



Investigating Population Genetic Diversity and Rhizosphere Microbiota of Central Apennines’ Artemisia eriantha

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Population Genetic Diversity
2.3. Microbiota 16S rRNA Gene Metabarcoding
2.4. Statistical Analysis
3. Results
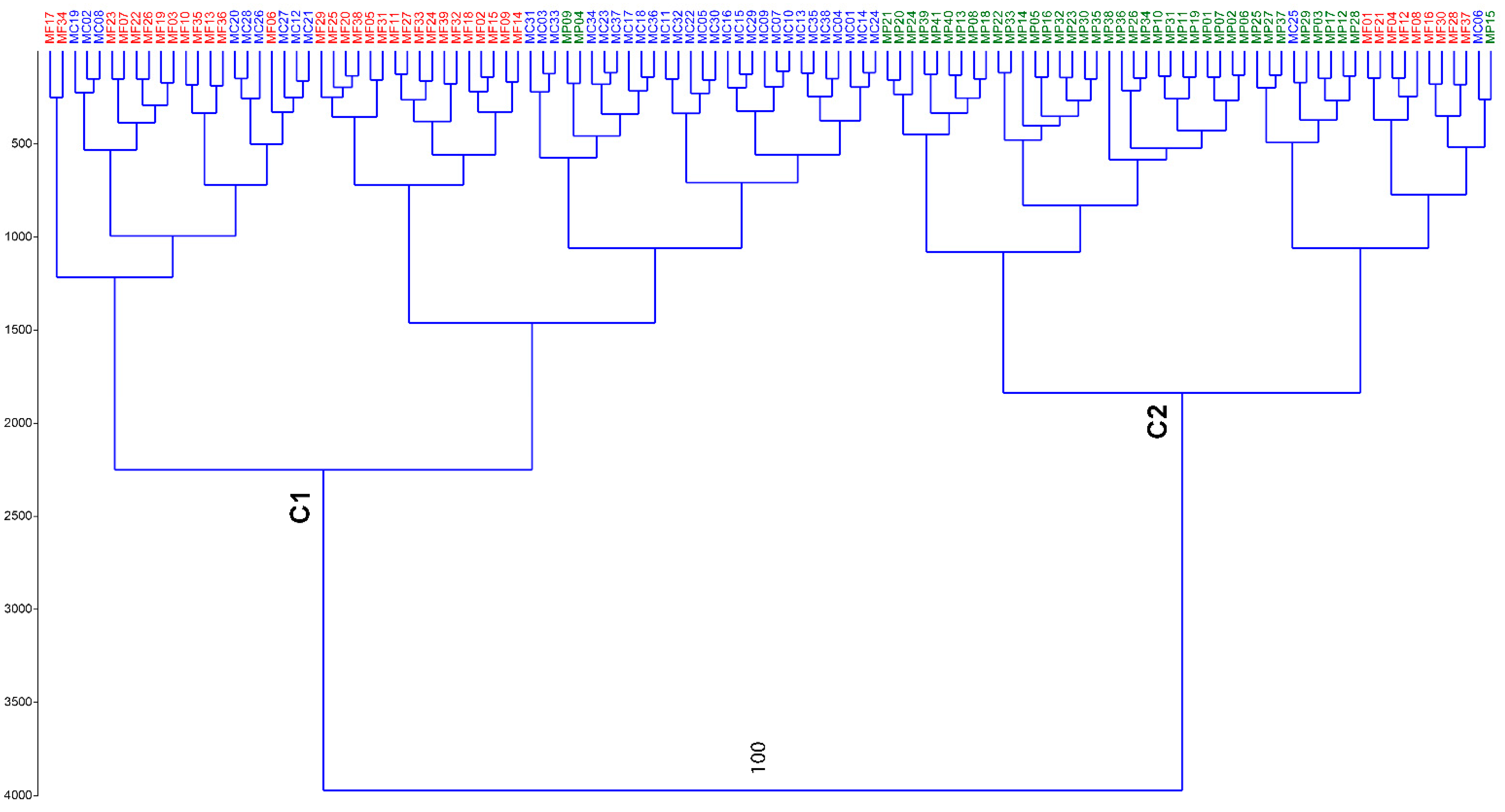
3.1. Genetic Characterization
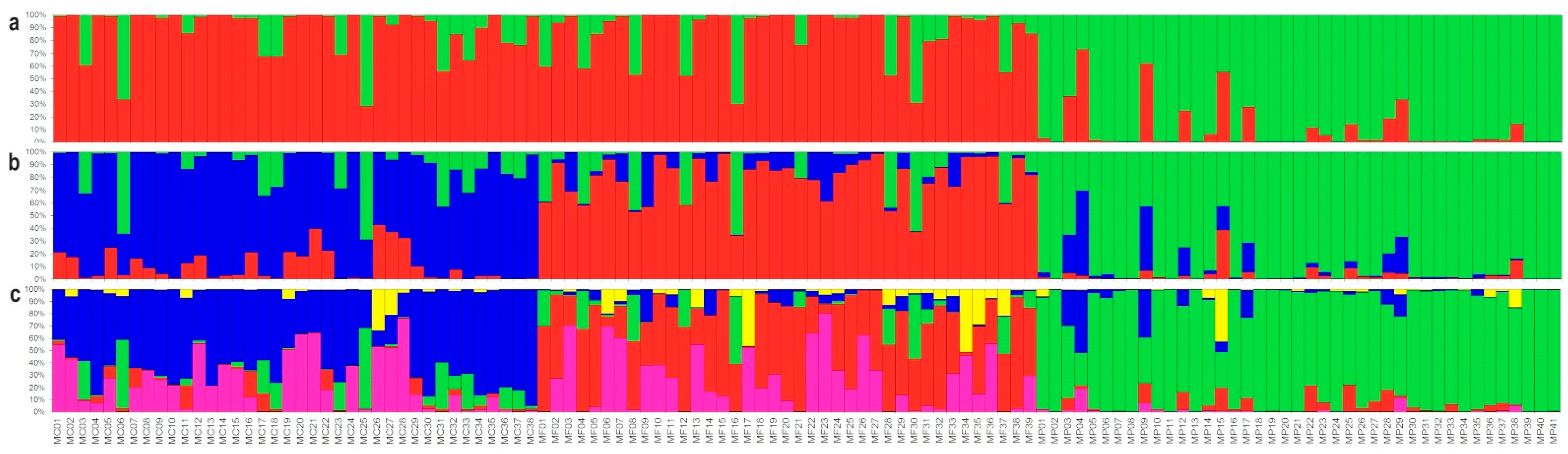
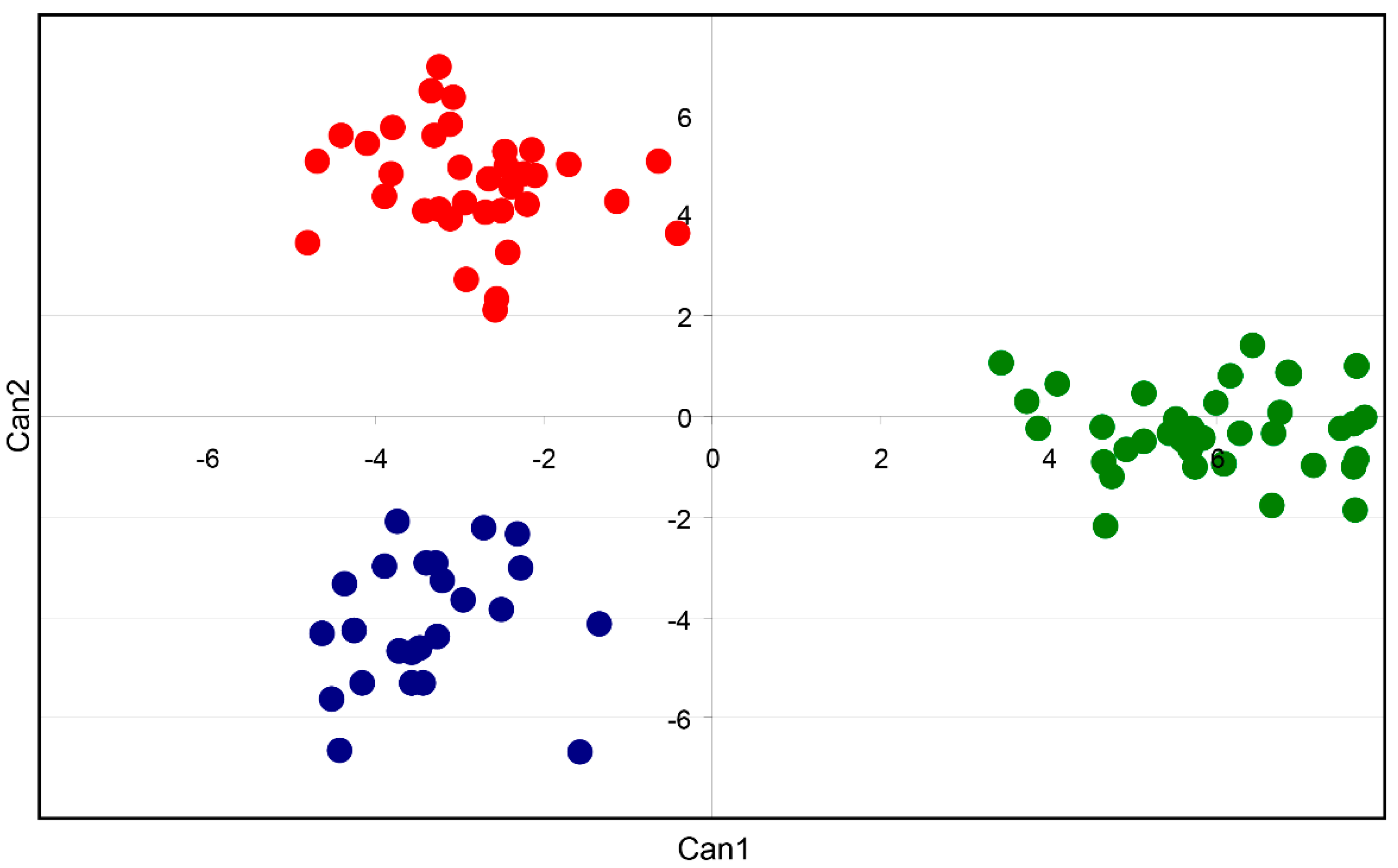
3.2. Genetic Structure
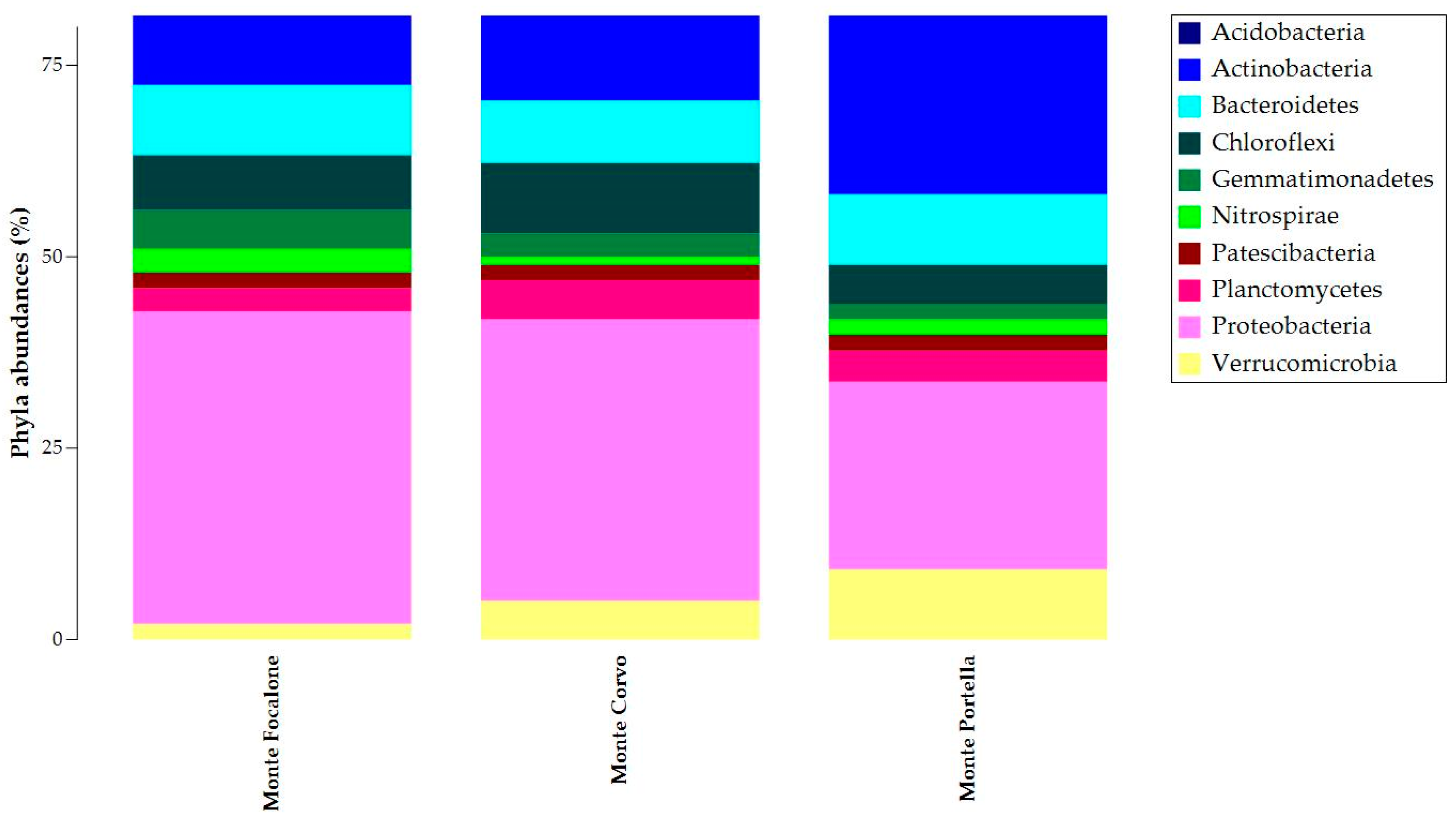
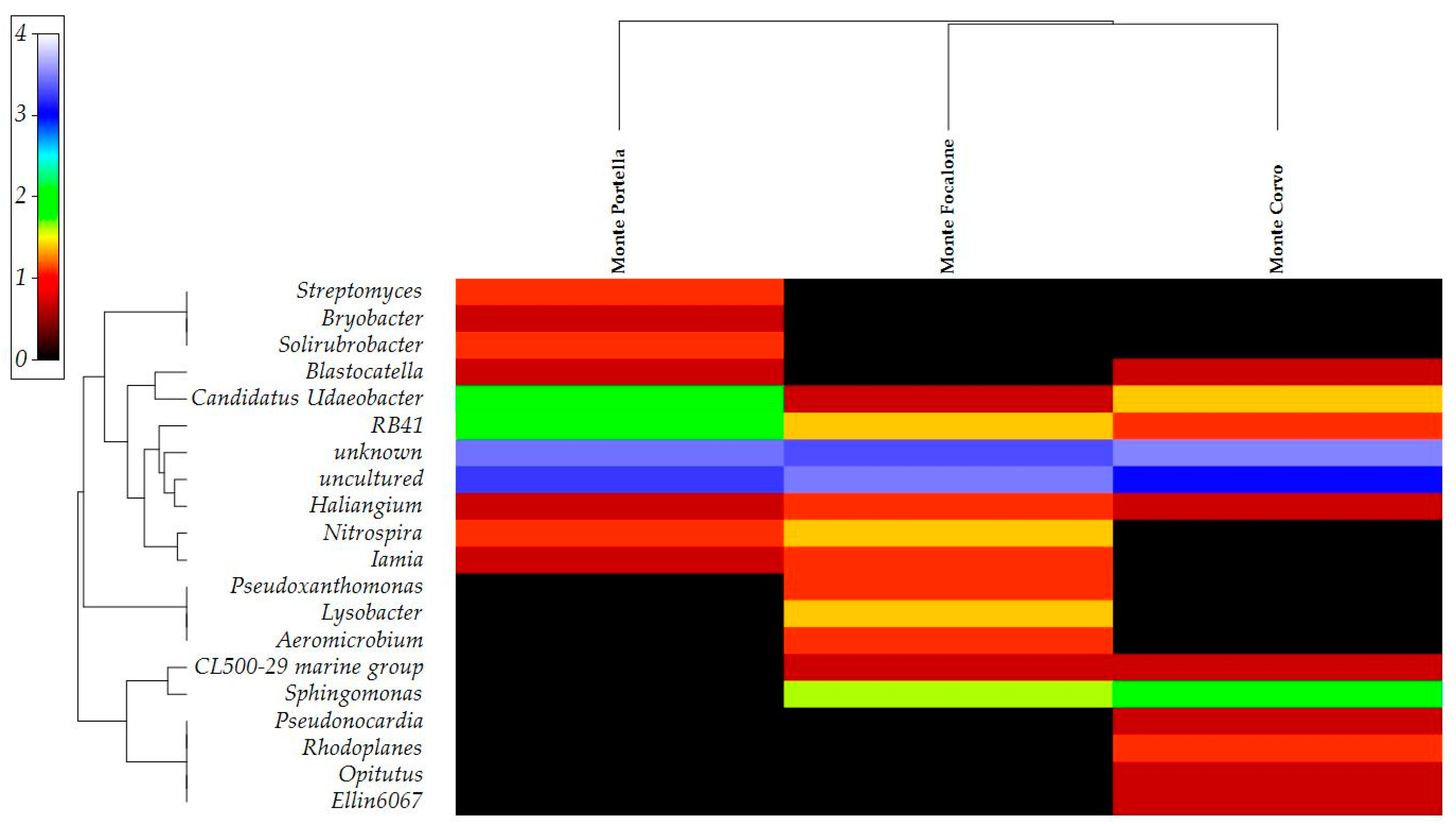
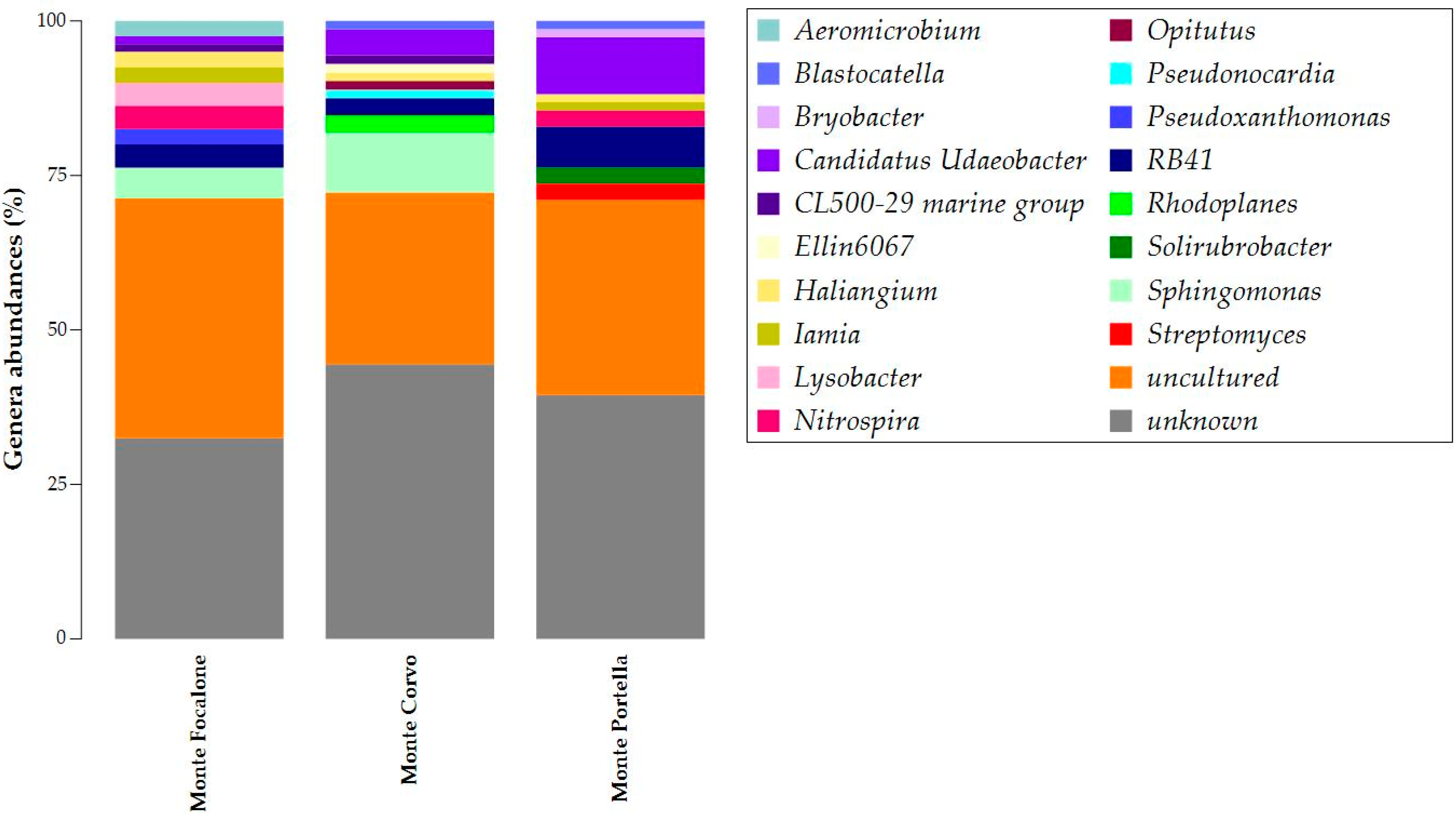
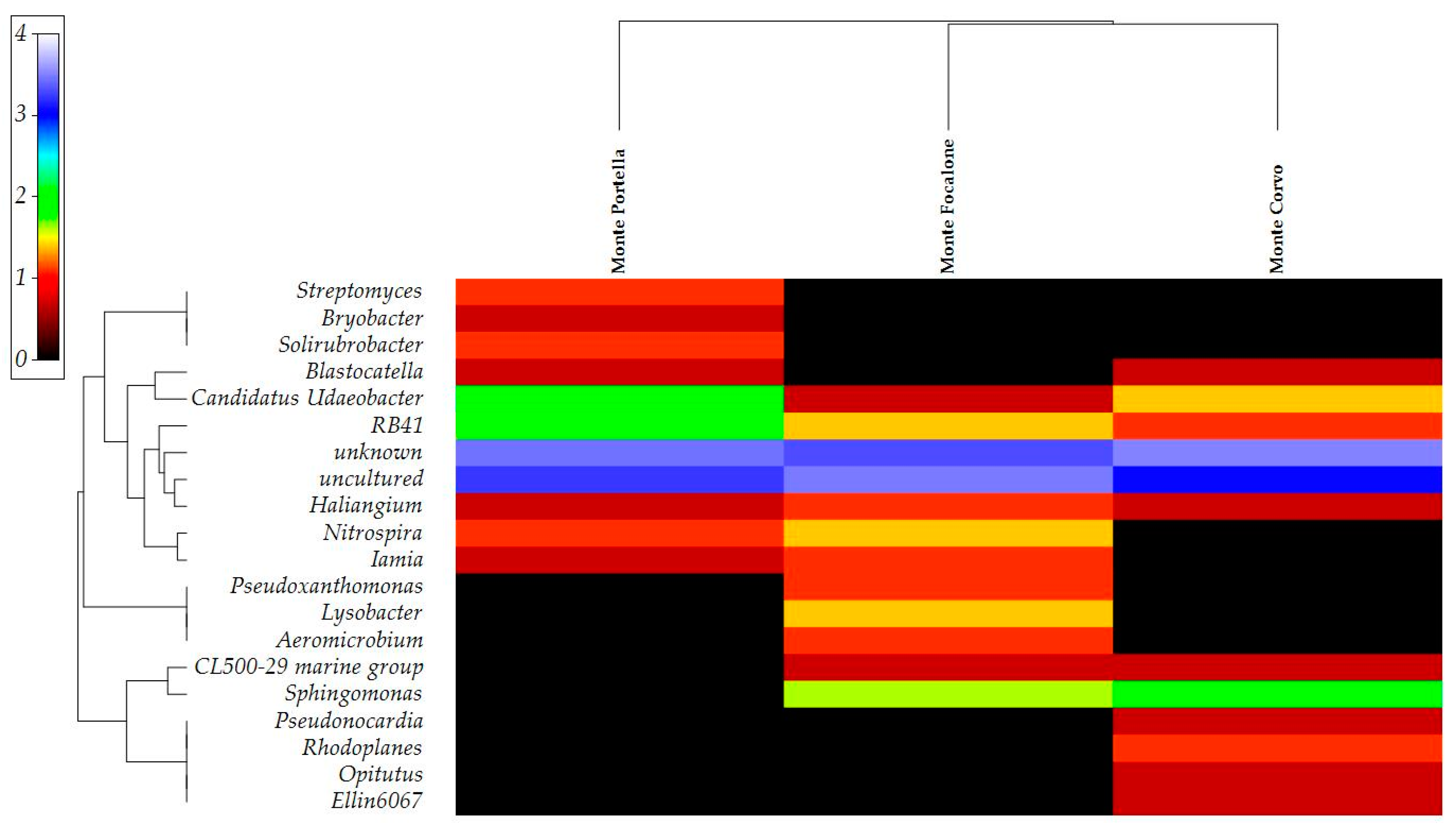
3.3. Microbiota 16S rRNA Gene Metabarcoding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ling, Y.R. On the System of the Genus Artemisia L. and the Relationship with Its Allies. Bull. Bot. Res. 1982, 2, 1–60. [Google Scholar]
- Wang, W.-M. On the Origin and Development of Artemisia (Asteraceae) in the Geological Past. Bot. J. Linn. Soc. 2004, 145, 331–336. [Google Scholar] [CrossRef]
- Graham, A. A Contribution to the Geological History of the Compositae. In Proceedings of the Kew International Compositae Conference 1994; Hind, D., Beentje, H., Eds.; Royal Botanic Gardens, Kew: London, UK, 1996; pp. 123–140. [Google Scholar]
- Yunfa, M.; Qingquan, M.; Xiaomin, F.; Xiaoli, Y.; Fuli, W.; Chunhui, S. Origin and Development of Artemisia (Asteraceae) in Asia and Its Implications for the Uplift History of the Tibetan Plateau: A Review. Quat. Int. 2011, 236, 3–12. [Google Scholar] [CrossRef]
- Weiss-Schneeweiss, H.; Emadzade, K.; Jang, T.-S.; Schneeweiss, G.M. Evolutionary Consequences, Constraints and Potential of Polyploidy in Plants. Cytogenet. Genome Res. 2013, 140, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Otto, S.P.; Whitton, J. Polyploid Incidence and Evolution. Annu. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [PubMed]
- de Storme, N.; Mason, A. Plant Speciation through Chromosome Instability and Ploidy Change: Cellular Mechanisms, Molecular Factors and Evolutionary Relevance. Curr. Plant Biol. 2014, 1, 10–33. [Google Scholar] [CrossRef]
- Pellicer, J.; Garcia, S.; Canela, M.Á.; Garnatje, T.; Korobkov, A.A.; Twibell, J.D.; Vallès, J. Genome Size Dynamics in Artemisia L. (Asteraceae): Following the Track of Polyploidy. Plant Biol. 2010, 12, 820–830. [Google Scholar] [CrossRef]
