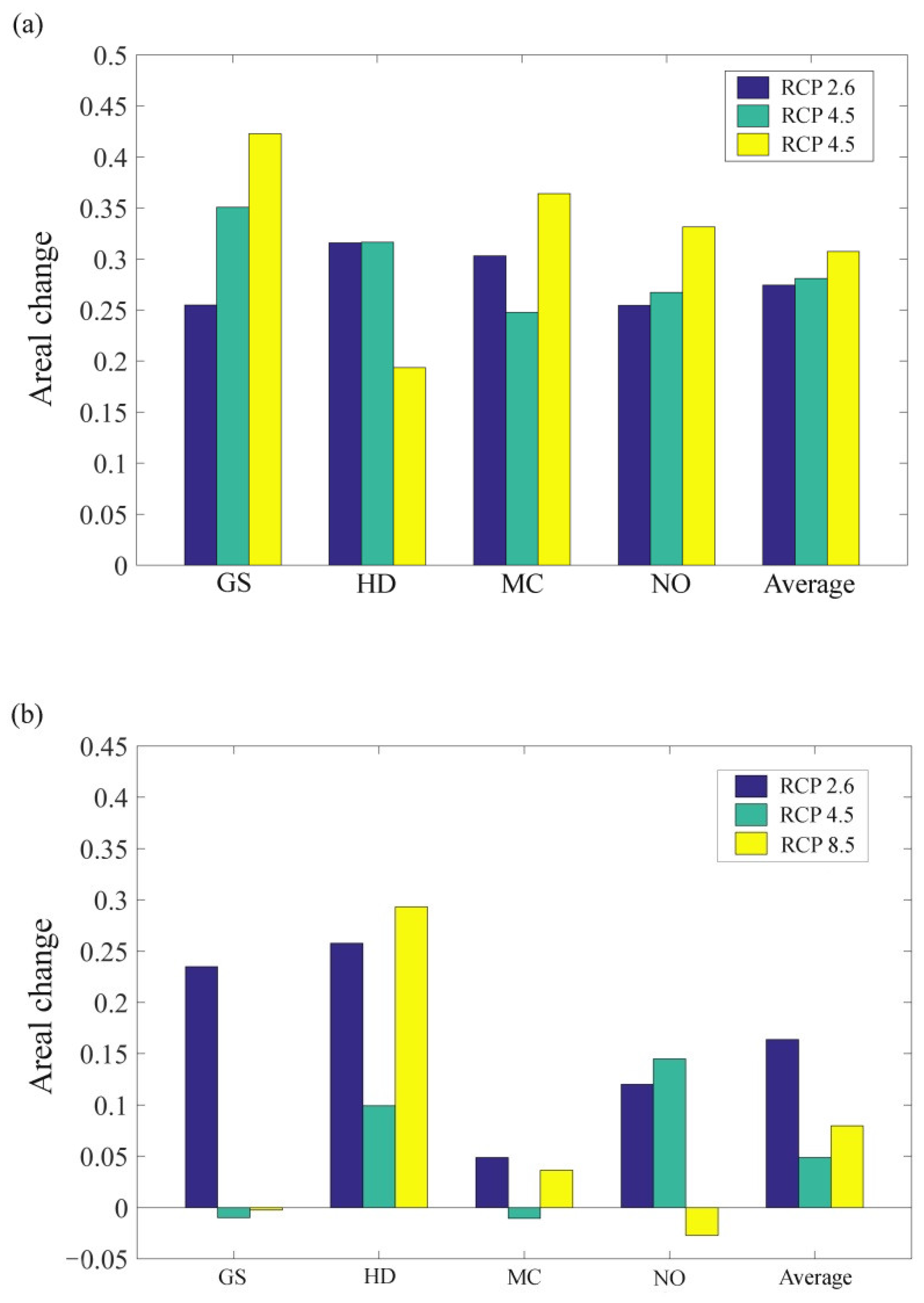
3.2. The Influence of CO2 Emission Scenarios on the Potential Future Distributions of A. inebrians
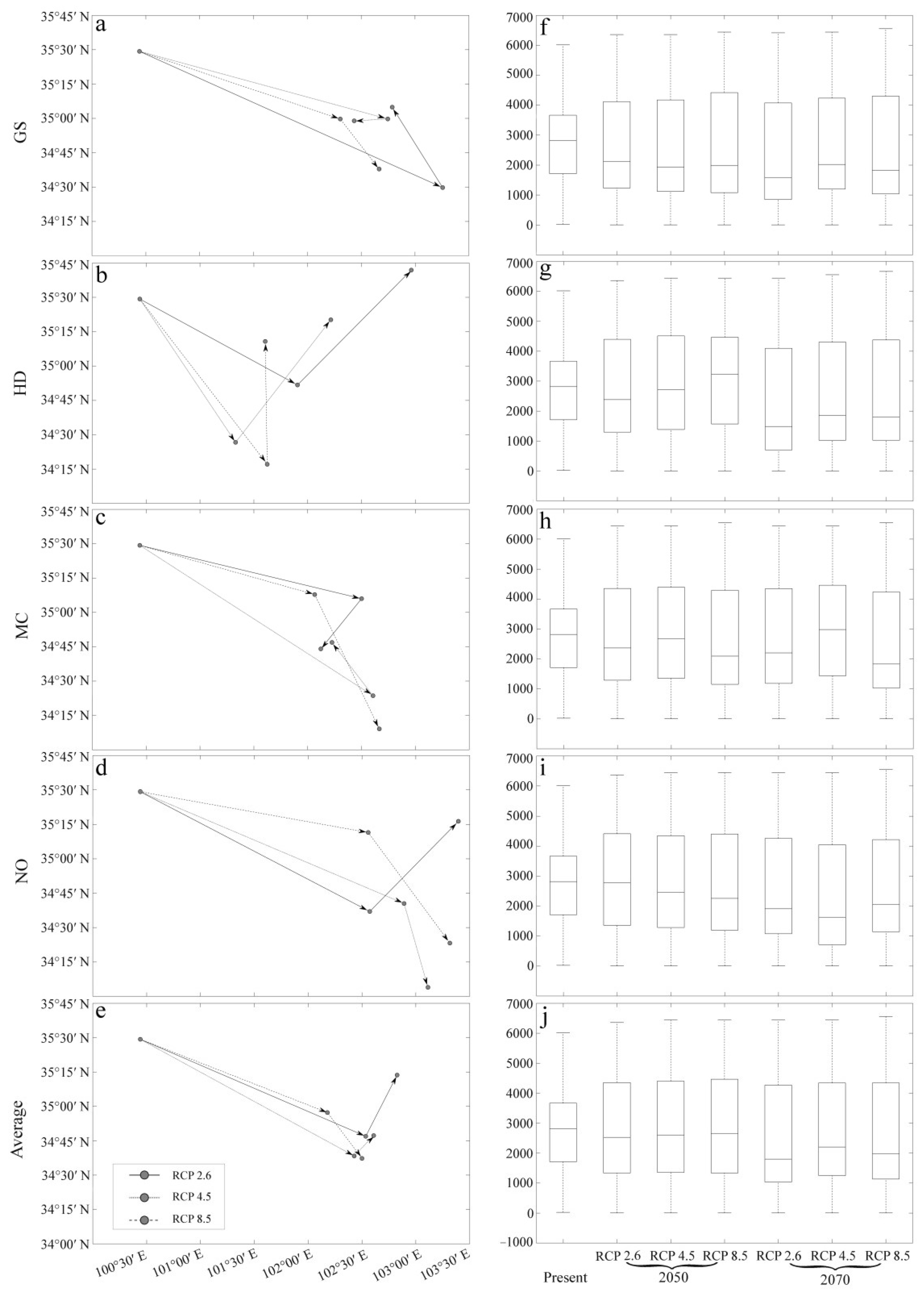
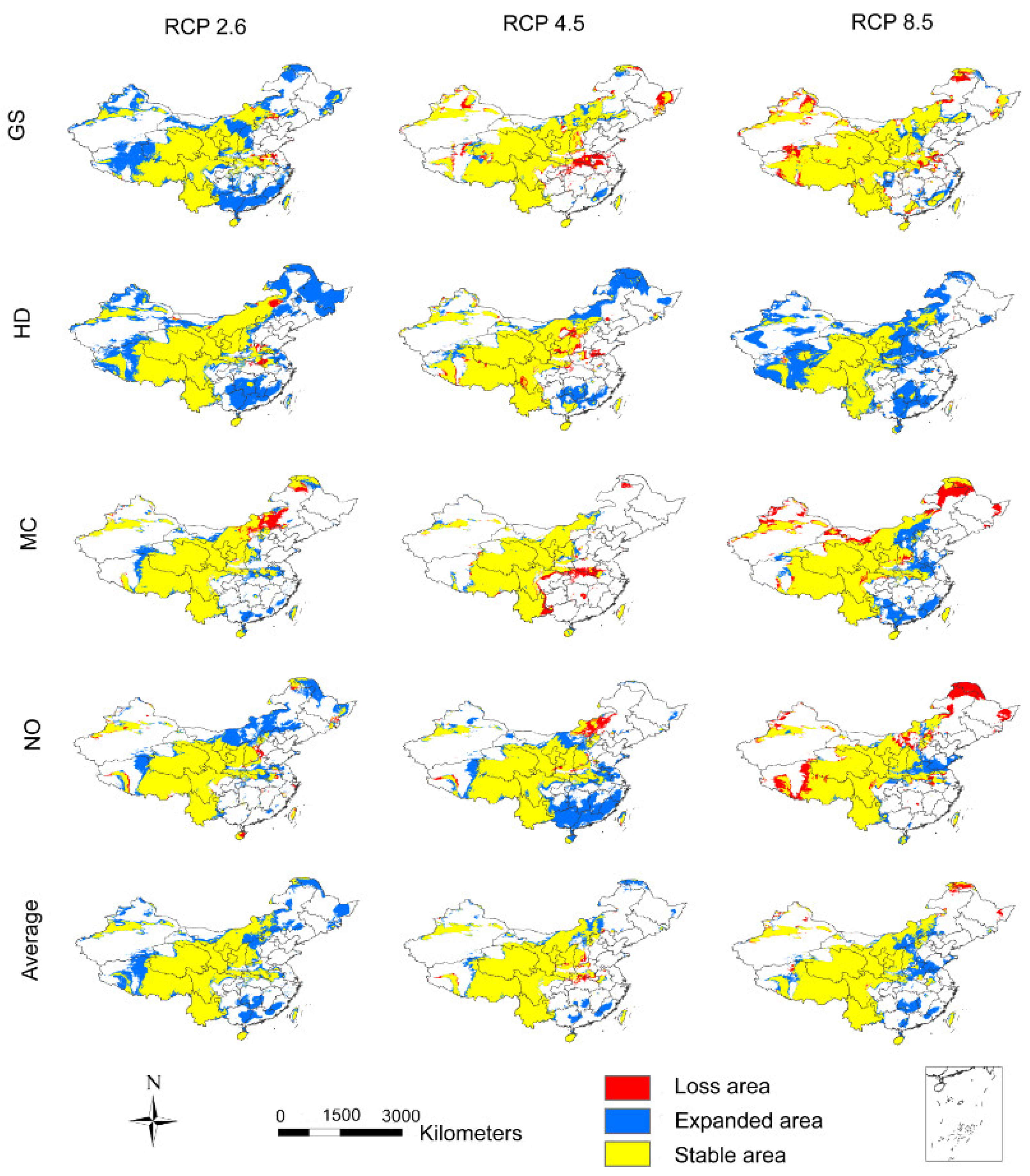
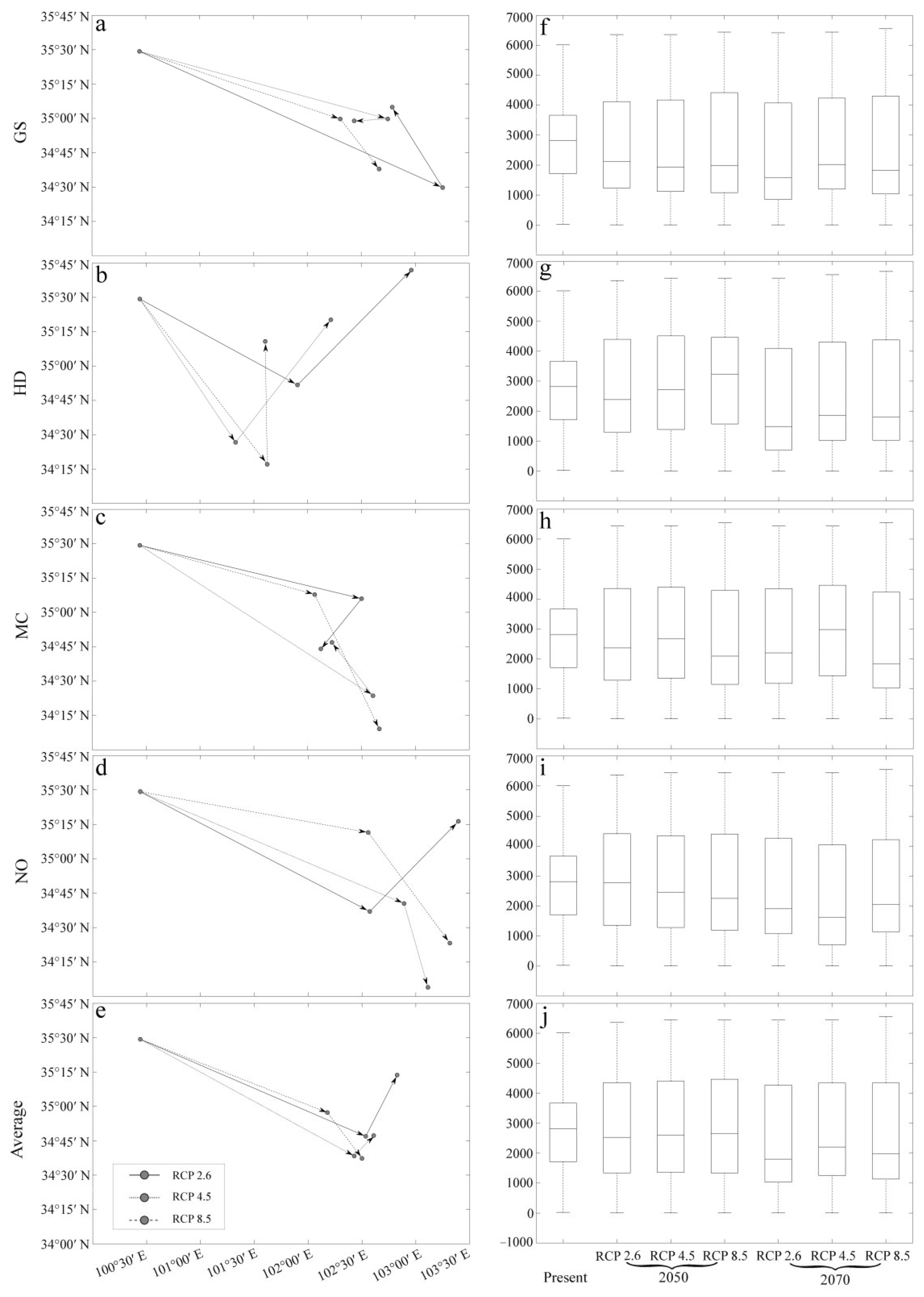
3.3. The Influence of CO2 Emission Scenarios on the Geographical Distribution Centroid and Average Elevation of the Adaptive Regions of A. inebrians
4. Discussions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Houghton, J.T.; Filho, L.G.M.; Callander, B.A.; Harris, N.; Kattenburg, A.; Maskell, K. Climate Change 1995: The Science of Climate Change; Cambridge University Press: Cambridge, UK, 1996; p. 584. [Google Scholar]
- Shahbaz, M.; Shahzad, S.J.H.; Mahalik, M.K. Is globalization detrimental to CO2 emissions in Japan? New threshold analysis. Environ. Model. Assess. 2017, 23, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Folland, C.K.; Boucher, O.; Colman, A.; Parker, D.E. Causes of irregularities in trends of global mean surfacetemperature since the late 19th century. Sci. Adv. 2018, 4, 5297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Mai, T.; Lovell, J. Impact of climate change, adaptation and potential mitigation to Vietnam agriculture. In Handbook of Climate Change Mitigation and Adaptation; Chen, W.-Y., Suzuki, T., Lackner, M., Eds.; Springer: New York, NY, USA, 2016; pp. 1–26. [Google Scholar]
- Bajwa, A.A.; Wang, H.; Chauhan, B.S.; Adkins, S.W. Effect of elevated carbon dioxide concentration on growth, productivity and glyphosate response of parthenium weed (Parthenium hysterophorus L.). Pest Manag. Sci. 2019, 75, 2934–2941. [Google Scholar] [CrossRef] [PubMed]
- IPPC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Harris, R.M.B.; Grose, M.R.; Lee, G.; Bindoff, N.L.; Porfirio, L.L.; Fox-Hughes, P. Climate projections for ecologists. WIREs Clim. Change 2014, 5, 621–637. [Google Scholar] [CrossRef]
