
Characteristic of Stomatal Conductance and Optimal Stomatal Behaviour in an Arid Oasis of Northwestern China



Abstract
:1. Introduction
2. Materials and Methods
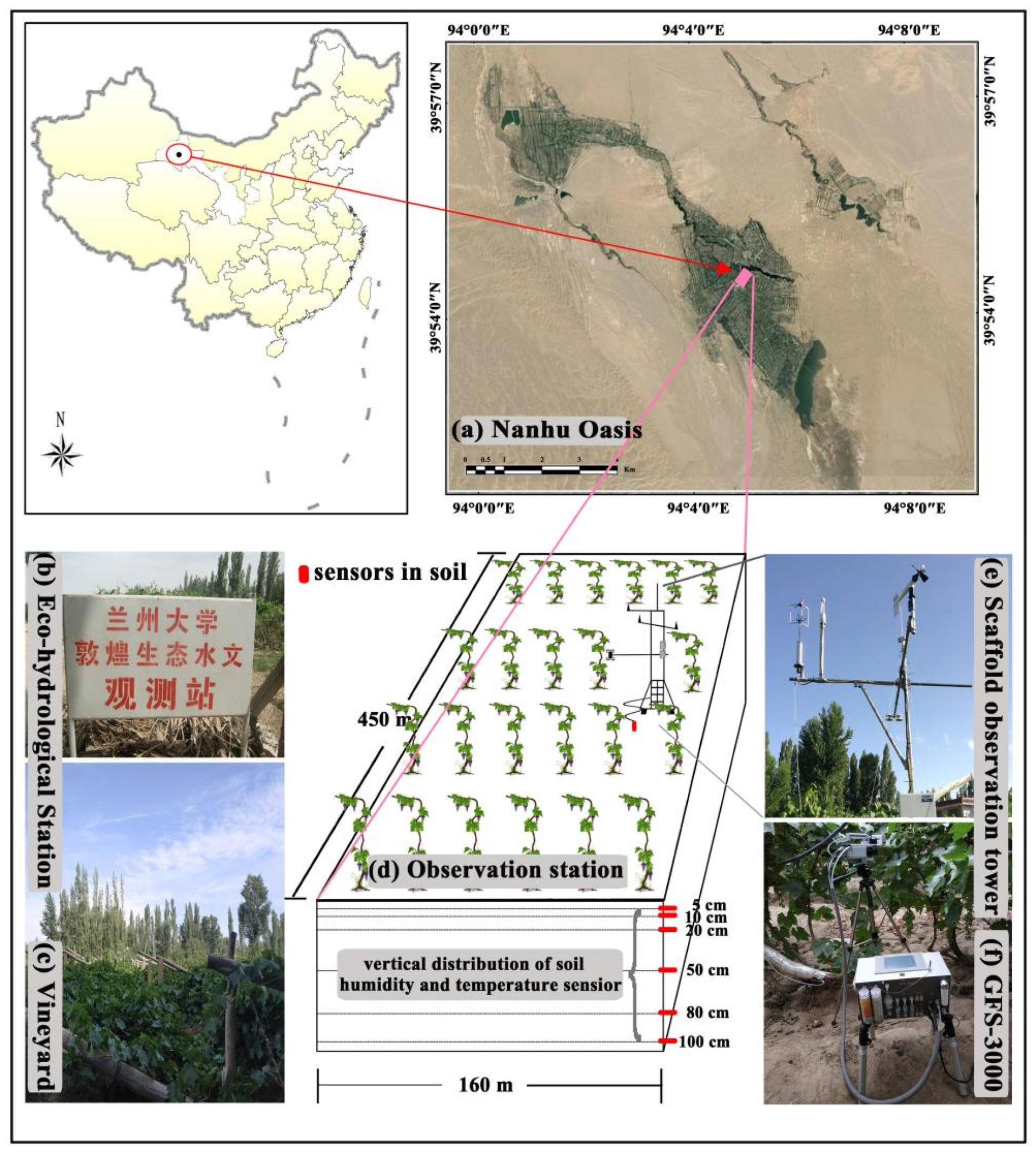
2.1. Site Description
2.2. Experimental Design and Field Measurements
2.3. Modeling Stomatal Conductance
3. Results
3.1. Environmental and Biological Variables
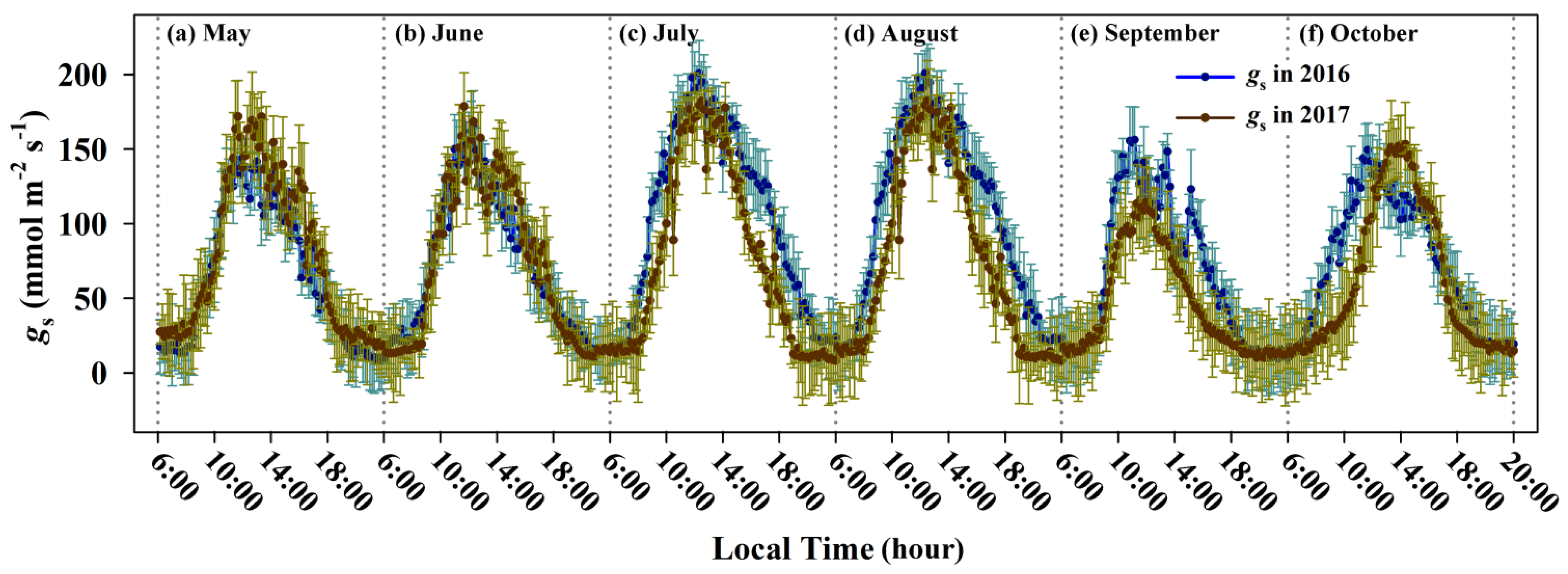
3.2. Diurnal Courses and Seasonal Variations of Stomatal Conductance
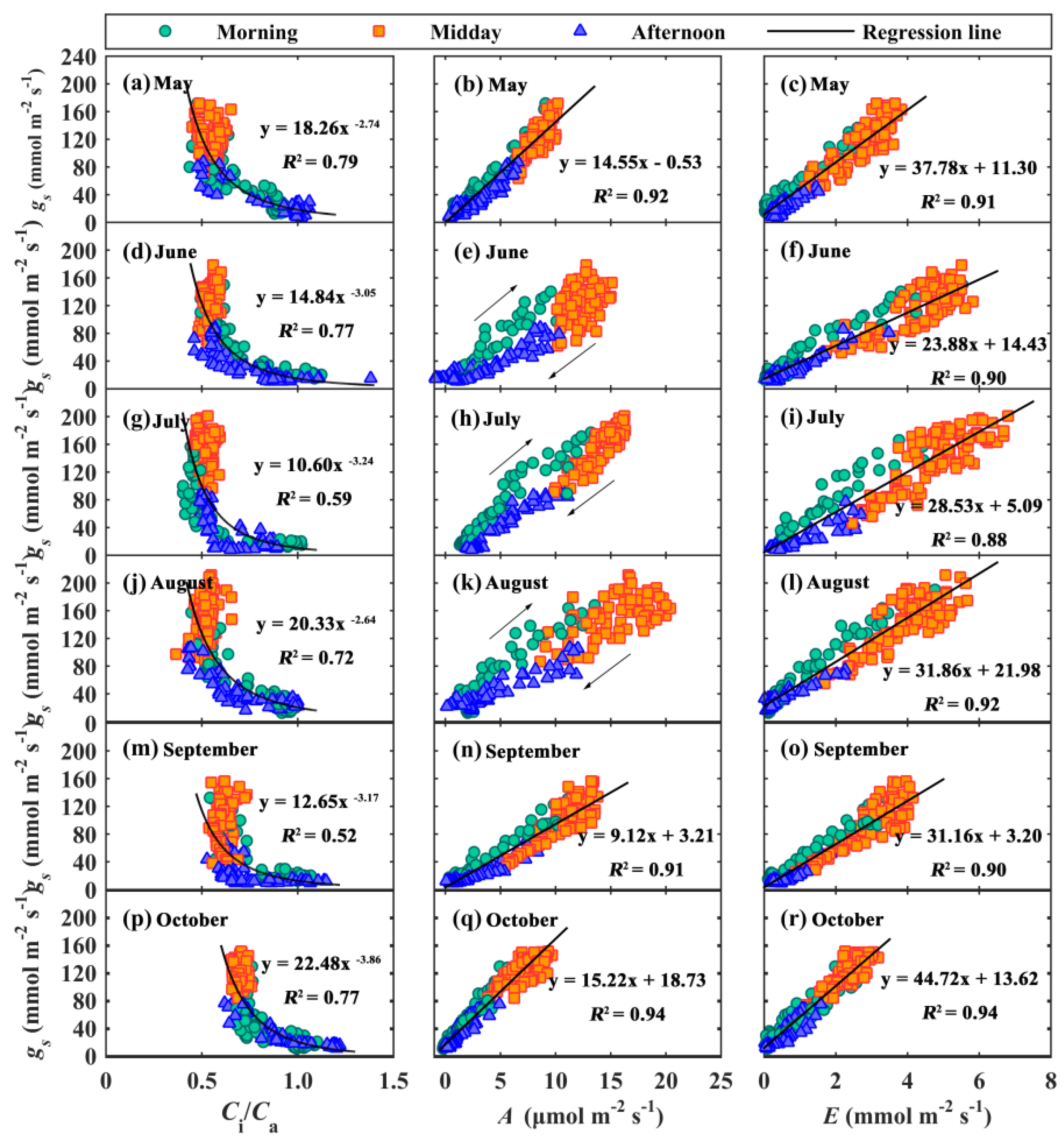
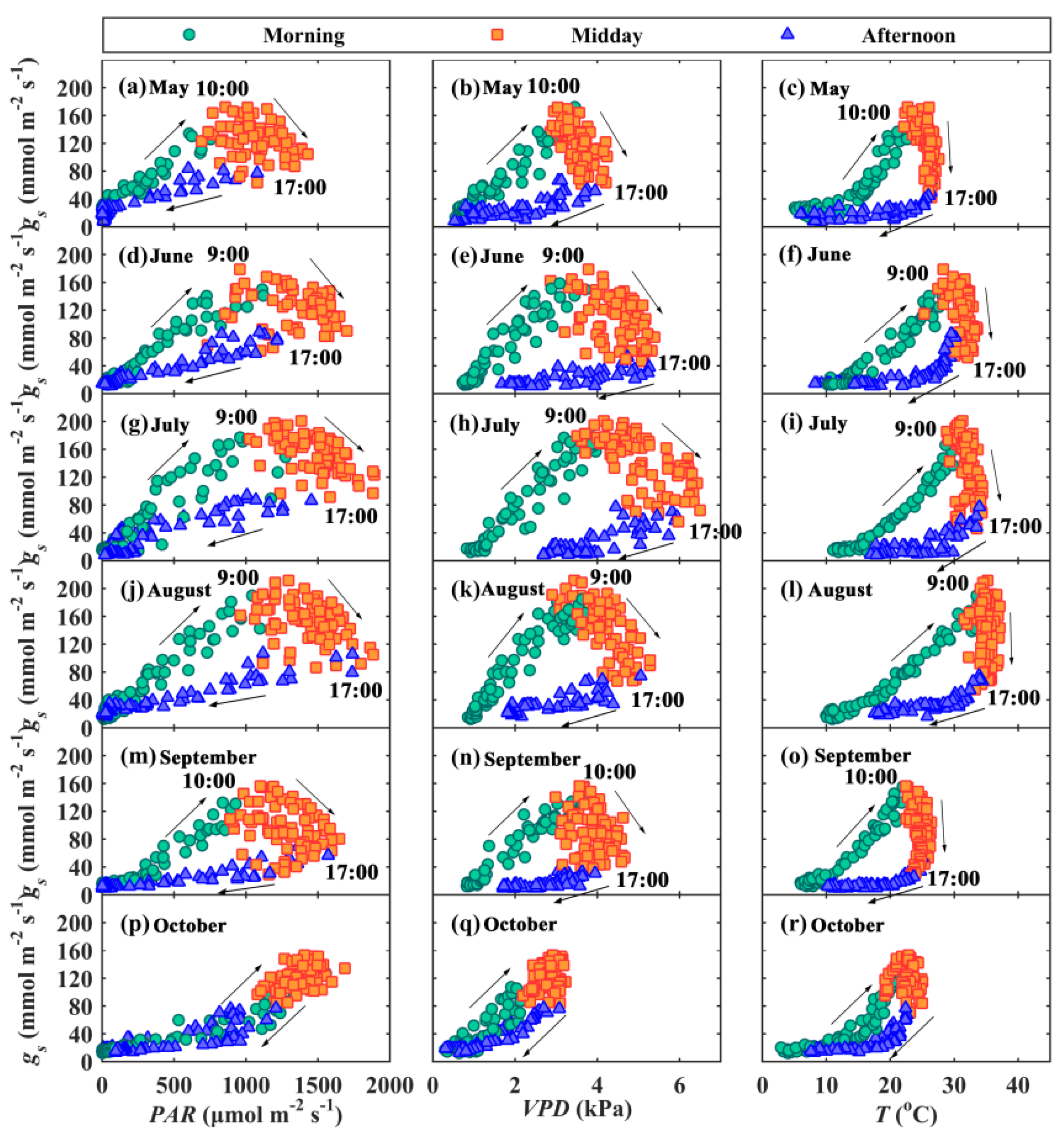
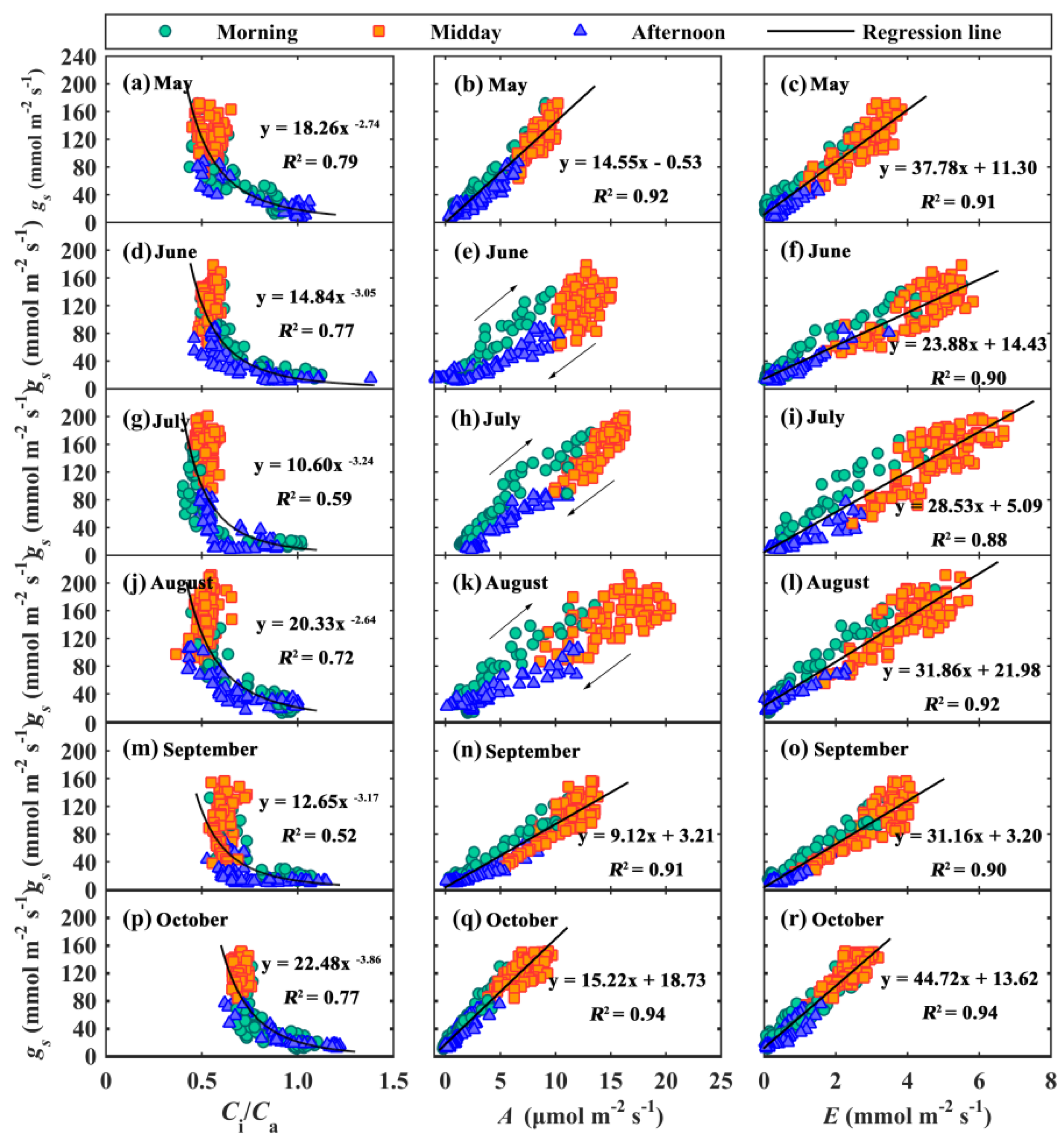
3.3. Hysteresis Loops between gs and Main Environmental and Biological Factors
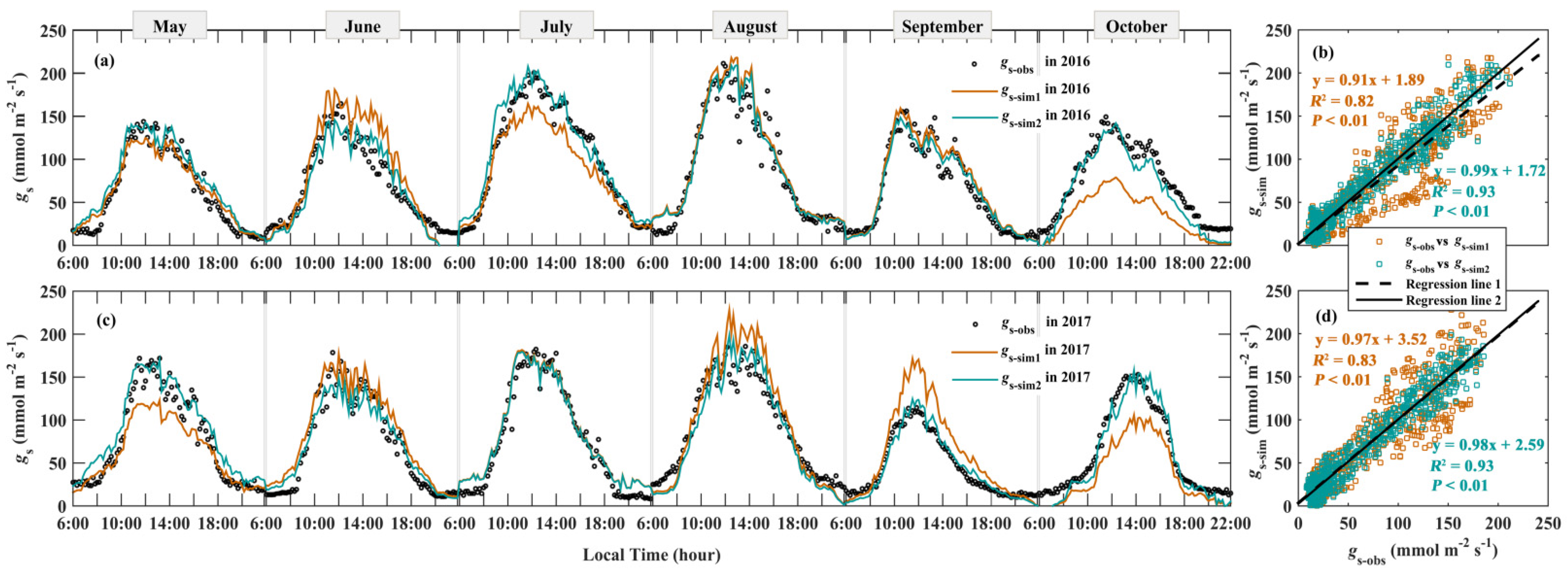
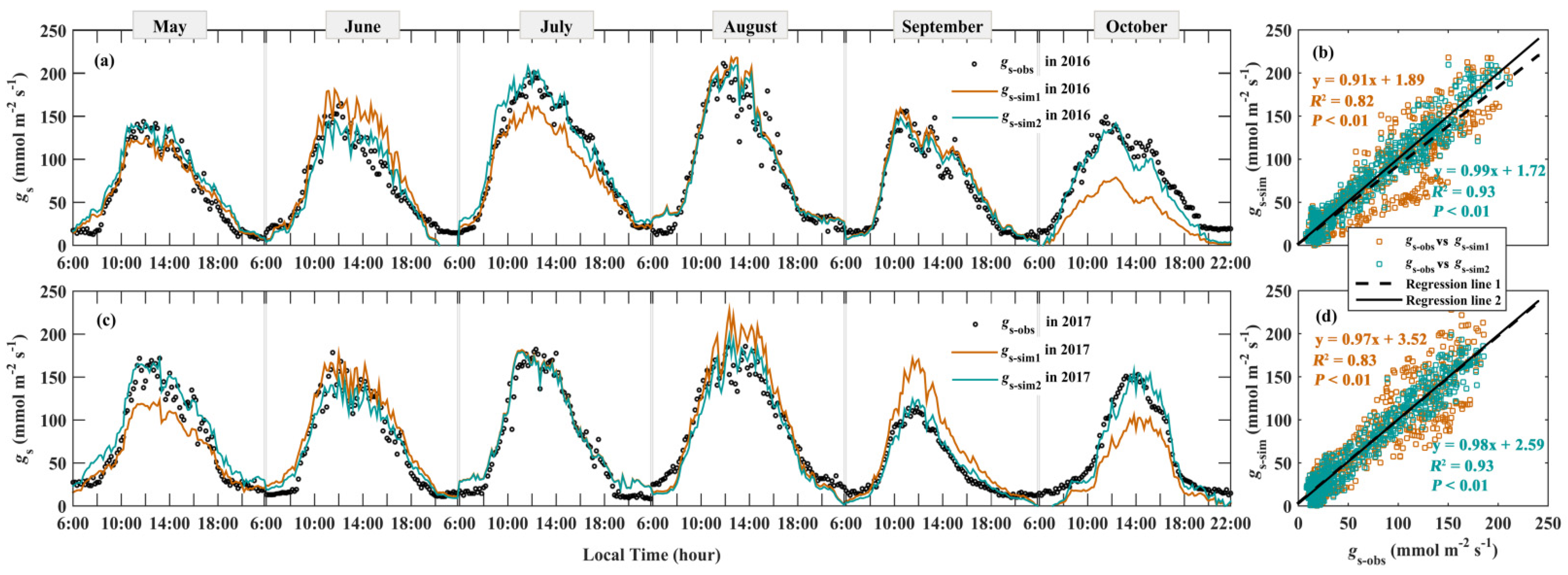
3.4. Modeling gs
4. Discussion
4.1. Hysteresis Loops between gs and Its Influencing Factors
4.2. Seasonal Variations of g1
4.3. Looking Forward
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, Y.S.; Medlyn, B.E.; Duursma, R.A.; Prentice, I.C.; Wang, H.; Baig, S.; Eamus, D.; de Dios, V.R.; Mitchell, P.; Ellsworth, D.S.; et al. Optimal stomatal behaviour around the world. Nat. Clim. Chang. 2015, 5, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Franks, P.J.; Bonan, G.B.; Berry, J.A.; Lombardozzi, D.L.; Holbrook, N.M.; Herold, N.; Oleson, K.W. Comparing optimal and empirical stomatal conductance models for application in Earth system models. Glob. Chang. Biol. 2018, 24, 5708–5723. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.A.; Beerling, D.J.; Franks, P.J. Stomata: Key players in the earth system, past and present. Curr. Opin. Plant Biol. 2010, 13, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Dewar, R.C. The Ball-Berry-Leuning and Tardieu-Davies stomatal models: Synthesis and extension within a spatially aggregated picture of guard cell function. Plant Cell Environ. 2002, 25, 1383–1398. [Google Scholar] [CrossRef]
- Ahsan, A.; Yaseen, A.A.; Yassine, C.; Ghazi, A.R.; Malik, A. Satellite-Based Water and Energy Balance Model for the Arid Region to Determine Evapotranspiration: Development and Application. Sustainability 2021, 13, 13111. [Google Scholar] [CrossRef]