- Brañas, M.O.; Xirau, J.V. Karyological Studies in Some Taxa of the Genus Artemisia (Asteraceae). Can. J. Bot. 1994, 72, 1126–1135. [Google Scholar] [CrossRef]
- Vallès, J.; McArthur, E.D. Artemisia Systematics and Phylogeny: Cytogenetic and Molecular Insights. In Shrubland Ecosystem Genetics and Biodiversity: Proceedings; McArthur, E.D., Fairbanks, D.J., Eds.; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2001; pp. 67–74. [Google Scholar]
- Vallès, J.; Garnatje, T.; Garcia, S.; Sanz, M.; Korobkov, A.A. Chromosome Numbers in the Tribes Anthemideae and Inuleae (Asteraceae). Bot. J. Linn. Soc. 2005, 148, 77–85. [Google Scholar] [CrossRef]
- McArthur, E.D.; Sanderson, S.C. Cytogeography and Chromosome Evolution of Subgenus Tridentatae of Artemisia (Asteraceae). Am. J. Bot. 1999, 86, 1754–1775. [Google Scholar] [CrossRef]
- Xirau, J.V.; Siljak-Yakovlev, S. Cytogenetic Studies in the Genus Artemisia L. (Asteraceae): Fluorochrome-Banded Karyotypes of Five Taxa, Including the Iberian Endemic Species Artemisia Barrelieri Besser. Can. J. Bot. 1997, 75, 595–606. [Google Scholar] [CrossRef]
- Sanz, M.; Schönswetter, P.; Vallès, J.; Schneeweiss, G.M.; Vilatersana, R. Southern Isolation and Northern Long-Distance Dispersal Shaped the Phylogeography of the Widespread, but Highly Disjunct, European High Mountain Plant Artemisia Eriantha (Asteraceae). Bot. J. Linn. Soc. 2014, 174, 214–226. [Google Scholar] [CrossRef]
- Pace, L.; Pellegrini, M.; Pannunzio, G.; Pirone, G. First Report of Fasciation Symptom in Artemisia Eriantha (Asteraceae), a Typical Orophyte of High-Altitude Cliffs, in Central Apennines (Italy). Plant Sociol. 2020, 57, 23–28. [Google Scholar] [CrossRef]
- Mucciarelli, M.; Maffei, M. Artemisia. In troduction to the Genus. In Artemisia; Wright, C.W., Ed.; Taylor & Francis: London, UK; New York, NY, USA, 2002; pp. 1–50. [Google Scholar]
- Reale, S.; Pace, L.; D’Archivio, A.A.; de Angelis, F.; Marcozzi, G. Volatiles Fingerprint of Artemisia Umbelliformis Subsp. Eriantha by Headspace-Solid Phase Microextraction GC–MS. Nat. Prod. Res. 2014, 28, 61–66. [Google Scholar] [CrossRef]
- Skowyra, M.; Gallego, M.; Segovia, F.; Almajano, M. Antioxidant Properties of Artemisia Annua Extracts in Model Food Emulsions. Antioxidants 2014, 3, 116–128. [Google Scholar] [CrossRef]
