
Plant Diversity Increased Arthropod Diversity and Crop Yield in Traditional Agroforestry Systems but Has No Effect on Herbivory


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Measuring Plant, Arthropod Diversity, and Crop Yield
2.3. Statistical Analysis
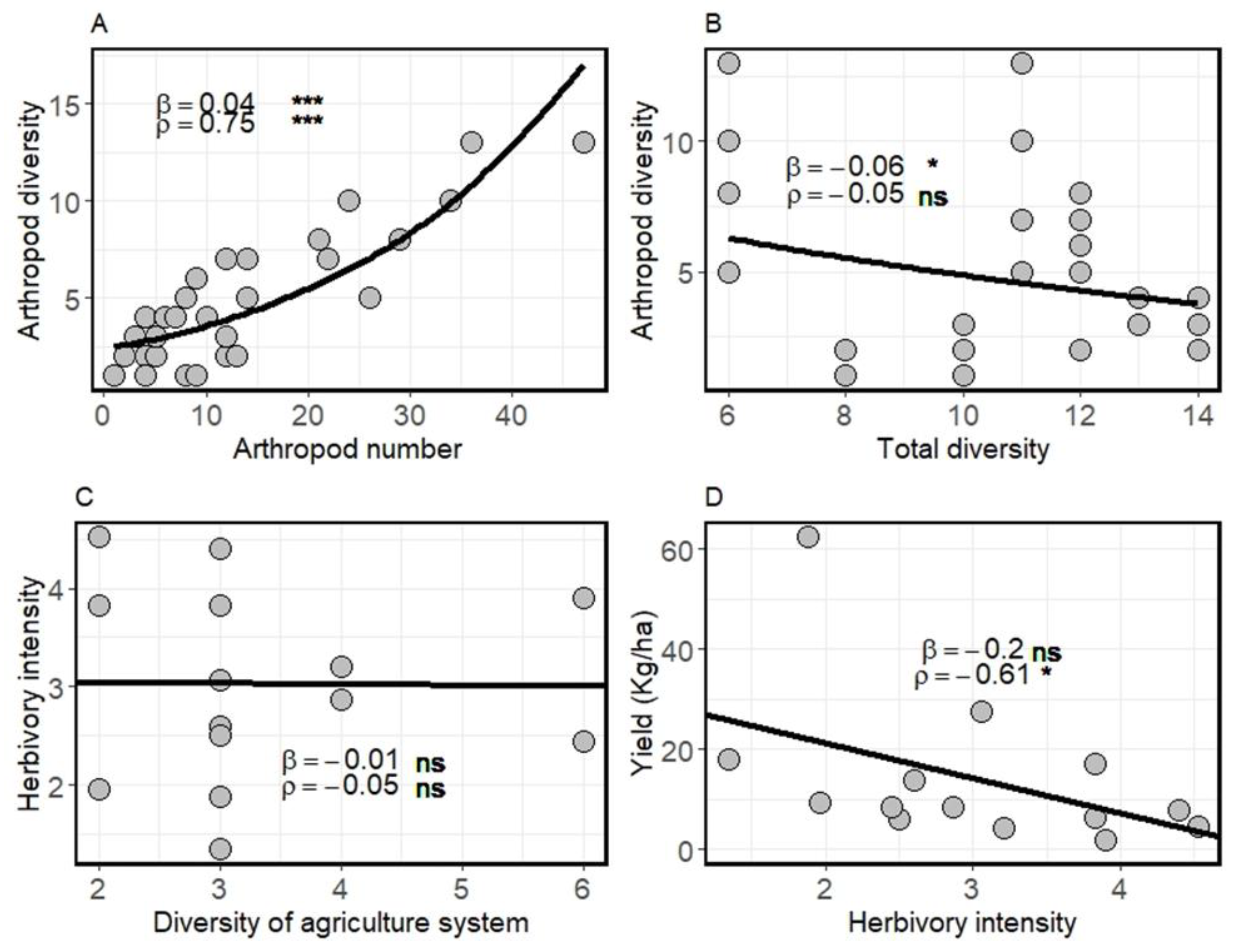
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, M.B. Ecological approaches and the development of “truly integrated” pest management. Proc. Natl. Acad. Sci. USA 1999, 96, 5944–5951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Research Council. Toward Sustainable Agricultural Systems in the 21st Century; The National Academies Press: Washington, DC, USA, 2010; ISBN 978-0-309-14896-2. [Google Scholar]
- Wood, S.A. Nutritional Functional Trait Diversity of Crops in South-Eastern Senegal. J. Appl. Ecol. 2018, 55, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, L.V.; Coolsaet, B.; Martin, A.; Mertz, O.; Pascual, U.; Corbera, E.; Dawson, N.; Fisher, J.A.; Franks, P.; Ryan, C.M. Social-ecological outcomes of agricultural intensification. Nat. Sustain. 2018, 1, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Jayne, T.; Snapp, S.; Place, F.; Sitko, N. Sustainable agricultural intensification in an era of rural transformation in Africa. Glob. Food Secur. 2019, 20, 105–113. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Database of the Food and Agricultural Organization (FAO) of the United Nations; FAO: Rome, Italy, 2011. [Google Scholar]
- Monfreda, C.; Ramankutty, N.; Foley, J.A. Farming the planet: 2. Geographic distribution of crop areas, yields, physiological types, and net primary production in the year 2000. Glob. Biogeochem. Cycles 2008, 22. [Google Scholar] [CrossRef]
- Ramankutty, N.; Evan, A.T.; Monfreda, C.; Foley, J.A. Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Glob. Biogeochem. Cycles 2008, 22. [Google Scholar] [CrossRef]
- Asner, G.P.; Powell, G.V.N.; Mascaro, J.; Knapp, D.E.; Clark, J.K.; Jacobson, J.; Kennedy-Bowdoin, T.; Balaji, A.; Paez-Acosta, G.; Victoria, E.; et al. High-Resolution Forest Carbon Stocks and Emissions in the Amazon. Proc. Natl. Acad. Sci. USA 2010, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, H.K.; Ruesch, A.S.; Achard, F.; Clayton, M.K.; Holmgren, P.; Ramankutty, N.; Foley, J.A. Tropical Forests Were the Primary Sources of New Agricultural Land in the 1980s and 1990s. Proc. Natl. Acad. Sci. USA 2010, 107, 16732–16737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamorro, L.; Masalles, R.M.; Sans, F.X. Arable weed decline in Northeast Spain: Does organic farming recover functional biodiversity? Agric. Ecosyst. Environ. 2016, 223, 1–9. [Google Scholar] [CrossRef]
- Hole, D.G.; Perkins, A.J.; Wilson, J.; Alexander, I.H.; Grice, P.V.; Evans, A.D. Does organic farming benefit biodiversity? Biol. Conserv. 2005, 122, 113–130. [Google Scholar] [CrossRef]
