1. Introduction
Terrestrial oligochaetes are earthworms, usually living on earth. In many temperate ecosystems, invertebrates are the most prominent animal biomass and significantly influence soil biological, chemical, and chemical characteristics. They influence the structure and composition of the above-ground plant population by altering soil structure and speeding up organic matter decomposition and nutrient cycling. Many people assume that there is only one kind of earthworm since they live in burrows and have a simple body structure. In fact, earthworms are a large and diverse group of burrowing annelids with over 6000 species mainly described in name and morphology, whereas their biology and ecology are unknown. Earthworms have a variety of life strategies and inhabit a variety of ecological niches. The ecosystem categorizes earthworms into three distinct groups: entogeic, epic, and endogeic, according to their feeding habits. Some organisms cannot be correctly assigned to any of these groups because they are difficult to define. Earthworms burrow deeper in agricultural soils than grassland and forest soils [
1,
2].
The earthworm is considered the critical environmental mediator capable of affecting earth functions and microbial activities by generating priming effects. Energy-rich mucus stimulates microorganisms and signals hormone-like molecules that affect plant genes’ expression. The “Sleeping Beauty Paradox” has been called a reciprocal relationship between earthworms and earth microbiota. The earthworm kiss, made in the drilosphere of readily assimilable glycoproteins, is triggered in sleeping microorganisms awaiting appropriate environmental conditions. Such a process helps microbial processes escalate over a short time (‘the hot moment’) and reverberate to a greater degree in limited soil region (‘the hot point’) at the bio-pores’ microscale aggregates at the level of soil and drilosphere. The earthworm’s impact on soil microbiota is significant and nutritionally substantial [
1,
2,
3,
4]. The effect of this, however, may rely on your food choice, range, dietary intake, assimilation, and digestion, helping earthworms digest microorganisms by reducing microbial biomass, particularly fungal biomass [
5,
6,
7]. It could also be used to pick or stimulate soil microbes that help them in soil digestion, as the earthworm bottom frequently lacks adequate enzymes. This process enriches soils with bacterial taxa-like bacteria that can break down organic matter in which earthworm nourishes or denitrifies bacteria that live under reduced intestinal oxygen conditions. Since microorganisms’ participation in these functions is essential, earthworm impact oil microbe communities is crucial in predicting and understanding the earthworm impact on ecosystem functions [
8,
9]. Thus, the earthworm impact on plant growth and nutrient cycling is not only direct but primarily and indirectly mediated by changes to the microbial community [
10,
11,
12,
13,
14].
Several variables positively and negatively impact the earthworm population dynamics, such as the total soil nitrogen, organic matter of soil, available potassium and phosphorous, soil texture, and often pH-controlled earthworm population. Natural system disturbances modify the soil biota habitat and render it undesirable in certain instances. The increased use of chemical fertilizers and pesticides threatens soil species. Soil cultivation can cause or expose earthworms to mechanical damage, and the machinery could, in some cases, produce antagonistic disorders for earthworm’s development, growth, and fertility. Numerous environmental factors contribute to mates’ death, earthworms, who work in the soil for millennia and make our soil fertile and profitable [
15,
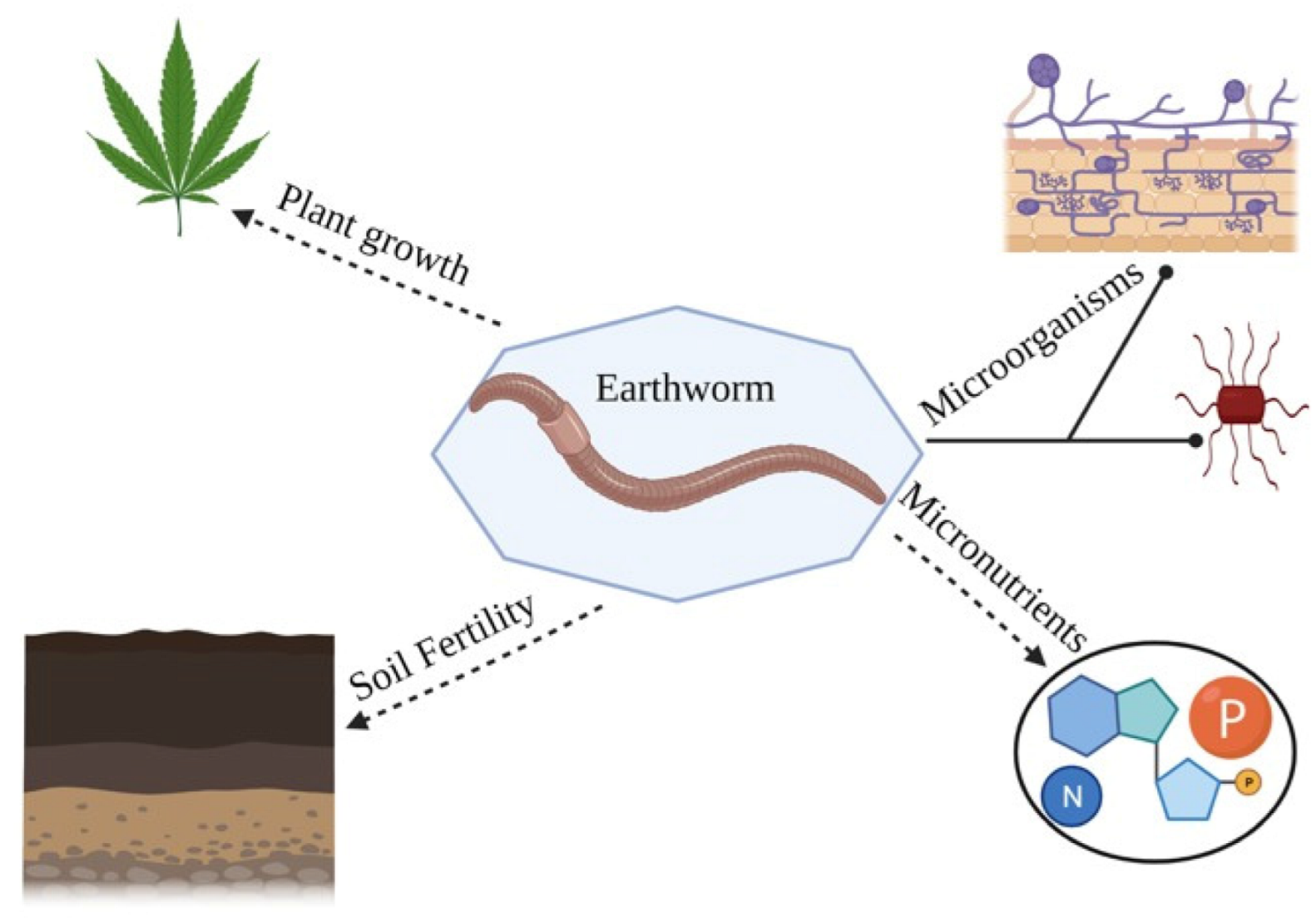
16]. These factors influence earthworms to maximize soil and sustainable production and maximum productivity for sustainable land management. An understanding of these factors is extremely important. Therefore, earthworms’ role in soil fertility and their population dynamics factors had been studied. Moreover, if any of the trends of the influence of the different functional groups on soil microorganism’s diversity (archaea, bacteria, and fungus), structure, abundance, and at the various sites (earthworm intestines, bulk soil, burrows, casts, rhizophorous, etc.) can be obtained from existing literature. Therefore, the present review aims to discuss the impact of earthworms on soil fertility, microbial population, and earthworm interaction among agriculture practices.