- Melguizo-Melguizo, D.; Diaz-de-Cerio, E.; Quirantes-Piné, R.; Švarc-Gajić, J.; Segura-Carretero, A. The Potential of Artemisia Vulgaris Leaves as a Source of Antioxidant Phenolic Compounds. J. Funct. Foods 2014, 10, 192–200. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Nguyen, H.T.; Islam, M.Z.; Obi, T.; Pothinuch, P.; Zar, P.P.K.; Hou, D.X.; Van Nguyen, T.; Nguyen, T.M.; Van Dao, C.; et al. Pharmacological Characteristics of Artemisia Vulgaris L. in Isolated Porcine Basilar Artery. J. Ethnopharmacol. 2016, 182, 16–26. [Google Scholar] [CrossRef]
- Martins, A.; Mignon, R.; Bastos, M.; Batista, D.; Neng, N.R.; Nogueira, J.M.F.; Vizetto-Duarte, C.; Custódio, L.; Varela, J.; Rauter, A.P. In Vitro Antitumoral Activity of Compounds Isolated from Artemisia Gorgonum Webb. Phytother. Res. 2014, 28, 1329–1334. [Google Scholar] [CrossRef]
- Afsar, S.K.; Rajesh Kumar, K.; Venu Gopal, J.; Raveesha, P. Assessment of Anti-Inflammatory Activity of Artemisia Vulgaris Leaves by Cotton Pellet Granuloma Method in Wistar Albino Rats. J. Pharm. Res. 2013, 7, 463–467. [Google Scholar] [CrossRef]
- Corrêa-Ferreira, M.L.; Verdan, M.H.; dos Reis Lívero, F.A.; Galuppo, L.F.; Telles, J.E.Q.; Alves Stefanello, M.É.; Acco, A.; de Oliveira Petkowicz, C.L. Inulin-Type Fructan and Infusion of Artemisia Vulgaris Protect the Liver against Carbon Tetrachloride-Induced Liver Injury. Phytomedicine 2017, 24, 68–76. [Google Scholar] [CrossRef]
- Sefi, M.; Fetoui, H.; Makni, M.; Zeghal, N. Mitigating Effects of Antioxidant Properties of Artemisia Campestris Leaf Extract on Hyperlipidemia, Advanced Glycation End Products and Oxidative Stress in Alloxan-Induced Diabetic Rats. Food Chem. Toxicol. 2010, 48, 1986–1993. [Google Scholar] [CrossRef]
- El-Tantawy, W.H. Biochemical Effects, Hypolipidemic and Anti-Inflammatory Activities of Artemisia Vulgaris Extract in Hypercholesterolemic Rats. J. Clin. Biochem. Nutr. 2015, 57, 33–38. [Google Scholar] [CrossRef]
- Pace, L.; Pellegrini, M.; Palmieri, S.; Rocchi, R.; Lippa, L.; del Gallo, M. Plant Growth-Promoting Rhizobacteria for in Vitro and Ex Vitro Performance Enhancement of Apennines’ Genepì (Artemisia Umbelliformis Subsp. Eriantha), an Endangered Phytotherapeutic Plant. Vitr. Cell. Dev. Biol.-Plant 2020, 56, 134–142. [Google Scholar] [CrossRef]
- Khela, S. Artemisia Umbelliformis. IUCN Red List. Threat. Species 2013, 2013, e.T202935A2758111. [Google Scholar]