- Bonan, G.B.; Williams, M.; Fisher, R.A.; Oleson, K.W. Modeling stomatal conductance in the earth system: Linking leaf water-use efficiency and water transport along the soil–plant–atmosphere continuum. Geosci. Model Dev. 2014, 7, 2193–2222. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Miner, G.L.; Bauerle, W.L.; Baldocchi, D.D. Estimating the sensitivity of stomatal conductance to photosynthesis: A review. Plant Cell Environ. 2017, 40, 1214–1238. [Google Scholar] [CrossRef]
- Franks, P.J.; Berry, J.A.; Lombardozzi, D.L.; Bonan, G.B. Stomatal Function across Temporal and Spatial Scales: Deep-Time Trends, Land-Atmosphere Coupling and Global Models. Plant Physiol. 2017, 174, 583–602. [Google Scholar] [CrossRef] [Green Version]
- Medlyn, B.E.; De Kauwe, M.G.; Lin, Y.S.; Knauer, J.; Duursma, R.A.; Williams, C.A.; Arneth, A.; Clement, R.; Isaac, P.; Limousin, J.M.; et al. How do leaf and ecosystem measures of water-use efficiency compare? New Phytol. 2017, 216, 758–770. [Google Scholar] [CrossRef] [Green Version]
- Ogle, K.; Reynolds, J.F. Desert dogma revisited: Coupling of stomatal conductance and photosynthesis in the desert shrub, Larrea tridentata. Plant Cell Environ. 2002, 25, 909–921. [Google Scholar] [CrossRef]
- Héroult, A.; Lin, Y.S.; Bourne, A.; Medlyn, B.E.; Ellsworth, D.S. Optimal stomatal conductance in relation to photosynthesis in climatically contrasting Eucalyptus species under drought. Plant Cell Environ. 2013, 36, 262–274. [Google Scholar] [CrossRef]
- Limousin, J.M.; Bickford, C.P.; Dickman, L.T.; Pangle, R.E.; Hudson, P.J.; Boutz, A.L.; Gehres, N.; Osuna, G.J.; Pockman, W.T.; Mcdowell, N.G. Regulation and acclimation of leaf gas exchange in a piñon–juniper woodland exposed to three different precipitation regimes. Plant Cell Environ. 2013, 36, 1812–1825. [Google Scholar] [CrossRef] [Green Version]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Glob. Chang. Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- Massman, W.J.; Kaufmann, M.R. Stomatal response to certain environmental factors: A comparison of models for subalpine trees in the Rocky Mountains. Agric. For. Meteorol. 1991, 54, 155–167. [Google Scholar] [CrossRef]
- Buckley, T.N.; Mott, K.A. Modelling stomatal conductance in response to environmental factors. Plant Cell Environ. 2013, 36, 1691–1699. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Wang, L.H.; Zhou, Q.; Huang, X.H. Stomatal and non-stomatal factors regulated the photosynthesis of soybean seedlings in the present of exogenous bisphenol A. Ecotoxicol. Environ. Saf. 2017, 145, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Sulman, B.N.; Roman, D.T.; Yi, K.; Wang, L.X.; Phillips, R.P.; Novick, K.A. High atmospheric demand for water can limit forest carbon uptake and transpiration as severely as dry soil. Geophys. Res. Lett. 2016, 43, 9686–9695. [Google Scholar] [CrossRef]
