
Estimation of Terrestrial Net Primary Productivity in the Yellow River Basin of China Using Light Use Efficiency Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
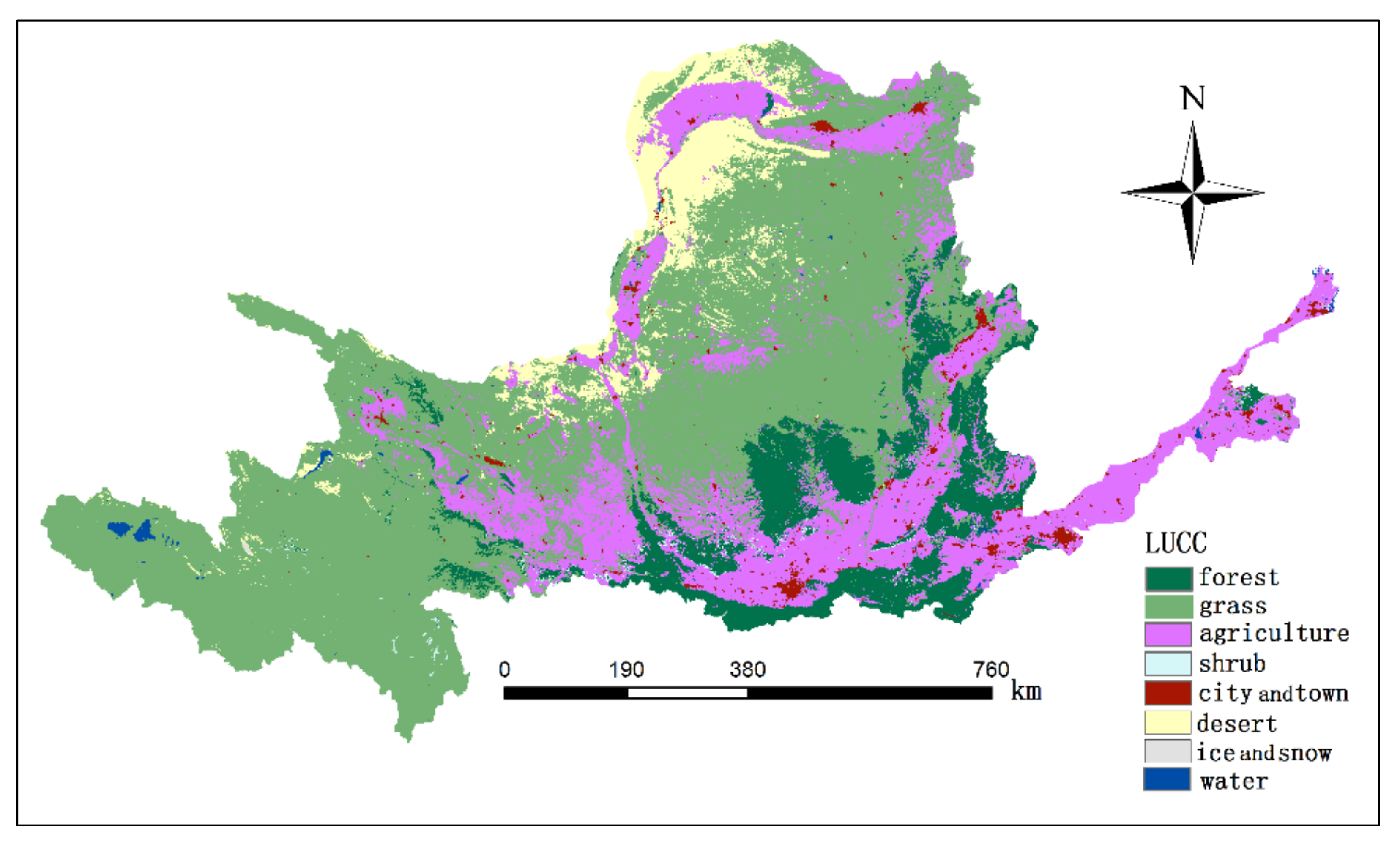
2.2. Data Preparation and Processing
2.3. Simulation of NPP
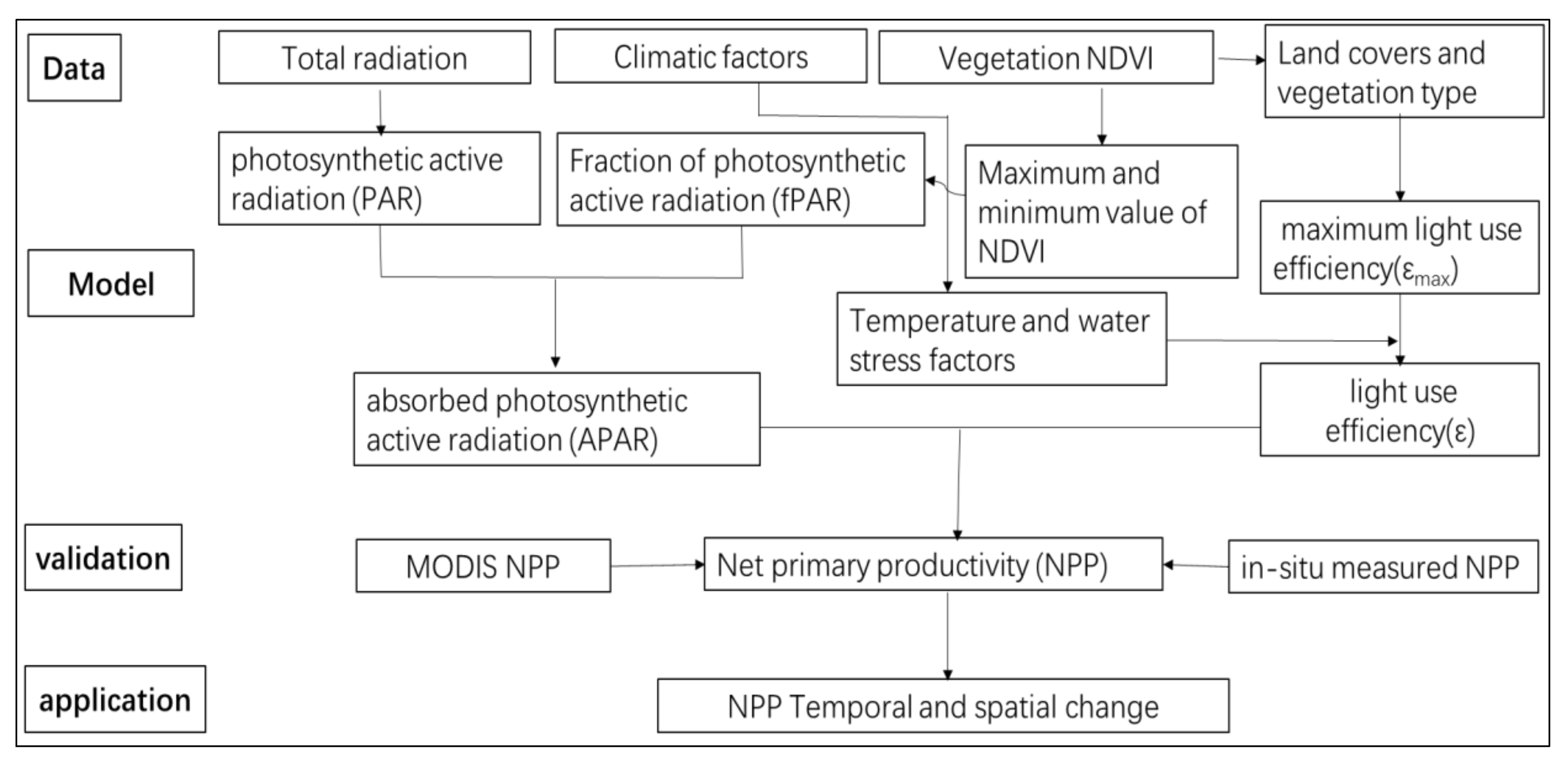
2.3.1. Analytical Framework
2.3.2. Simulation of NPP
2.3.3. APAR Estimation
2.3.4. FPAR Estimation
2.3.5. LUE Estimation
2.3.6. Temperature Stress Factors Estimation
2.3.7. Water Stress Factors Estimation
3. Results
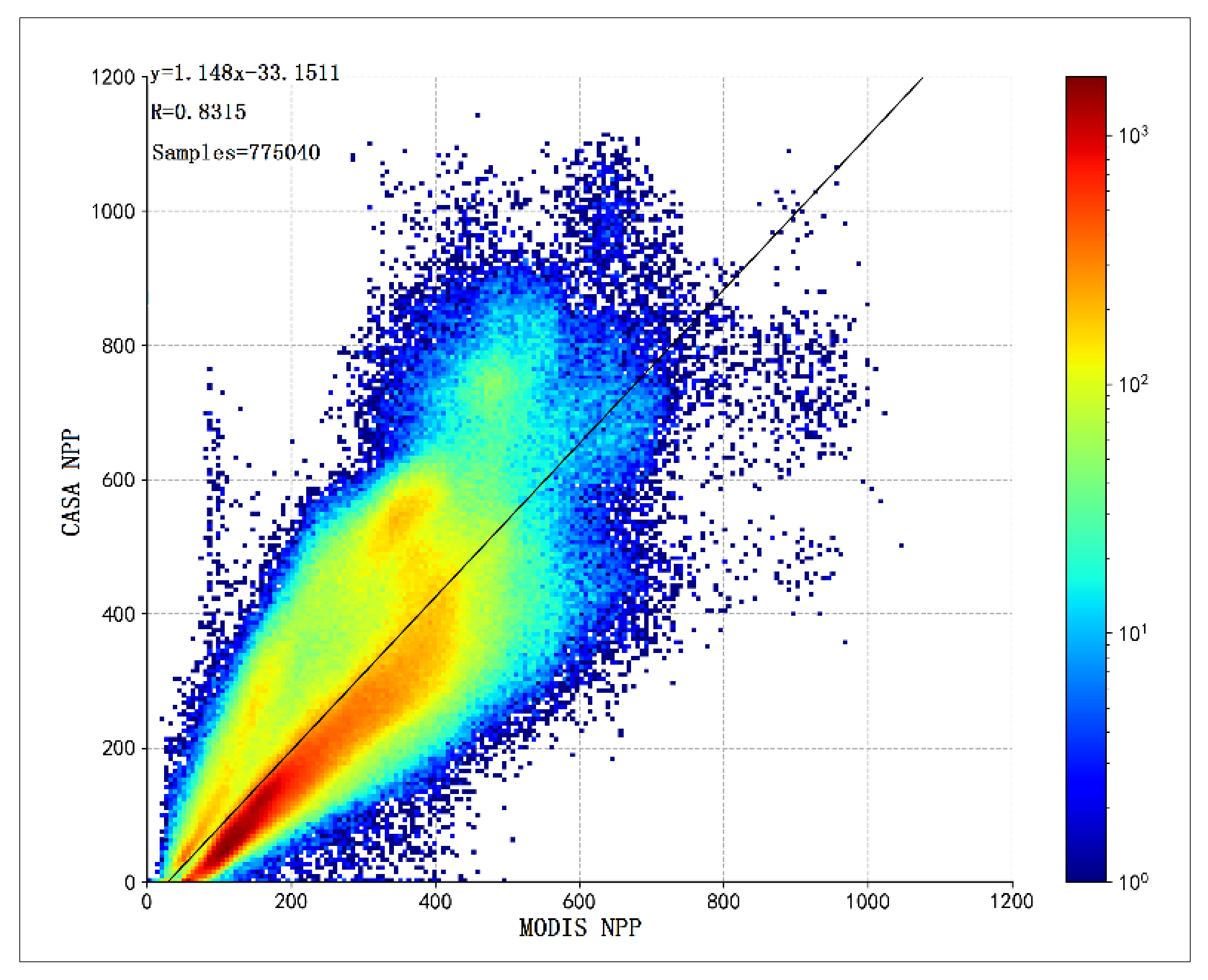
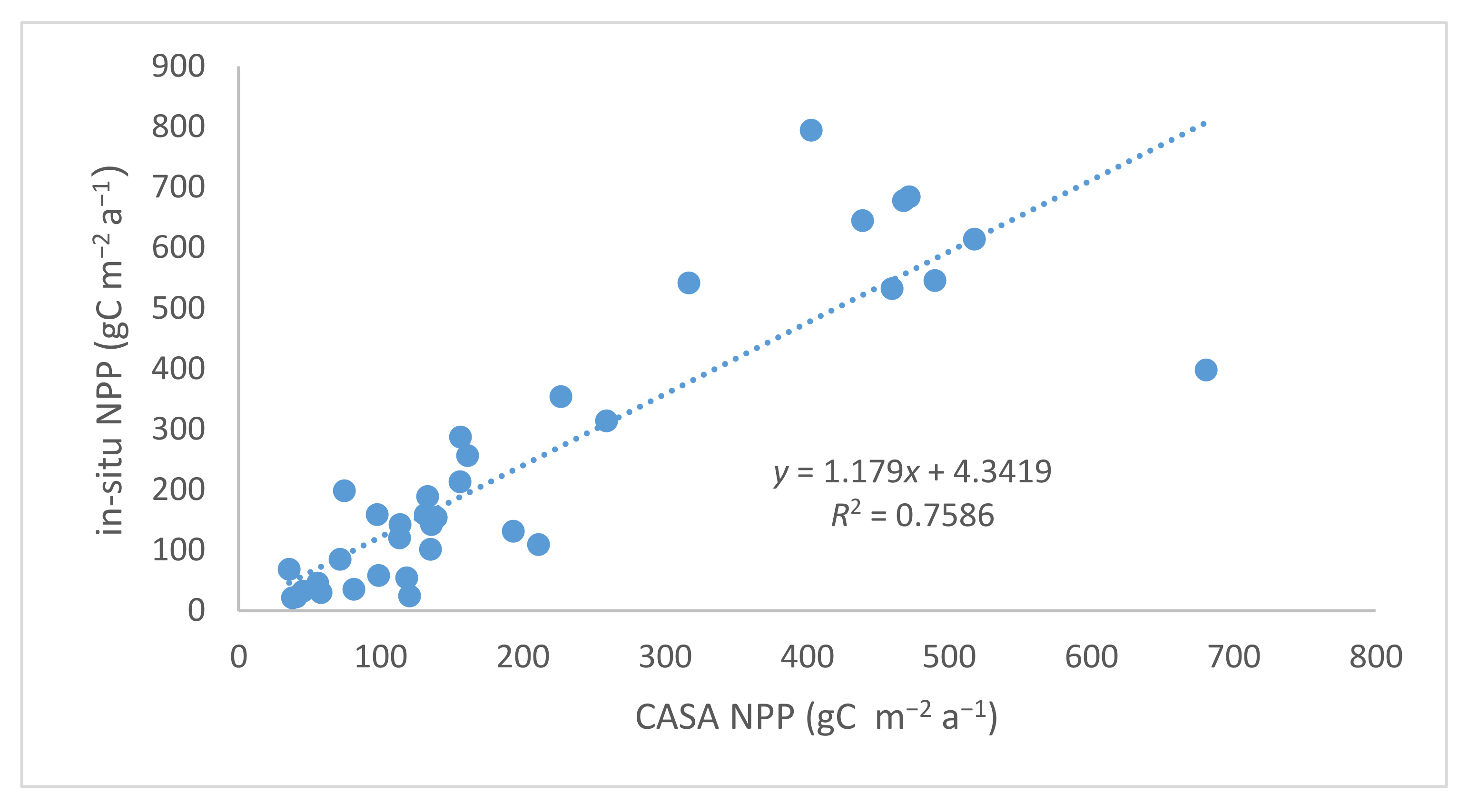
3.1. Validation of CASA Model NPP
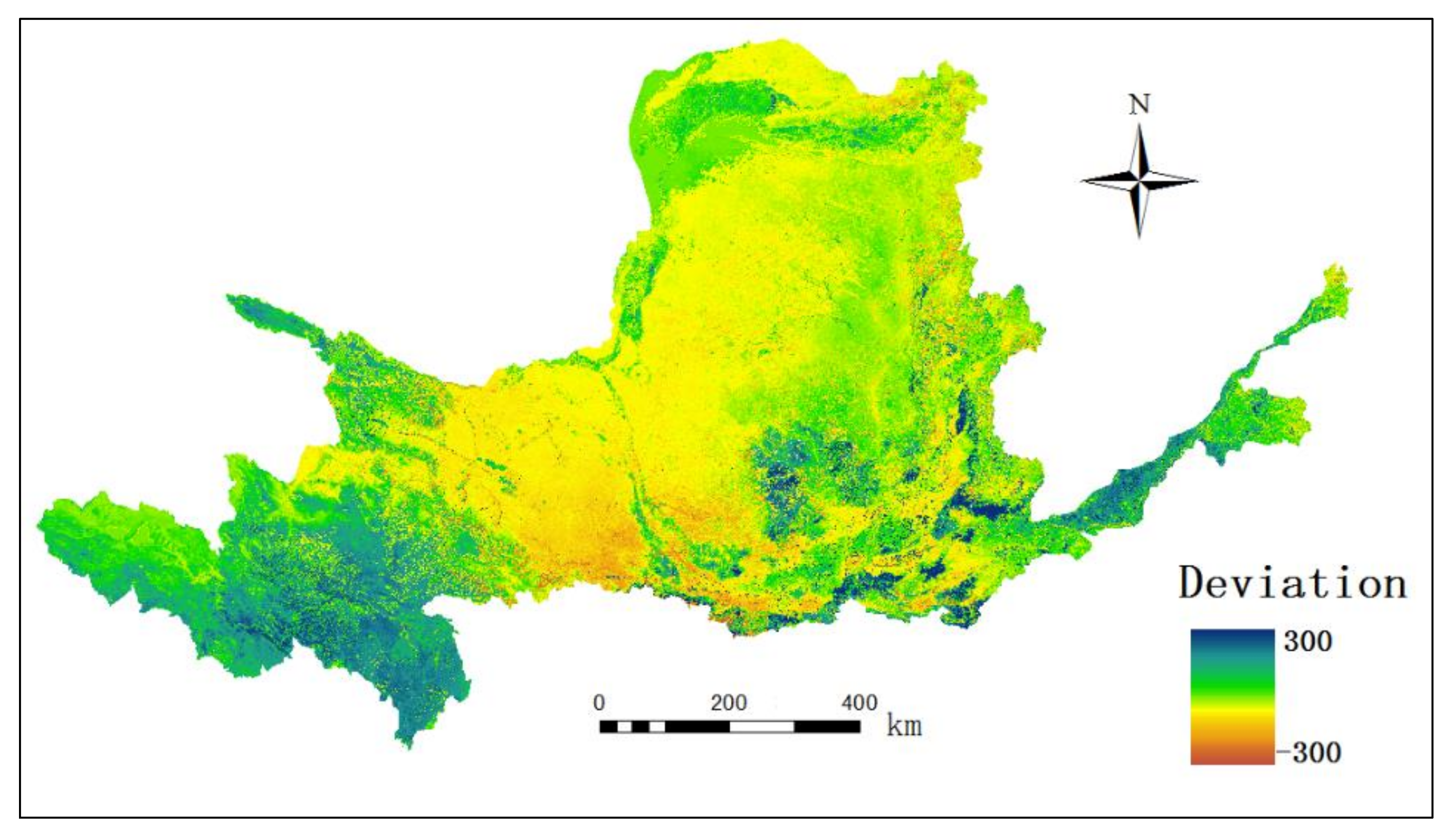
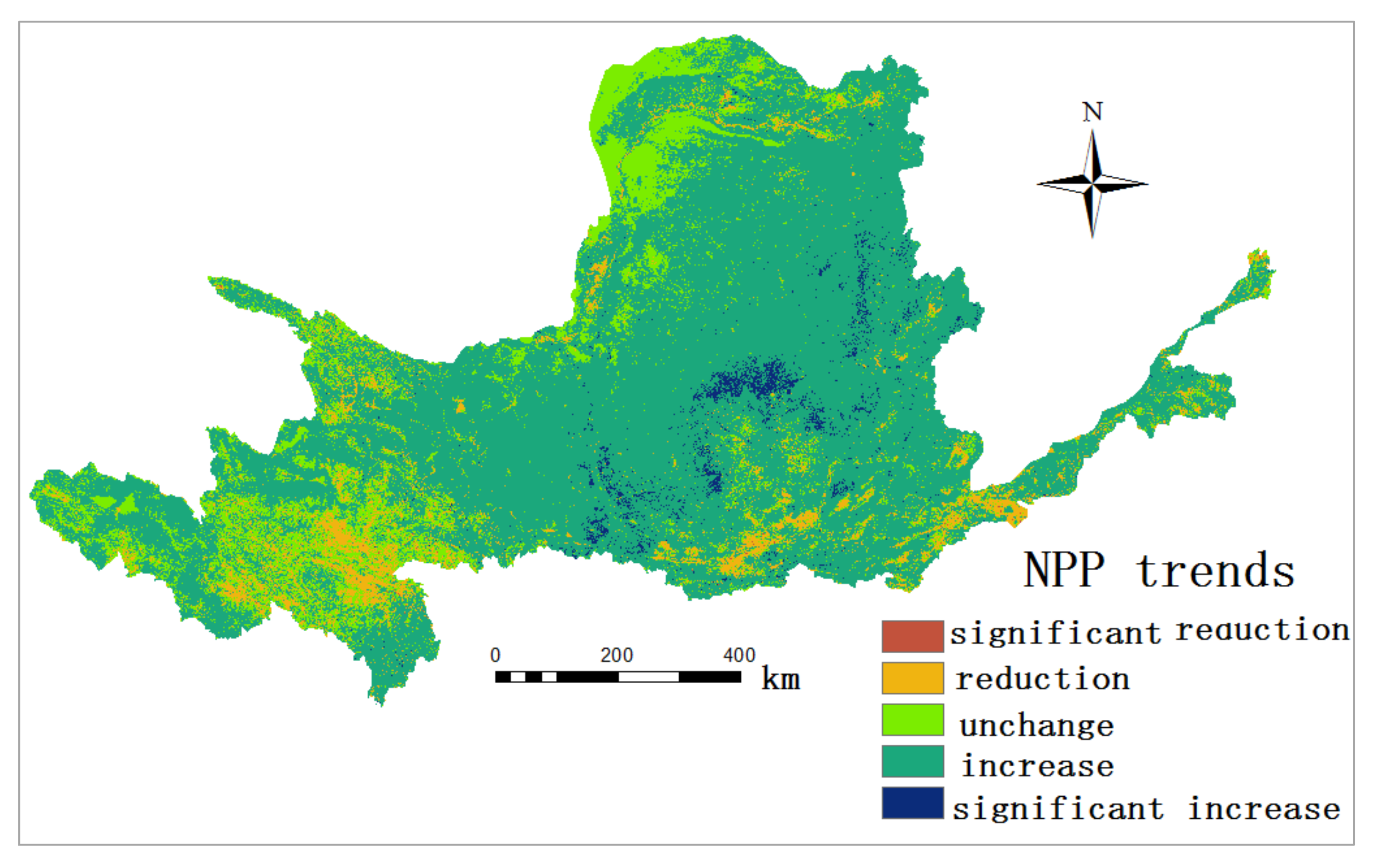
3.2. Spatial Distribution of NPP in the YRB
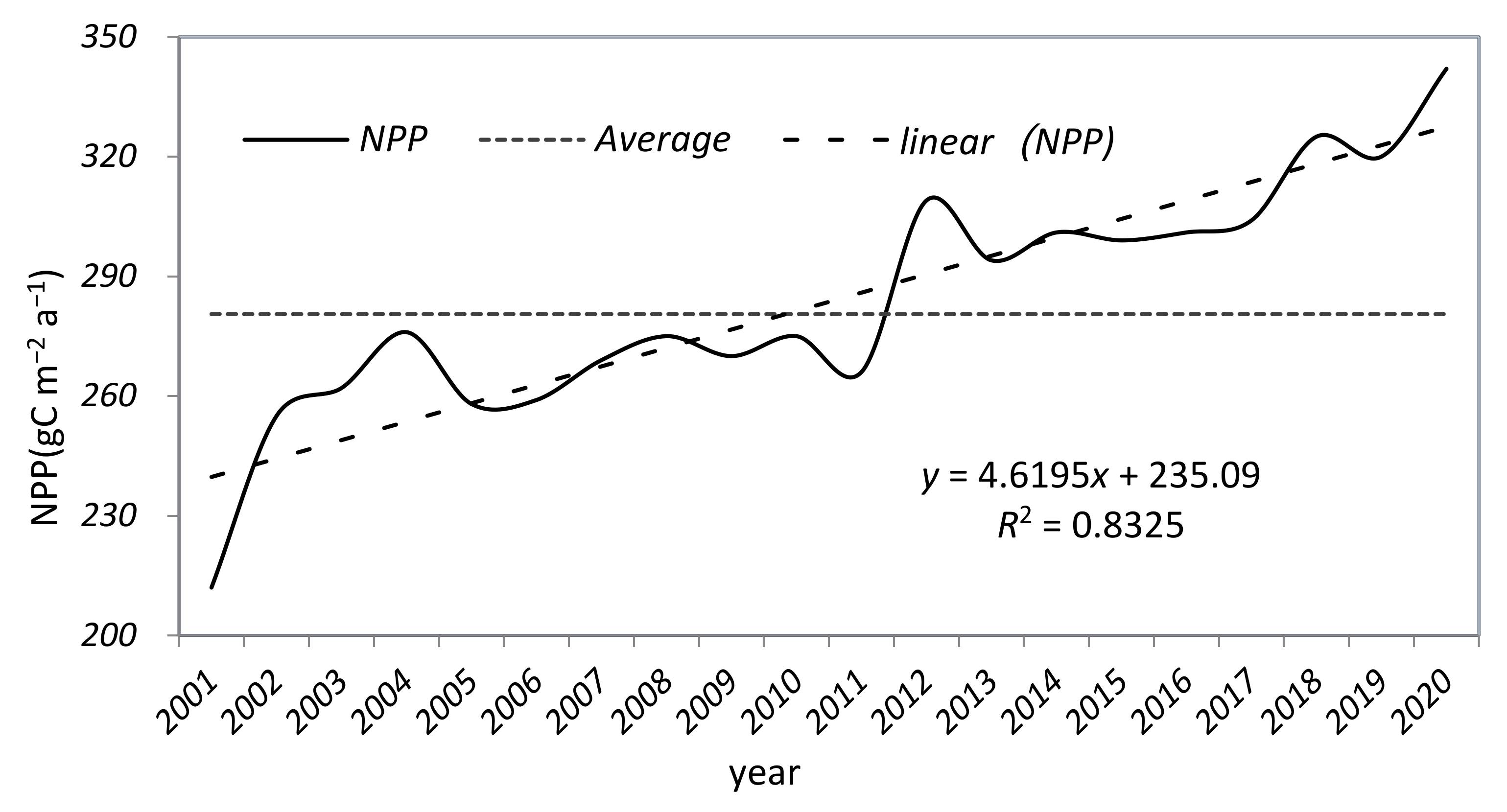
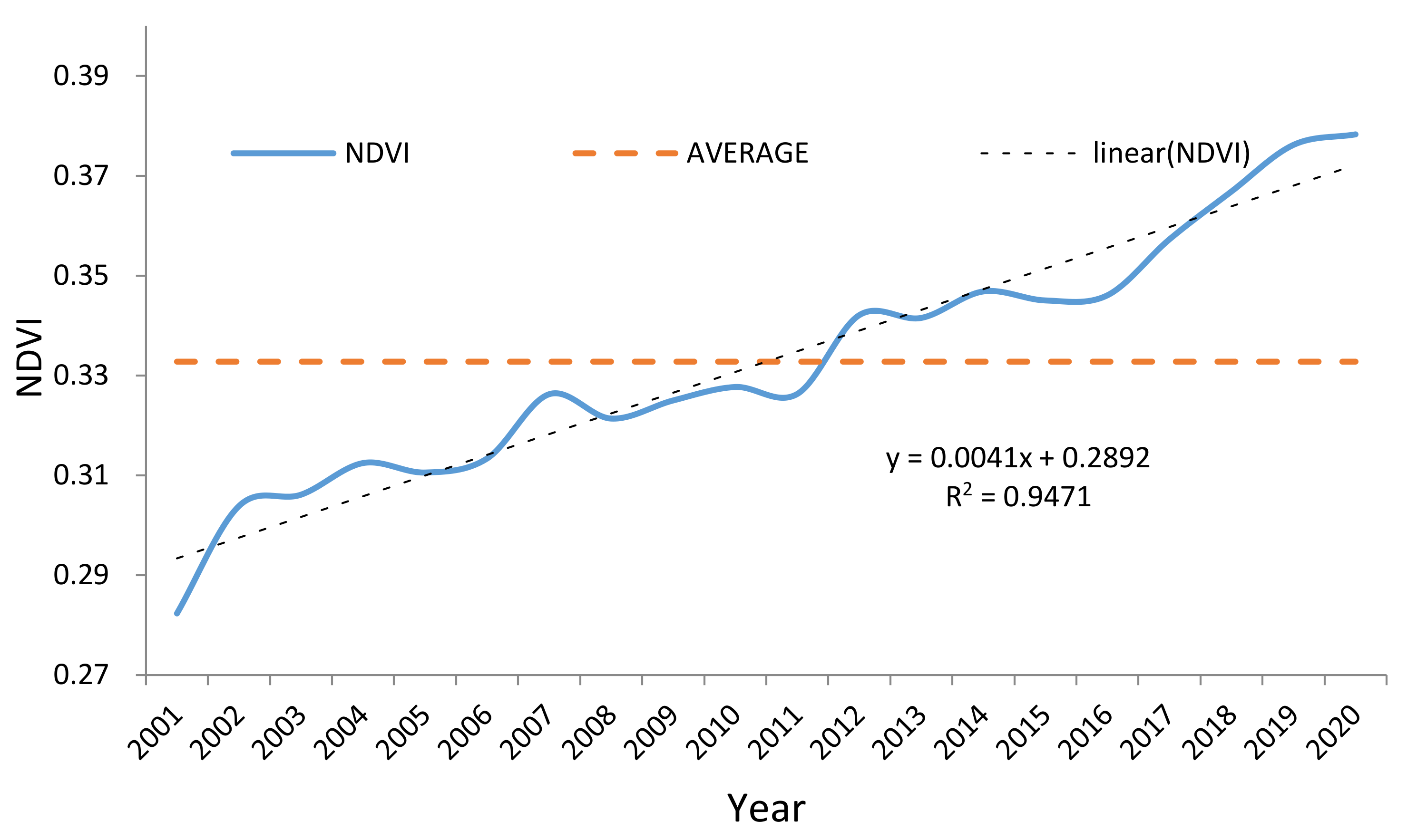
3.3. Spatial and Temporal and Variation of NPP in the YRB