- Adorni, M.; Ansaldi, M.; Ardenghi, N.M.G.; Armiraglio, S.; Bacchetta, G.; Bagella, S.; Bedini, G.; Bertani, G.; Bona, E.; Bonali, F. Schede per Una Lista Rossa Della Flora Vascolare e Crittogamica Italiana. Inf. Bot. Ital. 2013, 45, 115–193. [Google Scholar]
- Pieroni, A.; Giusti, M.E. Alpine Ethnobotany in Italy: Traditional Knowledge of Gastronomic and Medicinal Plants among the Occitans of the Upper Varaita Valley, Piedmont. J. Ethnobiol. Ethnomed. 2009, 5, 32. [Google Scholar] [CrossRef]
- Wu, W.-D.; Liu, W.-H.; Sun, M.; Zhou, J.-Q.; Liu, W.; Zhang, C.-L.; Zhang, X.-Q.; Peng, Y.; Huang, L.-K.; Ma, X. Genetic Diversity and Structure of Elymus Tangutorum Accessions from Western China as Unraveled by AFLP Markers. Hereditas 2019, 156, 8. [Google Scholar] [CrossRef]
- Coates, D.J.; Byrne, M.; Moritz, C. Genetic Diversity and Conservation Units: Dealing with the Species-Population Continuum in the Age of Genomics. Front. Ecol. Evol. 2018, 6, 165. [Google Scholar] [CrossRef]
- Garrido-Cardenas, J.A.; Mesa-Valle, C.; Manzano-Agugliaro, F. Trends in Plant Research Using Molecular Markers. Planta 2018, 247, 543–557. [Google Scholar] [CrossRef]
- Paun, O.; Schönswetter, P. Amplified Fragment Length Polymorphism: An Invaluable Fingerprinting Technique for Genomic, Transcriptomic, and Epigenetic Studies. Methods Mol. Biol. 2012, 862, 75–87. [Google Scholar] [CrossRef]
- Ling, N.; Wang, T.; Kuzyakov, Y. Rhizosphere Bacteriome Structure and Functions. Nat. Commun. 2022, 13, 836. [Google Scholar] [CrossRef] [PubMed]
- Xun, W.; Shao, J.; Shen, Q.; Zhang, R. Rhizosphere Microbiome: Functional Compensatory Assembly for Plant Fitness. Comput. Struct. Biotechnol. J. 2021, 19, 5487–5493. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Sansinenea, E. The Role of Beneficial Microorganisms in Soil Quality and Plant Health. Sustainability 2022, 14, 5358. [Google Scholar] [CrossRef]
- Sanon, A.; Andrianjaka, Z.N.; Prin, Y.; Bally, R.; Thioulouse, J.; Comte, G.; Duponnois, R. Rhizosphere Microbiota Interfers with Plant-Plant Interactions. Plant Soil 2009, 321, 259–278. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; el Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van De Lee, T.; Hornes, M.; Friters, A.; Pot, J.; Paleman, J.; Kuiper, M.; et al. AFLP: A New Technique for DNA Fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef]