- Meinshausen, M.; Smith, S.J.; Calvin, K.; Daniel, J.S.; Kainuma, M.L.T.; Lamarque, J.-F.; Matsumoto, K.; Montzka, S.A.; Raper, S.C.B.; Riahi, K.; et al. The RCP greenhouse gas concentrations and their extensions from 1765 to 2300. Clim. Change 2011, 109, 213–241. [Google Scholar] [CrossRef] [Green Version]
- van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F.; et al. The representative concentration pathways: An overview. Clim. Change 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Koo, K.A.; Park, S.U.; Kong, W.-S.; Hong, S.; Jang, I.; Seo, C. Potential climate change effects on tree distributions in the Korean Peninsula: Understanding model & climate uncertainties. Ecol. Model. 2017, 353, 17–27. [Google Scholar]
- Ernakovich, J.G.; Hopping, K.A.; Berdanier, A.B.; Simpson, R.T.; Kachergis, E.J.; Steltzer, H.; Wallenstein, M.D. Predicted responses of arctic and alpine ecosystems to altered seasonality under climate change. Glob. Change Biol. 2014, 20, 3256–3269. [Google Scholar] [CrossRef]
- Flagmeier, M.; Long, D.G.; Genney, D.R.; Hollingsworth, P.M.; Ross, L.C.; Woodin, S.J. Fifty years of vegetation change in oceanic-montane liverwort-rich heath in Scotland. Plant Ecol. Divers. 2014, 7, 457–470. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Sproull, G.J.; Quigley, M.F.; Sher, A.; González, E. Long-term changes in composition, diversity and distribution patterns in four herbaceous plant communities along an elevational gradient. J. Veg. Sci. 2015, 26, 552–563. [Google Scholar] [CrossRef]
- Walther, G.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.; Fromentin, J.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Change Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- Wróblewska, A.; Mirski, P. From past to future: Impact of climate change on range shifts and genetic diversity patterns of circumboreal plants. Reg. Environ. Change 2018, 18, 409–424. [Google Scholar] [CrossRef] [Green Version]