- Lin, C.J.; Gentine, P.; Huang, Y.F.; Guan, K.Y.; Kimm, H.; Zhou, S. Diel ecosystem conductance response to vapor pressure deficit is suboptimal and independent of soil moisture. Agric. For. Meteorol. 2018, 250–251, 21–34. [Google Scholar] [CrossRef]
- Tang, J.; Bolstad, P.V.; Ewers, B.E.; Desai, A.R.; Davis, K.J.; Carey, E.V. Sap flux-upscaled canopy transpiration, stomatal conductance, and water use efficiency in an old growth forest in the Great Lakes region of the United States. J. Geophys. Res. 2006, 111, G02009. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, Y.; Kumagai, T.O.; Yoshifuji, N.; Sato, T.; Tanaka, N.; Tanaka, K.; Suzuki, M.; Tantasirin, C. Environmental control of canopy stomatal conductance in a tropical deciduous forest in northern Thailand. Agric. For. Meteorol. 2015, 202, 1–10. [Google Scholar] [CrossRef]
- Liu, H.; Cohen, S.; Lemcoff, J.H.; Israeli, Y.; Tanny, J. Sap flow, canopy conductance and microclimate in a banana screenhouse. Agric. For. Meteorol. 2015, 201, 165–175. [Google Scholar] [CrossRef]
- Bai, Y.; Zhu, G.F.; Su, Y.H.; Zhang, K.; Han, T.; Ma, J.Z.; Wang, W.Z.; Ma, T.; Feng, L.L. Hysteresis loops between canopy conductance of grapevines and meteorological variables in an oasis ecosystem. Agric. For. Meteorol. 2015, 214–215, 319–327. [Google Scholar] [CrossRef]
- Bai, Y.; Li, X.Y.; Liu, S.M.; Wang, P. Modelling diurnal and seasonal hysteresis phenomena of canopy conductance in an oasis forest ecosystem. Agric. For. Meteorol. 2017, 246, 98–110. [Google Scholar] [CrossRef]
- Jarvis, P.G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Philos. Trans. R. Soc. Lond. Ser. B 1976, 273, 593–610. [Google Scholar] [CrossRef]
- Ball, M.C.; Woodrow, I.E.; Berry, J.A. A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions. In Progress in Photosynthesis Research; Biggins, J., Ed.; Martinus Nijhoff Publishers: Dordrecht, The Netherlands, 1987; pp. 221–224. [Google Scholar] [CrossRef]
- Leuning, R. Modelling stomatal behaviour and photosynthesis of Eucalyptus grandis. Aust. J. Plant Physiol. 1990, 17, 159–175. [Google Scholar] [CrossRef]