- Humbert, J.-Y.; Dwyer, J.; Andrey, A.; Arlettaz, R. Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: A systematic review. Glob. Change Biol. 2015, 22, 110–120. [Google Scholar] [CrossRef]
- Melts, I.; Lanno, K.; Sammul, M.; Uchida, K.; Heinsoo, K.K.T. Toward a mechanistic understanding of the effects of nitrogen and phosphorus additions on grassland diversity. J. Appl. Ecol. 2018, 32, 65–72. [Google Scholar]
- Puig-Montserrat, X.; Stefanescu, C.; Torre, I.; Palet, J.; Fàbregas, E.; Dantart, J.; Arrizabalaga, A.; Flaquer, C. Effects of organic and conventional crop management on vineyard biodiversity. Agric. Ecosyst. Environ. 2017, 243, 19–26. [Google Scholar] [CrossRef]
- Wilson, J.D.; Morris, A.J.; Arroyo, B.E.; Clark, S.C.; Bradbury, R.B. A review of the abundance and diversity of invertebrate and plant foods of granivorous birds in northern Europe in relation to agricultural change. Agric. Ecosyst. Environ. 1999, 75, 13–30. [Google Scholar] [CrossRef]
- Zimmerer, K.S. The compatibility of agricultural intensification in a global hotspot of smallholder agrobiodiversity (Bolivia). Proc. Natl. Acad. Sci. USA 2013, 110, 2769–2774. [Google Scholar] [CrossRef] [Green Version]
- Mikos, V.; Pulleman, M. European Policy Review: Functional agrobiodiversity supporting sustainable agriculture. J. Nat. Conserv. 2014, 22, 193–194. [Google Scholar] [CrossRef]
- Mertz, O.; Mertens, C.F. Land Sparing and Land Sharing Policies in Developing Countries—Drivers and Linkages to Scientific Debates. World Dev. 2017, 98, 523–535. [Google Scholar] [CrossRef]
- Grass, I.; Loos, J.; Baensch, S.; Batáry, P.; Librán-Embid, F.; Ficiciyan, A.; Klaus, F.; Riechers, M.; Rosa, J.; Tiede, J.; et al. Land-sharing/-sparing connectivity landscapes for ecosystem services and biodiversity conservation. People Nat. 2019, 1, 262–272. [Google Scholar] [CrossRef]
- Latini, A.O.; Silva, D.P.; Souza, F.M.L.; Ferreira, M.C.; de Moura, M.S.; Suarez, N.F. Reconciling coffee productivity and natural vegetation conservation in an agroecosystem landscape in Brazil. J. Nat. Conserv. 2020, 57, 125902. [Google Scholar] [CrossRef]
- Carney, J.A. Subsistence in the Plantationocene: Dooryard gardens, agrobiodiversity, and the subaltern economies of slavery. J. Peasant. Stud. 2020, 48, 1075–1099. [Google Scholar] [CrossRef]
- Kremen, C.; Miles, A. Ecosystem Services in Biologically Diversified versus Conventional Farming Systems: Benefits, Externalities, and Trade-Offs. Ecol. Soc. 2012, 17, 40. [Google Scholar] [CrossRef]
- Finckh, M.R.; Gacek, E.S.; Goyeau, H.; Lannou, C.; Mundt, C.C.; Munk, L.; Merz, U.; Nadziak, J.; Newton, A.; De Vallavieille-Pope, C.; et al. Cereal variety and species mixtures in practice, with emphasis on disease resistance. Science 2000, 20, 813–837. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, R.; Vasconcelos, H.L.; Groc, S.; Camacho, G.P.; Frizzo, T.L.M. The importance of remnants of natural vegetation for maintaining ant diversity in Brazilian agricultural landscapes. Biodivers. Conserv. 2013, 22, 983–997. [Google Scholar] [CrossRef]