- Peters, K.; Breitsameter, L.; Gerowitt, B. Impact of climate change on weeds in agriculture: A review. Agron. Sustain. Dev. 2014, 34, 707–721. [Google Scholar] [CrossRef] [Green Version]
- Patterson, D.T. Weeds in a changing climate. Weed Sci. 1995, 43, 685–700. [Google Scholar] [CrossRef]
- McDonald, A.; Riha, S.; DiTommaso, A.; DeGaetano, A. Climate change and the geography of weed damage: Analysis of U.S. maize systems suggests the potential for significant range transformations. Agric. Ecosyst. Environ. 2009, 130, 131–140. [Google Scholar] [CrossRef]
- Ziska, L.H.; Goins, E.W. Elevated atmospheric carbon dioxide and weed populations in glyphosate treated soybean. Crop Sci. 2006, 46, 1354–1359. [Google Scholar] [CrossRef]
- Jabran, K.; Dogan, M.N. High carbon dioxide concentration and elevated temperature impact the growth of weeds but do not change the efficacy of glyphosate. Pest Manag. Sci. 2018, 74, 766–771. [Google Scholar] [CrossRef]
- Saebø, A.; Mortensen, L. Influence of elevated atmospheric CO2 concentration on common weeds in Scandinavian agriculture. Acta Agric. Scand. 1998, 48, 138–143. [Google Scholar]
- Singh, R.P.; Singh, R.K.; Singh, M.K. Impact of climate and carbon dioxide change on weeds and their management-a Review. Indian J. Weed Sci. 2011, 43, 1–11. [Google Scholar]
- Williams, A.L.; Wills, K.E.; Janes, J.K.; Schoor, J.K.V.; Newton, P.C.D.; Hovenden, M.J. Warming and free-air CO2 enrichment alter demographics in four co-occurring grassland species. New Phytol. 2007, 176, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, X.Z.; Li, C.J.; Swoboda, G.A.; Schardl, C.L. Two distinct Epichloë species symbiotic with Achnatherum inebrians, drunken horse grass. Mycologia 2015, 107, 863–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huoman, S. Achnatherum Inebrians and its prevented and cured measures. Pratacult. Sci. 1992, 9, 36–37. [Google Scholar]
- Li, C.-J.; Gao, J.-H.; Ma, B. Seven diseases of drunken horse grass (Achnatherum inebrians) in China. Pratacult. Sci. 2003, 21, 51–53. [Google Scholar]