2. Microorganism and Earthworms
The soil organic matter (SOM) breakdown effect of earthworms has already been studied. While it is well understood in organic matter (OM) Turnover that soil fauna is of significant importance, the indirect influence on microbial communities, microorganisms, and soil fauna complex interactions are less understood. Microorganisms mainly achieve the breakdown of OM biochemicals. At the same time, earthworms can affect microbial decomposers’ actions by directly grazing microorganisms and the nearby area prone for microbial attacks after OM is comminuted. Microorganisms can sometimes serve as a food source for earthworms. The quantities ingested and ability to assimilate micro-ate and to digest bio-organisms differ with worm species, food substrates and the existence of an ecosystem in which they reside, which can greatly affect how much and what is assimilated by earthworms [
17,
18,
19,
20]. The results of the ingestion, digestion and relaxation of OM decomposition and microorganism’s species are directly affected by earthworms via the process of gut-association. After microorganisms are passed through the intestine of earthworms (some resistant bacteria, primarily protozoan and fungal spores), they are the inoculum for the new colonization of earthworm casts (EC) by microbial. Some bacteria are activated in the intestine, while others are unchallenged.
The microbial composition of the earthworms’ intestinal contents represents the ingested soil ingested. Besides, biomass, the numbers, and microbial communities’ behavior in the gut of earthworms are also shown to be different from those in undistributed soil [
8,
17,
18,
21,
22]. Singleton et al. [
23] studied intestinal and earthworm casts and found Pseudomonas, Penibacillus, Actinobacterium, Burkholderia, and Spiroplasm. Some bacteria are known to degrade hydrocarbons, such as Acidobacterium, Alkalinenes, and Pseudomonas. However, a 98 percent decrease in the total coliform density after the pig slurry passed through the
E. epigeic earthworm intestines.
There is a selective decrease in the
Escherichia coli BJ 18 coli in cattle dung in the gut of different earthworms of a genus,
Lumbricus [
17,
24,
25]. There are competitive interactions between those eaten in the gut and the endosymbiosis microorganisms. Indeed, it has been found that earthworm mid-gut fluid has a selective suppressive function and activates certain soil microorganisms.
Meanwhile, improvements in the intestinal bacterial population have been observed as a result of food resources. These changes have not obscured the distinctive characteristic that distinguishes the different ecological groups [
1,
26,
27]. The Fungi Spores (
alternaria alternata), for example, that survived in the medium good environments, began to germinate in fresh excrement and grew active. The destiny of the microorganism via the earthworm digestive tube is an important element for deterioration and development of the soil microbial environment. Freshly, rapid, and standardized microbiota shifts in gut selective impacts on ingested microorganisms in their presence and abundance have been observed [
28]. These particular effects can change the direction of decomposition, likely by changing the composition of microbial communities involved in decomposition. Previous studies aimed to evaluate intestinal transit’s impact on different organic residues’ biomass, enzyme activities, and microbial communities. Earthworms could change microbial physiology in the community and trigger enzyme activity during pig slurry vermicomposting [
18,
29,
30,
31,
32]. Studies involving six earthworm species and over ten liter soil and fungus species have shown that earthworms prefer to digest quickly, developing fungal species typically associated with early decomposition phases [
1,
33].
3. Earthworms Modify Microbial Communities in Soils
Earthworms can have a neutral, harmful, or positive impact on the microbial richness and diversity of ecosystems, for example, depending on the earthworm species and the microhabitat which is considered whether the study is focused on intestinal earthworms, plasters or soil. Neutral effects in earthworm bacterial communities have not been shown by the introduction of
Endogeal Aporrectode trapezoids on all soils to the sum of bacterial OTUs (operational taxonomic units) or Chao1 (Chao1 rishness estimator) [
34]. The burrows containing four species of Lumbricus relative to bulk soil are beneficial for their bacterial wealth and diversity. However, these authors have demonstrated that earthworms lack fungal diversity. In the vermicomposting process the influence of earthworms on soil microbial diversity has been also examined. Eudril sp. Epigeic earthworms, in particular. earthworms epigeic. Uh, or E. Uh, or Fetidas, which demonstrated that different time scales were taken into account in the analysis of bacterial diversity, increased the diversity of the substrate during its first vermicomposting phases at least [
35,
36,
37].
When considering the earthworm gut and casts, conflicting findings were observed. Earthworms have adversely affected bacterial wealth in the gut and casts. Bacterial species richness (estimated from cultivable bacteria) has been reduced by epigeic
Eisenia gut. Epigeic soil intakes have also decreased the microbial diversity seen in the intestines and casts of the
Eudrilus spa.
L. rubellus. Such a decline in micro biodiversity was due more precisely to the enrichment of bacterial taxa capable of degrading benzoic and aromatic compounds in the earthworm cast groups after soil intake [
32,
33,
36,
38].
In general, these studies show that Egert et al. [
39] had only minor variations between micro-habitats and earthworms that vary in their effect on microbial populations. Bacteria and archaea population structure in intestines, casting, and surrounding soil for the earthworm
anecic (Lumbrics Terrestris) property. On the other hand, the gut microbiome differed from bulk soil for the same species. The contrasting results illustrated here [
40] may partly explain the different techniques used for microbial diversity research. Whereas some findings are obtained using Terminal Restriction Fragment Length Polymorphism (T-RFLP) of bacterial 16S rRNA clone libraries, the resolution of diversity estimates has been stepped up with other studies. Earthworms often have a global effect on soil conditions upon their microbial soil contents [
33,
36,
39,
41]. This is especially relevant for the quality of nutrients. The effects of the earthworm epigeic
E. E. fetida on the biomass and composition of soil microbial population, evaluated through PLFA (Phospholipid fatty acids) and culturally capable bacteria counts, were more pronounced in low-nutrient areas where the bacterium stimulated earthworm intestines [
38] (
Figure 1).
