
Mycorrhiza: An Ecofriendly Bio-Tool for Better Survival of Plants in Nature
by
 , ,
, ,
Mamta Dhiman
1 ,
,
Lakshika Sharma
1,
Prashant Kaushik
2,
Abhijeet Singh
1 and
Madan Mohan Sharma
1,* 1
Department of Biosciences, Manipal University Jaipur, Dehmi Kalan, Near JVK Toll Plaza, Jaipur-Ajmer Expressway, Jaipur 303007, Rajasthan, India
2
Instituto de Conservación y Mejora de la Agrodiversidad Valenciana, Universitat Politècnica de València, 46022 Valencia, Spain
*
Author to whom correspondence should be addressed.
Sustainability 2022, 14(16), 10220; https://doi.org/10.3390/su141610220
Submission received: 27 June 2022
/
Revised: 9 August 2022
/
Accepted: 10 August 2022
/
Published: 17 August 2022
(This article belongs to the Special Issue Study on Influencing Factors of Sustainable Crop Production)
Abstract
:Modern agriculture is currently enduring rapid changes in defiance of the continuing increase of the global population and the various consequent environmental challenges. Crop quality is becoming as important as crop yield and can be characterized by several parameters. Extensive use of chemical fertilizers leads to food safety concerns globally; hence, the use of mycorrhizal symbionts have proven to be beneficial for the sustainable growth of the agricultural cropping system. Microflora inhabiting the soil entails various ecological interactions which are associated with agricultural performances. Amongst these microflora, mycorrhizal fungi are the critical suppliers of nutrients, with restricted diffusion capacities of minerals such as phosphorus, nitrate, zinc, sulfur etc. Mycorrhizae are the obligatory biotrophs that depend upon their host plant for the nutritional requirements. They act as the key contributors to sustainable agro-ecological enforcement and impact globally on the eco-systemic processes. These soil inhabitants devote themselves to the continuous nutrient flow and extemporize resistance against various environmental stresses like drought, flood, metal toxicity, salinity, etc. This review briefly highlights the taxonomic co-evolution, factors affecting mycorrhizal behaviors (phytohormonal regulation), and the concise mechanistic approach (improved water status, photosystems, stomatal conductance, ionic uptake, C & N fixation) to combat various environmental stresses (biotic/abiotic). Plant growth regulators play a crucial role in this symbiotic establishment with the plant roots. Auxins, brassinosteroids, and strigolactones are responsible for the establishment of mycorrhizal association. On the other hand, ethylene, abscisic acid, and jasmonic acids can promote or downregulate this process in the plants. Whereas, gibberellic acids and salicylic acids negatively impact on mycorrhizal association. The hormonal homeostasis (in response to fungal associations) leads to the activation of transcriptional and signaling cascades which ensues various physio-morphological changes for the benefit of the plant. The role of phytohormones in the regulation of plant-fungus mutualism, and the impact of mycorrhization on the activation of molecular and transcriptional cascades, have been described along with the potential applications of agricultural produce and soil rehabilitation.
1. Introduction
Microbial interactions in the rhizosphere are crucial for nutrient recycling, plant growth, and biotic/abiotic stress reduction in forest and agricultural ecosystems. These interactions vary from one plant species to other, at both the inter and intra-specific scales [1]. Among the various microorganisms (bacteria & fungi) involved in the rhizospheric activities, mycorrhizal fungi exhibit the exceptional feature of dwelling partly inside as well as outside the plant roots. The term mycorrhizae comes from the Greek word ‘mykes’ and ‘rhiza’, meaning ‘fungus’ and ‘root’ respectively, which was first applied to the association of trees with fungal symbionts [2]. Mycorrhizal fungi, which are members of Glomeromycota, are common on the landscape and associate with over 80% of plants in a diversity of managed (agricultural) and unmanaged (natural) ecosystems [3]. These fungal symbionts solely rely on the host system to fulfill their carbon requirements, and in exchange provide numerous benefits to the plant system in terms of sustainable nutritional flow, improved plant development, productivity, yield, stress tolerance, water uptake, enzymatic antioxidants accumulation, and soil fertility, etc. [3,4,5]. These microbial communities are enticed towards their symbiotic partners in response to some signaling factors in the form of root exudates released by the plants [6]. Moreover, in response to the mycorrhizal symbiosis, physicochemical as well as molecular alterations in plants leads to improved plant growth, where phytohormones impart considerable impact in regulating the overall process [7]. Mycorrhization benefits plants by up-regulating the catalytic activities of soil enzymes (such as phosphatases, dehydrogenase, nitrogenase, etc.), assisting in the breakdown of complex organic compounds of soil, and positively influencing other microbes present in the rhizosphere for improved nutrients uptake. Activation of these mechanisms, in turn, provides the ability to withstand drought stress, alleviate salinity, helps with micronutrient absorption and better water absorption, and defense systems in the plants [7]. Owing to these benefits, mycorrhizae have gained a lot of consideration towards multidisciplinary research and have huge applications in agriculture as bio-fertilizers, in fuel production due to the increased plant biomass, and in soil rehabilitation, phytoextraction, and phytoremediation, etc. The impact of mycorrhiza on plant survival in extreme environmental conditions, certain factors (phytohormones) that lead in the successful colonization and their mechanistic approach, and other potential applications are discussed in this paper.
This review aims to provide a better understanding about the plant-mycorrhizal interaction which supports sustained plant growth in the agricultural and forest biomes. These associations have been specified as crucial for regenerating over-exploited or lost forest covers. Due to these characteristics, they can be used as bio-tools for the conservation of many overlooked plant species that are significant to various commercial industries.
2. Methodology
A systematic review of relevant literature was conducted to find articles of relevance to the objective of the study. Keywords such as mycorrhizal association, rhizospheric soil microflora, plant–fungal symbiosis, bioremediation, abiotic stress, soil rehabilitation etc. were used to search the data. More than 200 review and research articles published in peer-reviewed journals were studied, and the most suitable were considered to represent the data in the form of a review article. Literature reviews for this report was searched using different databases such as Research gate, Google Scholar, Scopus, Web of Science and reference book records.
3. History
Arbuscule-like structures evolved during the Ordovician Period in the land plant fossils about 430 million years ago, which specifies mycorrhizal growth and successful colonization of land plants [8,9]. Based on the Rhynie chert evaluation, structural resemblances of subterranean organ fossils, and further molecular verifications in available literature concluded that the origin of AMF was in between the Ordovician and Devonian periods. In addition, co-evolution of these mycorrhizal fungi with plants suggests there are advantages of these symbiotic associations in providing increased strength to each other against diverse environmental conditions [6,8]. AMF are relatively older than nitrogen-fixing symbionts and have likely developed by co-opting mycorrhizal signaling components [10]. Originally, arbuscular mycorrhizae were placed under the zygomycetes classification, considering their morphological features of spores as taxonomic indicators. Lately, these taxonomic markers have been replaced by analyzing the molecular variations through rRNA (small subunit) sequencing. A taxonomic assessment using these molecular markers has led to the re-classification of all the AM fungi into a new phylum i.e., Glomeromycota (the sister clade to Ascomycota and Basidiomycota) [11]. Rhizophagus irregularis is the first mycorrhizal fungi with a completely sequenced genome and is the most studied strain in the research field [12]. The classification of AM fungi has been well described from the very beginning up until recent times. It was divided into different timeline periods as: (a) 1845–1974 (the discovery period), (b) 1975–1989 (alpha taxonomy), (c) 1990–2000 (the cladistics period), and (d) 2001–2012 (the phylogenetic period) by [13]. Mycorrhizal evolution is considered to be one of the major revolutions in the development of global land flora. Endomycorrhizae are the most ancient and abundant of the symbiotic associations and are documented based upon fossil records [8,14]. Endomycorrhizae classified under the phylum Glomeromycota are also designated as ‘vesicular-arbuscular mycorrhiza’ (VAM) because of the presence of intracellular structures such as vesicles (storage structures) and arbuscules (branched tree-like structures within roots) (Figure 1). However, due to the ephemeral nature of arbuscular structures, they might often be missing or difficult to observe in the roots collected from soil (owing to the age and color of the roots) [15]. On the other hand, vesicles are present in most of the subsections of mycorrhizal symbionts (except some members of Endogonaceae), therefore, the more appropriate terminology given to this group is ‘arbuscular mycorrhizae’ (AM) [16]. Depending on the morphological and colonization patterns, AM are categorized into “Arum” and “Paris” types (Figure 1). The Arum type describes the linear intercellular spread of hyphal structures within the host roots that form a ramified tree-like arrangement—arbuscules (inside the infected cell) and infection spreads through the side branches penetrating the cortex. The Paris type, on the other hand, describes thick and coiled hyphal growth intracellularly, and infection proceeds from cell to cell through the cortex [15,17]. Another type of AM spread is the “Intermediate” type, where characteristics of both Arum and Paris are present in the infection [6].
After AM infection spread, cortical cells endure structural modifications and develop a periarbuscular membrane (PAM) outside the fungal hyphae. PAM supports trading of nutrients and photosynthetic material between both the symbiotic partners through an “inter-facial apoplastic compartment” (IAC; the gap between arbuscules and PAM) [6]. The majority of mycorrhizae are arbuscular mycorrhizae, which involves the monophyletic Glomales and a broad range of herbaceous and woody plants [14]. Despite their abundance and wide range of relationship with plant species (>80% of terrestrial flora), AMF has shown low species diversity. AMF have high functional diversity because different combinations of host plants and AMF have distinct effects on the numerous aspects of symbiosis [17]. AM are obligatory symbionts and rely on the respective partner for the fulfilment of their carbon requirements, thus the establishment of AM cultures are not possible without any host plant [18]. AM fungi belong to nine genera: Gigaspora, Scutellospora, Glomus, Acaulospora, Entrophospora, Archaeospora, Gerdemannia, Paraglomus and Geosiphon, the only known fungal endosymbiosis with cyanobacteria [19].
These associations are true cosmopolitans and are apparent in all of the ecosystems from tropical forests, Arabic deserts, Arctic regions to elevated Himalayan regions, with few exceptions [20]. Moreover, AM fungi are the key contributors to the defense against various biotic and biotic stresses (Table 1).
The establishment of ectomycorrhiza (ECM) symbiosis occurs through higher fungi mycelia, taxonomically placed under Basidiomycota predominantly, and a few under Ascomycota [51,52]. The most important characteristic of these associations is the non-septate basidium of spore-producing symbionts. These associations usually establish in rootlets of woody flora (bushes or trees) of temperate and some tropic regions. They are mostly categorized under the myrtle family (Myrtaceae), beech family (Fagales), birch family (Betulaceae), rock rose family (Cistaceae), pine family (Pinaceae), willow family (Salicaceae), Dipterocarpaceae and to a lesser extent they are also found in Nyctaginaceae & Polygonaceae [53]. Ectomycorrhizae represent less than 5% of the mycorrhizal associations known in vascular plants, but are ubiquitous in the Pinaceae. Plants under this family dominantly inhabit the diverse climatic conditions and nutrient-deficient soil which are the key influencers of ectomycorrhizal associations. These ECM symbionts are the key players maintaining the nutritional flow through the forests by extending their mycelial network through both the host system and surrounding soils [51]. Ectomycorrhiza form ‘mycelium mantle’ surrounding the host root system (short lateral roots) which further occupies the epidermal and cortical cells without penetrating through the cortex. This results in highly branched mycelial structure to form ‘Hartig net’ [53]. Hartig net plays a critically important role in mutualism by forming the plant-fungus interface through which exchange of the nutrient material between both partners takes place. Besides, mycelia emerging through the root mantle are absolute in hunting the nutrient substances from inundating soil. The ECM colonizing plant range is relatively smaller than AM, despite the fact that owing to the larger forest canopy conquered by these associations, their economic value is steadily growing [19]. The most suitable biological habitat for these associations is parched, non-calcareous, sandy soil regions, swamps, and the lowlands of the Northern Hemisphere [54]. The first impression of such associations is not clear however, because of the obscure preservation in the form of fossil remains attributable to their ephemeral nature, minuscular dimensions, and delicate tissues. Acknowledging that the molecular clock indicates that their first existence is supposed to be from the early Cretaceous period, almost 130 mya [55]. They have evolved as derivatives of saprotrophs and the pattern of their association with the host has evolved autonomously many times [14]. Moreover, the permineralized flora of Princeton chert is apparent for the existence of ectomycorrhizal associations unveiling the ECM structures that are the Hartig net, mantle, and extra-matrical hyphae networks (Figure 1). These findings verify the rise of ECM at least 50 mya [56].
4. Role of Phytohormones in Regulating the Development of Plant-Fungus Symbiotic Association
Phytohormones are the key regulators of all the physicochemical, molecular, and phenotypic expressions of plants. These factors, in very small fractions, facilitate the signal transduction in response to different environmental stimuli. This in turn regulates plant growth, stress resistance, pathogenic resistance, and nutrient flow, etc. [57]. Based on the cellular environment, these molecules can act both in down-regulating or in a synergistic manner. They have been categorized as developmental {auxins (Aux)/cytokinin (Cyt)/gibberellins (GA)/brassinosteroids (BR)/strigolactones (SLs)} and stress resistant {(salicylates (SA)/ethylene (ET) /abscisic acid (ABA)/jasmonates (JA)} hormones. Although, stress-relieving hormones are not limited to this characteristic, and their interaction with growth-regulating hormones control various growth responses and vice-versa [57]. These hormonal interactions flow in a sequential fashion mainly includes: signal input (SI; differential buildup of phytohormones after receiving environmental stimuli), signal processing (SP; a triggered cascade of repressors/stimulators/transcription factors post ligand- receptor binding), and signal output (SO; in the form of phenotypic response post transcriptional changes) [57,58]. Various behavioral responses by different plant hormones have been addressed further with their effects on mycorrhizal colonization (Figure 2). Additionally, significant effects of protein-protein interaction has been evidenced in these regulatory responses [58]. DELLA & JAZ are the key proteins that regulate the GA & Jas pathways, respectively and provide defense and growth benefits to the plants. These interactive regulations (by DELLA & JAZ) are not limited to defense hormones but with other factors also (Aux, ethylene, light, etc.) [59,60].
4.1. Auxin
Several reports are available describing the contribution of the plant hormone to the formation of mycorrhizal associations since the first evidence of auxin involvement in ectomycorrhizal symbiosis was published [61]. Although, auxins are well known for their ability to regulate the plant root system, their role and action mechanism involved in symbiotic association still needs to be explored [62]. It has been reported that the auxin molecules released by the ECM variety enable the successful establishment of plant-mycorrhiza association that results in an improved root system [63]. Also, in Terfezia boudieri and Cistus incanus, auxins and P (phosphorus) govern the type of symbiotic associations (AM or ECM) [64]. Various studies have indicated certain impacts of auxins on inoculation and the colonization abilities of fungal symbiosis. The exogenous use of auxins in mycorrhizae associating plants viz. Papaver croceum and Quercus robur influences greater ECM establishment. Contrary to this, Aux transport inhibitors (TIBA; 2,3,5-triiodobenzoic acid & NPA; 1-N-naphthylphtalamic acid) hamper the mycorrhization process [65,66]. Auxins play a vital role in regulating the structural parameters of roots, which is subjective to the exudation and colonization by the fungal partners [67]. Higher auxin concentrations results in various structural variations in the roots such as attenuated gravitropic progression of taproot, generation of lateral roots to facilitate more colonization [63,68]. Fungal Aux stimulating lateral root proliferation in plants provides more infection sites for the fungal colonization, since the infection occurs closer to the root cap area with a higher probability of mycorrhization. Besides, these small molecules are evident for the increased arbuscular development by activation of various transcriptional mechanisms [6,39,40].
4.2. Strigolactone
Strigolactones (SLs) are plant hormones exuded by plants in the rhizosphere as signaling molecules [69]. They are carotenoid-derived molecules, like the other phytohormones in their action mechanism [10]. Originally, they were discovered as germination stimulating factors of parasitic weed Striga lutea and Orobanche exudates. A few decades later, their role in hyphal branching and the symbiotic association between plant mycorrhiza were witnessed [70,71]. These molecules travel upwardly in plants and down-regulate branching in plants. Their basic action mechanism in symbiosis is identical to the other plant growth regulators, that is, hormone-derived proteolysis [70]. SLs are specifically reported to trigger hyphae formation and branching in mycorrhizal species of Gigasporaceae & Glomeraceae families [10]. Mycorrhizae exhibit extremely sensitive behavior for SLs (GR24; synthetic analog of SL) and hence very low concentrations (approx. 10 nM) are sufficient to establish a symbiotic connection. These molecules recruit the nuclear division, mitochondrial expansion and promote the catalytic performance of NADH dehydrogenases with high ATP production, which are the essential parameters for hyphal growth in Gigaspora rosea [72]. For these activities, the fundamental structure of the intact tricyclic (ABC) lactones and a butanolide (D) ring (attached through an ether linkage) is critical and consistent [10,69]. However, both methylation and demethylation processes are imperative in the varying fungal growth response. Also, the study suggested that the developmental parameters such as hyphal branching may not always signify the symbiotic association between both the partners [73]. Additionally, SLs elicit the production of fungal exudates (myc factors i.e., lipochito- oligosaccharides & chito-oligosaccharides) for the enhanced fungal activity in the symbiotic association process [74]. Due to negligible stability in soil, strigolactones establish a gradient surrounding the roots which provide direction to the mycorrhiza [75]. It has also been reported that the extent of colonization in Petunia mutant plants (SL exporter muted) gets abridged due to the blocked exudation of SLs (orobanchol). Before the colonization process, the growing hyphal tip attaches to the root surface after differentiation into the hyphopodium or appressorium. Formation of a pre-penetration apparatus by the plant cells underneath the hyphopodia takes place for the fungal permeation into the plant. After penetration, fungal hyphae grow inter or intracellularly through the cortical cell layer and form arbuscules which supports nutrient exchange between both partners [76].