- Monteith, J.L. A reinterpretation of stomatal responses to humidity. Plant Cell Environ. 1995, 18, 357–364. [Google Scholar] [CrossRef]
- Cowan, I.R. Oscillations in stomatal conductance and plant functioning associated with stomatal conductance: Observations and a model. Planta 1972, 106, 185–219. [Google Scholar] [CrossRef] [PubMed]
- Hills, A.; Chen, Z.H.; Amtmann, A.; Blatt, M.R.; Lew, V.L. OnGuard, a computational platform for quantitative kinetic modeling of guard cell physiology. Tree Physiol. 2012, 159, 1026–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prentice, I.C.; Dong, N.; Gleason, S.M.; Maire, V.; Wright, I.J. Balancing the costs of carbon gain and water transport: Testing a new theoretical framework for plant functional ecology. Ecol. Lett. 2014, 17, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Sellers, P.J.; Randall, D.A.; Collatz, G.J.; Berry, J.A.; Field, C.B.; Dazlich, D.A.; Zhang, C.; Collelo, G.D.; Bounoua, L. A revised land surface parameterization (SiB2) for atmospheric GCMs. Part 1: Model formulation. J. Clim. 1996, 9, 676–705. [Google Scholar] [CrossRef]
- Bonan, G.B.; Doney, S.C. Limate, ecosystems, and planetary futures: The challenge to predict life in Earth system models. Science 2018, 359, eaam8328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowan, I.R.; Farquhar, G.D. Stomatal function in relation to leaf metabolism and environment. In Integration of Activity in the Higher Plant; Jennings, D.H., Ed.; Cambridge University Press: Cambridge, UK, 1977; pp. 471–505. [Google Scholar] [CrossRef]
- Knauer, J.; Zaehle, S.; Medlyn, B.E.; Reichstein, M.; Williams, C.A.; Migliavacca, M.; De Kauwe, M.G.; Werner, C.; Keitel, C.; Kolari, P.; et al. Towards physiologically meaningful water-use efficiency estimates from eddy covariance data. Glob. Chang. Biol. 2018, 24, 694–710. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.R.; Tanner, C.B.; Bennett, J.M. Water-use efficiency in crop production. BioScience 1984, 34, 36–40. [Google Scholar] [CrossRef]
- Ye, Z.; Fu, A.; Zhang, S.; Yang, Y. Suitable scale of an oasis in different scenarios in an arid region of china: A case study of the ejina oasis. Sustainability 2020, 12, 2583. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Li, W.; Yang, G.; Yan, K.; He, X.; Li, F.; Gao, Y.; Tian, L. Moisture, temperature, and salinity of a typical desert plant (Haloxylon ammodendron) in an arid oasis of northwest china. Sustainability 2021, 13, 1908. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, R.H. Water vapor exchange between soil and atmosphere over a Gobi surface near an oasis in Summer. J. Appl. Meteorol. 2004, 43, 1917–1928. [Google Scholar] [CrossRef]