4. Soil Structure and Activity of Earthworm: Implications for Soil and Water Systems
For most soil functions, including soil fertility, soil structure is crucial. Contribution of earthworms to soil and development structure via the humus development, mineral weather, and mixing of the components to produce stable aggregates, that is to say, organo-mineral complexes accumulated either on the soil profile or surface. The hydraulic and mechanical soil properties are also influenced by their burrowing activities, creating macro-pores that significantly impact water penetration and are essential to supplying plants with water and regulating surface ripples and erosion [
42].
Earthworm behaviors such as drought reaction, feeding, or avoidance of predators, cold temperatures, and soil oxygenation are powered by burrowing. Depending on the ecological community of earthworms, pore morphology varies.
Anecic earthworms dig wide, vertically oriented galleries (higher than 1 mm in diameter), reaching into the soil at depths greater than 1 m. In the vertical direction, no preferentially driven Endogeic earthworm galleries can be found. The burrow is smaller in diameter and not as profound as
Anecic burrows [
43]. In the litter and first few centimeters of the soil, epigeic earthworms live; therefore, there is little impact on soil macroporosity. Ernst et al. [
44] showed that ecological classes of Earthworms control soil water because, instead of burying,
L. rubellus leaves litre on a soil surface and avoids evaporation.
A. caliginosa caused higher rates of water penetration and faster discharges of water into the ground, possibly because of provisional burrows and constant repairs.
Earthworm burrows impair the water supply for crops. Studying greenhouse-growing rice, they have a detrimental effect in water deficiency treatment but a positive impact on the plant’s growth and, in the presence of the endogenous worm,
Reginaldia omodeoi, previously known as
Millsonia anomala [
45], there is a decreased water availability for this compact earthworm due to the lower soil water retention potential. However, in macro-pores formed by earthworms, preferential water flow occurred and was recorded for various soils: rice paddy flooring, temperate loamy flooring, and temperate clay flooring [
46,
47]. Preference flow raises the risk of nitrogen and pesticides leaching and eventually pollutes sub-surface and groundwater. Earthworms, however, have a generally favorable impact on soil porosity [
48,
49,
50].
Earthworm burrows may be a water supply to percolate the soil, but changes in water penetration may also impact the surface hydrological mechanism. In Ohio,
Anecic earthworm burrows decreased soil erosion by 50 percent due to the increased infiltration rate. Amynthas khami’s biogenic aggregates caused a 70% decrease in the runoff. Amynthas khami biogenic aggregates were responsible for a 75% reduction in runoff in Vietnam on an experimental field with a 40% slope [
51,
52]. Water penetration and soil macro-porosity are also increased by endogenous species, which appears to decrease runoff. However, some endogenic organisms have been shown to produce small-size casts that benefit the sealing of surfaces and lead to soil erosion. This effect was seen in
Pontoscolex corethrurus, a tropical earthworm.
It is nevertheless significant that, given the massive population increase in Brazil following the land use change from forest to pasture, this species was negatively impacted [
48,
51]. Species of earthworms that cause water-stable casts decrease their susceptibility to sprinkling and ruin, but they can reduce water penetration by increasing the surface volume density. Such inconsistencies between the impacts of earthworms on soil erosion, penetration of water, and soil structure, may be because this impact depends on the design of the soil and also on the following: (1) the available organic matter quantity on the soil surface, (2) the earthworm species, (3) the abundance of earthworm, and (4) the rainfall system [
53].
Typically, burrowing and casting practices in temperate and tropical soils are effectively in control of soil erosion.
Anecic earthworms have increased their soil roughness in temperate climates, enhanced through organic residues and “middens” forming, reducing the rushing surface. In Finland, surface rinses were negatively linked to the dry biomass of
L. Terrestris during precipitation events.
Anecic earthworms have been detected as a significant independent variable that causes ruin and erosion, and erosion rates have been degraded exponentially in three soil treatments, with reduced, enhanced, or entire earthworm populations [
42]. Endogeic and
Anecic casts increase soil structural stability on the soil’s surface and increase erosion resistance. Significant soil quantities can be described, such as 2 to 10 kg m
−2 in temperate climates, corresponding to a layer of earthworms 5 to 25 mm thick. Although some soil degradation cases are reported in the literature due to compacting earthworm species, earthworms seem to increase soil’s structural stability and soil porosity [
42,
54].
5. Soil Fertility and Earthworm’s Role
The EW have long been known to affect soil fertility; they were discovered in 1881, and Darwin’s book The Formation of Vegetable discusses their habits and influences. Further studies were published thereafter. Land macro-invertebrate are a key element in the processing, disrupting or producing biogenic structures that improve soil fever and productivity of organic matter (SOMs) and the dynamics of nutrients [
39,
41,
48]. SOM is a major active carbon reservoir as well as a central component of soil fertility. These ingredients promote soil structure and water retention, neutralize cations, and buffering pH [
44,
49]. The soil aggregates were proposed as structural units for that aim because it was thought that soil holds together plant growth and decomposition processes and helps the nutrient cycle [
12,
16,
19,
20]. The EWs are the main component of soil fauna communities. They are often linked to fertility and productivity in cultivated tropical soil organic matter. Through the regulation of mineralization and humification processes in such an environment, invertebrate populations, especially EWs, can play a vital role in SOM dynamics. EWs differ in ecological categories on the soil biological process and on the level of fertility. Anecic species are actively involved in deep mineral soil layers, endogetic species are present in the highest mineral soil layer and epigeic species are present on the ground surface. Most of these categories are combined to ensure that soil fertility is maintained [
22,
30].
EWs perform important functions by producing aggregates and openings (biostructure) to help provide nutrients (N, K, and Ca) as well as serving as pores (biostructure). The impact of EWs on dynamics of organic matter differs dependent on the space scales and time used. Endogetic EWs increase initial SOM turnover in humid tropical environments by indirectly influencing soil C as microbial activity enters. EWs were also reported to increase the addition of macro aggregates from the cover crop C as well as to micro aggregates consisting of macro aggregates. As a result of the increased organic C and N transfer into soil aggregates, EWs may be able to help agricultural systems stabilize and accumulate SOM. Furthermore, EWs improve nitrogen mineralization by influencing the microbial population both directly and indirectly [
12,
17,
20].