4.3. Gibberellin
Gibberellins (GA) are associated with different plant growth stages such as seed germination (breaking seed dormancy), pollen growth, root-shoot elongation, and flower induction, etc. These molecules trigger the signaling cascade which is responsible for the degradation of DELLA TFs (transcription factors) because of poly-ubiquitination by another TF-E3 ubiquitin ligases [77]. Hence, loss of function of the DELLA-TFs is key to express the GA induced response [78]. Based on the available reports, notable evidences have suggested negative impacts of the GA application on AM symbiosis in plant species like (Pisum sativum, Lotus japonicas, Solanum lycopersicum, Oryza sativa, Medicago truncatula, and Triticum etc. [40,50,51]. Although, the rise in GA concentration in AM associated plant Lotus japonicus roots have been reported and increased, GA levels may regulate the hyphal density in the roots by maintaining the arbuscule formation [79]. The pea variety (na-1), which are GA deficient due to the inactive ent-kauremoic enzyme, exhibit high mycorrhizal colonization than the wild type P. sativum plants which authenticates the previous findings [80]. Moreover, stable expression of the DELLA protein (della1-Δ18; non-degradable/ resistant to GA due to the absence of the DELLA domain) is significant in successfully developing mycorrhizae in L. japonicas and M. truncatula [78,79]. Further, GA signaling initiates the formation of arbuscular structures and not the hyphae branching. This is evident in the DELLA mutant forms of rice plant (slr1), P. sativum & M. truncatula (della1 & della2) showing a drastic decline in the arbuscular count (than in other fungal structures) [81,82]. On the other hand, overexpressing Della factors SLR-YFP (O. sativa), Rht1 and Rht2 (Triticum sp.) resulted in a rise in these structures [83].
4.4. Abscisic Acid (ABA)
ABA plays a crucial role in plants with regards to stress management [84,85]. In stress (drought) conditions, ABA levels increase to induce stomatal closure in leaves of mycorrhizae treated plants like Glycine max and Solanum lycopersicum roots [86,87]. ABA also influences mycorrhization in a dose-dependent manner and supports this perception in S. lycopersicum & M. truncatula. Lower concentrations of ABA in plants like S. lycopersicum ABA mutant (sitens) and M. truncatula ABA mutant (PP2A) have shown declined arbuscular branching and fungal penetration into roots, respectively [35]. Besides, in sitens mutant variety, a reduction in AM can be rescued by the application of exogenous ABA. ABA cross-interaction with others suppresses GA (DELLA) cascade, thereby increasing arbuscular formation [75].
There are only a few studies about ABA’s role in AM development and apart from the discussed assumptions, further clarification about the molecular mechanism is still needed [75].
4.5. Jasmonate (JA)
Jasmonic acid, a key defense phytohormone active against both biotic and abiotic stress, has significant role in mycorrhizal symbiosis [75]. JA has been reported to exhibit both positive as well as negative to neutral responses for mycorrhizal symbiosis, where JA cascade is triggered in response to a myc-fungus infection which resulted in enhanced synthesis of JA precursor and genes [88]. Negative consequences on mycorrhizal colonization are due to increased JA in S. lycopersicum, T. majus, O. sativa, and C. papaya, by activating the plant’s defense system and rise in Ca2+ spiking. Similarly, mutant rice plants (cpm2; JA deficient) were reported to have increased mycorrhizal colonization in roots [75,88]. In tomato mutant (def-1) plants (JA deficient), mycorrhizal colonization exhibited positive impacts such as improved resistance to biotic stress (via Spodoptera littoralis) compared to non-inoculated plants (wild type & constitutively JA producing). With JA-accumulating plants, alleviated colonization has been reported [46]. Contrary to this, tomato plants conferring progressive effect of JA on mycorrhizal colonization have also been reported where suppressed JA levels resulted in a delayed mycorrhization process, and vice-versa [89,90]. However, further clarification about such contrasting effect is still needed. One of the possible mechanisms reported for the regulated JA functioning has been the environmental factors (i.e., light conditions). Thus, JA homeostasis plays a crucial role in optimal mycorrhizal colonization [75].
4.6. Brassinosteroid
Brassinosteroids (BRs) are steroidal hormones which regulate different aspects of plant growth such as the development of flower parts (stamen and pollen) through cellular expansion and elongation, vascular system development, fruit ripening, shoot elongation, and pathogenic defense systems, etc. [40,65,66]. Few reports are available which state the role of BRs in mycorrhizal symbiosis in plants such as rice, wheat, tomato, pea etc. In recent findings, it has been concluded that deficiencies of BRs (caused by a mutation in BR synthesizing genes) resulted decline in mycorrhization in mutant rice (brd2-1), pea (lk) and tomato (dx) plants [91,92,93]. Whereas, foliar nourishment of synthetic BRs leads to the improved mycorrhization in wheat [94]. Some studies have suggested the elevated sucrose transport (through a SlSUT2 transporter) and its increased availability to the fungus as a significant factor for improved mycorrhization [91,92]. Recently, it has been reported that the deficient BRs responses and the silencing of the SlSUT2 transporter also decreases mycorrhization in tomatoes, along with reduced pollen development [95]. Further work to demonstrate a detailed understanding behind BRs activity is still required [78].
4.7. Ethylene
Ethylene (ET), a stress hormone, plays an important role in many physiological activities in plants [96]. This volatile phytohormone imparts both positive as well as negative controls on plant growth such as promoted seed germination, fruit ripening, and cell senescence [75]. Very few reports describing the role of ET in mycorrhizal development are available, revealing the regulation of plant immunity and crucial in interactions of the plant with symbiotic or pathogenic microbes. According to the literature surveyed, the elevated concentration of ET is evident in downregulating the mycorrhization process [93]. In a report, this effect was found consistent in the M. truncatula mutant (ein2, ET insensitive) where increased mycorrhizal development in response to ET insensitivity was reported [97]. On the other hand, the ET-insensitive pea plant did not favor mycorrhizal development, neither did it suppress AM development when supplemented with ethylene [93]. In the tomato mutant (ET overproducing), the reduction in the roots colonized with mycorrhizal fungi has also been reported [93]. Moreover, different morphological effects in response to the ethylene have been observed, where ET-ABA interactions down-regulate the intra-radical colonization without affecting hyphopodia on the root [98]. In other plant species like P. sativum, O. sativa, and L. japonicas, unusual hyphopodial structures were observed as an effect of ethylene with restricted root entry by mycorrhizal species [99,100]. The most probable reason suggested for these ethylene-induced effects has been targeting factors involved in Ca2+ spiking cascade and the activation of transcriptional factors and enzymes (amino cyclopropane carboxylate oxidase) involved in the defense system in response to colonization by Glomus fasciculatum [75,98]. It has also been reported that ET signaling is associated with phosphate starvation [101]. Although further research is needed to clarify whether ET reduction occurs to promote intra-radical mycorrhization or because of increased P nutrition by symbiosis [75].
4.8. Salicylic Acid (SA)
SA has been regarded as a stress phytohormone which stimulates endogenous signaling cascade to acquire systemic resistance against pathogens. Signaling also gets induced during mycorrhizal symbiosis, which results in an undesirable impact on mycorrhizal colonization with the host root [6,75]. In tobacco plants (transgenic; exhibit constitutive SA synthesis) due to continuous synthesis of SA, alleviation in colonization has been observed. On the other hand, decreased SA concentrations due to SA hydroxylase activity promoted the colonization [102]. Like ABA, SA reduces mycorrhizal colonization in rice, although no observable effects on hyphopodium development have been reported [103]. Additionally, the SA introduction into rice effects the efficiency of fungal association by lowering the colonization in roots without disturbing the development of appressorium, which indicates indirect influence of SA on fungal growth [6]. Besides, the rise in SA concentration in defective P. sativa (for myc-symbiosis) has been reported. Contrary to this, transgenic tobacco (down-regulated SA production) maintained an increased colonization efficiency [102]. In this context, the suppressing effects of SA on fungal penetration into the host via roots has been suggested [104]. Conclusively, based on the literature available, the application of SA in higher concentrations may reduce or delay the process of successful mycorrhizal colonization in plants. There are only a few reports even now and therefore, the role of SA in mycorrhizal symbiosis still requires further investigation.
5. Applications of Mycorrhizal Symbiosis to the Ecosystem
5.1. Positive Impacts on Plant Growth and Nutritional Requirements
The most prominent assistance provided by symbiotic association of plant-mycorrhizae is to improve growth through the sustainable and enhanced supply of micronutrients. The most evident nutrients involved in this phenomenon is Phosphorous (P) which has additional benefits such as carbon assimilation, regulated enzymatic activities, water retention, and improved soil quality which leads to a positive impact on plant growth [105,106]. AMF are associated with the regulated flow of water and nutrients in exchange of carbohydrates from the host [106]. The mycorrhizal association modifies the morphology of the host roots and improves water-mineral uptake from the rhizosphere [107,108,109]. These associations show varying colonization patterns and capacities depending upon the plant species [110]. AM symbiosis also regulate rhizospheric enzymes such as urease, glucosidases, dehydrogenase, nitrogenase, phosphatase, catalase, peroxidase and soil polyphenol oxidases to provide better soil antioxidant activities [111,112,113,114].
Rhizospheric enzymes improve soil aggregation by hydrolysis and the activation of non-available organic matter in soil, the transfer of nutrients within or between the plants, stabilizing mycorrhizal products like hydrophobins, polysaccharides, glomalin related soil proteins and other extracellular composites, and chelating toxic substances in the rhizosphere [114]. Increased phosphatase activities by mycorrhizal association amplifies levels of phosphorus release from the soil organic matter, hence enhanced translocation of nutrients from the soil to the host plant. In addition, the pattern of intra-radical and extra-radical hyphal structures influence the phosphorus metabolisms among AM species [115]. Conclusively, most of the plants in the natural environment depend on mycorrhizal associations for their nourishment, and these associations have been reported for the transport of about 50% of fixed N and 90% of P into the plant [116,117].
5.2. AMF and Mineral Nutrition
Mycorrhizal symbiosis has gained significant attention with regards to agricultural sustainability due to its characteristic properties of mineral nutrients uptake, utilization, translocation, and how it acts as a biocontrol instrument to the plants. As mentioned in previous sections, they exhibit a critical mediator between the roots and soil, where the soil nutrients acquired by fungal partners get moved to the plant partner in exchange for the photosynthetic carbon produce. Mycelial extensions on the roots’ surface help plants to capture nutrients more efficiently by increasing the surface area, and hence maximum the absorption of soil minerals [118]. The mycorrhizal association triggers the transfer of minerals such as phosphate, ammonium, nitrate, zinc, copper, potassium, sulfur, etc. with the help of various transporters (Table 2) [21,32]. Phosphate transporters (PTs) present in the mycorrhizal fungi due to their high affinity have been extensively studied for their functional and molecular characteristics imparting nutritional benefits towards plant development [4,117]. AM associations have also been reported to promote P uptake cascade in plants, by triggering expression of some phosphate transporters in many plant species such as in M. truncatula (MtPT4), A. sinicus (AsPT1) and O. sativa (OsPT11) [119,120,121]. In this way, phosphate accumulated via mycelial absorption (an active process) is accessible to the plants. These transporter proteins are considered to be indicators of mycorrhizal symbiosis embedded on periarbuscular membranes (PAM) (Figure 3) [122]. Other plant transporter genes for micro and macronutrients like ammonium (AMs), sulfur (SULTF), zinc (ZIPs), nitrate (NPF), potassium (KTs) etc. have also been identified in mycorrhized plants. These transport systems are coupled with a positive impact on arbuscular development as well as a regulatory response to the plant homeostasis [119,123]. In addition, potassium (K+) plays a significant role in plant physiological processes and a symbiotic association with fungus not only increases the potassium supply, but also provide resistance against drought stress to the plant. Potassium accessibility in soil, however, is of concern due to their high mineral adsorption characteristics. Although, these (K+) transporters are associated with myc-symbiosis, their significant physiological involvements have been less explored [123]. Moreover, myc-inoculation into the agricultural sites could soon possibly be an effective method for improved crop productivity, nutritional flow, and regulation of symbiotic associations [116].
5.3. AMF as Bio-Fertilizer
Generally, bio-fertilizers are substances which include microbial population and when applied to the soil, result in improved plant growth by promoting mineral nutrition uptake, water supply, protection against biotic/abiotic stresses, and soil quality. In particular, the fungal microorganisms (due to thin hyphal structures) have emerged as extremely proficient networks with the capabilities of nutrient acquisition from soil inaccessible to the plant roots [135]. Hence, mycorrhizal symbiosis is promising in alleviating limitations related to nutrient uptake [116]. It is also a very interesting fact that the plants invest almost a hundred times of the energy (in C form) required to produce a root than a single hypha which further travels beyond the exhausted nutritional regions of the soil for sustainable nutrient supply. These inferences support the cost-effective nature of the mycorrhizal symbiosis [136]. Mycorrhizal symbiosis is propitious for improved soil texture and other physicochemical properties that result in aggregate formation (in dry or wet conditions), improved soil catalytic performances, proper aeration because of hyphal entanglement, balanced soil pH, etc. Fungal hyphae penetrating deep into the soil form a mesh-like hold upon soil particles and result in micro and macroaggregates formation [137]. Glomalin, the fungal exudate is associated with the formation of these aggregates and helps to hold the soil matrix [136]. These aggregates ultimately provide: (a) protection against soil erosion through heavy wind and water flow, (b) porous texture to the soil, (c) carbon fixation by protecting the organic matter decay by other microbial populations and (d) soil moisture regulation [137,138]. In a recent study, it has been indicated that Glomalin related proteins (product of AMF) help in the restoration of eroded lands by increased soil aggregation and organic carbon sequestration [139]. A variety of mycorrhizal biofertilizers are available on the market (such as Rootplus, Vamstar, Myko-win, Rutmy, Farrata, VAM, Mycoxol, etc.) and have been used widely in agriculture for higher crop yield, production, and soil fertility.
5.4. Mitigation of Biotic & Abiotic Stress
Harsh environmental conditions (abiotic stress) and pathogenic attack (biotic stress) are the major intimidations to global agricultural produce. Negative consequences of these stresses can impede plant growth, nutritional inequities, physiological ailments, ionic toxicity, and cause hormonal imbalance. To overcome the negative consequences, plant adopt several physiological, morphological, structural, and biochemical modifications to alleviate stress [39,47]. A mutualistic association with soil microorganisms promises a stress-tolerant approach towards improved plant defense [26]. From previous reports, it appears that myc-plants exhibit more efficiency in growing under stress conditions [23,28,48]. Reports are available describing stress resistance via: (a) regulated ionic uptake for improved osmoregulation (P↑, N↑, Mn, K↑, Na↓, etc.), (b) up-regulated photosynthetic performance, (c) alleviated oxidative stress, (d) enhanced soil catalytic (mainly phosphatases) activities (for improved availability of mineral elements), (e) the dilution effect on harmful salts/minerals, (f) hormonal balancing, (g) regulation of plant-fungus aquaporin and mineral transporter genes, and (h) elevated water status (Figure. 3) [31,36,45,47]. Different mitigation responses for various biotic/abiotic stresses have been listed (Table 3). However, the mitigation mechanisms for various stress conditions have been debatable and, depending upon the associated myc-plant species, mitigation responses may vary.