- Sperry, J.S.; Meinzer, F.C.; McCulloh, K.A. Safety and efficiency conflicts in hydraulic architecture: Scaling from tissues to trees. Plant Cell Environ. 2008, 31, 632–645. [Google Scholar] [CrossRef] [PubMed]
- De Kauwe, M.G.; Medlyn, B.E.; Zaehle, S.; Walker, A.P.; Dietzem, C.; Hickler, T.; Jain, A.K.; Luo, Y.Q.; Parton, W.J.; Prentice, I.C.; et al. Forest water use and water use efficiency at elevated CO2: A model-data intercomparison at two contrasting temperate forest FACE sites. Glob. Chang. Biol. 2013, 19, 1759–1779. [Google Scholar] [CrossRef]
- Zhu, G.F.; Li, X.; Zhang, K.; Ding, Z.Y.; Han, T.; Ma, J.Z.; Huang, C.L.; He, J.H.; Ma, T. Multi-model ensemble prediction of terrestrial evapotranspiration across north China using Bayesian model averaging. Hydrol. Process. 2016, 30, 2861–2879. [Google Scholar] [CrossRef]
- Merilo, E.; Jõesaar, I.; Brosché, M.; Kollist, H. To open or to close: Species-specific stomatal responses to simultaneously applied opposing environmental factors. New Phytol. 2014, 202, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Kołodziejek, J. Growth and competitive interaction between seedlings of an invasive Rumex confertus and of cooccurring two native Rumex species in relation to nutrient availability. Nature 2019, 9, 3298. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Zhou, G.; He, Q. Critical leaf water content for maize photosynthesis under drought stress and its response to rewatering. Sustainability 2021, 13, 7218. [Google Scholar] [CrossRef]
- Ono, K.; Maruyama, A.; Kuwagata, T.; Mano, M.; Takimoto, T.; Hayashi, K.; Hasegawa, T.; Miyata, A. Canopy-scale relationships between stomatal conductance and photosynthesis in irrigated rice. Glob. Chang. Biol. 2013, 19, 2209–2220. [Google Scholar] [CrossRef]
- Yan, W.M.; Zhong, Y.Q.W.; Shangguan, Z.P. Contrasting responses of leaf stomatal characteristics to climate change: A considerable challenge to predict carbon and water cycles. Glob. Chang. Biol. 2017, 23, 3781–3793. [Google Scholar] [CrossRef] [PubMed]