- Theron, K.J.; Gaigher, R.; Pryke, J.S.; Samways, M.J. Abandoned fields and high plant diversity support high spider diversity within an agricultural mosaic in a biodiversity hotspot. Biodivers. Conserv. 2020, 29, 3757–3782. [Google Scholar] [CrossRef]
- Arnold, S.E.J.; Elisante, F.; Mkenda, P.A.; Tembo, Y.L.B.; Ndakidemi, P.A.; Gurr, G.M.; Darbyshire, I.A.; Belmain, S.R.; Stevenson, P.C. Beneficial insects are associated with botanically rich margins with trees on small farms. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Galloway, A.D.; Seymour, C.L.; Gaigher, R.; Pryke, J.S. Organic farming promotes arthropod predators, but this depends on neighbouring patches of natural vegetation. Agric. Ecosyst. Environ. 2021, 310, 107295. [Google Scholar] [CrossRef]
- Letourneau, D.K.; Armbrecht, I.; Rivera, B.S.; Lerma, J.M.; Carmona, E.J.; Daza, M.C.; Trujillo, A.R.; Escobar, S.; Riviera, L.; Galindo, V.; et al. Does plant diversity benefit agroecosystems? A synthetic review. Ecol. Appl. 2011, 21, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Carvalheiro, L.G.; Veldtman, R.; Shenkute, A.G.; Tesfay, G.B.; Pirk, C.; Donaldson, J.; Nicolson, S. Natural and within-farmland biodiversity enhances crop productivity. Ecol. Lett. 2011, 14, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Guyot, V.; Castagneyrol, B.; Vialatte, A.; Deconchat, M.; Jactel, H. Tree diversity reduces pest damage in mature forests across Europe. Biol. Lett. 2016, 12, 20151037. [Google Scholar] [CrossRef] [Green Version]
- Hajjar, R.; Jarvis, D.I.; Gemmill-Herren, B. The utility of crop genetic diversity in maintaining ecosystem services. Agric. Ecosyst. Environ. 2008, 123, 261–270. [Google Scholar] [CrossRef]
- Mundt, C.C. Use of multiline cultivars and cultivar mixtures for disease management. Annu. Rev. Phytopathol. 2002, 40, 381–410. [Google Scholar] [CrossRef] [Green Version]
- Ewel, J.J. Natural systems as models for the design of sustainable systems of land use. Agrofor. Syst. 1999, 45, 1–21. [Google Scholar] [CrossRef]
- Jackson, W. Natural systems agriculture: A truly radical alternative. Agric. Ecosyst. Environ. 2002, 88, 111–117. [Google Scholar] [CrossRef]
- Lewis, W.J.; van Lenteren, J.C.; Phatak, S.C.; Tumlinson, J.H. A total system approach to sustainable pest management. Proc. Natl. Acad. Sci. USA 1997, 94, 12243–12248. [Google Scholar] [CrossRef] [Green Version]
- Kardol, P.; Fanin, N.; Wardle, D. Long-term effects of species loss on community properties across contrasting ecosystems. Nature 2018, 557, 710–713. [Google Scholar] [CrossRef] [PubMed]