5.5. Potential Applications in Phytoremediation
Heavy metal accumulation (Pb, Cd, Hg, Al, Cu, Zn, Cr, etc.) in soil due to natural or human activities has been a serious and persistent environmental threat [143,145,147,158,170]. Owing to their non-degradable nature, these toxic substances pollute the natural resources and the food chains which ultimately reflects adverse effects on the atmospheric, aquatic as well as terrestrial ecosystems. Various chemical and physical remediation techniques are there to overcome the negative effects of heavy metals, while bio-remediation techniques have proved to be more promising in terms of cost-effectiveness and maintaining the soil fertility (by preventing serious soil degradation) [72,122,123]. Some of these heavy metals can be excreted from the body, while others accumulate successively depending upon the exposure, dosage, route, etc. and exhibit chronic behavior [170]. Ubiquitous distribution and abundance of Pb has been one of the hazardous effects on the environment, imposing serious harm to plant growth [146]. Some plant species survive in the presence of heavy metals in the soil, which specifies the presence of some expanded mechanistic approach to adapt in such polluted environments, also called phytoremediation. These plants are designated as hyper-accumulators due to this promising characteristic of accumulating high amounts of heavy metals within the tissue [171,172]. Plants involved in detoxification of soil pollutants usually show sluggish growth, taking a longer time in soil cleaning. While, in combination of mycorrhizal symbiosis plants exhibit a high growth rate, increased biomass, phytochemical activities, and therefore, eradication of soil pollutants at a high rate [173]. However, heavy metals as soil contaminants also hamper colonization and spore formation of mycorrhizal fungi due to increment in root exudate production that limits the supply of carbon sources to the fungal symbionts [174,175]. Mycorrhizal association improves phytoremediation efficiencies. The plant species undergo biological modifications such as increased upward translocation of essential minerals (Zn/Cu) and holding harmful metals (Pb/Cd) in the roots to protect the plant which allows them to survive in extreme abiotic stress [9]. There have been some mechanistic behaviors signifying the removal of toxic substances: metal ion immobilization within mycorrhizal structures; blocking the metal uptake by conversion into non-toxic or ineffective complexes in rhizosphere via chelation, bonding with other biomolecules and precipitation; segregation inside the mycorrhizal vacuole or arbuscules; cytosolic accumulation using biopolymers; stimulating or enhancing antioxidant activities to prevent cellular damage; use of membrane transporters towards or against the concentration gradient for metal translocation; metal ions diffusion to an alleviated response; enhancing nutrient flow to the host; increasing enzymatic efficiencies of soil; stimulating root exudation and up-regulating rhizospheric activities [147,167,169,176]. Further, the impact of mycorrhizal association with hyper-accumulating plant species on phytoremediation efficiencies is depicted (Table 3).
5.6. Enhanced Biological Produce and Agricultural Profitability
Myc-association plays a vital role in managing sustainable plant growth, in addition to improved responses to changing and stressful environmental conditions. As evidenced, the applications of different mycorrhizal species such as G. coronatum, G. mosseae, G. decipiens have been reported for their increased biological yield (cobs per plant, grains, in maize) [110]. Also, these associations have shown enhanced nutritive values through the production of organic (sugars, amino acids) and secondary metabolites (flavonoids, carotenoids, phytochemicals, and volatile organic complexes) [95,111,116,133]. They are responsible for the enhanced C and N fixation, soil fertility, and texture, high food storage, thus a cost-effective approach for the farmers [3]. Another significant factor is the quality yield production, which has been also reported to be enhanced and accompanied by myc-fungi inoculation [81,116,134]. Based on the reported facts, soil microbiota directly influences agricultural profitability [105].
6. Future Prospects
The majority of world flora is associated with the mycorrhizal interactions which contribute to the nutritional or non-nutritional benefits to the host. Based on a systematic literature survey, it can be concluded that mycorrhiza are the key regulators of sustainable ecological performance and contribute to the global flora conservation. To overcome the negative effects of stressful environments, a wide range of mycorrhizal community such as Funneliformis mosseae, Rhizophagus irregularis, Glomuse tunicatum, Glomus intraradices, Glomus mosseae, Acaulospora scrobiculata, and Claroideoglomus etunicatum etc. have been observed to successfully mitigate detrimental effects by stimulating the plant’s defense system. Mycorrhizae, in response to biotic or abiotic stresses, persuade various plant mechanisms such as the activation of defensive proteins (glomalin), toxins (phenolics and alkaloids), hormonal homeostasis, antioxidants (glutathione, carotenoid), and volatile compounds production (prevents from pathogenic attack) etc. The mechanistic approach behind these effects has been described as (a) the elevated uptake of mineral salts (due to activation of plant and fungal transporter proteins present in the epidermis, root hairs, PAM and extraradical hyphae, respectively; (b) up-regulation of phytohormones; (c) induction of aquaporin genes for increased water uptake; (d) dilution of heavy metal toxicity by increased plant biomass; (e) osmoregulation by producing sugars and amino acids; (f) photosystem (PS) improvisation, etc. Additionally, considering their role in the rehabilitation of contaminated soil, increased soil fertility, stress tolerance or mitigation, improved biological produce, activation of beneficial soil microflora, etc., a huge commercial revenue can be obtained in agricultural terms. While phytohormones are well known to regulate the plant root system and increase stress resistance, their role and action mechanism involved in controlled symbiotic association still needs to be explored. In addition, species’ richness of these symbionts and specificity with the host is lesser known due to studies carried out with a limited number of myc-species. Therefore, further studies towards complete understanding of diverse mechanisms underlying mycorrhizal symbiosis are yet to be conducted. In addition, investigation towards the synergistic effects with other microbial moieties can have a great impact. Moreover, the inclination towards ecological habitats is quite diverse, which is of greater interest to the ecological investigations. Transcription and functional analysis have, up to a certain extent, improved the basic understanding of the association between both partners and the taxonomic classification of the mycorrhizal moiety. Although the molecular profiling of mycorrhizae has elevated in the last decade, further investigations are required for a better understanding of developmental and functional molecular strategies associated with mycorrhizal association in plants.
Author Contributions
Conceptualization, M.M.S. and M.D.; methodology, M.D., L.S. and M.M.S.; validation, M.M.S., P.K. and A.S.; formal analysis, M.M.S., P.K. and A.S.; investigation, M.D.; data curation, M.D.; writing—original draft preparation, M.D.; writing—review and editing, M.D. and M.M.S.; supervision, M.M.S.; project administration, M.M.S. and A.S.; funding acquisition, M.M.S. and A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded jointly by NMPB (Govt. of India) and Manipal University Jaipur (research project vide Pr. No. R&D/RAJ03/201-17); UGC, India (SRF fellowship).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors express gratitude to University Grants Commission, New Delhi, India, for financial support (in the form of SRF) sanctioned to Mamta Dhiman; National Medicinal Plants Board, Ministry of AYUSH, New Delhi, Government of India, and Manipal University Jaipur for financial support in the form of research project (vide Pr. No. R&D/RAJ03/201-17); and infrastructural support by MUJ.
Conflicts of Interest
The authors declare no conflict to interest.
Abbreviations:
VAM, vascular arbuscular mycorrhiza; rRNA, ribosomal ribonucleic acid; PAM, periarbuscular membrane; AMF, arbuscular mycorrhizal fungi; ECM, ectomycorrhiza; Myc, mycorrhizal ABA, Abscisic acid; Et, Ethylene; GA, Gibberellin; JA, jasmonic acid; SL, strigolactone; SA, salicylic acid.
References
- Schweitzer, J.A.; Bailey, J.K.; Fischer, D.G.; LeRoy, C.J.; Lonsdorf, E.V.; Whitham, T.G.; Hart, S.C. Plant-soil-microorganism interactions: Heritable relationship between plant genotype and associated soil microorganisms. Ecology 2008, 89, 773–781. [Google Scholar] [CrossRef]
- Frank, B. Über die auf Wurzelsymbiose Beruhende Ernährung Gewisser Bäume Durch Unterirdische Pilze; Berichte der Deutschen Botanischen Gesellschaft: Berlin, Germany, 1885. [Google Scholar]
- Bernardo, L.; Carletti, P.; Badeck, F.W.; Rizza, F.; Morcia, C.; Ghizzoni, R.; Rouphael, Y.; Colla, G.; Terzi, V.; Lucini, L. Metabolomic responses triggered by arbuscular mycorrhiza enhance tolerance to water stress in wheat cultivars. Plant Physiol. Biochem. 2019, 137, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular mycorrhizal fungi as natural biofertilizers: Let’s benefit from past successes. Front. Microbiol. 2016, 6, 1559. [Google Scholar] [CrossRef] [PubMed]
- Abdelhameed, R.E.; Metwally, R.A. Mitigation of salt stress by dual application of arbuscular mycor-rhizal fungi and salicylic acid. Agrochimica 2018, 62, 353–366. [Google Scholar] [CrossRef]
- Liao, D.; Wang, S.; Cui, M.; Liu, J.; Chen, A.; Xu, G. Phytohormones regulate the development of Arbuscular mycorrhizal symbiosis. Int. J. Mol. Sci. 2018, 19, 3146. [Google Scholar] [CrossRef]
- Rao, A.V.; Tak, R. Growth of different tree species and their nutrient uptake in limestone mine spoil as influenced by Arbuscular mycorrhizal (AM)-fungi in Indian arid zone. J. Arid Environ. 2002, 51, 113–119. [Google Scholar] [CrossRef]
- Remy, W.; Taylor, T.N.; Hass, H.; Kerp, H. Four hundred-million-year-old vesicular arbuscular mycorrhizae. Proc. Natl. Acad. Sci. USA 1994, 91, 11841–11843. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, Y.; Ghosh, A.; Song, Y.; Chen, H.; Tang, M. Assessment of Arbuscular mycorrhizal fungi status and heavy metal accumulation characteristics of tree species in a lead–zinc mine area: Potential applications for phytoremediation. Environ. Sci. Pollut. Res. 2015, 22, 13179–13193. [Google Scholar] [CrossRef]
- Gutjahr, C.; Parniske, M. Cell and Developmental Biology of Arbuscular Mycorrhiza Symbiosis. Annu. Rev. Cell Dev. Biol. 2013, 29, 593–617. [Google Scholar] [CrossRef]
- Harrison, M.J. Signaling in the Arbuscular mycorrhizal symbiosis. Annu. Rev. Microbiol. 2005, 59, 19–42. [Google Scholar] [CrossRef]
- Wipf, D.; Krajinski, F.; van Tuinen, D.; Recorbet, G.; Courty, P.E. Trading on the arbuscular mycorrhiza market: From arbuscules to common mycorrhizal networks. New Phytol. 2019, 223, 1127–1142. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Molecular phylogeny and systematics of Glomeromycota: Methods and limitations. Plant Arch. 2018, 18, 1091–1101. [Google Scholar]
- Hlbbett, D.S.; Gilbert, L.B.; Donoghue, M.J. Evolutionary instability of ectomycorrhizal symbioses in basidiomycetes. Nature 2000, 407, 506–508. [Google Scholar] [CrossRef] [PubMed]
- Brundrett, M. Diversity and classification of mycorrhizal associations. Biol. Rev. Camb. Philos. Soc. 2004, 79, 473–495. [Google Scholar] [CrossRef] [PubMed]
- Harrison, M.J. The Arbuscular mycorrhizal symbiosis. In Plant-Microbe Interactions; Springer: Berlin/Heidelberg, Germany, 1997; pp. 1–34. [Google Scholar]
- Prasad, R.; Bhola, D.; Akdi, K.; Cruz, C.; Sairam, K.; Tuteja, N.; Varma, A. Introduction to mycorrhiza: Historical development. In Mycorrhiza-Function, Diversity, State of the Art; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–7. [Google Scholar]
- Chagnon, P.-L. Ecological and evolutionary implications of hyphal anastomosis in Arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 2014, 88, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Varma, A. Symbiosis: The art of living. In Symbiotic Fungi; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–28. [Google Scholar]
- Kehri, H.K.; Akhtar, O.; Zoomi, I.; Pandey, D. Arbuscular mycorrhizal Fungi: Taxonomy and its Systematics. Int. J. Life Sci. Res. 2018, 6, 58–71. [Google Scholar]
- Chen, J.; Zhang, H.; Zhang, X.; Tang, M. Arbuscular mycorrhizal symbiosis alleviates salt stress in black locust through improved photosynthesis, water status, and K+/Na+homeostasis. Front. Plant Sci. 2017, 8, 1739. [Google Scholar] [CrossRef]