- Shi, Z. Important Poisonous Plants of China Grassland; China Agricultural Press: Beijing, China, 1997. [Google Scholar]
- Li, X.; Ren, J.; Feng, K.; Lei, T.; Ar, Y.; Zhang, X. Ecological control method of Achnatherum inebrians. Pratacult. Sci. 1996, 5, 14–17. [Google Scholar]
- Vilà, M.; Beaury, E.M.; Blumenthal, D.M.; Bradley, B.A.; Ibanez, I. Understanding the combined impacts of weeds and climate change on crops. Environ. Res. Lett. 2021, 16, 034043. [Google Scholar] [CrossRef]
- Albright, T.P.; Chen, H.; Chen, L.; Guo, Q. The ecological niche and reciprocal prediction of the disjunct distribution of an invasive species: The example of Ailanthus altissima. Biol. Invasions 2010, 12, 2413–2427. [Google Scholar] [CrossRef]
- Medley, K.A. Niche shifts during the global invasion of the Asian tiger mosquito, Aedes albopictus Skuse (Culicidae), revealed by reciprocal distribution models. Glob. Ecol. Biogeogr. 2010, 19, 122–133. [Google Scholar] [CrossRef]
- Peterson, A.T. Predicting the geography of species’ invasions via ecological niche modeling. Q. Rev. Biol. 2003, 78, 419–433. [Google Scholar] [CrossRef] [Green Version]
- Olivera, L.; Minghetti, E.; Montemayor, S.I. Ecological niche modeling (ENM) of Leptoglossus clypealis a new potential global invader: Following in the footsteps of Leptoglossus occidentalis? Bull. Entomol. Res. 2021, 111, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.J.; Gilbert, C.C.; Kamilar, J.M. Ecological niche modeling of the genus Papio. Am. J. Phys. Anthropol. 2018, 166, 812–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolanowska, M.; Rewicz, A.; Baranow, P. Ecological niche modeling of the pantropical orchid Polystachya concreta (Orchidaceae) and its response to climate change. Sci. Rep. 2020, 10, 14801. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G. Species’ distribution modeling for conservation educators and practitioners. Lesson Conserv. 2010, 3, 54–89. [Google Scholar]