- SSpehn, E.M.; Hector, A.; Joshi, J.; Scherer-Lorenzen, M.; Schmid, B.; Bazeley-White, E.; Beierkuhnlein, C.; Caldeira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; et al. Ecosystem effects of biodiversity manipulations in european grasslands. Ecol. Monogr. 2005, 75, 37–63. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Reich, P.B.; Knops, J.; Mielke, T.; Wedin, D.; Lehman, C. Diversity and Productivity in a Long-Term Grassland Experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shipley, B. Cause and Correlation in Biology: A User’s Guide to Path Analysis, Structural Equations and Causal Inference; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Grace, J.B. Structural Equation Modeling and Natural Systems; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar] [CrossRef]
- Boetzl, F.A.; Krimmer, E.; Krauss, J.; Steffan-Dewenter, I. Agri-environmental schemes promote ground-dwelling predators in adjacent oilseed rape fields: Diversity, species traits and distance-decay functions. J. Appl. Ecol. 2018, 56, 10–20. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Beyond Fragmentation: The Continuum Model for Fauna Research and Conservation in Human-Modified Landscapes. Oikos 2006, 112, 473–480. [Google Scholar] [CrossRef]
- ASECNA. Données Climatiques, Station de Natitingou, Bénin; ASECNA: Cotonou, Benin, 2010. [Google Scholar]
- Houehanou, T.D.; Assede, E.P.S.; Habiyaremye, F.M.; Sogbohossou, E.A.; Kouton, M.; Agbani, P.; Yaoitcha, A.S.; Gbeffe, A.K.; Houinato, M.R.B. La Réserve de La Biosphère de La Pendjari (Bénin) Guide et Lexique Pour Le Suivi Des Parcours Naturels: Habitats, Faune et Flore; Institut Royal des Sciences Naturelles de Belgique (IRSNB): Bruxelle, Belgique, 2017. [Google Scholar]
- Nago, A.G.S.; Amahowe, I.; Zannou, O.; Houessou, L.; Ahononga, F.; N’Séra, P.; Sinsin, B.; Kouton, M.; Kidjo, F.; Sahilou, S. Diversité, abondance et densité des populations de faune dans la Réserve de Biosphère de la Pendjari (Nord Bénin). Ann. UP Série Sci. Nat. Agron. 2016, 6, 10–25. [Google Scholar]
- Faure, P.; Volkoff, B. Some Factors Affecting Regional Differentiation of the Soils in the Republic of Benin (West Africa). Catena 1998, 32, 281–306. [Google Scholar] [CrossRef]
- M’Woueni, D.; Gaoue, O.G.; Balagueman, R.O.; Biaou, H.S.S.; Natta, A.K. Road mediated spatio-temporal tree decline in traditional agroforests in an African biosphere reserve. Glob. Ecol. Conserv. 2019, 20, e00796. [Google Scholar] [CrossRef]
- Téhou, A.C.; Amahowé, O.I.; Mensah, G.A. Importance des aires protégées du complexe des parcs du W, d’Arly et de la Pendjari pour les communautés riveraines et le grand public en République du Bénin. Ann. Sci. Agron. 2012, 16, 235–249. [Google Scholar]