- Aroca, R.; Ruiz-Lozano, J.M.; Zamarreño, Á.M.; Paz, J.A.; García-Mina, J.M.; Pozo, M.J.; López-Ráez, J.A. Arbuscular mycorrhizal symbiosis influences strigolactone production under salinity and alleviates salt stress in lettuce plants. J. Plant Physiol. 2013, 170, 47–55. [Google Scholar] [CrossRef]
- Khalloufi, M.; Martínez-Andújar, C.; Lachaâl, M.; Karray-Bouraoui, N.; Pérez-Alfocea, F.; Albacete, A. The interaction between foliar GA3 application and Arbuscular mycorrhizal fungi inoculation improves growth in salinized tomato (Solanum lycopersicum L.) plants by modifying the hormonal balance. J. Plant Physiol. 2017, 214, 134–144. [Google Scholar] [CrossRef]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.B.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd Allah, E.F. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef]
- Hajiboland, R.; Dashtebani, F.; Aliasgharzad, N. Physiological responses of halophytic C4 grass Aeluropus littoralis to salinity and Arbuscular mycorrhizal fungi colonization. Photosynthetica 2015, 53, 572–584. [Google Scholar] [CrossRef]
- Parvin, S.; Van Geel, M.; Yeasmin, T.; Verbruggen, E.; Honnay, O. Effects of single and multiple species inocula of Arbuscular mycorrhizal fungi on the salinity tolerance of a Bangladeshi rice (Oryza sativa L.) cultivar. Mycorrhiza 2020, 30, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Liu, X.; Shan, L.; Wu, Q.; Liu, Y.; Zhang, Z.; Ma, F.; Li, C. Dopamine and Arbuscular mycorrhizal fungi act synergistically to promote apple growth under salt stress. Environ. Exp. Bot. 2020, 178, 104159. [Google Scholar] [CrossRef]
- Hadian-Deljou, M.; Esna-Ashari, M.; Mirzaie-asl, A. Alleviation of salt stress and expression of stress-responsive gene through the symbiosis of Arbuscular mycorrhizal fungi with sour orange seedlings. Sci. Hortic. 2020, 268, 109373. [Google Scholar] [CrossRef]
- Shahvali, R.; Shiran, B.; Ravash, R.; Fallahi, H.; Banović Đeri, B. Effect of symbiosis with Arbuscular mycorrhizal fungi on salt stress tolerance in GF677 (peach × almond) rootstock. Sci. Hortic. 2020, 272, 109535. [Google Scholar] [CrossRef]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Polispitak, K.; Thongpoem, P.; Singh, H.P.; Cha-um, S. Alleviation of Salt Stress in Upland Rice (Oryza sativa L. ssp. indica cv. Leum Pua) Using Arbuscular mycorrhizal Fungi Inoculation. Front. Plant Sci. 2020, 11, 348. [Google Scholar] [CrossRef]
- Liu, C.; Dai, Z.; Cui, M.; Lu, W.; Sun, H. Arbuscular mycorrhizal fungi alleviate boron toxicity in Puccinellia tenuiflora under the combined stresses of salt and drought. Environ. Pollut. 2018, 240, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Volpe, V.; Chitarra, W.; Cascone, P.; Volpe, M.G.; Bartolini, P.; Moneti, G.; Pieraccini, G.; Di Serio, C.; Maserti, B.; Guerrieri, E.; et al. The Association With Two Different Arbuscular mycorrhizal Fungi Differently Affects Water Stress Tolerance in Tomato. Front. Plant Sci. 2018, 9, 1480. [Google Scholar] [CrossRef]
- Pavithra, D.; Yapa, N. Arbuscular mycorrhizal fungi inoculation enhances drought stress tolerance of plants. Groundw. Sustain. Dev. 2018, 7, 490–494. [Google Scholar] [CrossRef]
- Saia, S.; Amato, G.; Frenda, A.S.; Giambalvo, D.; Ruisi, P. Influence of arbuscular mycorrhizae on biomass production and nitrogen fixation of berseem clover plants subjected to water stress. PLoS ONE 2014, 9, e90738. [Google Scholar] [CrossRef]
- Chitarra, W.; Pagliarani, C.; Maserti, B.; Lumini, E.; Siciliano, I.; Cascone, P.; Schubert, A.; Gambino, G.; Balestrini, R.; Guerrieri, E. Insights on the impact of Arbuscular mycorrhizal symbiosis on tomato tolerance to water stress. Plant Physiol. 2016, 171, 1009–1023. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, G.; Erice, G.; Aroca, R.; Chaumont, F.; Ruiz-Lozano, J.M. Enhanced drought stress tolerance by the Arbuscular mycorrhizal symbiosis in a drought-sensitive maize cultivar is related to a broader and differential regulation of host plant aquaporins than in a drought-tolerant cultivar. Front. Plant Sci. 2017, 8, 1056. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Tomar, R.S.; Jajoo, A. Arbuscular mycorrhizal fungi (AMF) protects photosynthetic apparatus of wheat under drought stress. Photosynth. Res. 2019, 139, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Polanco, M.; Sánchez-Romera, B.; Aroca, R.; Asins, M.J.; Declerck, S.; Dodd, I.C.; Martínez-Andújar, C.; Albacete, A.; Ruiz-Lozano, J.M. Exploring the use of recombinant inbred lines in combination with beneficial microbial inoculants (AM fungus and PGPR) to improve drought stress tolerance in tomato. Environ. Exp. Bot. 2016, 131, 47–57. [Google Scholar] [CrossRef]
- Mathur, S.; Jajoo, A. Arbuscular mycorrhizal fungi protects maize plants from high temperature stress by regulating photosystem II heterogeneity. Ind. Crops Prod. 2020, 143, 111934. [Google Scholar] [CrossRef]
- Mathur, S.; Sharma, M.P.; Jajoo, A. Improved photosynthetic efficacy of maize (Zea mays) plants with Arbuscular mycorrhizal fungi (AMF) under high temperature stress. J. Photochem. Photobiol. B Biol. 2018, 180, 149–154. [Google Scholar] [CrossRef]
- Cabral, C.; Ravnskov, S.; Tringovska, I.; Wollenweber, B. Arbuscular mycorrhizal fungi modify nutrient allocation and composition in wheat (Triticum aestivum L.) subjected to heat-stress. Plant Soil 2016, 408, 385–399. [Google Scholar] [CrossRef]
- Hajiboland, R.; Joudmand, A.; Aliasgharzad, N.; Tolrá, R.; Poschenrieder, C. Arbuscular mycorrhizal fungi alleviate low-temperature stress and increase freezing resistance as a substitute for acclimation treatment in barley. Crop Pasture Sci. 2019, 70, 218–233. [Google Scholar] [CrossRef]
- Li, S.; Yang, W.; Guo, J.; Li, X.; Lin, J.; Zhu, X. Changes in photosynthesis and respiratory metabolism of maize seedlings growing under low temperature stress may be regulated by Arbuscular mycorrhizal fungi. Plant Physiol. Biochem. 2020, 154, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pasbani, B.; Salimi, A.; Aliasgharzad, N.; Hajiboland, R. Colonization with Arbuscular mycorrhizal fungi mitigates cold stress through improvement of antioxidant defense and accumulation of protecting molecules in eggplants. Sci. Hortic. 2020, 272, 109575. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; Mehralian, M. Arbuscular mycorrhizal Fungi Inoculation to Enhance Chilling Stress Tolerance of Watermelon. Gesunde Pflanz. 2020, 72, 171–179. [Google Scholar] [CrossRef]
- Formenti, L.; Rasmann, S. Mycorrhizal fungi enhance resistance to herbivores in tomato plants with reduced jasmonic acid production. Agronomy 2019, 9, 131. [Google Scholar] [CrossRef]
- Song, Y.Y.; Ye, M.; Li, C.Y.; Wang, R.L.; Wei, X.C.; Luo, S.M.; Zeng, R. Sen Priming of Anti-Herbivore Defense in Tomato by Arbuscular mycorrhizal Fungus and Involvement of the Jasmonate Pathway. J. Chem. Ecol. 2013, 39, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Balestrini, R.; Rosso, L.C.; Veronico, P.; Melillo, M.T.; De Luca, F.; Fanelli, E.; Colagiero, M.; Di Fossalunga, A.S.; Ciancio, A.; Pentimone, I. Transcriptomic responses to water deficit and nematode infection in mycorrhizal tomato roots. Front. Microbiol. 2019, 10, 1807. [Google Scholar] [CrossRef] [PubMed]
- Marquez, N.; Giachero, M.L.; Gallou, A.; Debat, H.J.; Declerck, S.; Ducasse, D.A. Transcriptome analysis of mycorrhizal and nonmycorrhizal soybean plantlets upon infection with Fusarium virguliforme, one causal agent of sudden death syndrome. Plant Pathol. 2019, 68, 470–480. [Google Scholar] [CrossRef]
- Hao, Z.; Fayolle, L.; van Tuinen, D.; Chatagnier, O.; Li, X.; Gianinazzi, S.; Gianinazzi-Pearson, V. Local and systemic mycorrhiza-induced protection against the ectoparasitic nematode Xiphinema index involves priming of defence gene responses in grapevine. J. Exp. Bot. 2012, 63, 3657–3672. [Google Scholar] [CrossRef]
- Anderson, I.C.; Cairney, J.W.G. Ectomycorrhizal fungi: Exploring the mycelial frontier. FEMS Microbiol. Rev. 2007, 31, 388–406. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010; ISBN 0080559344. [Google Scholar]
- Balestrini, R.; Kottke, I. Structure and development of ectomycorrhizal roots. In Molecular Mycorrhizal Symbiosis; John Wiley & Sons: Hoboken, NJ, USA, 2016; ISBN 9781118951446. [Google Scholar]
- Agerer, R. Exploration types of ectomycorrhizae: A proposal to classify ectomycorrhizal mycelial systems according to their patterns of differentiation and putative ecological importance. Mycorrhiza 2001, 11, 107–114. [Google Scholar] [CrossRef]
- Berbee, M.L.; Taylor, J.W. Dating the evolutionary radiations of the true fungi. Can. J. Bot. 1993, 71, 1114–1127. [Google Scholar] [CrossRef]
- Lepage, B.A.; Currah, R.S.; Stockey, R.A.; Rothwell, G.W. Fossil ectomycorrhizae from the middle Eocene. Am. J. Bot. 1997, 84, 410–412. [Google Scholar] [CrossRef]
- Pozo, M.J.; López-Ráez, J.A.; Azcón-Aguilar, C.; García-Garrido, J.M. Phytohormones as integrators of environmental signals in the regulation of mycorrhizal symbioses. New Phytol. 2015, 205, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Shabek, N.; Zheng, N. Plant ubiquitin ligases as signaling hubs. Nat. Struct. Mol. Biol. 2014, 21, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Ballaré, C.L. Light Regulation of Plant Defense. Annu. Rev. Plant Biol. 2014, 65, 335–363. [Google Scholar] [CrossRef]
- Xu, H.; Liu, Q.; Yao, T.; Fu, X. Shedding light on integrative GA signaling. Curr. Opin. Plant Biol. 2014, 21, 89–95. [Google Scholar] [CrossRef] [PubMed]
- MacDougal, D.T.; Dufrenoy, J. Mycorrhizal Symbiosis In Aplectrum, Corallorhiza and Pinus. Plant Physiol. 1944, 19, 440–465. [Google Scholar] [CrossRef]
- Sitrit, Y.; Roth-Bejerano, N.; Kagan-Zur, V.; Turgeman, T. Pre-symbiotic Interactions Between the Desert Truffle Terfezia boudieri and Its Host Plant Helianthemum sessiliflorum. In Desert Truffles Phylogeny, Physiology, Distribution and Domestication; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Felten, J.; Kohler, A.; Morin, E.; Bhalerao, R.P.; Palme, K.; Martin, F.; Ditengou, F.A.; Legué, V. The ectomycorrhizal fungus laccaria bicolor stimulates lateral root formation in poplar and arabidopsis through auxin transport and signaling. Plant Physiol. 2009, 151, 1991–2005. [Google Scholar] [CrossRef]
- Zaretsky, M.; Kagan-Zur, V.; Mills, D.; Roth-Bejerano, N. Analysis of mycorrhizal associations formed by Cistus incanus transformed root clones with Terfezia boudieri isolates. Plant Cell Rep. 2006, 25, 62–70. [Google Scholar] [CrossRef]
- Gay, G.; Normand, L.; Marmeisse, R.; Sotta, B.; Debaud, J.C. Auxin overproducer mutants of Hebeloma cylindrosporum Romagnesi have increased mycorrhizal activity. New Phytol. 1994, 128, 645–657. [Google Scholar] [CrossRef]
- Herrmann, S.; Oelmüller, R.; Buscot, F. Manipulation of the onset of ectomycorrhiza formation by indole-3-acetic acid, activated charcoal or relative humidity in the association between oak microcuttings and Piloderma croceum: Influence on plant development and photosynthesis. J. Plant Physiol. 2004, 161, 509–517. [Google Scholar] [CrossRef]
- Gutjahr, C.; Paszkowski, U. Multiple control levels of root system remodeling in Arbuscular mycorrhizal symbiosis. Front. Plant Sci. 2013, 4, 204. [Google Scholar] [CrossRef]
- Splivallo, R.; Fischer, U.; Göbel, C.; Feussner, I.; Karlovsky, P. Truffles regulate plant root morphogenesis via the production of auxin and ethylene. Plant Physiol. 2009, 150, 2018–2029. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Atarashi, T.; Kuse, M.; Sugimoto, Y.; Takikawa, H. Concise synthesis of heliolactone, a non-canonical strigolactone isolated from sunflower. Biosci. Biotechnol. Biochem. 2020, 84, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.; Ruyter-Spira, C.; Bouwmeester, H.J. Strigolactones and root infestation by plant-parasitic Striga, Orobanche and Phelipanche spp. Plant Sci. 2011, 180, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Matsuzaki, K.I.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in Arbuscular mycorrhizal fungi. Nature 2005, 435, 824–827. [Google Scholar] [CrossRef] [PubMed]
- Besserer, A.; Puech-Pagès, V.; Kiefer, P.; Gomez-Roldan, V.; Jauneau, A.; Roy, S.; Portais, J.C.; Roux, C.; Bécard, G.; Séjalon-Delmas, N. Strigolactones stimulate Arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biol. 2006, 4, e226. [Google Scholar] [CrossRef]
- Taulera, Q.; Lauressergues, D.; Martin, K.; Cadoret, M.; Servajean, V.; Boyer, F.D.; Rochange, S. Initiation of Arbuscular mycorrhizal symbiosis involves a novel pathway independent from hyphal branching. Mycorrhiza 2020, 30, 491–501. [Google Scholar] [CrossRef]
- Genre, A.; Chabaud, M.; Balzergue, C.; Puech-Pagès, V.; Novero, M.; Rey, T.; Fournier, J.; Rochange, S.; Bécard, G.; Bonfante, P.; et al. Short-chain chitin oligomers from Arbuscular mycorrhizal fungi trigger nuclear Ca2+ spiking in Medicago truncatula roots and their production is enhanced by strigolactone. New Phytol. 2013, 198, 190–202. [Google Scholar] [CrossRef]
- Das, D.; Gutjahr, C. Role of phytohormones in arbuscular mycorrhiza development. In The Model Legume Medicago Truncatula; Wiley: Hoboken, NJ, USA, 2020. [Google Scholar]