- Scafaro, A.P.; Xiang, S.; Long, B.M.; Bahar, N.A.; Weerasinghe, L.K.; Creek, D.; Evans, J.R.; Reith, P.B.; Atkin, O.K. Strong thermal acclimation of photosynthesis in tropical and temperate wet-forest tree species: The importance of altered Rubisco content. Glob. Chang. Biol. 2017, 23, 2783–2800. [Google Scholar] [CrossRef]
- Wu, C.; Wang, X.; Wang, H.; Ciais, P.; Peñuelas, J.; Myneni, R.B.; Desai, A.R.; Gough, C.M.; Gonsamo, A.; Black, A.T.; et al. Contrasting resp-onses of autumn-leaf senescence to daytime and night-time warming. Nat. Clim. Chang. 2018, 8, 1092–1096. [Google Scholar] [CrossRef] [Green Version]
- Drake, J.E.; Tjoelker, M.G.; Vårhammar, A.; Medlyn, B.E.; Reich, P.B.; Leigh, A.; Pfautsch, S.; Blackman, C.J.; López, R.; Aspinwall, M.J.; et al. Trees tolerate an extreme heatwave via sustained transpirational cooling and increased leaf thermal tolerance. Glob. Chang. Biol. 2018, 24, 2390–2402. [Google Scholar] [CrossRef] [PubMed]
- Marchin, R.M.; Broadhead, A.A.; Bostic, L.E.; Dunn, R.R.; Hoffmann, W.A. Stomatal acclimation to vapor pressure d-eficit doubles transpiration of small tree seedlings with warming. Plant Cell Environ. 2016, 39, 2221–2234. [Google Scholar] [CrossRef] [PubMed]
- Duursma, R.A.; Barton, C.V.M.; Lin, Y.S.; Medlyn, B.E.; Eamus, D.; Tissue, D.T.; Ellsworth, D.S.; McMurtrie, R.E. The peaked response of transpiration rate to vapour pressure deficit in field conditions can be explained by the temperature optimum of photosynthesis. Agric. For. Meteorol. 2014, 189, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Zhao, W.; Zhang, Z.; Su, Y. Sap flow and tree conductance of shelter-belt in arid region of China. Agric. For. Meteorol. 2006, 138, 132–141. [Google Scholar] [CrossRef]
- Jones, H.G. Plant and Microclimate: A Quantitative Approach to Environmental Plant Physiology, 3rd ed.; Cambridge University Press: Cambridge, UK, 2014; pp. 135–144. [Google Scholar] [CrossRef]
- Wang, S.T.; Zhu, G.F.; Xia, D.S.; Ma, J.Z.; Han, T.; Ma, T.; Zhang, K.; Shang, S.S. The characteristics of evapotranspiration and crop coefficients of an irrigated vineyard in arid Northwest China. Agric. Water Manag. 2019, 212, 388–398. [Google Scholar] [CrossRef]
- Rodrigues, T.R.; Vourlitis, G.L.; Lobo, F.D.A.; Santanna, F.B.; Arruda, P.H.Z.D.; Nogueira, J.D.S. Modeling canopy conductance under contrasting seasonal conditions for a tropical savanna ecosystem of south central Mato Grosso, Brazil. Agric. For. Meteorol. 2016, 23, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Tie, Q.; Hua, H.C.; Tian, F.Q.; Guan, H.D.; Lin, H. Environmental and physiological controls on sap flow in a subh-umid mountainous catchment in North China. Agric. For. Meteorol. 2017, 240–241, 46–57. [Google Scholar] [CrossRef]