- Gaoue, O.G. Determinant Factors for the Integrated Management of Pendjari Hunting Reserve Northem Benin. Engineer Thesis, University of Abomey-Calavi, Abomey-Calavi, Benin, 2000. [Google Scholar]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Lundin, O.; Ward, K.L.; Williams, N.M. Identifying native plants for coordinated habitat management of arthropod pollinators, herbivores and natural enemies. J. Appl. Ecol. 2019, 56, 665–676. [Google Scholar] [CrossRef]
- Arnold, J.E.; Egerer, M.; Daane, K.M. Local and Landscape Effects to Biological Controls in Urban Agriculture—A Review. Insects 2019, 10, 215. [Google Scholar] [CrossRef] [Green Version]
- Egerer, M.; Liere, H.; Lucatero, A.; Philpott, S.M. Plant Damage in Urban Agroecosystems Varies with Local and Landscape Factors. Ecosphere 2020, 11, e03074. [Google Scholar] [CrossRef] [Green Version]
- Barratt, B.I.P.; Derraik, J.G.B.; Rufaut, C.G.; Goodman, A.J.; Dickinson, K.J.M. Morphospecies as a substitute for Coleoptera species identification, and the value of experience in improving accuracy. J. R. Soc. New Zealand 2003, 33, 583–590. [Google Scholar] [CrossRef]
- Biaou, H.S.S.; Natta, A.; Dicko, A. Typologie des systèmes agroforestiers et leurs impacts sur la satisfaction des besoins des populations rurales au Bénin. Bull. Rech. Agron. Bénin 2016, 43–56. Available online: ://www.slire.net/download/2371/article_5complet_brab_n_sp_cial_esr_d_cembre_2016_biaou_et_al_typologie_des_syst_mes_agroforestiers.pdf (accessed on 24 December 2021).
- Guillou, M.; Rogel-Gaillard, C. Agricultures et biodiversité: Des futurs liés. Ann. Mines Responsab. Environ. 2012, 68, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Traore, M.; Nacro, H.; Doamba, W.; Tabo, R.; Nikiema, A. Effets de doses variées du tourteau de Jatropha curcas sur la productivité du mil (variété HKP) en condition pluviale en Afrique de l’Ouest. Tropicultura 2015, 33, 19–25. [Google Scholar]
- Canty, A.J. Resampling Methods in {R}: The boot Package. R News 2002, 2, 2–7. [Google Scholar]
- Rosseel, Y. Iavaan: An R package for structural equation modeling. J. Stat. Soft. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Bollen, K.A.; Pearl, J. Eight Myths About Causality and Structural Equation Models. In Handbook of Causal Analysis for Social Research; Springer: Cham, Switzerland, 2013; pp. 301–328. [Google Scholar] [CrossRef]
- Grace, J.B.; Irvine, K.M. Scientist’s guide to developing explanatory statistical models using causal analysis principles. Ecology 2019, 101, e02962. [Google Scholar] [CrossRef] [PubMed]
- Delaney, A.; Dembele, A.; Nombré, I.; Lirasse, F.G.; Marshall, E.; Nana, A.; Stout, J.C.; Vickery, J.; Tayleur, C. Local-scale tree and shrub diversity improves pollination services to shea trees in tropical West African parklands. J. Appl. Ecol. 2020, 57, 1504–1513. [Google Scholar] [CrossRef]
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered Mutualisms: The Conservation of Plant-Pollinator Interactions. Annu. Rev. Ecol. Syst. 1998, 29, 83–112. [Google Scholar] [CrossRef]