- Pimprikar, P.; Gutjahr, C. Transcriptional Regulation of Arbuscular Mycorrhiza Development. Plant Cell Physiol. 2018, 59, 678–695. [Google Scholar] [CrossRef]
- Davière, J.M.; Achard, P. Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [Google Scholar] [CrossRef]
- McGuiness, P.N.; Reid, J.B.; Foo, E. The role of gibberellins and brassinosteroids in nodulation and Arbuscular mycorrhizal associations. Front. Plant Sci. 2019, 10, 269. [Google Scholar] [CrossRef]
- Takeda, N.; Handa, Y.; Tsuzuki, S.; Kojima, M.; Sakakibara, H.; Kawaguchi, M. Gibberellins interfere with symbiosis signaling and gene expression and alter colonization by Arbuscular Mycorrhizal fungi in Lotus Japonicus. Plant Physiol. 2015, 167, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Foo, E. Auxin influences strigolactones in pea mycorrhizal symbiosis. J. Plant Physiol. 2013, 170, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Floss, D.S.; Levy, J.G.; Lévesque-Tremblay, V.; Pumplin, N.; Harrison, M.J. DELLA proteins regulate arbuscule formation in Arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2013, 110, E5025–E5034. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Luo, D.; Zhang, X.; Liu, J.; Wang, W.; Jin, Y.; Dong, W.; Liu, J.; Liu, H.; Yang, W.; et al. A della protein complex controls the Arbuscular mycorrhizal symbiosis in plants. Cell Res. 2014, 24, 130–133. [Google Scholar] [CrossRef]
- Carbonnel, S.; Gutjahr, C. Control of arbuscular mycorrhiza development by nutrient signals. Front. Plant Sci. 2014, 5, 462. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef]
- Finkelstein, R. Abscisic Acid Synthesis and Response. Arab. B. 2013, 11, e0166. [Google Scholar] [CrossRef]
- Martín-Rodríguez, J.A.; Huertas, R.; Ho-Plágaro, T.; Ocampo, J.A.; Čková, V.T.; Tarkowská, D.; Ludwig-Müller, J.; García-Garrido, J.M. Gibberellin-abscisic acid balances during arbuscular mycorrhiza formation in tomato. Front. Plant Sci. 2016, 7, 1273. [Google Scholar] [CrossRef]
- Meixner, C.; Ludwig-Müller, J.; Miersch, O.; Gresshoff, P.; Staehelin, C.; Vierheilig, H. Lack of mycorrhizal autoregulation and phytohormonal changes in the supernodulating soybean mutant nts1007. Planta 2005, 222, 709–715. [Google Scholar] [CrossRef]
- Gutjahr, C.; Siegler, H.; Haga, K.; Iino, M.; Paszkowski, U. Full establishment of Arbuscular mycorrhizal symbiosis in rice occurs independently of enzymatic jasmonate biosynthesis. PLoS ONE 2015, 10, e0123422. [Google Scholar] [CrossRef]
- Isayenkov, S.; Mrosk, C.; Stenzel, I.; Strack, D.; Hause, B. Suppression of allene oxide cyclase in hairy roots of Medicago truncatula reduces jasmonate levels and the degree of mycorrhization with Glomus intraradices. Plant Physiol. 2005, 139, 1401–1410. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, R.; Schaarschmidt, S.; Hause, B. Repeated leaf wounding alters the colonization of Medicago truncatula roots by beneficial and pathogenic microorganisms. Plant, Cell Environ. 2012, 35, 1344–1357. [Google Scholar] [CrossRef] [PubMed]
- Bitterlich, M.; Krügel, U.; Boldt-Burisch, K.; Franken, P.; Kühn, C. Interaction of brassinosteroid functions and sucrose transporter SlSUT2 regulate the formation of arbuscular mycorrhiza. Plant Signal. Behav. 2014, 9, e970426. [Google Scholar] [CrossRef] [PubMed]
- Bitterlich, M.; Krügel, U.; Boldt-Burisch, K.; Franken, P.; Kühn, C. The sucrose transporter SlSUT2 from tomato interacts with brassinosteroid functioning and affects arbuscular mycorrhiza formation. Plant J. 2014, 78, 877–889. [Google Scholar] [CrossRef] [PubMed]
- Foo, E.; McAdam, E.L.; Weller, J.L.; Reid, J.B. Interactions between ethylene, gibberellins, and brassinosteroids in the development of rhizobial and mycorrhizal symbioses of pea. J. Exp. Bot. 2016, 67, 2413–2424. [Google Scholar] [CrossRef] [PubMed]
- Tofighi, C.; Khavari-Nejad, R.A.; Najafi, F.; Razavi, K.; Rejali, F. Responses of wheat plants to interactions of 24-epibrassinolide and Glomus mosseae in saline condition. Physiol. Mol. Biol. Plants 2017, 23, 557–564. [Google Scholar] [CrossRef]
- Hansch, F.; Jaspar, H.; von Sivers, L.; Bitterlich, M.; Franken, P.; Kühn, C. Brassinosteroids and sucrose transport in mycorrhizal tomato plants. Plant Signal. Behav. 2020, 15, 1714292. [Google Scholar] [CrossRef]
- Zhang, W.; Hu, Y.; Liu, J.; Wang, H.; Wei, J.; Sun, P.; Wu, L.; Zheng, H. Progress of ethylene action mechanism and its application on plant type formation in crops. Saudi J. Biol. Sci. 2020, 27, 1667–1673. [Google Scholar] [CrossRef]
- Varma Penmetsa, R.; Uribe, P.; Anderson, J.; Lichtenzveig, J.; Gish, J.C.; Nam, Y.W.; Engstrom, E.; Xu, K.; Sckisel, G.; Pereira, M.; et al. The Medicago truncatula ortholog of Arabidopsis EIN2, sickle, is a negative regulator of symbiotic and pathogenic microbial associations. Plant J. 2008, 55, 580–595. [Google Scholar] [CrossRef]
- Fracetto, G.G.M.; Peres, L.E.P.; Lambais, M.R. Gene expression analyses in tomato near isogenic lines provide evidence for ethylene and abscisic acid biosynthesis fine-tuning during arbuscular mycorrhiza development. Arch. Microbiol. 2017, 199, 787–798. [Google Scholar] [CrossRef]
- Geil, R.D.; Peterson, R.L.; Guinel, F.C. Morphological alterations of pea (Pisum sativum cv. Sparkle) arbuscular mycorrhizas as a result of exogenous ethylene treatment. Mycorrhiza 2001, 11, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Gutjahr, C.; Banba, M.; Croset, V.; An, K.; Miyao, A.; An, G.; Hirochika, H.; Imaizumi-Anraku, H.; Paszkowski, U. Arbuscular Mycorrhiza-Specific Signaling in Rice Transcends the Common Symbiosis Signaling Pathway. Plant Cell 2008, 20, 2989–3005. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xie, Y.; Wang, H.; Ma, X.; Yao, W.; Wang, H. Light and ethylene coordinately regulate the phosphate starvation response through transcriptional regulation of PHOSPHATE STARVATION RESPONSE1. Plant Cell 2017, 29, 2269–2284. [Google Scholar] [CrossRef] [PubMed]
- Herrera Medina, M.J.; Gagnon, H.; Piché, Y.; Ocampo, J.A.; García Garrido, J.M.; Vierheilig, H. Root colonization by Arbuscular mycorrhizal fungi is affected by the salicylic acid content of the plant. Plant Sci. 2003, 164, 993–998. [Google Scholar] [CrossRef]
- Blilou, I.; Ocampo, J.A.; García-Garrido, J.M. Induction of LTP (lipid transfer protein) and Pal (phenylalanine ammonia-lyase) gene expression in rice roots colonized by the Arbuscular mycorrhizal fungus Glomus mosseae. J. Exp. Bot. 2000, 51, 1969–1977. [Google Scholar] [CrossRef]
- Gutjahr, C.; Paszkowski, U. Weights in the balance: Jasmonic acid and Salicylic acid signaling in root-biotroph interactions. Mol. Plant-Microbe Interact. 2009, 22, 763–772. [Google Scholar] [CrossRef]
- Rillig, M.C.; Aguilar-Trigueros, C.A.; Camenzind, T.; Cavagnaro, T.R.; Degrune, F.; Hohmann, P.; Lammel, D.R.; Mansour, I.; Roy, J.; van der Heijden, M.G.A. Why farmers should manage the Arbuscular mycorrhizal symbiosis. New Phytol. 2019, 222, 1171–1175. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Ericoid mycorrhizas. In Mycorrhizal Symbiosis; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Dhiman, M.; Sharma, L.; Singh, A.; Sharma, M.M. Exsitu Conservation Using In vitro Methods of an Endangered Plant Sterculia urens Roxb.: A High Volume Trade Plant for Gum Karaya. Ind. Crops Prod. 2020, 158, 113015. [Google Scholar] [CrossRef]
- Sharma, S.; Anand, G.; Singh, N.; Kapoor, R. Arbuscular mycorrhiza augments arsenic tolerance in wheat (Triticum aestivum L.) by strengthening antioxidant defense system and thiol metabolism. Front. Plant Sci. 2017, 8, 906. [Google Scholar] [CrossRef]
- Zhang, Q.; Gong, M.; Liu, K.; Chen, Y.; Yuan, J.; Chang, Q. Rhizoglomus intraradices Improves Plant Growth, Root Morphology and Phytohormone Balance of Robinia pseudoacacia in Arsenic-Contaminated Soils. Front. Microbiol. 2020, 11, 1428. [Google Scholar] [CrossRef]
- Singh, M.; Beura, K.; Pradhan, A.K.; Rakshit, R.; Lal, M. Ability of arbuscular mycorrhiza to promote growth of maize plant and enzymatic activity of an alluvial soil. J. Appl. Nat. Sci. 2015, 7, 1029–1035. [Google Scholar] [CrossRef]
- Pritsch, K.; Garbaye, J. Enzyme secretion by ECM fungi and exploitation of mineral nutrients from soil organic matter. Ann. For. Sci. 2011, 68, 25–32. [Google Scholar] [CrossRef]
- Gianfreda, L. Enzymes of importance to rhizosphere processes. J. Soil Sci. Plant Nutr. 2015, 15, 283–306. [Google Scholar] [CrossRef]
- Wang, F. Occurrence of Arbuscular mycorrhizal fungi in mining-impacted sites and their contribution to ecological restoration: Mechanisms and applications. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1901–1957. [Google Scholar] [CrossRef]
- Lü, L.H.; Wu, Q.S. Mitigation of replant disease by mycorrhization in horticultural plants: A review. Folia Hortic. 2018, 30, 269–282. [Google Scholar] [CrossRef]
- Boddington, C.L.; Dodd, J.C. Evidence that differences in phosphate metabolism in mycorrhizas formed by species of Glomus and Gigaspora might be related to their life-cycle strategies. New Phytol. 1999, 142, 531–538. [Google Scholar] [CrossRef]
- Nouri, E.; Breuillin-Sessoms, F.; Feller, U.; Reinhardt, D. Correction: Phosphorus and nitrogen regulate Arbuscular mycorrhizal symbiosis in Petunia hybrida. PLoS ONE 2015, 10, e0127472. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef]
- Odoh, C.K.; Eze, C.N.; Obi, C.J.; Anyah, F.; Egbe, K.; Unah, U.V.; Akpi, U.K.; Adobu, U.S. Fungal Biofertilizers for Sustainable Agricultural Productivity. In Agriculturally Important Fungi for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2020; pp. 199–225. [Google Scholar]
- Harrison, M.J.; Dewbre, G.R.; Liu, J. A phosphate transporter from Medicago truncatula involved in the acquisition of phosphate released by Arbuscular mycorrhizal fungi. Plant Cell 2002, 14, 2413–2429. [Google Scholar] [CrossRef]
- Paszkowski, U.; Kroken, S.; Roux, C.; Briggs, S.P. Rice phosphate transporters include an evolutionarily divergent gene specifically activated in Arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2002, 99, 13324–13329. [Google Scholar] [CrossRef]
- Xie, X.; Huang, W.; Liu, F.; Tang, N.; Liu, Y.; Lin, H.; Zhao, B. Functional analysis of the novel mycorrhiza-specific phosphate transporter AsPT1 and PHT1 family from Astragalus sinicus during the Arbuscular mycorrhizal symbiosis. New Phytol. 2013, 198, 836–852. [Google Scholar] [CrossRef] [PubMed]
- Bapaume, L.; Reinhardt, D. How membranes shape plant symbioses: Signaling and transport in nodulation and arbuscular mycorrhiza. Front. Plant Sci. 2012, 3, 223. [Google Scholar] [CrossRef] [PubMed]
- Garcia, K.; Zimmermann, S.D. The role of mycorrhizal associations in plant potassium nutrition. Front. Plant Sci. 2014, 5, 337. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.D.; Hu, X.C.; Wu, Q.S.; Yang, T.Y.; Srivastava, A.K.; Zhang, D.J.; Gao, X.B.; Kuča, K. Mycorrhizas promote P acquisition of tea plants through changes in root morphology and P transporter gene expression. S. Afr. J. Bot. 2021, 137, 455–462. [Google Scholar] [CrossRef]
- Ma, X.; Li, X.; Ludewig, U. Arbuscular mycorrhizal colonization outcompetes root hairs in maize under low phosphorus availability. Ann. Bot. 2021, 127, 155–166. [Google Scholar] [CrossRef]
- Volpe, V.; Giovannetti, M.; Sun, X.G.; Fiorilli, V.; Bonfante, P. The phosphate transporters LjPT4 and MtPT4 mediate early root responses to phosphate status in non mycorrhizal roots. Plant Cell Environ. 2016, 39, 660–671. [Google Scholar] [CrossRef]
- Chorianopoulou, S.N.; Sigalas, P.P.; Tsoutsoura, N.; Apodiakou, A.; Saridis, G.; Ventouris, Y.E.; Bouranis, D.L. Regulation of sulfur homeostasis in mycorrhizal maize plants grown in a fe-limited environment. Int. J. Mol. Sci. 2020, 21, 3249. [Google Scholar] [CrossRef]
- Senovilla, M.; Abreu, I.; Escudero, V.; Cano, C.; Bago, A.; Imperial, J.; González-Guerrero, M. MtCOPT2 is a Cu+ transporter specifically expressed in Medicago truncatula mycorrhizal roots. Mycorrhiza 2020, 30, 781–788. [Google Scholar] [CrossRef]
- Wang, S.; Chen, A.; Xie, K.; Yang, X.; Luo, Z.; Chen, J.; Zeng, D.; Ren, Y.; Yang, C.; Wang, L.; et al. Functional analysis of the OsNPF4.5 nitrate transporter reveals a conserved mycorrhizal pathway of nitrogen acquisition in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 16649–16659. [Google Scholar] [CrossRef]
- Drechsler, N.; Courty, P.-E.; Brulé, D.; Kunze, R. Identification of arbuscular mycorrhiza-inducible Nitrate Transporter 1/Peptide Transporter Family (NPF) genes in rice. Mycorrhiza 2018, 28, 93–100. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Cavagnaro, T.R.; Watts-Williams, S.J. The effects of soil phosphorus and zinc availability on plant responses to mycorrhizal fungi: A physiological and molecular assessment. Sci. Rep. 2019, 9, 14880. [Google Scholar] [CrossRef] [PubMed]