- Zhu, G.; Liu, G.; Bu, W.; Gao, Y. Ecological niche modeling and its applications in biodiversity conservation. Biodivers. Sci. 2013, 21, 90–98. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Xu, Q.; Luo, Y. A maximum entropy model predicts the potential geographic distribution of Sirex noctilio. Forests 2020, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Gibson, L.; Barrett, B.; Burbidge, A. Dealing with uncertain absences in habitat modelling: A case study of a rare ground-dwelling parrot. Divers. Distrib. 2007, 13, 704–713. [Google Scholar] [CrossRef]
- Peterson, A.T.; Pape, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of GARP and Maxent. Ecography 2007, 30, 550–560. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Bagchi, R.; Crosby, M.; Huntley, B.; Hole, D.G.; Butchart, S.H.M.; Collingham, Y.; Kalra, M.; Rajkumar, J.; Rahmani, A.; Pandey, M.; et al. Evaluating the effectiveness of conservation site networks under climate change: Accounting for uncertainty. Glob. Change Biol. 2013, 19, 1236–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuiller, W. Patterns and uncertainties of species’ range shifts under climate change. Glob. Change Biol. 2004, 10, 2020–2027. [Google Scholar] [CrossRef]
- Baker, D.J.; Hartley, A.J.; Butchart, S.H.; Willis, S.G. Choice of baseline climate data impacts projected species’ responses to climate change. Glob. Change Biol. 2016, 22, 2392–2404. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Wang, G.; Innes, J.; Nitschke, C.; Kang, H. Climatic niche models and their consensus projections for future climates for four major forest tree species in the Asia–Pacific region. For. Ecol. Manag. 2016, 360, 357–366. [Google Scholar] [CrossRef]
- Hortal, J.; Jiménez-Valverde, A.; Gómez, J.F.; Lobo, J.M.; Baselga, A.J.O. Historical bias in biodiversity inventories affects the observed environmental niche of the species. Oikos 2010, 117, 847–858. [Google Scholar] [CrossRef]