- Cnops, G.; den Boer, B.; Gerats, A.; van Montagu, M.; Lijsebettens, M. van Chromosome Landing at the Arabidopsis TORNADO1 Locus Using an AFLP-Based Strategy. Mol. Gen. Genet. 1996, 253, 32–41. [Google Scholar] [CrossRef]
- Mizrahi-Man, O.; Davenport, E.R.; Gilad, Y. Taxonomic Classification of Bacterial 16S RRNA Genes Using Short Sequencing Reads: Evaluation of Effective Study Designs. PLoS ONE 2013, 8, e53608. [Google Scholar] [CrossRef]
- Vaccarelli, I.; Matteucci, F.; Pellegrini, M.; Bellatreccia, F.; del Gallo, M. Exploring Microbial Biosignatures in Mn-Deposits of Deep Biosphere: A Preliminary Cross-Disciplinary Approach to Investigate Geomicrobiological Interactions in a Cave in Central Italy. Front. Earth Sci. 2021, 9, 590257. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research—An Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Huff, D.R.; Peakall, R.; Smouse, P.E. RAPD Variation within and among Natural Populations of Outcrossing Buffalograss [Buchloë Dactyloides (Nutt.) Engelm.]. Theor. Appl. Genet. 1993, 86, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Rosenblatt, M. Remarks on Some Nonparametric Estimates of a Density Function. Ann. Math. Stat. 1956, 27, 832–837. [Google Scholar] [CrossRef]
- Parzen, E. On Estimation of a Probability Density Function and Mode. Ann. Math. Stat. 1962, 33, 1065–1076. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT User’s Guide, Version 8; SAS Institute Inc.: Cary, NC, USA, 1999. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software Structure: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; VonHoldt, B.M. STRUCTURE HARVESTER: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Bahrndorff, S.; Alemu, T.; Alemneh, T.; Lund Nielsen, J. The Microbiome of Animals: Implications for Conservation Biology. Int. J. Genom. 2016, 2016, 1–7. [Google Scholar] [CrossRef]
- Blekhman, R.; Goodrich, J.K.; Huang, K.; Sun, Q.; Bukowski, R.; Bell, J.T.; Spector, T.D.; Keinan, A.; Ley, R.E.; Gevers, D.; et al. Host Genetic Variation Impacts Microbiome Composition across Human Body Sites. Genome Biol. 2015, 16, 191. [Google Scholar] [CrossRef]
- Kinross, J.M.; Darzi, A.W.; Nicholson, J.K. Gut Microbiome-Host Interactions in Health and Disease. Genome Med. 2011, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Western, D. Human-Modified Ecosystems and Future Evolution. Proc. Natl. Acad. Sci. USA 2001, 98, 5458–5465. [Google Scholar] [CrossRef] [PubMed]