- Watts-Williams, S.J.; Cavagnaro, T.R. Arbuscular mycorrhizal fungi increase grain zinc concentration and modify the expression of root ZIP transporter genes in a modern barley (Hordeum vulgare) cultivar. Plant Sci. 2018, 274, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wei, S.; Hu, W.; Xiao, L.; Tang, M. Arbuscular mycorrhizal fungus rhizophagus irregularis increased potassium content and expression of genes encoding potassium channels in Lycium barbarum. Front. Plant Sci. 2017, 8, 440. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, J.; Liu, J.; Cui, M.; Huang, Y.; Tian, Y.; Chen, A.; Xu, G. The potassium transporter slhak10 is involved in mycorrhizal potassium uptake. Plant Physiol. 2019, 180, 465–479. [Google Scholar] [CrossRef]
- Drew, E.A.; Murray, R.S.; Smith, S.E.; Jakobsen, I. Beyond the rhizosphere: Growth and function of Arbuscular mycorrhizal external hyphae in sands of varying pore sizes. Plant Soil 2003, 251, 105–114. [Google Scholar] [CrossRef]
- Rooney, D.C.; Killham, K.; Bending, G.D.; Baggs, E.; Weih, M.; Hodge, A. Mycorrhizas and biomass crops: Opportunities for future sustainable development. Trends Plant Sci. 2009, 14, 542–549. [Google Scholar] [CrossRef]
- Augé, R.M. Arbuscular mycorrhizae and soil/plant water relations. Can. J. Soil Sci. 2004, 84, 373–381. [Google Scholar] [CrossRef]
- Driver, J.D.; Holben, W.E.; Rillig, M.C. Characterization of glomalin as a hyphal wall component of Arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2005, 37, 101–106. [Google Scholar] [CrossRef]
- Zhang, J.; Li, J.; Ma, L.; He, X.; Liu, Z.; Wang, F.; Chu, G.; Tang, X. Accumulation of glomalin-related soil protein benefits soil carbon sequestration: Tropical coastal forest restoration experiences. Land Degrad. Dev. 2022, 33, 1541–1551. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, X.; Sun, Y.; Wu, Z.; Li, T.; Hu, Y.; Lv, J.; Li, G.; Zhang, Z.; Zhang, J.; et al. Chromium immobilization by extra- and intraradical fungal structures of Arbuscular mycorrhizal symbioses. J. Hazard. Mater. 2016, 316, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.L.; Chen, B.D.; Sun, Y.Q.; Ren, B.H.; Zhang, X.; Wang, Y.S. Chromium resistance of dandelion (Taraxacum platypecidum Diels.) and bermudagrass (Cynodon dactylon [Linn.] Pers.) is enhanced by arbuscular mycorrhiza in Cr(VI)-contaminated soils. Environ. Toxicol. Chem. 2014, 33, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Arias, J.A.; Peralta-Videa, J.R.; Ellzey, J.T.; Viveros, M.N.; Ren, M.; Mokgalaka-Matlala, N.S.; Castillo-Michel, H.; Gardea-Torresdey, J.L. Plant growth and metal distribution in tissues of Prosopis Juliflora-velutina grown on chromium contaminated soil in the presence of Glomus deserticola. Environ. Sci. Technol. 2010, 44, 7272–7279. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Wu, S.C.; Kuek, C.; Khan, A.G.; Wong, M.H. The role of mycorrhizae associated with vetiver grown in Pb-/ Zn-contaminated soils: Greenhouse study. Restor. Ecol. 2007, 15, 60–67. [Google Scholar] [CrossRef]
- Chen, B.D.; Li, X.L.; Tao, H.Q.; Christie, P.; Wong, M.H. The role of arbuscular mycorrhiza in zinc uptake by red clover growing in a calcareous soil spiked with various quantities of zinc. Chemosphere 2003, 50, 839–846. [Google Scholar] [CrossRef]
- Arriagada, C.; Pereira, G.; García-Romera, I.; Ocampo, J.A. Improved zinc tolerance in Eucalyptus globulus inoculated with Glomus deserticola and Trametes versicolor or Coriolopsis rigida. Soil Biol. Biochem. 2010, 42, 118–124. [Google Scholar] [CrossRef]
- Arriagada, C.A.; Herrera, M.A.; Ocampo, J.A. Contribution of Arbuscular mycorrhizal and saprobe fungi to the tolerance of Eucalyptus globulus to Pb. Water. Air. Soil Pollut. 2005, 166, 31–47. [Google Scholar] [CrossRef]
- Maldaner, J.; Steffen, G.P.K.; Saldanha, C.W.; Steffen, R.B.; Tabaldi, L.A.; Missio, E.L.; De Morais, R.M.; Flores, R. Combining tolerant species and microorganisms for phytoremediation in aluminium-contaminated areas. Int. J. Environ. Stud. 2020, 77, 108–121. [Google Scholar] [CrossRef]
- Merlos, M.A.; Zitka, O.; Vojtech, A.; Azcón-Aguilar, C.; Ferrol, N. The Arbuscular mycorrhizal fungus Rhizophagus irregularis differentially regulates the copper response of two maize cultivars differing in copper tolerance. Plant Sci. 2016, 253, 68–76. [Google Scholar] [CrossRef]
- Ruscitti, M.; Arango, M.; Beltrano, J. Improvement of copper stress tolerance in pepper plants (Capsicum annuum L.) by inoculation with Arbuscular mycorrhizal fungi. Theor. Exp. Plant Physiol. 2017, 29, 37–49. [Google Scholar] [CrossRef]
- Li, J.; Liang, H.; Yan, M.; Chen, L.; Zhang, H.; Liu, J.; Wang, S.; Jin, Z. Arbuscular mycorrhiza fungi facilitate rapid adaptation of Elsholtzia splendens to copper. Sci. Total Environ. 2017, 599–600, 1462–1468. [Google Scholar] [CrossRef]
- Meier, S.; Cornejo, P.; Cartes, P.; Borie, F.; Medina, J.; Azcón, R. Interactive effect between Cu-adapted Arbuscular mycorrhizal fungi and biotreated agrowaste residue to improve the nutritional status of Oenothera picensis growing in Cu-polluted soils. J. Plant Nutr. Soil Sci. 2015, 178, 126–135. [Google Scholar] [CrossRef]
- Wu, J.T.; Wang, L.; Zhao, L.; Huang, X.C.; Ma, F. Arbuscular mycorrhizal fungi effect growth and photosynthesis of Phragmites australis (Cav.) Trin ex. Steudel under copper stress. Plant Biol. 2020, 22, 62–69. [Google Scholar] [CrossRef]
- Santana, N.A.; Ferreira, P.A.A.; Tarouco, C.P.; Schardong, I.S.; Antoniolli, Z.I.; Nicoloso, F.T.; Jacques, R.J.S. Earthworms and mycorrhization increase copper phytoextraction by Canavalia ensiformis in sandy soil. Ecotoxicol. Environ. Saf. 2019, 182, 109383. [Google Scholar] [CrossRef]
- Santana, N.A.; Rabuscke, C.M.; Soares, V.B.; Soriani, H.H.; Nicoloso, F.T.; Jacques, R.J.S. Vermicompost dose and mycorrhization determine the efficiency of copper phytoremediation by Canavalia ensiformis. Environ. Sci. Pollut. Res. 2018, 25, 12663–12677. [Google Scholar] [CrossRef]
- Cornejo, P.; Meier, S.; García, S.; Ferrol, N.; Durán, P.; Borie, F.; Seguel, A. Contribution of inoculation with Arbuscular mycorrhizal fungi to the bioremediation of a copper contaminated soil using Oenothera picensis. J. Soil Sci. Plant Nutr. 2017, 17, 14–21. [Google Scholar] [CrossRef]
- Fiqri, A.; Utomo, W.H.; Handayanto, E. Effect of Arbuscular mycorrhizal fungi on the potential of three wild plant species for phytoextraction of mercury from small-scale gold mine tailings. J. Degrad. Min. Lands Manag. 2016, 3, 551–558. [Google Scholar] [CrossRef]
- Chamba, I.; Rosado, D.; Kalinhoff, C.; Thangaswamy, S.; Sánchez-Rodríguez, A.; Gazquez, M.J. Erato polymnioides—A novel Hg hyperaccumulator plant in ecuadorian rainforest acid soils with potential of microbe-associated phytoremediation. Chemosphere 2017, 188, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Leudo, A.M.; Cruz, Y.; Montoya-Ruiz, C.; Delgado, M.D.P.; Saldarriaga, J.F. Mercury Phytoremediation with Lolium perenne-Mycorrhizae in Contaminated Soils. Sustainability 2020, 12, 3795. [Google Scholar] [CrossRef]
- Shabani, L.; Sabzalian, M.R.; Mostafavi pour, S. Arbuscular mycorrhiza affects nickel translocation and expression of ABC transporter and metallothionein genes in Festuca arundinacea. Mycorrhiza 2016, 26, 67–76. [Google Scholar] [CrossRef]
- Akib, M.A.; Mustari, K.; Kuswinanti, T.; Syaiful, S.A.; Syatrawati; Kumalawati, Z. Nickel (Ni) reduction in Sorowako post-mining soil through application of mycorrhiza Acaulospora sp. associated with Canavalia ensiformis L. J. Microb. Syst. Biotechnol. 2019, 1, 30–37. [Google Scholar] [CrossRef]
- Alam, M.Z.; Anamul Hoque, M.; Ahammed, G.J.; Carpenter-Boggs, L. Arbuscular mycorrhizal fungi reduce arsenic uptake and improve plant growth in Lens culinaris. PLoS ONE 2019, 14, e0211441. [Google Scholar] [CrossRef] [PubMed]
- Orłowska, E.; Godzik, B.; Turnau, K. Effect of different Arbuscular mycorrhizal fungal isolates on growth and arsenic accumulation in Plantago lanceolata L. Environ. Pollut. 2012, 168, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Shahabivand, S.; Maivan, H.Z.; Mahmoudi, E.; Soltani, B.M.; Sharifi, M.; Aliloo, A.A. Antioxidant activity and gene expression associated with cadmium toxicity in wheat affected by mycorrhizal fungus. Zemdirbyste 2016, 103, 53–60. [Google Scholar] [CrossRef]
- Liu, L.; Li, J.; Yue, F.; Yan, X.; Wang, F.; Bloszies, S.; Wang, Y. Effects of Arbuscular mycorrhizal inoculation and biochar amendment on maize growth, cadmium uptake and soil cadmium speciation in Cd-contaminated soil. Chemosphere 2018, 194, 495–503. [Google Scholar] [CrossRef]
- Abdelhameed, R.E.; Metwally, R.A. Alleviation of cadmium stress by Arbuscular mycorrhizal symbiosis. Int. J. Phytoremediat. 2019, 21, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Ho, S.H.; Zhu, S.; Ma, F.; Wu, J.; Yang, J.; Wang, L. Adaptive response of Arbuscular mycorrhizal symbiosis to accumulation of elements and translocation in Phragmites australis affected by cadmium stress. J. Environ. Manag. 2017, 197, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Janoušková, M.; Pavlíková, D.; Vosátka, M. Potential contribution of Arbuscular mycorrhiza to cadmium immobilisation in soil. Chemosphere 2006, 65, 1959–1965. [Google Scholar] [CrossRef]
- Garg, N.; Kaur, H. Response of Antioxidant Enzymes, Phytochelatins and Glutathione Production Towards Cd and Zn Stresses in Cajanus cajan (L.) Millsp. Genotypes Colonized by Arbuscular Mycorrhizal Fungi. J. Agron. Crop Sci. 2013, 199, 118–133. [Google Scholar] [CrossRef]
- Babadi, M.; Zalaghi, R.; Taghavi, M. A non-toxic polymer enhances sorghum-mycorrhiza symbiosis for bioremediation of Cd. Mycorrhiza 2019, 29, 375–387. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef]
- Padmavathiamma, P.K.; Li, L.Y. Phytoremediation technology: Hyper-accumulation metals in plants. Water. Air. Soil Pollut. 2007, 184, 105–126. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, Y.; Han, X.; Chiu, T.Y.; Ghosh, A.; Chen, H.; Tang, M. The roles of arbuscular mycorrhizal fungi (AMF) in phytoremediation and tree-herb interactions in Pb contaminated soil. Sci. Rep. 2016, 6, 20469. [Google Scholar] [CrossRef] [PubMed]
- Joner, E.J.; Leyval, C. Time-course of heavy metal uptake in maize and clover as affected by root density and different mycorrhizal inoculation regimes. Biol. Fertil. Soils 2001, 33, 351–357. [Google Scholar] [CrossRef]
- Ortega-Larrocea, M.P.; Siebe, C.; Estrada, A.; Webster, R. Mycorrhizal inoculum potential of Arbuscular mycorrhizal fungi in soils irrigated with wastewater for various lengths of time, as affected by heavy metals and available P. Appl. Soil Ecol. 2007, 37, 129–138. [Google Scholar] [CrossRef]
- Spagnoletti, F.; Carmona, M.; Gómez, N.E.T.; Chiocchio, V.; Lavado, R.S. Arbuscular mycorrhiza reduces the negative effects of M. phaseolina on soybean plants in arsenic-contaminated soils. Appl. Soil Ecol. 2017, 121, 41–47. [Google Scholar] [CrossRef]
- Mishra, A.; Bhattacharya, A.; Mishra, N. Mycorrhizal symbiosis: An effective tool for metal bioremediation. In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbes in Soil, Crop and Environmental Sustainability; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 9780128182581. [Google Scholar]
Figure 1.
Schematic diagram for mycorrhizal colonization stages of (A) Endomycorrhizae; (a) pre- symbiotic communication between fungal and root exudates; (b) hyphopodium development for fungal entry; (c) hyphal penetration into the cortex; (d) elongation of intraradical hyphae; (e) hyphae branching & arbuscule formation; and (B) Ectomycorrhizae; (f) mycelium or mantle sheath covering epidermis and cortical cells; (g) intraradicle elongation into cortical layer without penetrating cells (Hartig net).
Figure 1.
Schematic diagram for mycorrhizal colonization stages of (A) Endomycorrhizae; (a) pre- symbiotic communication between fungal and root exudates; (b) hyphopodium development for fungal entry; (c) hyphal penetration into the cortex; (d) elongation of intraradical hyphae; (e) hyphae branching & arbuscule formation; and (B) Ectomycorrhizae; (f) mycelium or mantle sheath covering epidermis and cortical cells; (g) intraradicle elongation into cortical layer without penetrating cells (Hartig net).
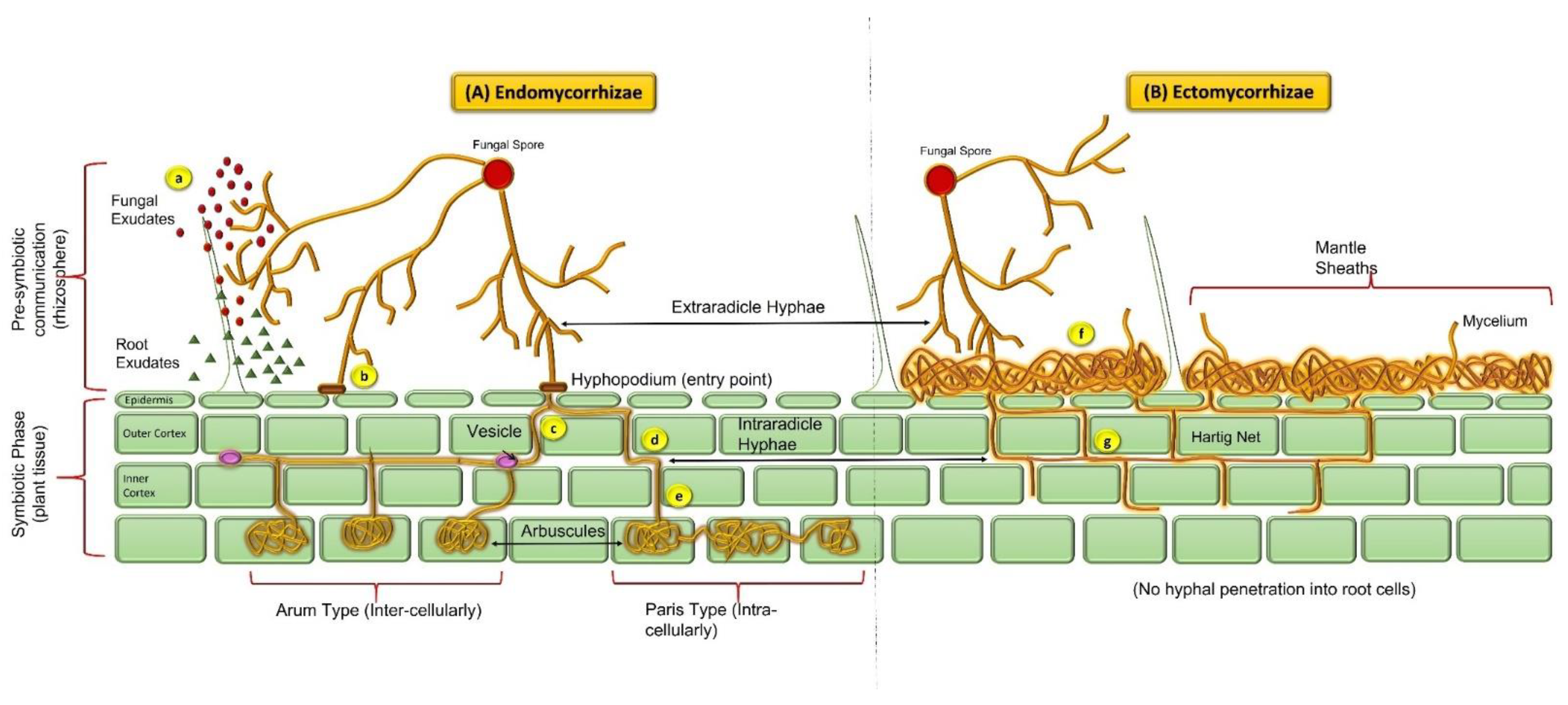
Figure 2.
Impact of various hormones on symbiotic phase regulation and suggested mode of action.
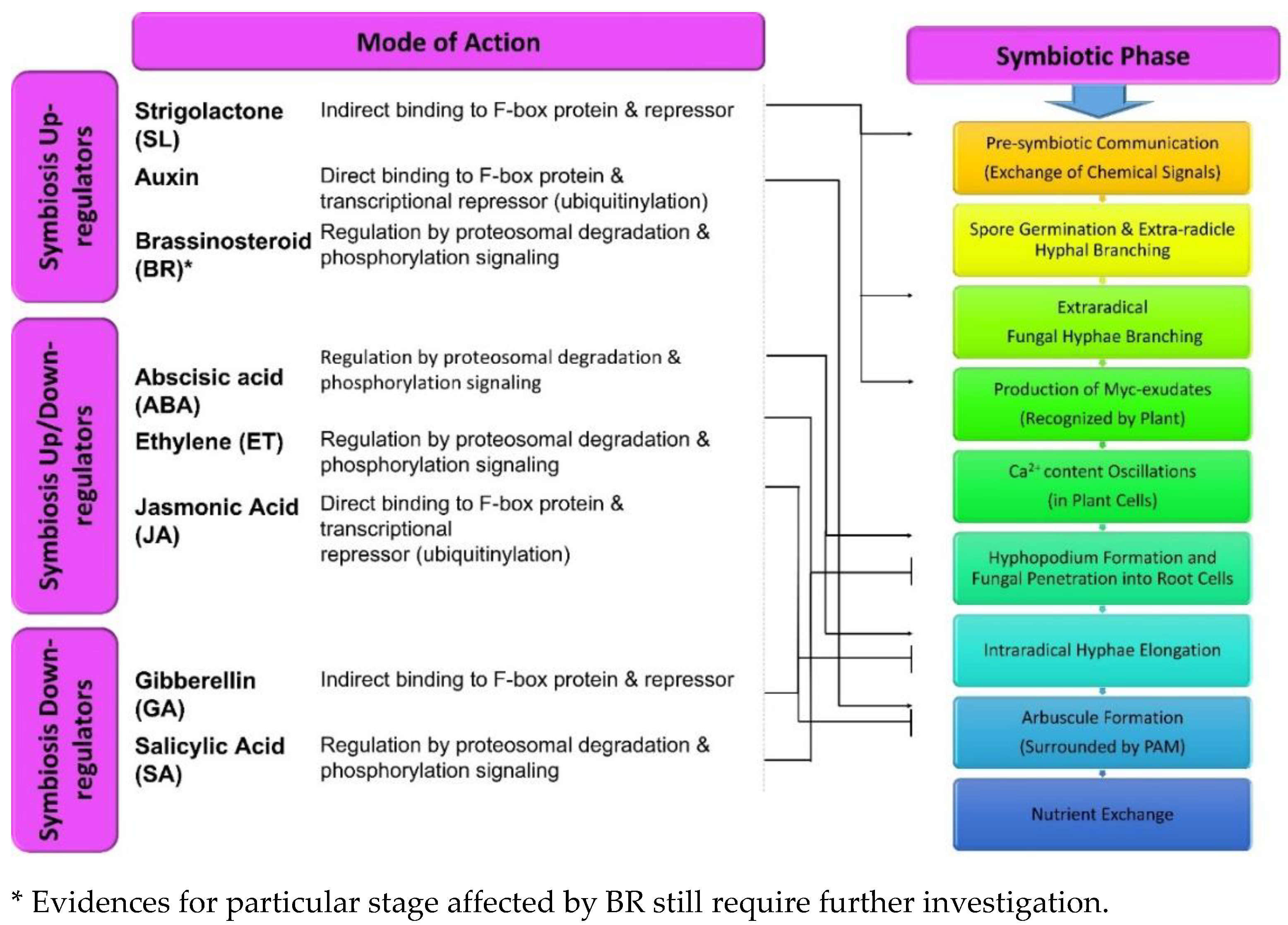
Figure 3.
The effect of mycorrhizal associations on plant growth and restoration of soil: alleviated nutrient supply, poor root network and impaired plant growth without mycorrhizal exposure (left), and rhizospheric extraradical hyphae extension deep into inaccessible soils (soil aggregation), elevation in nutrient uptake (ionic exchange by arbuscule formation through IFA) and improve plant growth (right).
Figure 3.
The effect of mycorrhizal associations on plant growth and restoration of soil: alleviated nutrient supply, poor root network and impaired plant growth without mycorrhizal exposure (left), and rhizospheric extraradical hyphae extension deep into inaccessible soils (soil aggregation), elevation in nutrient uptake (ionic exchange by arbuscule formation through IFA) and improve plant growth (right).