- Kang, B.H.; Shim, S.I.; Lee, S.G.; Shin, H.W. Physiological and ecological studies on the seed dormancy of dominant weed species in Korea. Korean J. Environ. Agric. 1993, 12, 193–207. [Google Scholar]
- Guan, X.; Ramaswamy, H.; Zhang, B.; Lin, B.; Wang, S. Influence of moisture content, temperature and heating rate on germination rate of watermelon seeds. Sci. Hortic. 2020, 272, 109528. [Google Scholar] [CrossRef]
- Tian, Z.-H.; Shen, G.-H. Advance on regulation of seed dormancy and germination of weeds. Acta Agric. Shanghai 2015, 31, 137–141. [Google Scholar]
- Yu, X.-J.; Chen, B.-J.; Shi, S.-L.; Wei, G.-B.; Man, Y.-R.; Ma, Y.-L. Effect of temperature and moisture condition on seed germination of Achnatherum Inebriants (Hance) Keng. Acta Agrestia Sin. 2009, 17, 218–221. [Google Scholar]
- Cowie, B.W.; Venter, N.; Witkowski, E.; Byrne, M.J. Implications of elevated carbon dioxide on the susceptibility of the globally invasive weed, parthenium hysterophorus, to glyphosate herbicide. Pest Manag. Sci. 2020, 76, 2324–2332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhu, Y.; Li, Y.; Ni, Y. Effects of atmospheric CO2 doubling on the climate. Sci. Meteorol. Sin. 1994, 14, 16–22. [Google Scholar]
- Jentsch, A.; Kreyling, J.; Boettcher-Treschkow, J.; Beierkuhnlein, C. Beyond gradual warming: Extreme weather events alter flower phenology of European grassland and heath species. Glob. Change Biol. 2009, 15, 837–849. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.K.; Ohlemuller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Kräuchi, N.; Gao, S.P. Global warming: Can existing reserves really preserve current levels of biological diversity? J. Integr. Plant Biol. 2006, 48, 255–259. [Google Scholar] [CrossRef]
- Midgley, G.F.; Hannah, L.; Millar, D.; Thuiller, W.; Booth, A. Developing regional and species-level assessments of climate change impacts on biodiversity in the Cape Floristic Region. Biol. Conserv. 2003, 112, 87–97. [Google Scholar] [CrossRef]