- Sergaki, C.; Lagunas, B.; Lidbury, I.; Gifford, M.L.; Schäfer, P. Challenges and Approaches in Microbiome Research: From Fundamental to Applied. Front. Plant Sci. 2018, 9, 1205. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S. A Review on Genetic Diversity of Wild Plants by Using Different Genetic Markers. Pure Appl. Biol. 2012, 1, 68–71. [Google Scholar] [CrossRef]
- Rundle, H.D.; Nosil, P. Ecological Speciation. Ecol. Lett. 2005, 8, 336–352. [Google Scholar] [CrossRef]
- Wang, I.J.; Bradburd, G.S. Isolation by Environment. Mol. Ecol. 2014, 23, 5649–5662. [Google Scholar] [CrossRef]
- de Kort, H.; Prunier, J.G.; Ducatez, S.; Honnay, O.; Baguette, M.; Stevens, V.M.; Blanchet, S. Life History, Climate and Biogeography Interactively Affect Worldwide Genetic Diversity of Plant and Animal Populations. Nat. Commun. 2021, 12, 516. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, L.; Xie, X.; Yang, J.; Li, Y.; Zhang, H. Genetic Diversity of Wild Populations of Rheum Tanguticum Endemic to China as Revealed by ISSR Analysis. Biochem. Syst. Ecol. 2010, 38, 264–274. [Google Scholar] [CrossRef]
- Elmeer, K.; Elkhgkheg, A. Genetic Diversity of Artemisia Herba-Alba in Libyan Green Mountain. Iran. J. Sci. Technol. Trans. A Sci. 2019, 43, 1507–1512. [Google Scholar] [CrossRef]
- Shafie, M.S.B.; Hasan, S.M.Z.; Shah, R.M. Study of Genetic Variability of Wormwood Capillary (Artemisia Capillaris) Using Inter Simple Sequence Repeat (ISSR) in Pahang Region, Malaysia. Plant Omics 2009, 2, 127. [Google Scholar]
- Badr, A.; El-Shazly, H.H.; Helail, N.S.; Ghanim, W. el Genetic Diversity of Artemisia Populations in Central and North Saudi Arabia Based on Morphological Variation and RAPD Polymorphism. Plant Syst. Evol. 2012, 298, 871–886. [Google Scholar] [CrossRef]
- Baetz, U.; Martinoia, E. Root Exudates: The Hidden Part of Plant Defense. Trends Plant Sci. 2014, 19, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R.; Gamalero, E. Recent Developments in the Study of Plant Microbiomes. Microorganisms 2021, 9, 1533. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and Heritability of the Maize Rhizosphere Microbiome under Field Conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef] [Green Version]