- Song, B.Z.; Wu, H.Y.; Kong, Y.; Zhang, J.; Du, Y.L.; Hu, J.H.; Yao, Y.C. Effects of intercropping with aromatic plants on the diversity and structure of an arthropod community in a pear orchard. BioControl 2010, 55, 741–751. [Google Scholar] [CrossRef]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.-S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the Evidence for Biodiversity Effects on Ecosystem Functioning and Services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [Green Version]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher Levels of Multiple Ecosystem Services Are Found in Forests with More Tree Species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef]
- Plath, M.; Dorn, S.; Riedel, J.; Barrios, H.; Mody, K. Associational Resistance and Associational Susceptibility: Specialist Herbivores Show Contrasting Responses to Tree Stand Diversification. Oecologia 2012, 169, 477–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sholes, O.D.V. Effects of Associational Resistance and Host Density on Woodland Insect Herbivores. J. Anim. Ecol. 2008, 77, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.B. Resilience in Agriculture through Crop Diversification: Adaptive Management for Environmental Change. BioScience 2011, 61, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Geoff, M.G.; Stephen, D.; Wratten, J.M.L. Multi-function agricultural biodiversity: Pest management and other benefits. Basic Appl. Ecol. 2003, 4, 107–116. [Google Scholar]
- Morales, M.B.; Winqvist, C.; Geiger, F.; Bengtsson, J.; Berendse, F.; Weisser, W.; Emmerson, M.; Ceryngier, P.; Liira, J.; Tscharntke, T.; et al. Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland. Basic Appl. Ecol. 2010, 11, 97–105. [Google Scholar] [CrossRef]
- Nicholls, C.I.; Altieri, M.A. Plant Biodiversity Enhances Bees and Other Insect Pollinators in Agroecosystems. A Review. Agron. Sustain. Dev. 2013, 33, 257–274. [Google Scholar] [CrossRef] [Green Version]
- Randlkofer, B.; Obermaier, E.; Hilker, M.; Meiners, T. Vegetation complexity—The influence of plant species diversity and plant structures on plant chemical complexity and arthropods. Basic Appl. Ecol. 2010, 11, 383–395. [Google Scholar] [CrossRef]
- Perner, J.; Wytrykush, C.; Kahmen, A.; Buchmann, N.; Egerer, I.; Creutzburg, S.; Odat, N.; Audorff, V.; Weisser, W. Effects of Plant Diversity, Plant Productivity and Habitat Parameters on Arthropod Abundance in Montane European Grasslands. Ecography 2005, 28, 429–442. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Change 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Cilas, C.; Babin, R.; Avelino, J. Tropical Crop Pests and Diseases in a Climate Change Setting—A Few Examples. In Climate Change and Agriculture Worldwide; Springer: Cham, Switzerland, 2016; pp. 73–82. [Google Scholar] [CrossRef] [Green Version]