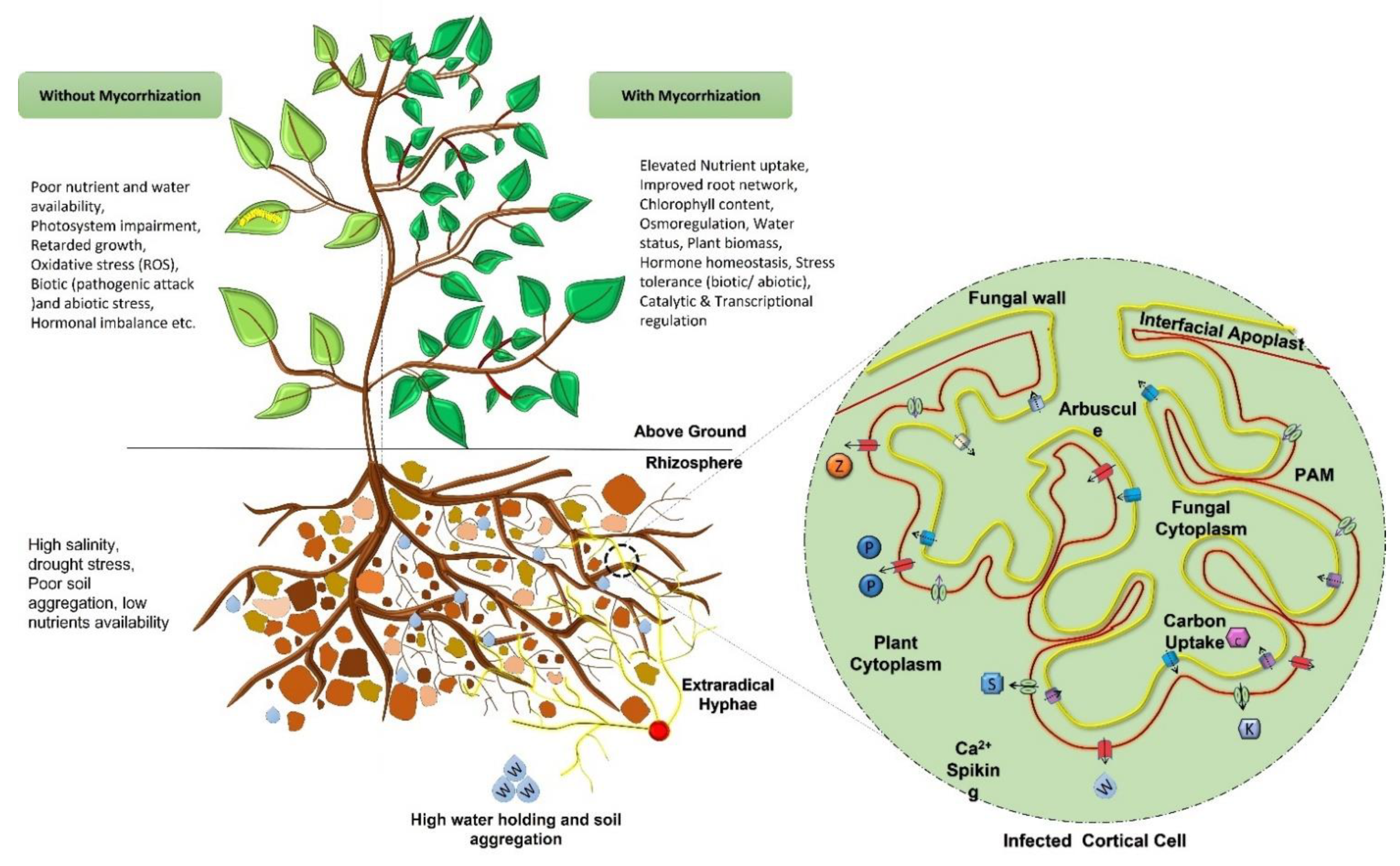

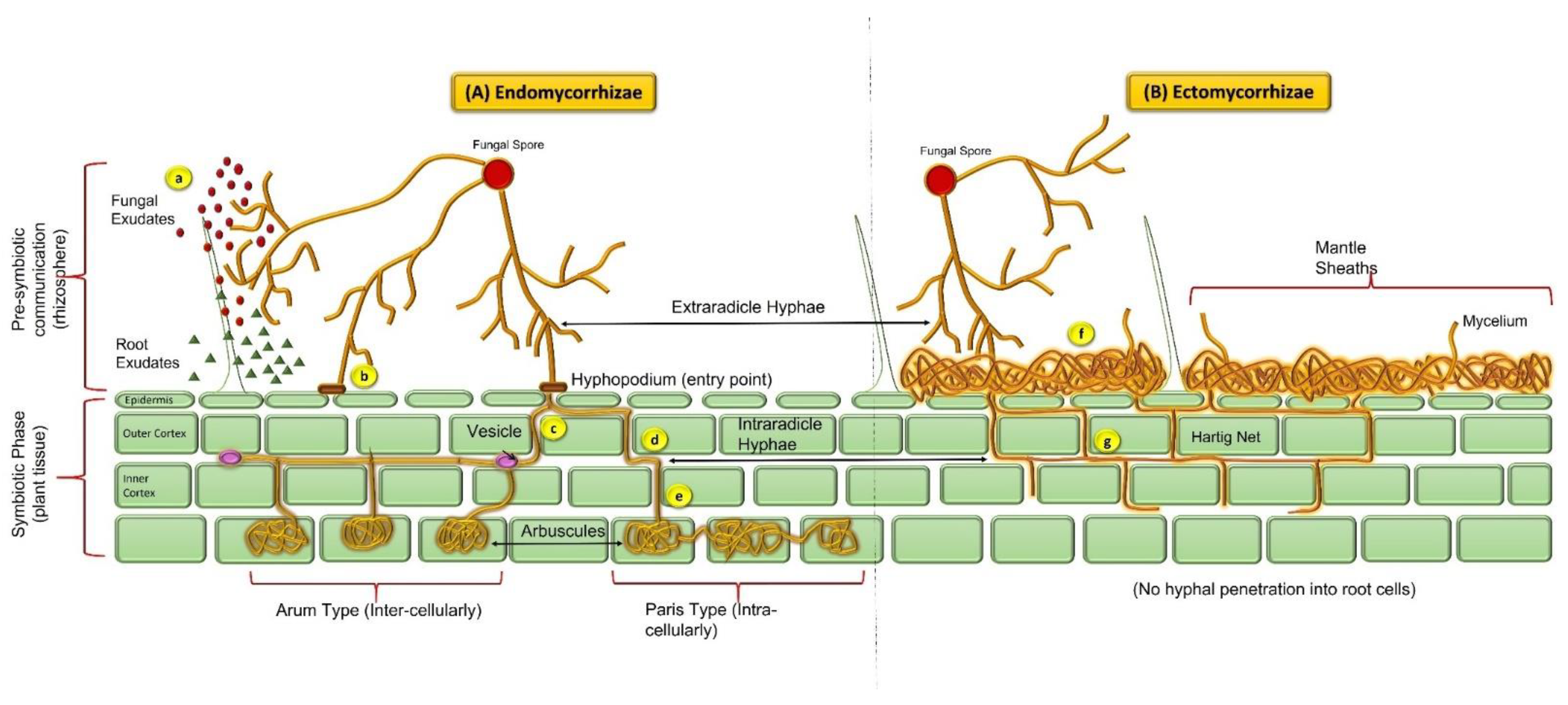{kind=link}
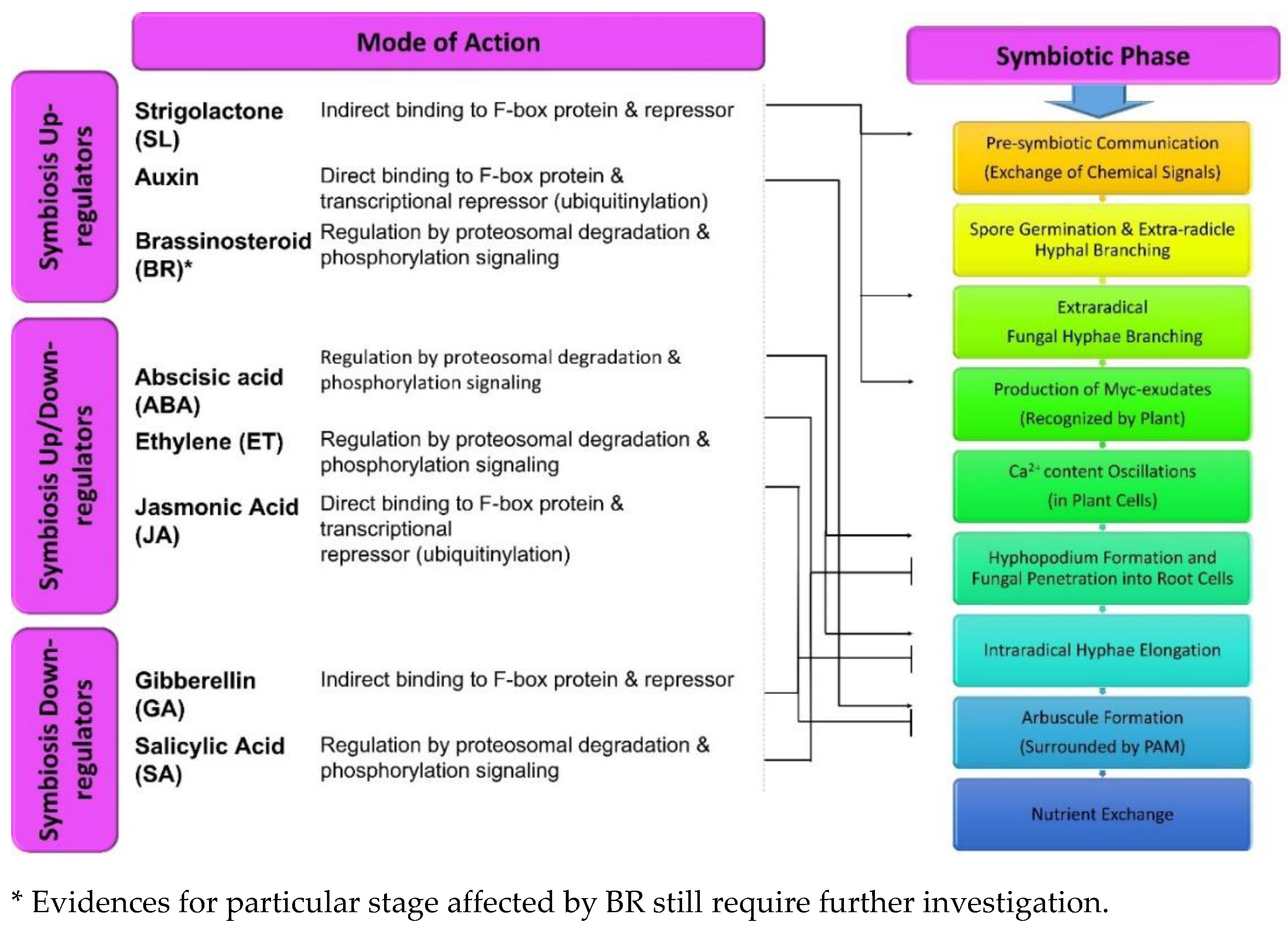{kind=link}
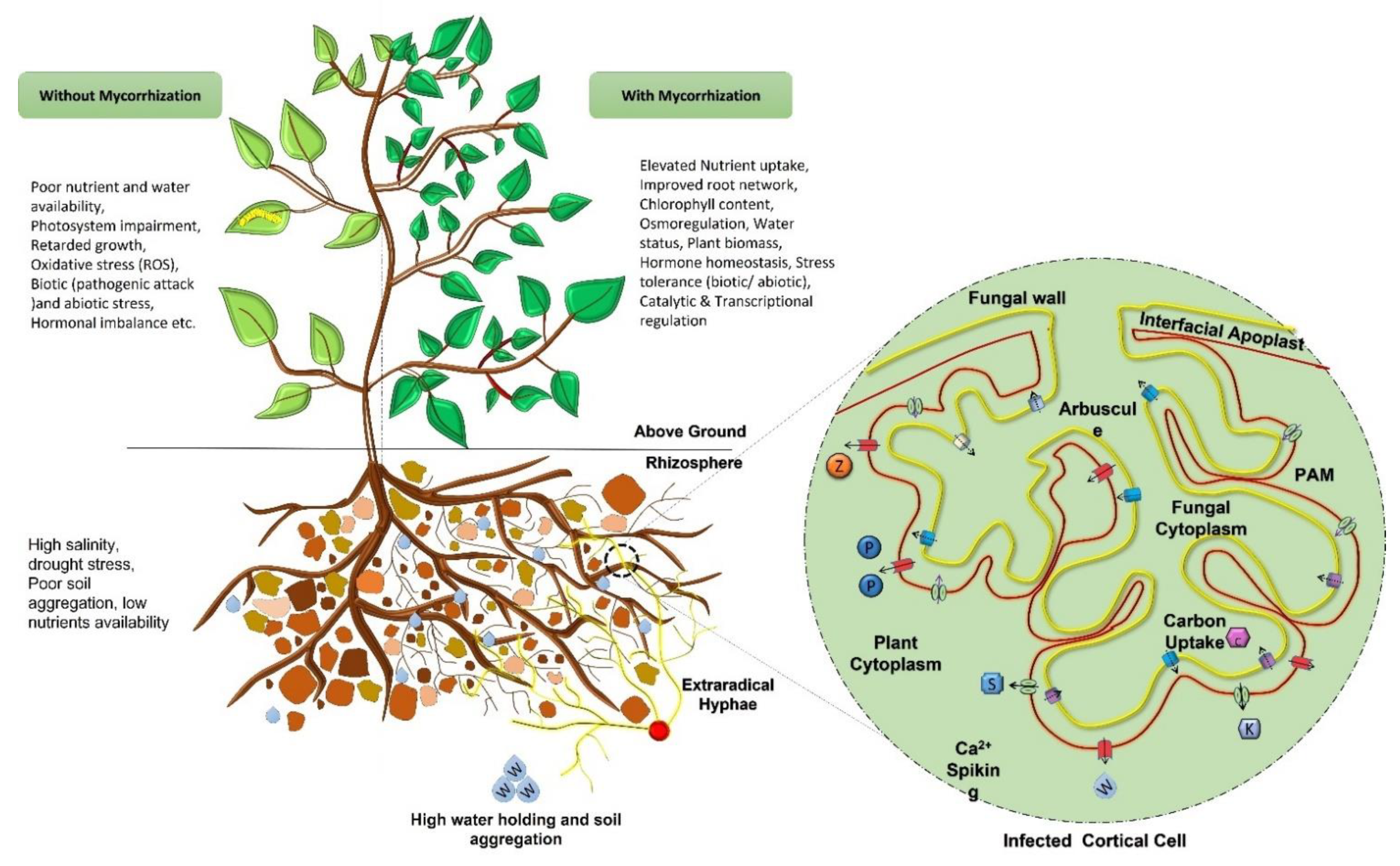{kind=link}
Table 1.
Effect of mycorrhizal symbiosis on stress tolerance.