The studies have shown that more EW activity than total input was produced by adding recycled crop residues, organic and inorganic manure, slashed vegetation, and weeds by soil nitrogen for plants. The study found that an enormous increase in soil pH is an important part of the EWs. EWs have an impact on N cycling based on the type of cropping system and the fertilizer used (mineral versus organic). In addition, the EWs can increase the availability of nutrients in a system with reduced human impact, in tillage, less mineral fertilizer and less organic matter. The role of EWs in improving the fertility of soils is old science, but scientific results better explain this today. More details have been reviewed on this topic. Lumbricidas, which includes Lumbricus genus, Aporrectodea and several others, are the main family of EWs in the enhancement of agricultural land. Lumbricides come from Europe and have been transported to many parts of the world through human activities [
35,
36].
Earthworms aid fertility in many ways. For example, the earthworm moves nutrients deep into the soil and leaves them in the top layer. A vermicompost forage provides a soil food source to help earthworms do this job and brings organic matter into the soil in the process. The composting process uses different techniques that allow the decomposition of organic matter and the nutrients to become available for the plants while keeping the soil intact. When improved structure, improved soil mixing, and greater organic matter production are employed, earthworms contribute to increase the availability of nutrients for plants. many worms consume much of the substrate but contain a small percentage (5–10%). The rest is excreted as earthworm-like casts, which are high in nitrogen, potassium, micronutrients, and beneficial soil microbes. Earthworm gut bacteria consume harmful chemicals and also breakdown organic waste. Auxin is a plant hormone that stimulate root growth and mass. Due to microorganisms found in the intestine of earthworms and in their excrement, the process of nitrogen fixation is relatively high in the soil. The percentage of nitrogenase casts is also higher, promoting greater nitrification in the soil. higher levels of nitrogen fixation in soils contribute to more nitrogen in the casts [
55,
56].
Most soil functions, including soil fertility, are dependent on soil structure. Through the formation of humans, weathering, and mixing, the Earthworms contribute to soil structure and formation and to the development of stable aggregates, for example, complexes of organo-mineral deposition in soil surfaces in the soil profile [
16]. They also have an effect on soil mechanical and hydraulic properties as a result of their burrowing operations, which generate macropores that significantly affect water infiltration and are thus essential to water supply and surface runoff and erosion control [
42].
The interaction of physical forces such as contents of soil water, large biota behavior such as roots of plant and earthworms, organic matter in the soil, and tillage all contribute to the soil structure development from the micro to macro scale. Due to their low assimilation ability, the earthworm has the most noticeable effect on reorganizing soil structure when moving through it, feeding on it, and releasing material [
10,
11].
Depending on the species and the soil interaction, earthworms may either make the soil compact or loose. The majority of soil structure physical improvements through the production of casts in vertical and horizontal burrows are caused by anecic and endogetic species. Cast production modifies the density of soil bulk by integrating organic matter into the soil. For example,
Reginaldia omodeoi, an endogetic earthworm, has increased the density of soil bulk from 1.24 g cm
−3 to 1.31 g cm
−3 and 1.37 g cm
−3 in 2 distinct studies [
2].
Bulk density significantly increased from 1.12 to 1.23 g cm
−3 as well. In the presence of a tropical endogeic earthworm,
Pontoscolex corethrurus, a decrease in porosity from 58 percent to 53 percent was also observed. The interaction of soil de-compacting (small Eudrilidae family) and compacting (R. omodeoi). soils contributed to the maintenance of the soil structure in a long-lasting tropical field experiment. Earthworms also affect soil aggregate size distribution. The de-compacting earthworm (for example,
Millsonia anomala) destroyed the compacting macroaggregates while the de-compacting earthworms also played a similar role with the casts. This shows that the action of earthworms in regulation of soil structure dynamics is widely variable. Compacting earthworms (for example,
R. omodeoi, yams and maize crops) increased from 30 percent to 54 percent and 25 percent to 42 percent, respectively, the relative proportion of aggregates >2 mm [
14,
15,
16,
17].
It is estimated that earthworms produce casts in the rates of 40 to 100 tons, and therefore make a huge contribution to building stable aggregates of soil [
18]. In just 2 months, 18–42 per cent of the soil material could be converted into macro-aggregates [
19]. Earthworms clearly improve the porosity of the soil as well. In a study, in comparison with therapies in which earthworms were present, the fine (<0.4 mm) part of soil aggregates was dominant during treatment with no population of earthworms [
15]. The effect was dependent on the balance between cast and soil degradation and the endogeic earthworm modified the porosity of soil to control water flow in land [
20] (
Figure 2).
6. Nitrogen and Phosphorus in the Soil and Earthworms
Earthworms increase organic matter mineralization, thus increasing the amount of available nitrogen in the soil. A great deal of nitrogen will enter terrestrial ecosystems as a result of the decomposition of the Earthworm biomass. It is estimated that about 60–70 kg of nitrogen has been returned to the soil each year in English Lumberland, woodland in England by means of
Lumbricus Terrestris decomposition. During the decomposition of the earthworm’s tissue, nitrogen quickly breaks down into a mineral, decomposed, and the mineral fertilizes the soil. the comparison to the body tissue, whereas Devine et al. [
57] found that 70% of nitrogen mineralization in earthworms took place in 20–10 days. The important aspect of the studies conducted by Satchell and Devine, however, is that they showed the pace of soil conversion as different for different kinds of tissue, such as different species of worms. Most activity occurs in the castings as a result of the use of earthworm gut and earthworm intestines, rather than the fertilizer in the soil [
57,
58].
Earthworms are heterotrophic species that help plants and other heterotrophic organisms degrade organic matter and molecules. Earthworms speed up the decomposition of organic matter by comminuting it to increase its usable surface area. Some organic compounds, in the form of small organic compounds or mineral nutrients, are released into the atmosphere after digestion. Plant nutrients, in particular, such as nitrogen (N), are frequently recycled. New earthworm activity aids in the dissolution of in organic matter. They release nitrogen as part of their bodily waste, and make new casts, which contain NH4
+, allin, and allin, in their urine, which are broken down by the soil organisms, and they supply earth that gets absorbed by mucus in their intestinal secretion, which is obtained from their bodies [
46,
47,
56,
57,
58].