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bargali, S.; Singh, S. Aspect of productivity and nutrient cycling in an 8-year old Eucalyptus plantation in a moist plain area adjacent to Central Himalaya, India. Can. J. For. Res. 1991, 21, 1365–1372. [Google Scholar] [CrossRef]
- Bargali, S.; Singh, S. dynamics, storage and flux of nutrients in an aged eucalypt plantation in Central Himalaya. Oecologia Mont. 1995, 4, 9–14. [Google Scholar]
- Odum, E. Fundamentals of Ecology; Saunders: Philadelphia, PA, USA, 1971. [Google Scholar]
- Liet, H.; Whittaker, R. Primary Productivity of Biosphere (Editors’ Preface); Springer: New York, NY, USA, 1975. [Google Scholar]
- Zhang, L.-X.; Zhou, D.-C.; Fan, J.-W.; Hu, Z.-M. Comparison of four light use efficiency models for estimating terrestrial gross primary production. Ecol. Model. 2015, 300, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Srikanta, S. Modeling terrestrial ecosystem productivity of an estuarine ecosystem in the Sundarban Biosphere Region, India using seven ecosystem models. Ecol. Model. 2017, 356, 73–90. [Google Scholar]
- Liu, Z.; Wang, L.; Wang, S. Comparison of different GPP models in China using MODIS image and Chinaflux data. Remote Sens. 2014, 6, 10215–10231. [Google Scholar] [CrossRef] [Green Version]
- Ruimy, A.; Dedieu, G.; Saugier, B. TURC: A diagnostic model of continental gross primary productivity and net primary productivity. Glob. Biogeochem. Cycles 1996, 10, 269–285. [Google Scholar] [CrossRef]
- Dong, X.; Yang, W.; Ulgiati, U.; Yan, M.; Zhang, X. The impact of human activities on natural capital and ecosystem services of natural pastures in North Xinjiang, China. J. Ecol. Model. 2012, 225, 28–39. [Google Scholar] [CrossRef]