- Kraus, E.C.; Stout, M.J. Direct and Indirect Effects of Herbicides on Insect Herbivores in Rice, Oryza sativa. Sci. Rep. 2019, 9, 6998. [Google Scholar] [CrossRef] [PubMed]
- Kughur, P.G. The Effects of Herbicides on Crop Production and Environment in Makurdi Local. J. Sustain. Develop. Africa 2012, 14, 206–216. [Google Scholar]
- Harvey, R. Gros plan sur les parcs agroforestiers. Sahel Agroforesterie 2008, 11–12, 1–16. [Google Scholar]
- Gallé, R.; Happe, A.; Baillod, A.B.; Tscharntke, T.; Batáry, P. Landscape configuration, organic management, and within-field position drive functional diversity of spiders and carabids. J. Appl. Ecol. 2018, 56, 63–72. [Google Scholar] [CrossRef]
- Stephenson, A.G. Fruit Set, Herbivory, Fruit Reduction, and the Fruiting Strategy of Catalpa Speciosa (Bignoniaceae). Ecology 1980, 61, 57–64. [Google Scholar] [CrossRef]
- Kozłowski, J. Optimal Allocation of Resources to Growth and Reproduction: Implications for Age and Size at Maturity. Trends Ecol. Evol. 1992, 7, 15–19. [Google Scholar] [CrossRef]
- Marquis, R.J. Leaf Herbivores Decrease Fitness of a Tropical Plant. Science 1984, 226, 537–539. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Chiariello, N.R.; Coley, P.D.; Pitelka, L.F. Allocating Resources to Reproduction and Defense. BioScience 1987, 37, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.Y.; Zielinski, R.E.; Zangerl, A.R.; Crofts, A.R.; Berenbaum, M.R.; Delucia, E.H. The Differential Effects of Herbivory by First and Fourth Instars of Trichoplusia Ni (Lepidoptera: Noctuidae) on Photosynthesis in Arabidopsis Thaliana. J. Exp. Bot. 2006, 57, 527–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Scientific Name | Family | Order | Trophic Group |
|---|---|---|---|
| Oplostomus fuligineus | Scarabaeidae | Coleoptera | Omnivore |
| Ctenicera destructor | Elateridae | Coleoptera | Herbivore |
| Gryllus sp. | Grillidae | Orthoptera | Herbivore |
| Smicronyx fulvus | Curculionidae | Coleoptera | Herbivore |
| Zonocerus variegatus | Pyrgomorphideae | Orthoptera | Herbivore |
| Toxoptera citricida | Aphidideae | Hemiptera | Herbivore |
| Blaniulus guttulatus | Blaniulidae | Diplopoda | Herbivore (roots) |
| Locusta migratoria | Acridideae | Orthoptera | Herbivore |
| Cerotoma arcuata | Chrysomelidae | Coleoptera | Herbivore |
| Stenaptinus insignis | Carabidae | Coleoptera | Herbivore |
| Tachyura parvula | Carabidae | Coleoptera | Necrophage |
| Polydesmus spp. | Polydesmidae | Diplopoda | Herbivore (roots) |
| Caterpillar | Papilionidae | Lepidoptera | Herbivore |
| Trichopluchia ni | Noctuidae | Lepidoptera | Herbivore |
| Schistocerca gregaria | Acrididae | Orthoptera | Herbivore |
| Ichnemonide | Ichneumonidae | Hymenoptera | Carnivore |