Phosphorous is an essential plant nutrient that is involved in the accumulation and conversion of energy in living cells’ metabolic reactions. It also encourages early vegetative growth and leads to grain crop maturity. Although P is a vital factor in growth of plant, it is the second nutrient that limits growth of plant following nitrogen. P is less water soluble and comparably more mobile than other key soil nutrients, especially nitrogen, and available for plants. The weak availability of P in soil is responsible for soil pho, ion antagonism and metal concentrations (Ca, Fe and Al), which may precipitate with P ions [
55].
The casts contain more P than the soil without earthworms available in the surrounding area. As a result of this, Nuutinen et al. (1998) concluded that earthworms had a positive correlation with soil P accumulation in the soil. Increased phosphatase activity in the casts could be responsible for the increase in available P in earthworm casts. These increases in the amount of available P in earthworm digestibility may be due to increased phosphatase activity in the products found in the products. The authors also cited carboxyl groups present in large amounts in the intestine (6.8 and 6.4 in the anterior and posterior, and 5.4 in the other parts of the intestine), as well as baroyl which is associated with mucus secreted compounds, blocking and competing for P sorption. The chemical form of P and its concentration can be changed by ingesting and thoroughly mixing soil in the intestinal tract of earthworms. Inorganic phosphorus was produced four times faster in the casts than it was in the surface soil. Kuczak et al. (2006) estimated the total available P reserve forestry system, pasture and secondary forests could comprise 41, 38, 2 and 26 kg ha−1 casts, respectively.
Many studies have focused on small-scale processes in casting and burrowing, and in particular, on P. Extrapolation of results achieved at the scale of the environment is therefore tricky. The effect of earthworks on the cycling of nutrients is consequently significant. In addition to this spatial dimension, time variance is another obstacle for future models to better understand earthworms’ entire function. [
1]. These authors highlight the drilosphere as a ‘dynamic sphere’ in which earthworms affect soil and change continuously in time and space, depending on the cycles of earthworm’s activity and their size, the horizontal and vertical distribution of earthworms according to biotic and abiotic factors etc. This description can be generalized to P, and several environmental levels are combined better to understand the effect of earthworms on the P cycle.
7. Earthworms’ Effects on Crop Production, Agriculture Practices and Population of Earthworms
For millions of years, earthworms have inhabited the soil and constitute the most terrestrial ecosystems, with the most abundant underground biomass; therefore, earthworms and plants may coevolve. The advantages of earthworms were recognized for plant growth about a century ago. As a result, 46 and 54 percent of the studies were extensively reviewed by different authors regarding earthworm impact on crop production and health [
58]. In 246 experiments in tropical countries, Brown et al. [
21,
58] examined and concluded that in 53 per cent of the study, the bio-mass production with and without earthworms differed less than 20 per cent. The biomass production difference without and with earthworms in 53 percent of the trials was less than 20 percent. The presence of earthworms decreased biomass production by more than 20% in 4% of the studies, suggesting that earthworms are detrimental to plant development [
22,
59,
60,
61,
62]. In the remaining 43 percent of studies where earthworms enhanced plant growth, there was more than a 20 percent increase in biomass production. In the presence of earthworms, the difference in biomass production has been identified as being responsible for several environmental factors. Soil types, especially soil texture and carbon content, significantly determine plant yield variation [
22]. Since earthworms are so sensitive to the presence of pH, highly acidic soils give the greatest increase in production of biomass, as stated Laossi et al. [
63]. Functional plant groups are also an important driver: Earthworms produce greater biomass than annual species in perennial species, especially trees, whereas the presence of earthworms can sometimes have negative effects on the biomass of leguminous plants [
22].
Many studies, recording 83 cases located in temperate countries, have shown that over ground production in 79 percent of earthworm cases increased significantly, while 9 percent significantly decreased [
30,
64,
65]. However, as seen in Amazon, earthworms’ additions over soil carriage ability will lead to soil compaction. High earthworm abundance has a detrimental impact on crop production that is not well known. In addition, earthworms’ beneficial influence on plant growth could be due to five mechanisms. I i.e., (i) Increased porosity and aggregation of the soil, altering plant supply with water and oxygen, and (ii) increasing SOM (soil organic matter) mineralization, increasing plant availability with nutrients. Interactions with other species are one of three additional mechanisms: (iii) insect and parasite biocontrol; (iv) symbiotic stimulation; and (v) development by stimulating microbial activity of plant growth regulation systems [
22].
For pest biocontrol, earthworms could be successful. Earthworms
Aporrectodea rosea and
A. trapezoids decreased all intake severity due to soil-borne fungal pathogen, and earthworm
R. omodeoi minimized damage to rained rice plants caused by plant-parasitic nematodes
Heterodera sacchari. Earthworms have affected plant growth and nutrient levels of aphids. A herbivore’s success is known to rely heavily on the nitrogen levels of plant tissue. As documented above, changes in plant growth caused by earthworms are often linked to an increase in plant tissue nitrogen content. Earthworms’ impacts on plant growth are also likely to spread into the herbivore system. In reality, earthworms have been shown to promote the reproduction of aphids on herbs and legumes. However, the effects of earthworms on above-ground herbivores are probably different according to the type of soil and dependent on plant species. The reduction in aphid reproduction only in plants was probably caused by changes in herbal protection compounds that show that herbivores below ground and herbivores affect herbivores. Herbivorous success is known to heavily rely on plant tissue nitrogen, and as documented above, improvements in plant growth mediated by earthworm are often associated with increased plant tissue nitrogen levels. Earthworms can alter plant nitrogen levels by increasing the abundance of nitrogen in the soil. In contrast, the increase in the availability of nutrients in the presence of earthworms can increase plant resistance. Thus, the influence of earthworms on plant growth probably spread to the herbivore system. In reality, earthworms stimulate the reproduction of aphids have been documented. Furthermore, the earthworms’ impact on above-ground herbivores probably differs from the soil and relies on plant species. The decrease in aphid reproduction, probably only on plants before, was due to changes in plant defense compounds, which indicate that it is below ground herbivores and plant defense against herbivores affected by them. In several studies, stimulation of the plant performance of earthworms has been reported. Recently, it has been shown that earthworm-induced changes in plant performance can also impact plant food aphids. The effect of earthworms in aphids is caused by changes in the plant; because they feed aphids on phloem sap they are susceptible to changes in the quality of the host plant, especially in the content of nitrogen. For example, fertilizer use leads to an improvement in the reproduction of aphid species to increase soluble nitrogen levels in plants. [
45,
66,
67,
68,
69].