- Bouffaud, M.-L.; Poirier, M.-A.; Muller, D.; Moënne-Loccoz, Y. Root Microbiome Relates to Plant Host Evolution in Maize and Other Poaceae. Environ. Microbiol. 2014, 16, 2804–2814. [Google Scholar] [CrossRef]
- Szoboszlay, M.; Lambers, J.; Chappell, J.; Kupper, J.V.; Moe, L.A.; McNear, D.H. Comparison of Root System Architecture and Rhizosphere Microbial Communities of Balsas Teosinte and Domesticated Corn Cultivars. Soil Biol. Biochem. 2015, 80, 34–44. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; del Rio, T.G.; et al. Defining the Core Arabidopsis Thaliana Root Microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; ver Loren van Themaat, E.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E.; et al. Revealing Structure and Assembly Cues for Arabidopsis Root-Inhabiting Bacterial Microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The Diversity and Biogeography of Soil Bacterial Communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef]
- Fudou, R.; Jojima, Y.; Iizuka, T.; Yamanaka, S. Haliangium Ochraceum Gen. Nov., Sp. Nov. and Haliangium Tepidum Sp. Nov.: Novel Moderately Halophilic Myxobacteria Isolated from Coastal Saline Environments. J. Gen. Appl. Microbiol. 2002, 48, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Zhao, J.; Ren, T.; Liu, G. Correlation between Soil Microbial Communities and Tobacco Aroma in the Presence of Different Fertilizers. Ind. Crops Prod. 2020, 151, 112454. [Google Scholar] [CrossRef]
- Qiu, M.; Zhang, R.; Xue, C.; Zhang, S.; Li, S.; Zhang, N.; Shen, Q. Application of Bio-Organic Fertilizer Can Control Fusarium Wilt of Cucumber Plants by Regulating Microbial Community of Rhizosphere Soil. Biol. Fertil. Soils 2012, 48, 807–816. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and Interactions in Plant Growth Promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Oberhofer, M.; Hess, J.; Leutgeb, M.; Gössnitzer, F.; Rattei, T.; Wawrosch, C.; Zotchev, S.B. Exploring Actinobacteria Associated With Rhizosphere and Endosphere of the Native Alpine Medicinal Plant Leontopodium Nivale Subspecies Alpinum. Front. Microbiol. 2019, 10, 2531. [Google Scholar] [CrossRef]
- Wei, L.; Ouyang, S.; Wang, Y.; Shen, X.; Zhang, L. Solirubrobacter Phytolaccae Sp. Nov., an Endophytic Bacterium Isolated from Roots of Phytolacca Acinosa Roxb. Int. J. Syst. Evol. Microbiol. 2014, 64, 858–862. [Google Scholar] [CrossRef]