| Dysdercus sp. | Pyrrhocoridae | Heteroptera | Nectarivore |
| Vanessa cardui | Nymphalidae | Lepidoptera | Nectarivore |
| Diabrotica virgifera | Chrysomelidae | Lepidoptera | Herbivore |
| Blaniulus sp. | Blaniulidae | Myriapoda | Detritivore |
| Schistocerca sp. | Acrididae | Orthoptera | Herbivore |
| Helicoverpa armigera | Noctuidae | Lepidoptera | Herbivore |
| Nymphalis polychloros (larvae) | Nymphalinae | Lepidoptera | Herbivore |
| Geotrupes stercorarius | Geotrupidae | Coleoptera | Coprophage |
| Arthropod Abundance | Arthropod Richness | Crop Yield (kg/ha) | Total Plant Richness | Crop Richness | Herbivory Rate (%) | |
|---|---|---|---|---|---|---|
| Minimum | 1 | 1 | 0.5 | 6 | 2 | 1.34 |
| Median | 9.5 | 4 | 550 | 11.5 | 3.5 | 2.96 |
| Mean | 13.2 | 4.7 | 828.9 | 10.8 | 3.7 | 3.02 |
| Maximum | 47 | 13 | 6250 | 14.0 | 6 | 4.52 |
| SD | 11.2 | 3.4 | 1155.9 | 2.5 | 1.03 | 0.97 |
| Sample size | 32 | 32 | 32 | 32 | 32 | 14 |
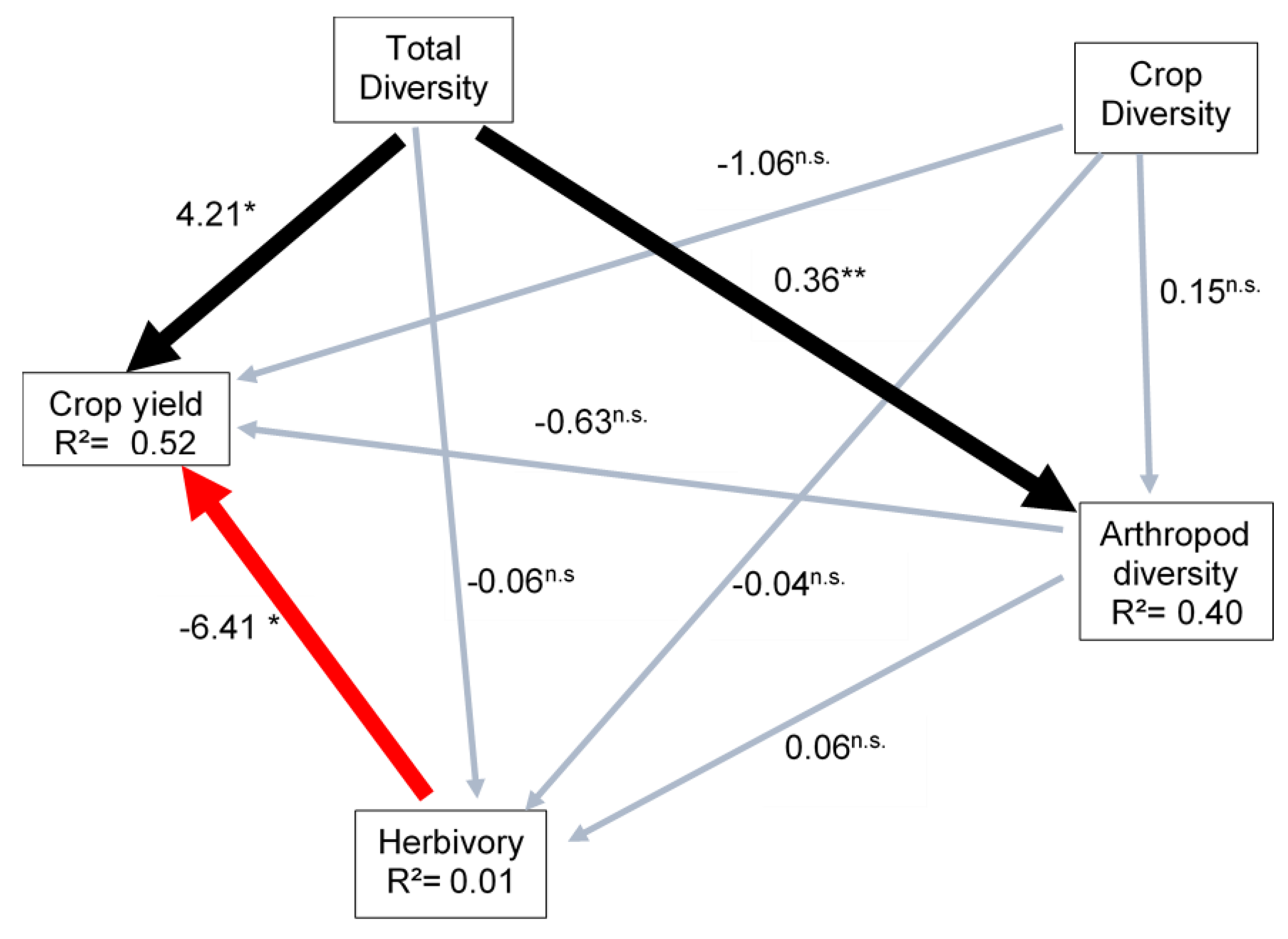
| Causal Relationships in the SEM | Estimate | SE | z | p | Std.lv | Std.all |
|---|---|---|---|---|---|---|
| Herbivory~Arthropod diversity | 0.064 | 0.281 | 0.228 | 0.820 | 0.064 | 0.078 |
| Herbivory~Crop diversity | −0.039 | 0.213 | −0.184 | 0.854 | −0.039 | −0.052 |
| Herbivory~Total diversity | −0.059 | 0.162 | −0.366 | 0.714 | −0.059 | −0.130 |
| Crop yield~Herbivory | −6.407 | 2.937 | −2.181 | 0.029 | −6.407 | −0.402 |
| Crop yield~Arthropod diversity | −0.635 | 3.099 | −0.205 | 0.838 | −0.635 | −0.049 |
| Crop yield~Crop diversity | −1.063 | 2.348 | −0.453 | 0.651 | −1.063 | −0.088 |
| Crop yield~Total diversity | 4.210 | 1.784 | 2.360 | 0.018 | 4.210 | 0.540 |
| Arthropod diversity~Total diversity | 0.363 | 0.119 | 3.062 | 0.002 | 0.363 | 0.654 |
| Arthropod diversity~Crop diversity | 0.148 | 0.199 | 0.747 | 0.455 | 0.148 | 0.160 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
N’Woueni, D.K.; Gaoue, O.G. Plant Diversity Increased Arthropod Diversity and Crop Yield in Traditional Agroforestry Systems but Has No Effect on Herbivory. Sustainability 2022, 14, 2942. https://doi.org/10.3390/su14052942
N’Woueni DK, Gaoue OG. Plant Diversity Increased Arthropod Diversity and Crop Yield in Traditional Agroforestry Systems but Has No Effect on Herbivory. Sustainability. 2022; 14(5):2942. https://doi.org/10.3390/su14052942
Chicago/Turabian StyleN’Woueni, Daniel K., and Orou G. Gaoue. 2022. "Plant Diversity Increased Arthropod Diversity and Crop Yield in Traditional Agroforestry Systems but Has No Effect on Herbivory" Sustainability 14, no. 5: 2942. https://doi.org/10.3390/su14052942
APA StyleN’Woueni, D. K., & Gaoue, O. G. (2022). Plant Diversity Increased Arthropod Diversity and Crop Yield in Traditional Agroforestry Systems but Has No Effect on Herbivory. Sustainability, 14(5), 2942. https://doi.org/10.3390/su14052942