It was found that in the presence of earthworms, plant growth regulators production was at higher rate. Signaling molecules such as auxin or ethylene formed in earthworm casts could include these compounds, as shown by
Arabidopsis thalian loss of function mutant and earthworm research on plants and their defense. A stimulation of indole acetic acid (an auxin compound) produced by cultivable bacteria was also demonstrated. Most of the earthworm’s reports are optimistic about plant growth and health but seem controlled [
70,
71,
72].
Earthworm biomass and behavior can be greatly influenced by agricultural practices such as crop rotation, fertilization, use of pesticides, lime application, drainage, tillage, and irrigation. The numerous studies investigating the tillage impacts on earthworms concluded that in clay loam soils, populations of earthworms were decreased by deep plowing and intensive tilling. In sandy loams, the tillage results were complex and based on many aspects, including earthworm species in the soil. Earthworm abundance was encouraged by no-tillage management systems. Nevertheless, populations appear to rebound from less extreme agriculture forms within one year, given the disturbance is not repeated [
55]. Once a year, tillage impact on the population of earthworms was found to be less disruptive than that of birds feeding on earthworms [
67]. The tillage effects on the earthworm
Aporrectodea turgida indicated that tillage-induced disruption is likely to have a greater influence on biomass and earthworm populations than the availability of food in cold, humid agro-systems. Among other factors, mechanical weeding is responsible for ecosystem disruptions, physical damage to earthworms, and reproductive disruption [
73].
High input levels characterize agricultural systems. Organic crop inputs drive biological activity in agricultural soils. The contribution of organic materials from organic fertilizers, manure applications, crop cover, or crop residues has a positive effect on the behavior of the soil’s biological population, structure and scale. The organic fertilizers’ solid materials derived from animal and plant sources have been reported to increase earthworm populations. However, most chemicals influence earthworms by increasing plant production indirectly and, subsequently, rising plant residues, which remain on-field after harvesting. In the decomposition rate of surface residues, OM distribution over the soil profile, and the modification of soil physical properties, earthworms play an essential role. The management of OM is important for developing a sustainable earthworm populations and for ensuring that amelioration occurs in soil that is returned to the environment.
SOM and earthworm populations may also be affected by the conversion of grassland into arable land. Indeed, a sharp decrease in the abundance of earthworms after converting grassland into arable soil has been observed. However, converting arable land to grassland stimulated the species’ wealth and quantity even in the second year after its conversion [
59,
63,
74].
8. Population and Community Levels of Earthworms as Well as Factors Influencing Earthworms Population
Earthworms impact the physical and chemical composition of soil and contribute to organic and mineral soil transfers. Parameters such as the population of earthworm and size of the community, rate of reproduction rate, mortality, survival, and growth have apparent effects on casting and burrowing operations on the plot scale. Besides, concerning bioturbation procedures, the ecological and functional community of earthworms is very obviously important, mainly depending on the location and availability of food supplies. The earthworms are divided into detritivores, epigeic and
Anecic, which usually feed at or near the soil surface mainly on plant litter, and geophages (endogeic species), feeding more in-depth in the soil profile and ingesting large quantities of soil. As a result, the spatial distribution of populations and groups of earthworm fields varies widely and is, in general, closely related to the soil. A recent study by Feller et al. [
64] showed that the amount of soil created in various temperate ecosystems in different temperate ecosystems by worms ran from 2.2 t ha-1 years to 1 year and contributed to the aggregate distribution of earthworms [
15,
52,
74].
Nevertheless, as Bohlen et al. [
66] pointed out, most studies have focused on earthworm surface casts, which can be a small share of total cast production above and underground dejections. The spatial distribution of field earthworm populations and communities varies a lot and is usually closely linked to soil properties. These studies are to be conducted both in laboratory experiments and at the field level to provide an overview of how earthworms influence nutrient cycling worldwide. Unfortunately, regardless of the spatiotemporal factor, the phosphorous balance, the amount of P that is returned to the earth through earthworm activities and the total nutrient fluxes by earthworm biomass, remains misunderstood.
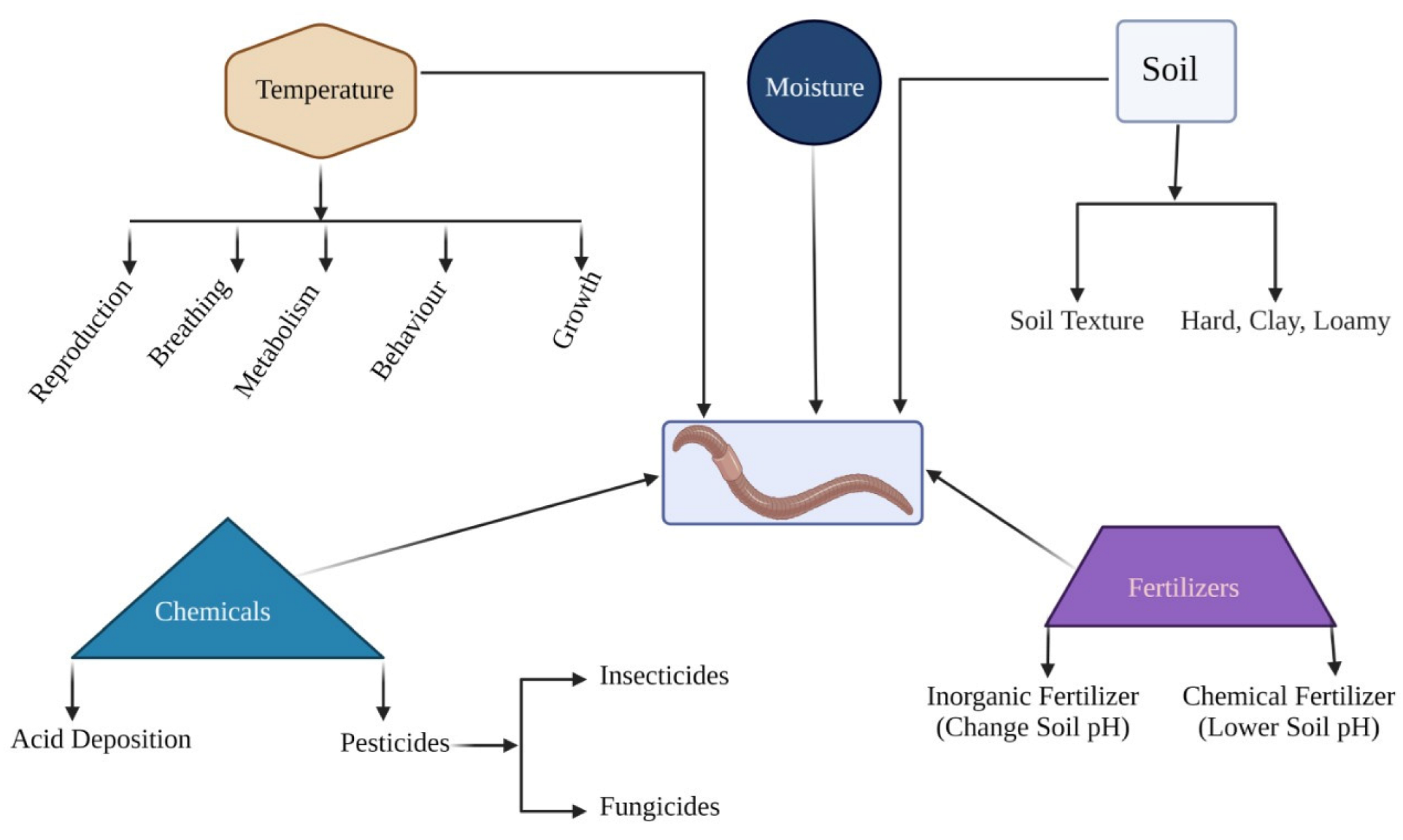
Several environmental factors influence the density, abundance, and distribution of earthworms. The most crucial factors that regularize the population of earthworms are the quality of organic soil, the soil type, the soil moisture content, the temperature of the soil and pH. The wealth and distribution of earthworms are greatly influenced by climate and biotic factors.