- Gignoux, J.; Fritz, H.; Abbadie, L.; Loreau, M. Which functional processes control the short-term effect of grazing on net primary production in grasslands? Oecologia 2001, 129, 114–124. [Google Scholar]
- Xu, Y.; Xiao, F.; Liao, Y. Assessment of grassland ecosystem service value in response to climate change in China. Diversity 2022, 14, 160. [Google Scholar] [CrossRef]
- Nemani, R.; Keeling, C.; Hashimoto, H. Climate driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Li, X.; Xian, Y.; Chai, Y.; Li, M.; Yang, K.; Qiu, X. Assessing the Net Primary Productivity dynamics of the desert steppe in Northern China during the past 20 Years and its response to climate. Sustainability 2022, 14, 5581. [Google Scholar] [CrossRef]
- Matsushita, B.; Tamura, M. Integrating remotely sensed data with an ecosystem model to estimate net primary productivity in East Asia. Remote Sens. Environ. 2001, 81, 58–66. [Google Scholar] [CrossRef]
- Jones, M.; Running, S.; Kimball, J. Terrestrial primary productivity indicators for inclusion in the National Climate Indicators System. Clim. Chang. 2018, 15, 1855–1868. [Google Scholar] [CrossRef]
- Khalifa, M.; Elagib, N.; Ribbe, L. Spatio-temporal variations in climate, primary productivity and efficiency of water and carbon use of the land cover types in Sudan and Ethiopia. Sci. Total Environ. 2018, 624, 790–806. [Google Scholar] [CrossRef]
- Prince, S.; Goward, S. Global primary production: A remote sensing approach. J. Biogeog. 1995, 22, 815–835. [Google Scholar] [CrossRef]
- Li, T.; Li, M.; Ren, F.; Tian, L. Estimation and Spatio-Temporal Change Analysis of NPP in Subtropical Forests: A Case Study of Shaoguan, Guangdong, China. Remote Sens. 2022, 14, 2541. [Google Scholar] [CrossRef]