- Aasamaa, K.; Sõber, A. Stomatal sensitivities to changes in leaf water potential, air humidity, CO2 concentration and light intensity, and the effect of abscisic acid on the sensitivities in six temperate deciduous tree species. Environ. Exp. Bot. 2011, 71, 72–78. [Google Scholar] [CrossRef]
- Song, Q.H.; Fei, X.H.; Zhang, Y.P.; Sha, L.Q.; Liu, Y.T.; Zhou, W.J.; Wu, C.S.; Lu, Z.Y.; Luo, K.; Gao, J.B.; et al. Water use efficiency in a primary subtropical evergreen forest in Southwest China. Sci. Rep. 2017, 7, 43031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kauwe, M.G.; Kala, J.; Lin, Y.S.; Pitman, A.J.; Medlyn, B.E.; Duursma, R.A.; Abramowitz, G.; Wang, Y.P.; Miralles, D.G. A test of an optimal stomatal conductance scheme within the CABLE land surface model. Geosci. Model Dev. 2015, 8, 431–452. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xiao, J.; He, B.; Arain, M.A.; Beringer, J.; Desai, A.R.; Emmel, C.; Hollinger, D.Y.; Krasnova, A.; Mammarella, I.; et al. Solar-induced chlorophyll fluorescence is strongly correlated with terrestrial photosynthesis for a wide variety of biomes: First global analysis based on OCO-2 and flux tower observations. Glob. Chang. Biol. 2018, 24, 3990–4008. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Wolf, A.; Arango-Velez, A.; Choat, B.; Chmura, D.J.; Jansen, S.; Kolb, T.; Li, S.; Meinzer, F.C.; Pita, P.; et al. Woody plants optimise stomatal behaviour relative to hydraulic risk. Ecol. Lett. 2018, 21, 968–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Factor | Morning | Midday | Afternoon | |||
|---|---|---|---|---|---|---|---|
| Regression Line | R2 | Regression Line | R2 | Regression Line | R2 | ||
| May | PAR | gs = 0.15 × PAR + 22.72 | 0.94 | gs = −0.04 × PAR + 175.25 | 0.10 | gs = 0.06 × PAR + 18.91 | 0.89 |
| VPD | gs = 46.01 × VPD − 10.68 | 0.91 | gs = −37.91 × VPD + 258.42 | 0.33 | gs = 9.19 × e0.49VPD | 0.69 | |
| T | gs = 6.03 × T − 29.63 | 0.76 | gs = −11.45 × T + 418.74 | 0.40 | gs = 5.54 × e0.08T | 0.60 | |
| Jun | PAR | gs = 0.15 × PAR + 8.79 | 0.92 | gs = −0.04 × PAR + 183.36 | 0.17 | gs = 0.06 × PAR + 12.92 | 0.90 |