- Farda, B.; Djebaili, R.; Vaccarelli, I.; del Gallo, M.; Pellegrini, M. Actinomycetes from Caves: An Overview of Their Diversity, Biotechnological Properties, and Insights for Their Use in Soil Environments. Microorganisms 2022, 10, 453. [Google Scholar] [CrossRef]
- Dutta, S.; Na, C.S.; Lee, Y.H. Features of Bacterial Microbiota in the Wild Habitat of Pulsatilla Tongkangensis, the Endangered “Long-Sepal Donggang Pasque-Flower Plant,” Endemic to Karst Topography of Korea. Front. Microbiol. 2021, 12, 656105. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
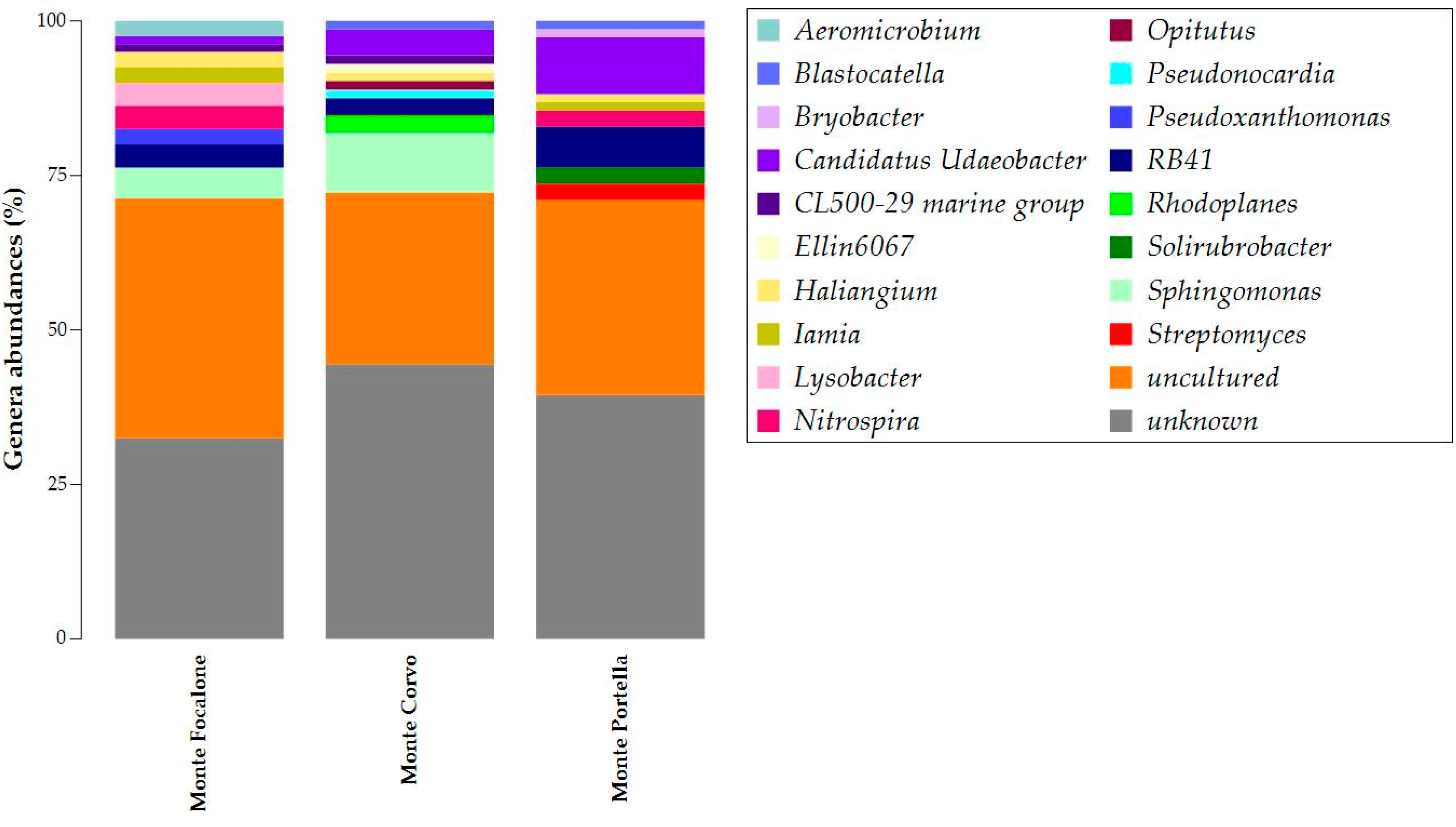

{kind=link}
{kind=link}
{kind=link}
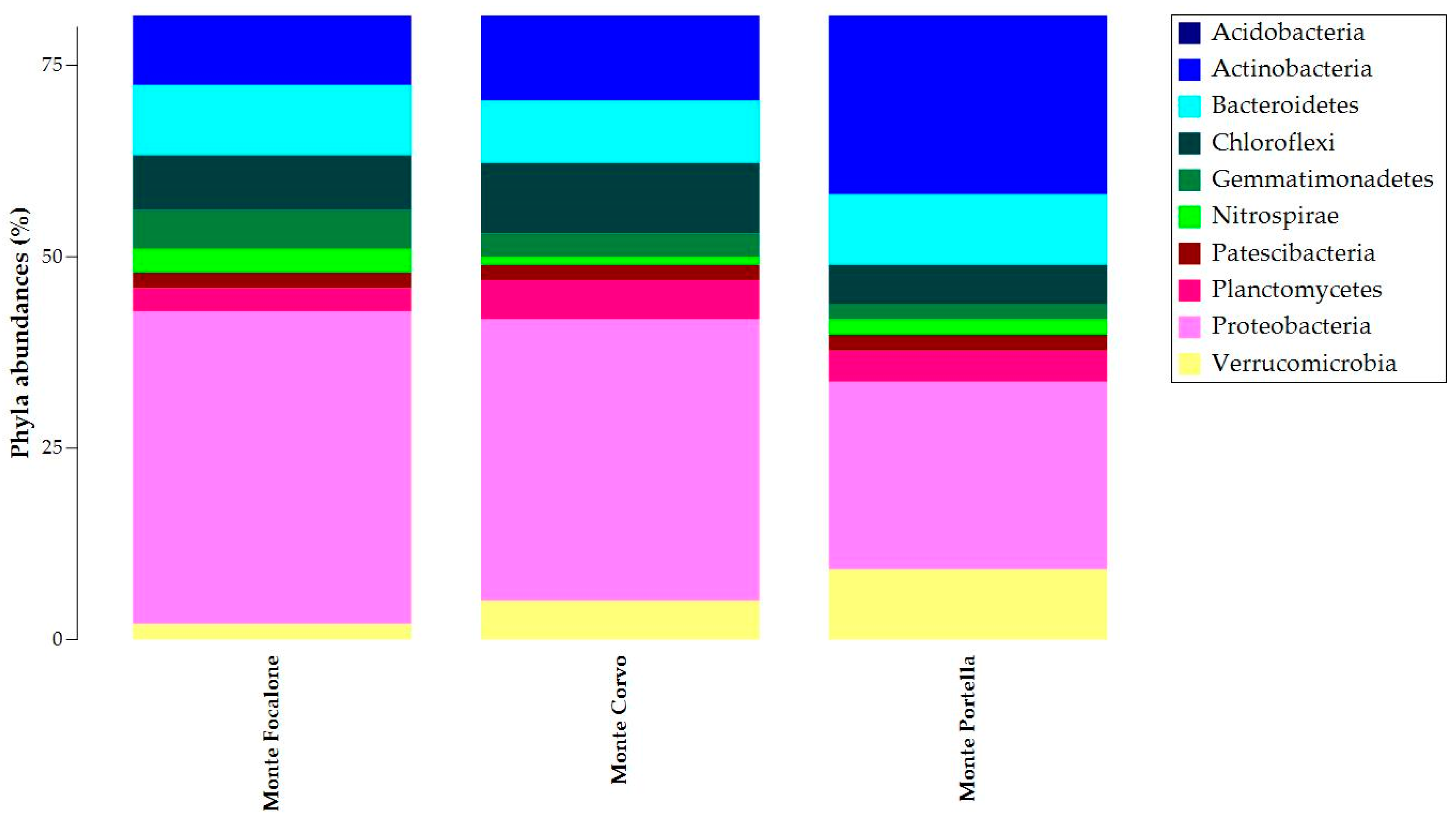{kind=link}
{kind=link}
{kind=link}
| Primer Combination * | Total Fragments | Polymorphic Fragments |
|---|---|---|
| M+AGC/E+CCA | 139 | 128 |
| M+AAC/E+CCA | 126 | 119 |
| M+AGA/E+CAG | 143 | 137 |
| M+ACG/E+CAC | 138 | 132 |
| M+AGA/E+CTG | 128 | 119 |
| M+ACG/E+CAA | 142 | 132 |
| Total | 816 | 767 |
| Richness (ASV Level) | Shannon H′ | Eveness e^H/S | Chao-1 | |
|---|---|---|---|---|
| Monte Focalone | 7713 | 5.57 | 0.59 | 446.4 |
| Monte Corvo | 6486 | 5.75 | 0.70 | 445 |
| Monte Portella | 3671 | 5.32 | 0.74 | 275 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russi, L.; Marconi, G.; Ferradini, N.; Farda, B.; Pellegrini, M.; Pace, L. Investigating Population Genetic Diversity and Rhizosphere Microbiota of Central Apennines’ Artemisia eriantha. Sustainability 2022, 14, 11405. https://doi.org/10.3390/su141811405
Russi L, Marconi G, Ferradini N, Farda B, Pellegrini M, Pace L. Investigating Population Genetic Diversity and Rhizosphere Microbiota of Central Apennines’ Artemisia eriantha. Sustainability. 2022; 14(18):11405. https://doi.org/10.3390/su141811405
Chicago/Turabian StyleRussi, Luigi, Gianpiero Marconi, Nicoletta Ferradini, Beatrice Farda, Marika Pellegrini, and Loretta Pace. 2022. "Investigating Population Genetic Diversity and Rhizosphere Microbiota of Central Apennines’ Artemisia eriantha" Sustainability 14, no. 18: 11405. https://doi.org/10.3390/su141811405
APA StyleRussi, L., Marconi, G., Ferradini, N., Farda, B., Pellegrini, M., & Pace, L. (2022). Investigating Population Genetic Diversity and Rhizosphere Microbiota of Central Apennines’ Artemisia eriantha. Sustainability, 14(18), 11405. https://doi.org/10.3390/su141811405