| Sr. No. | Stress Condition | Mycorrhizal sp. | Host Plant | Possible Mechanism | Reference |
|---|---|---|---|---|---|
| 1 | Salinity, 200 mM NaCl | Rhizophagus irregularis (Formerly Glomus intraradices) | R. pseudoacacia | Improved photosynthetic rate, PS-II photochemistry, water status, K+, Chloroplast (RppsbA, RppsbD, RprbcL)& transporter genes (RpSOS1, RpHKT1, RpSKOR) up-regulation, lower shoot:root Na+ content | [21] |
| 2 | 80 mM NaCl | Glomus intraradices | Lactuca sativa | Alleviates salt stress through improved stomata performance, photosystem (PS-II), Carotenoid deoxygenase gene (LsNCED2) induction, normalized ABA level and by altering the hormonal profiles (SLs induction) | [22] |
| 3 | 100 mM NaCl | Rhizophagus irregularis | Solanum lycopersicum | Elevated K+ and K+/Na+ ratio (prevention of metabolic processes disruption), regulated hormone synthesis & cross talk | [23] |
| 4 | 200 mM NaCl | Glomuse tunicatum, Glomus intraradices, Glomus mosseae | Cucumis sativus L. | Photosynthetic pigments regulation, enhanced antioxidant activities, osmolyte (proline& phenols) regulation, improved water status; regulated mineral uptake; reduced uptake of Na+. | [24] |
| 5 | 200 mM NaCl | Claroideoglomus etunicatum | Aeluropus littoralis | Overcome free radical formation by elevated antioxidant activity, high CO2 synthesis and nitrate assimilation | [25] |
| 6 | 120 mM NaCl | Funnelliformis mosseae, Acaulospora laevis, Gigaspora margarita | Oryza sativa L. | Rise in chlorophyll content, K+/Na+ ratio, photosynthesis, and dropped shoot/root Na+ ratio by limiting Na+ uptake and translocation. | [26] |
| 7 | 200 mM NaCl | Funneliformis mosseae | Malus domestica Borkh. | AMF in combination with dopamine help to maintain host cell membrane integrity, improves photosynthesis | [27] |
| 8 | 35 and 70 mM NaCl. | Glomus sp. mix (G. mosseae, G. intraradices, G. hoi) | Citrus aurantium L. | Elevation in plant growth, chlorophyll levels, improved water status, gas exchange capacities (increased photosynthetic rate, stomatal conductance and transpiration rate), enhanced oxidative stress defense system | [28] |
| 9 | 160 mM NaCl | R. intraradices and F. mosseae. | Prunusdulcis× Prunuspersica hybrid | Improved physiological parameters (chlorophyll, osmolytes that are soluble sugars and proline content to combat salt toxicity) and increased antioxidant enzymes activity compared to non-inoculated. F. mosseae elevated chlorophyll content more efficiently, whereas R. intraradices prevailed total sugars and proline content. | [29] |
| 10 | 150 mM NaCl | Glomus etunicatum, Glomusgeo sporum, and Glomus mosseae | Oryza sativa L. | Improved physiological parameters (chlorophyll, osmolytes that are soluble sugars and proline content to combat salt toxicity) and increased antioxidant enzymes activity | [30] |
| 11 | - | Funneliformis mosseae and Claroideoglomus etunicatum | Puccinellia tenuiflora | Increased P uptake, high antioxidant capacities, enhanced biomass to dilute the salt concentration, elevated K+/Na+ ratio, restricted Na+ translocation towards aerial parts. | [31] |
| 12 | 200 mM NaCl | Glomus monosporum, G. clarum, Gigasporanigra, and Acaulospora laevis | Vigna unguiculata L. | Elevated photosynthetic pigments, soluble sugar contents, ions accumulation and compartmentalization (maintained membrane integrity) and high enzymatic activities. | [5] |
| 13 | Drought Stress | Funneliformis mosseae (formerly Glomus mosseae) and Rhizophagus intraradices | Solanum lycopersicum | Stress tolerance varies depending upon the myc-species. F. mossaeae promoted volatile emission (VOC), high arbuscule formation in colonization regions R. intraradices is more efficient towards P uptake (upregulated P transporters; LePT4,5), high plant performance to lower water dispersal by adopting a compact structure (high internode/height ratio), high water utilization efficiency | [32] |
| 14 | Drought Stress | AMF | Glycine max L. | Increased water holding capacity, photosynthesics, osmoregulation | [33] |
| 15 | Drought Stress | Rhizophagus irregularis (formerly Glomus intraradices) and Funneliformis mosseae (formerly G. mosseae) | Trifolium alexandrinum L. | Enhanced nutrient uptake, increase in phosphorus acquisition, defense against oxidative stress, increased N2 fixation, sufficient availability of the photosynthates | [34] |
| 16 | Drought Stress | Funneliformis mosseae and Rhizophagus intraradices | Solanum lycopersicum | Aquaporin genes regulation; LeNIP3;1 (overexpressed), LeNIP3;1 & LeTIP2;3 (suppressed) by F. mosseae, and RiAQPF1 & 2 (overexpressed) by R. intraradices, elevated stomatal density, activation of LOX (lipoxygenase) genes, increased antioxidant activity, proline content (osmoregulation) | [35] |
| 17 | Drought Stress | Funneliformis mosseae | Triticum durum Desf., Triticum aestivum L. | Positive impact on root metabolome, high C fixation, high P sugar accumulation, osmoregulatory effects, anti-oxidative behavior, regulated phytohormone profile | [3] |
| 18 | Drought Stress | Rhizophagus irregularis | Zea mays | Efficiency of photosystem II, membrane stability, osmotic regulation via accumulation of soluble sugars and plant biomass production. Root hydraulic conduction via down-regulating aquaporin genes (ZmPIP1;6, ZmPIP2;2, and ZmTIP4;1) | [36] |
| 19 | Drought Stress | Myc-mix. (Rhizophagus intraradices + Funneliformis mosseae + F. geosporum) | Triticum aestivum | Elevation in photosynthetic pigments, high Mg uptake, C fixation (photosynthate) and biomass; improved water status; enhanced PSI & PSII photochemistry | [37] |
| 20 | - | Rhizophagus irregularis | Solanum lycopersicum | Promoted photosynthesis, improved C fixation, osmoregulation and root hydraulic conductivity via enhanced aquaporin. | [38] |
| 21 | Temperature stress (43–44 °C) | Funneliformis sp. AMF | Zea mays | Up-regulated water transport and transpiration, regulated PSII heterogeneity, stomatal conductance | [39] |
| 22 | (44 °C) | Rhizophagus intraradices, Funneliformis mosseae, F. geosporum | Zea mays | Enhanced PSI & PSII photochemistry, high Mg2+ uptake. | [40] |
| 23 | (35 °C) | Rhizophagus irregularis, Funneliformis mosseae, Funneliformis geosporum, Claroideoglomus claroideum | Triticum aestivum L. | Increased photosynthetic yield, nutrient distribution and nutrient composition in roots, lowered the K/Ca ratio | [41] |
| 24 | (3–5 °C) | Glomus versiforme and Rhizophagus irregularis | Hordeum vulgare L. | Enhanced membrane stability, antioxidative capacity & phenolics metabolism Glomus sp. imparted more alleviation against cold stress. Rhizophagus found more efficient towards survival rate. | [42] |
| 25 | (15 °C) | Rhizophagus irregularis | Zea mays L. | Down-regulated PS-I & PS-II genes and decreased oxidative stress, enhanced C assimilation by metabolic upregulation, high ATP production by increased P concentration | [43] |
| 26 | (5–25 °C) | Funneliformis mosseae, Claroideoglomus etunicatum, Rhizophagus irregularis, and Diversispora versiformis) | Solanum melongena L. | Promoted photochemical, antioxidant activities, and maintained membrane integrity, proline and phenolics accumulation (protection against stress) | [44] |
| 27 | (4 ± 0.5 °C) | Glomus intraradices | Citrullus lanatus | Improved photosynthesis, induced peroxidase (POX) activity, restoring photosynthesis efficiency, released oxidative stress | [45] |
| 28 | Biotic stress Aphids (M. euphorbiae) | Rhizophagus intraradices | Solanum lycopersicum L. | Indirect defense via enzymatic release of methyl salicylate to attract parasitoid A.ervi | [32] |
| 29 | Spodoptera littoralis | Rhizophagus irregularis | Solanum lycopersicum L. | Enhanced nutrient acquisition, N2 fixation, defense activation | [46] |
| 30 | Caterpillar, Helicoverpaarimigera | Glomus mosseae | Solanum lycopersicum Mill. | Activation of stress responsive genes (LOXD, AOC, PI-I & II) in leaves, regulated JA cascade | [47] |
| 31 | Meloidogyne incognita (severe yield losses in tomato) | Rhizophagus intraradices | Solanum lycopersicum | Improved plant peroxidases for ROS scavenging, Upregulated flavonoid enzymes, modulation of pathogen related genes (LTP), phytohormonal regulation, increased glutathione transferases | [48] |
| 32 | Fusarium virguliforme | Rhizophagus irregularis | Glycine max | Peroxidase genes regulation, decreased. Down-regulation of several genes coding for glutathione-S-transferase (GST) | [49] |
| 33 | Xiphynema index | Rhizophagus intraradices | Grapevine rootstock SO4 (Vitis berlandieri × V.riparia) | Decreased down-regulation of several genes coding for glutathione-S-transferase (GST) | [50] |
Table 2.
Regulation of mineral nutrition via transporters in mycorrhizae associated plants.
| Sr. No. | Mineral | Mycorrhizal sp. | Plant sp. | Host Plant Transporters | Effect of Mycorrhizal Symbiosis | Reference |
|---|---|---|---|---|---|---|
| 1. | Phosphate | Claroideoglomus etunicatum | Camellia sinensis | CsPT1 & CsPT4 | AMF up-regulated root CsPT1 expression, while down-regulated the CsPT4 expression. AMF inoculation significantly promoted P acquisition capacity of tea plants, especially in roots through improving root growth and enhancing soil acid phosphatase activity and root CsPT1 expression. | [124] |
| Rhizophagus irregularis | Zea mays | ZmPht1;6 & ZmPht1;11 | AMF improved plant growth and Pi assimilation, AMF colonization strongly improved the nutritional status of the plants and increased the internal P concentration. ZmPht1;6 over expression at a high level in AMF-colonized roots. While less expressed ZmPht1;11 also stimulated by AMF colonization. | [125] | ||
| 2. | Gigaspora margarita or Funnelliformis mosseae | Lotus japonicus | LjPT4 | LjPT4 affects proper arbuscule formation on the fungal side and for improved Pi uptake on the plant side. | [126] | |
| 3. | Sulfur | Rhizophagus irregularis | Zea mays | ZmSULTR1.2a, ZmSULTR2.1, ZmSULTR3.5 | Upregulation of ZmSULTR1.2a & ZmSULTR2.1 in sulfur deprived conditions while downregulation of ZmSULTR3.5 in mycorrhized plants. | [127] |
| 4. | Copper | Rhizophagus irregularis | Medicago truncatula | MtCOPT2 | Preferential expression of MtCOPT2 during mycorrhizal symbiosis. | [128] |
| Nitrate | Rhizophagus irregularis | Oryza sativa, Zea mays, Sorghum bicolor, Medicago truncatula | OsNPF4.5, ZmNPF4.5, SbNPF4.5, MtNPF4.5 | Myc-symbiosis resulted in efficient up-regulation of OsNPF4.5, ZmNPF4.5 and SbNPF4.5, while slight induction of MtNPF4.5. | [129] | |
| Rhizophagus irregularis | Oryza sativa | OsNPF genes: NPF2.2/ PTR2, NPF1.3, NPF6.4 and NPF4.12 | Enhanced expression of nitrate transporter genes in mycorrhizal roots in nutrient dependent manner. | [130] | ||
| 5. | Ammonium | Rhizophagus irregularis | Oryza sativa | OsAM1, OsAM10, OsAM20, OsAM25 | Significant upregulation in roots via AMF symbiosis. | [130] |
| Rhizophagus irregularis | Oryza sativa | OsAMT3.1 | Up-regulation of OsAMT3.1 in rice mycorrhizal roots | [129] | ||
| 6. | Zinc | Rhizophagus irregularis | Medicago truncatula | MtZIP5, MtZIP2 | AMF symbiosis caused higher expression of MtZIP5 in poor rhizospheric Zn condition and reduction in MtZIP2 at elevated soil Zn concentration. | [131] |
| Rhizophagus irregularis/mock-inoculated | Hordeum vulgare | HvZIP3, HvZIP7, HvZIP8, HvZIP10, HvZI13 | Out of five transporters, HvZI13 found most significantly upregulated, HvZI3 & 8 upregulated also in Zn deficient conditions, while HvZI7 & 10 downregulated. | [132] | ||
| 7. | Potassium | Rhizophagus irregularis | Lycium barbarum Solanum lycopersicum | LbKT1, LbSKOR SlHAK10 | Regulated expression of LbKT1 and LbSKOR for varying water & potassium availability | [133,134] |
Table 3.
Influence of different mycorrhizal sp. on soil restoration by phytoremediation of toxic metals.
Table 3.
Influence of different mycorrhizal sp. on soil restoration by phytoremediation of toxic metals.
| Pollutant | Mycorrhizal Species | Plant Species | Possible Mechanism | Literature Cited |
|---|---|---|---|---|
| Chromium (Cr) | Rhizophagus irregularis | Daucuscarota | Reduced translocation, and immobilization of Cr6+ through EPS (extracellular polymers) production. distribution of Cr in roots | [140] |
| Rhizophagus irregularis | bermudagrass [Cynodondactylon (Linn.) | Cr absorption and immobilization by AM roots, Reduction of Cr6+ to Cr3+ within fungal structures, inhibited Cr flow from roots to shoots, | [141] | |
| Rhizophagus irregularis | Taraxacum platypecidum | Cr absorption and immobilization by AM roots, inhibit Cr translocation from roots to shoots, promoted plant growth | [141] | |
| Glomus deserticola | Prosopisjuli flora-velutina | Accumulation of Cr in vascular tissue and decreased the translocation of Cr into shoots | [142] | |
| Zinc (Zn) | Glomus mosseae & G. intraradices | Vetiver grass | Increased P uptake by the plant and improved overall growth (G. intraradices showed more rehabilitation capacity) | [143] |
| Glomu smosseae | Trifolium pratense | Zn accumulation in roots which decreases in shoots as the Zn conc. rises to its maximum, improved P sustenance | [144] | |
| Glomus deserticola | Eucalyptus globulus | Increased root to shoot metal accumulation, high metabolic activity, symbiotic effect of saprophytic fungal sp. on mycoremediation process | [145] | |
| Lead (Pb) | Glomus mosseae& G. intraradices | Vetiver grass | Increased P uptake by the plant and improved overall growth (G. mosseae showed more rehabilitation capacity) | [143] |
| Glomus mosseae and G. deserticola | Eucalyptus globulus | Promoted overall growth, mineral nutrition, chlorophyll production and enzymatic performances (which further increased due to synergistic effect of G. deserticola and T. koningii), enhanced Pb accumulation | [146] | |
| Aluminium | Pisolithus sp. | Schinusmolle | Phytoextraction or phytostbilization, Glomalin production supported chelation, rise in photochemical efficacy | [147] |
| Copper (Cu) | R. irregularis | Zea mays | Increased accumulation of total phytochelating content in shoots | [148] |
| Funneliformis mosseae; R. intraradices | Capsicum annuum | Cu Higher total dry weight and the leaf | [149] | |
| Arbascular Mycorrhizal Fungi (AMF) | Elsholtzia splendens | Increase in germination rate and the germination index of the seeds as well as the fresh weights of hypocotyl and radicle | [150] | |
| Claroideoglomus claroideum | Oenothera picensis | Protect plant from metal toxicity, enhance both plant establishment and nutrition | [151] | |
| R. irregularis | Phragmites australis | Stress tolerance via up-regulating photo systems membrane complexes, improved plant growth. | [152] | |
| Rhizoglomus clarum | Canavalia ensiformis | Alleviated amounts of Cu due to phytoextraction in addition to earthworms | [153] | |
| Rhizophagus clarus | Canavalia ensiformis | Alleviated amounts of Cu due to phytoextraction & phytostabilization in addition to bovine | [154] | |
| Claroideo glomu sclaroideum and | Oenothera picensis | Cu chelation with AM-secreted glomalin protein | [155] | |
| Mercury (Hg) | Glomussp.,Gigaspora sp. &Skutelespora sp. | Cyperus kyllingia, Lindernia crustacea, Paspalum conjugatum | P. conjugatum resulted maximum phytoextraction, while C.kyllingia exhibited maximum (Hg) tolerance | [156] |
| Native AM fungal morphotypes | Axonopus compressus, and Erato polymnioides | A. compressus ensued phythoextracting; Eratopolymnioides–Hg phytostabilization | [157] | |
| AMF | Lolium perenne | Decreased shoot:root (St:Rt) (Hg conc.), increased metal assimilation in roots | [158] | |
| Nickel (Ni) | Funneliformis mosseae (also named as Glomus mosseae) | Festuca arundinacea | Enhance expression of ABC transporters and metallothione induced metal intoxication, decreased metal translocation | [159] |
| Acaulospora sp. (indigenous) | Canavalia ensiformis | [160] | ||
| Arsenic (As) | AMF mix | Lens culinaris | Alleviated uptake by roots and shoots as an effect of mycorrhizal association | [161] |
| Rhizophagus intraradices (formerly named G. intraradices) | Plantago lanceolata | Down-regulating phosphate/arsenate transporters could assist plants to enhance the As tolerance | [162] | |
| Rhizoglomus intraradices & Glomus etunicatum | Triticum aestivum | Regulated P/As ratio, enhanced antioxidant production, holding As into non-toxic forms via increased production of biopolymers | [108] | |
| Rhizoglomus intraradices | Robiniapseudoacacia | Induced changes in root morphology, increased shoot-root dry weights, controlled phyto-hormone concentration etc. | [108] | |
| Acaulospora scrobiculata | Anadenantheraperegrina | Promoted P uptake lead to higher growth rates, As concentrations in the roots and shoots. | [109] | |
| Cadmium (Cd) | Funelliformis mosseae and Piriformos poraindica | T. aestivum | Biomass uplift, imposed catalytic activities for G-SH transferase, catalase, peroxidase etc., and antioxidant genes upregulation | [163] |
| Glomus intraradices | Zea mays | Mycorrhizae in association with biochar resulted alleviation in Cd accumulation in plant and restricted mobilization, soil rehabiliation | [164] | |
| Glomus monosporum, G. clarum, Gigaspora nigra, and Acaulospora laevis | Trigonella foenum-graecum | Decreased St: Rt Cd ratio, enhanced antioxidant activities | [165] | |
| Rhizophagus irregularis | Phragmites australis | Immobilization of Cd in roots, increased mineral uptake (Mn& P mainly) to survive Cd-toxicity | [166] | |
| Glomus intraradices, Glomus mosseae, Glomus claroideum, and Glomus geosporum | Nicotiana tabacum | Phyto stabilization of lead via immobilization in extraradical mycelial network | [167] | |
| Glomusmosseae | Cajanus ajan | Diminished oxidative disturbances (free radicle formation), high non-protein thiols (-SH) production and high antioxidant activities | [168] | |
| Claroideoglomus etunicatum | Sorghum bicolor | Increased the glomalin content for improved soil, Cd stabilization in mycorrhizal roots &phytoextraction (by shoots), high nutrient uptake | [169] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dhiman, M.; Sharma, L.; Kaushik, P.; Singh, A.; Sharma, M.M. Mycorrhiza: An Ecofriendly Bio-Tool for Better Survival of Plants in Nature. Sustainability 2022, 14, 10220. https://doi.org/10.3390/su141610220
AMA Style
Dhiman M, Sharma L, Kaushik P, Singh A, Sharma MM. Mycorrhiza: An Ecofriendly Bio-Tool for Better Survival of Plants in Nature. Sustainability. 2022; 14(16):10220. https://doi.org/10.3390/su141610220
Chicago/Turabian StyleDhiman, Mamta, Lakshika Sharma, Prashant Kaushik, Abhijeet Singh, and Madan Mohan Sharma. 2022. "Mycorrhiza: An Ecofriendly Bio-Tool for Better Survival of Plants in Nature" Sustainability 14, no. 16: 10220. https://doi.org/10.3390/su141610220
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.