8.1. Type of Soil
The degree to which earthworms affect their abundance and distribution is linked to the soil environments in which they exist. Soil texture affects earthworms’ populations because it affects other soil characteristics, for example, hydration, nutrients, and CEC (cation exchange capacity). In light and medium loam soils, more earthworms are observed than hard clay, sandy, and alluvial soils [
59]. Hendrix et al. [
67] documented the relationship between the soil’s salt content and the abundance of earthworms. Furthermore, Baker et al. [
68] found a week-long relationship between the soil’s clay content and the sum of
A. Squirrel,
A. caliginosa, A, rosea, and
A. trapezoids. However, for
A. caliginosa, these species found the most remarkable positive relation with the clay material (
Figure 3).
8.2. Moisture
Typically, earthworms require adequate humidity to grow and develop properly. Water comprises between 75–90 percent of the bodyweight of earthworms. They breathe through humid skin, and surface blood capillaries should receive adequate moisture for performing respiratory activity. Enough moisture in the soil measures the activity of earthworms. The activity of earthworms in wet soil is greater than dry soils, so desiccation must be protected.
Between species and various areas of the earth, the need for humidity varies. Earthworms have adopted distinct methods to cope with dry soil conditions; they migrate to lower layers of soil, diapause, or form cocoons resistant to drought. Between 60 and 70 percent of humidity is optimal for the growth and production of earthworms. For earthworms, enough moisture accompanied by heavy rainfall is fatal. Earthworms are forced to migrate to the soil’s surface, where predation and ultraviolet radiation harm them because dissolved oxygen is substituted with excess moisture and anaerobic conditions induced [
66,
67].
8.3. Temperature
The temperature affects the reproduction, behavior, metabolism, growth, and breathing of earthworms greatly. Earthworms may only survive a certain maximum temperature range and increasing temperatures above that limit can kill them. Earthworms are more able to tolerate wet and warm or dry conditions than either dry or warm conditions. From species to species, temperature tolerances and earthworm preferences vary. Cocoons’ incubation time, fertility, and temperature also affect the growth period from hatching to sexual maturity in earthworms. At higher temperatures, cocoons tend to hatch earlier. For the growth of the indigenous Lumbricidae population in Europe, 10–15 °C is the optimum temperature. The best conditions for earthworm activities are the night temperatures in the soil not reaching 10.5 °C [
67,
68,
70] (
Figure 3).
8.4. Effect of Fertilizers
There are varying impacts on earthworms by inorganic fertilizers at each location. Inorganic fertilizers help earthworms and are detrimental to them. When applied, inorganic fertilizers change the pH of the soil and shift vegetation type and quantity. Chemical fertilizers, on the one hand, reduce the number of earthworms by lowering pH, but they also increase the number of earthworms by raising vegetative production [
69]. With the application of superphosphate and lime to pasture, Edwards and Bohlen [
55] found a four-fold increase in the number of earthworms. Lime is also beneficial because many earthworm species prefer calcium-rich soils and avoid acidic soil conditions. With the application of superphosphate fertilizers, it has been recorded decreased earthworm numbers in grass plots. The use of nitrogenous fertilizers can also encourage the aggregation of a large number of earthworms. Nitro chalk introduction in large quantities to various pastures due to increased grass production recorded increased earthworm numbers [
64,
65]. In addition, organic fertilizers increase the number of earthworms as more chemical fertilizers increase in arable land with the same N. Organic manures encourage more earthworm growth since they serve as additional food for earthworms, making crop residues more appealing to earthworms that maintain a high C: N ratio [
67,
71].
8.5. Effect of Chemicals
In the soil, acid deposition, biphenyls, polychlorinated, heavy metals, and pesticides are agricultural chemicals. When present in soil, these chemicals have direct impacts on the abundance of earthworms and their distribution by interfering with the productivity of earthworms, reducing their activities and eventually leading to the death of earthworms. The effect of pesticides on earthworms is determined by the form and rates of pesticide usage, earthworm age, species, and prevailing environmental conditions. Using large-scale contact with pesticides in action, even in deep soil, kills [
67,
71].
There are records of fungicides and insecticides being specifically harmful to earthworms. However, the pesticides are most commonly applied after canopy closure to prevent earthworms from touching the chemicals. Pesticides are in the soil sprayed on to this treatment are safe to enter the ground or plant until the next time they are washed away, but when contaminated leaves or plants are dumped onto soil, they may be a danger to residents and livestock. More copper-based fungicides are harmful to earthworms, which are more common in organic farming. Fungicides based on copper, popular in organic farming, may be immune to earthworms. Copper oxychloride’s direct poisonous effect on earthworms in vineyards in South Africa indicated that copper in earthworms could accumulate. For example, paraquat at the commercial dose showed that the reduction in cast formation was better than glyphosate, also used at commercial dose compared to a control plot with no use of herbicides. Herbicide treatments are performed on the soil’s surface before planting or the emergence of weeds and plants to affect anecdotal and epiges feeding on surface litters [
69,
70].