- Wang, T.; Campbell, E.M.; O’Neill, G.A.; Aitken, S.N. Projecting future distributions of ecosystem climate niches: Uncertainties and management applications. For. Ecol. Manag. 2012, 279, 128–140. [Google Scholar] [CrossRef]
- Chen, M.; Lin, E. Global greenhouse gas emission mitigation under representative concentration pathways scenarios and challenges to China. Adv. Clim. Change Res. 2010, 80, 436–442. [Google Scholar]
- Baselga, A.; Araújo, M. Individualistic vs. community modelling of species distributions under climate change. Ecography 2010, 32, 55–65. [Google Scholar] [CrossRef]
- Phillips, S.J.; Miroslav, D.; Jane, E.; Graham, C.H.; Anthony, L.; John, L.; Simon, F. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Stockwell, D.R.B. Improving ecological niche models by data mining large environmental datasets for surrogate models. Ecol. Model. 2005, 192, 188–196. [Google Scholar] [CrossRef] [Green Version]
- Levsen, N.D.; Tiffin, P.; Olson, M.S. Pleistocene speciation in the genus Populus (salicaceae). Syst. Biol. 2012, 61, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sax, D.F.; Stachowicz, J.J.; Brown, J.H.; Bruno, J.F.; Dawson, M.N.; Gaines, S.D.; Grosberg, R.K.; Hastings, A.; Holt, R.D.; Mayfield, M.M. Ecological and evolutionary insights from species invasions. Trends Ecol. Evol. 2007, 22, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Keith, D.A.; HResit, A.A.; Wilfried, T.; Midgley, G.F.; Pearson, R.G.; Phillips, S.J.; Regan, H.M.; Araújo, M.B.; Rebelo, T.G. Predicting extinction risks under climate change: Coupling stochastic population models with dynamic bioclimatic habitat models. Biol. Lett. 2008, 4, 560–563. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.; Kumar, L.; Reid, N. Impacts of climate change and land-use on the potential distribution of an invasive weed: A case study of Lantana camara in Australia. Weed Res. 2012, 52, 391–401. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioclimatic Variables | Meaning of Variables |