| VPD | gs = 56.68 × VPD − 29.63 | 0.87 | gs = −17.61 × VPD + 202.82 | 0.26 | gs = 6.42 × e0.42VPD | 0.54 | |
| T | gs = 7.04 × T − 70.79 | 0.90 | gs = −5.42 × T + 296.24 | 0.19 | gs = 4.35 × e0.08T | 0.73 | |
| Jul | PAR | gs = 0.14 × PAR + 17.41 | 0.76 | gs = −0.02 × PAR + 176.02 | 0.02 | gs = 0.07 × PAR + 12.68 | 0.87 |
| VPD | gs = 50.66 × VPD − 33.76 | 0.90 | gs = −18.11 × VPD + 236.09 | 0.23 | gs = 2.18 × e0.59VPD | 0.74 | |
| T | gs = 7.14 × T − 78.97 | 0.91 | gs = −18.97 × T + 758.69 | 0.56 | gs = 1.04 × e0.12T | 0.64 | |
| Aug | PAR | gs = 0.15 × PAR + 19.09 | 0.93 | gs = −0.05 × PAR + 217.40 | 0.12 | gs = 0.04 × PAR + 23.60 | 0.88 |
| VPD | gs = 49.42 × VPD − 46.36 | 0.89 | gs = −24.43 × VPD + 246.48 | 0.28 | gs = 11.75 × e0.37VPD | 0.72 | |
| T | gs = 6.05 × T − 53.41 | 0.94 | gs = −1.88 × T + 215.87 | 0.01 | gs = 5.41 × e0.07T | 0.72 | |
| Sep | PAR | gs = 0.12 × PAR + 5.64 | 0.94 | gs = −0.02 × PAR + 113.27 | 0.01 | gs = 0.03 × PAR + 10.05 | 0.81 |
| VPD | gs = 48.43 × VPD − 27.72 | 0.95 | gs = −11.93 × VPD + 136.54 | 0.03 | gs = 3.53 × e0.57VPD | 0.70 | |
| T | gs = 8.19 × T − 47.95 | 0.94 | gs = −3.05 × T + 166.36 | 0.01 | gs = 3.60 × e0.09T | 0.69 | |
| Oct | PAR | gs = 16.45 × e0.0014PAR | 0.94 | - | - | gs = 16.01 × e0.0013PAR | 0.79 |
| VPD | gs = 11.58 × e0.92VPD | 0.83 | - | - | gs = 11.43 × e0.70VPD | 0.92 | |
| T | gs = 8.88 × e0.12T | 0.94 | - | - | gs = 5.66 × e0.10T | 0.72 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, T.; Feng, Q.; Yu, T.; Yang, X.; Zhang, X.; Li, K. Characteristic of Stomatal Conductance and Optimal Stomatal Behaviour in an Arid Oasis of Northwestern China. Sustainability 2022, 14, 968. https://doi.org/10.3390/su14020968
Han T, Feng Q, Yu T, Yang X, Zhang X, Li K. Characteristic of Stomatal Conductance and Optimal Stomatal Behaviour in an Arid Oasis of Northwestern China. Sustainability. 2022; 14(2):968. https://doi.org/10.3390/su14020968
Chicago/Turabian StyleHan, Tuo, Qi Feng, Tengfei Yu, Xiaomei Yang, Xiaofang Zhang, and Kuan Li. 2022. "Characteristic of Stomatal Conductance and Optimal Stomatal Behaviour in an Arid Oasis of Northwestern China" Sustainability 14, no. 2: 968. https://doi.org/10.3390/su14020968
APA StyleHan, T., Feng, Q., Yu, T., Yang, X., Zhang, X., & Li, K. (2022). Characteristic of Stomatal Conductance and Optimal Stomatal Behaviour in an Arid Oasis of Northwestern China. Sustainability, 14(2), 968. https://doi.org/10.3390/su14020968