In laboratory studies, it is difficult to determine the toxicity of pesticides to earthworms because successful rates, such as the use of pesticides in the field, are generally low compared to the toxic levels used in laboratory studies. Lethal effects are rarely observed if products are used under field conditions at standard application rates. However, depending on the earthworm species and the product used, a sub-lethal impact on reproduction and growth can occur. After five weeks of exposure to fenamiphos (used as a nematicide and an insecticide) and endosulfan (insecticide), with no significant reduction in growth, a significant reduction in growth with methiocarb (molluscicide, acaricide, insecticide and used as a repellent for bird) and ridomil (fungicide) has been recorded. Therefore, the inference that can be drawn from the available literature is that most pesticide toxicity data are based on
E. Fetida’s answer in standard tests of optimum population density with well-fed earthworms. Improved eco-toxicological risk assessments include information on the types of pesticides used in the environment and earthworm population exposure rates while considering fluctuating environmental conditions [
72] (
Figure 3).
9. Conclusions
Managers need to consider the following limitations in ecosystem management before using earthworms: external constraints imposed by the internal boundaries of the socio-economic system, ecosystem biological and physical properties, and those correlated with ecosystems’ multi-functionality. For instance, earthworm abundance in surrounding regions needs to be understood before planning an area devoid of earthworms. Socio-economic studies should be undertaken when a plan for introducing organic matter is designed to decide whether imports from other areas are required, to ensure that transport costs are not too high in terms of financial and carbon costs, and to determine whether other production sectors, such as industry, forestry or agriculture, can compete. Tools developed in industrial ecology, such as territorial metabolism or life-cycle analysis, may help address these questions [
71].
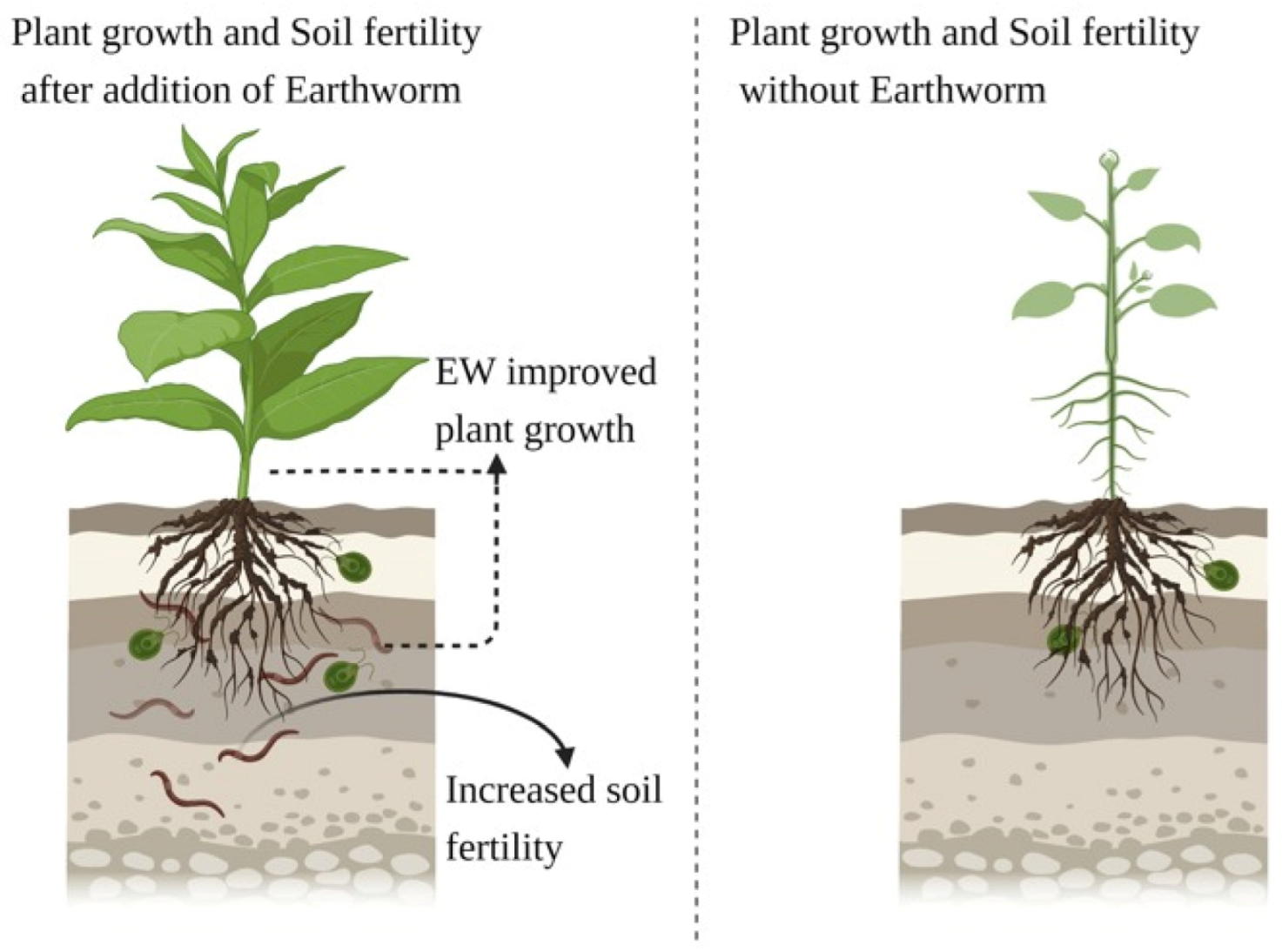
In soil ecosystems, earthworms are significant biological factors. They are adaptive to cultivation techniques and can therefore be used as soil health bioindicators. Earthworms have been proposed as possible measures of farmers’ sustainable activities, thus optimizing various farming systems. In general, the effects of earthworms on soil fertility and plant growth are positive. They strengthen the structure of the soil and stabilize SOM fractions in their casts. In the short term, mineralization is increased, which makes mineral nutrients accessible to plants. Earthworms trigger the release of molecules similar to plant growth plants. Tillage is usually harmful to earthworms, while SOM content increases the positive effect on earthworm populations. Considering the potential contribution of EWs to soil fertility management, there is a need to consider them in agroecosystem management decisions. The fact that EWs can specifically affect soil fertility that may be of great importance to increase sustainable land use in naturally degraded ecosystems and agroecosystems. Proper earthworm management may sustain crop yields while fertilizer inputs could be reduced. Nevertheless, earthworm studies will be addressed in future work on soil properties, vegetation types, climatic details, identification of earthworms on the species level, and the existence of other soil micro-fauna.



{kind=link}
{kind=link}
{kind=link}