|---|---|
| Bio02 | Mean Diurnal Range (Mean of monthly (max temp—min temp)) |
| Bio03 | Isothermality (Mean Diurnal Range/Temperature Annual Range) (×100) |
| Bio06 | Min Temperature of Coldest Month |
| Bio10 | Mean Temperature of Warmest Quarter |
| Bio15 | Precipitation Seasonality (Coefficient of Variation) |
| Bio16 | Precipitation of Wettest Quarter |
| Bio19 | Precipitation of Coldest Quarter |
| GCM | Code | Institution |
|---|---|---|
| GISS-E2-R | GS | NASA Goddard Institute for Space Studies |
| HadGEM2-AO | HD | National Institute of Meteorological Research/Korea Meteorological Administration |
| MIROC5 | MC | Atmosphere and Ocean Research Institute (The University of Tokyo), National Institute for Environmental Studies, and Japan Agency for Marine-Earth Science and Technology |
| NorESM1-M | NO | Norwegian Climate Centre |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.-M.; Jin, L.; Huang, L.; Wang, W.-T. The Future Climate under Different CO2 Emission Scenarios Significantly Influences the Potential Distribution of Achnatherum inebrians in China. Sustainability 2022, 14, 4806. https://doi.org/10.3390/su14084806
Jiang J-M, Jin L, Huang L, Wang W-T. The Future Climate under Different CO2 Emission Scenarios Significantly Influences the Potential Distribution of Achnatherum inebrians in China. Sustainability. 2022; 14(8):4806. https://doi.org/10.3390/su14084806
Chicago/Turabian StyleJiang, Jia-Min, Lei Jin, Lei Huang, and Wen-Ting Wang. 2022. "The Future Climate under Different CO2 Emission Scenarios Significantly Influences the Potential Distribution of Achnatherum inebrians in China" Sustainability 14, no. 8: 4806. https://doi.org/10.3390/su14084806
APA StyleJiang, J.-M., Jin, L., Huang, L., & Wang, W.-T. (2022). The Future Climate under Different CO2 Emission Scenarios Significantly Influences the Potential Distribution of Achnatherum inebrians in China. Sustainability, 14(8), 4806. https://doi.org/10.3390/su14084806