- Guo, B.; Wang, S.; Wang, M. Spatio-temporal variation of NPP from 1999 to 2015 in Zoige grassland wetland, China. J. Appl. Ecol. 2020, 31, 424–432. [Google Scholar]
- Potter, C.; Randerson, J.; Field, C.; Matson, P.; Vitousek, P.; Mooney, H.; Klooster, S. Terrestrial ecosystem production: A process model based on global satellite and surface data. Glob. Biogeochem. Cycles 1993, 7, 811–841. [Google Scholar] [CrossRef]
- Field, C.; Randerson, J.; Malmstrom, C. Global net primary production: Combining ecology and remote sensing. Remote Sens. Environ. 1995, 51, 74–88. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Fang, J.; Zhou, L.; Zhu, B.; Tan, K.; Tao, S. Changes in vegetation net primary productivity from 1982 to 1999 in China. Glob. Biogeochem. Cycles 2005, 19, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Zhu, H.; Shi, H.; Liu, X. Spatio-temporal distribution pattern of vegetation net primary productivity and its response to climate change in Buryatiya Republic, Russia. J. Resour. Ecol. 2011, 2, 257–265. [Google Scholar]
- Hao, L.; Wang, S.; Cui, X.; Zhai, Y. Spatiotemporal Dynamics of Vegetation Net Primary Productivity and Its Response to Climate Change in Inner Mongolia from 2002 to 2019. Sustainability 2021, 13, 13310. [Google Scholar] [CrossRef]
- Running, S.; Nemani, R.; Glassy, J. MODIS Daily Photosynthesis and Annual Net Primary Production Product (MOD17) Algorithm Theoretical Basis Document; Version 3.0; NASA: Washington, DC, USA, 1997; pp. 1–59.
- Manral, V.; Bargali, K.; Bargali, S.; Jhariya, M.; Padalia, K. Relationships between soil and microbial biomass properties and annual flux of nutrients in Central Himalayan forests, India. Land Degrad. Dev. 2002, 33, 1–12. [Google Scholar]
- Awasthi, P.; Bargali, K.; Bargali, S.; Jhariya, M. Structure and Functioning of Coriaria nepalensis Wall dominated Shrub lands in degraded hills of Kumaun Himalaya. I. Dry Matter Dynamics. Land Degrad. Dev. 2022, 33, 1474–1494. [Google Scholar] [CrossRef]
- Upper and Middle Yellow River Bureau. Introduction to Water and Soil Conservation in the Yellow River Basin; Yellow River Water Conservancy Press: Zhengzhou, China, 2011. [Google Scholar]
- Xiao, F.; Xu, Y.; Huang, D.; Liao, Y.; Yu, L. Impact of climate change on ecological security of the Yellow River Basin and its adaptation countermeasures. Ren. Yell. Riv. 2021, 43, 10–14. [Google Scholar]
- Zhao, M.; Heinsch, F.; Nemani, R. Improvements of the MODIS terrestrial gross and net primary production global data set. Remote Sens. Environ. 2005, 95, 164–176. [Google Scholar] [CrossRef]
- Turner, D.; Ritts, W.; Cohen, W. Evaluation of MODIS NPP and GPP products across multiple biomes. Remote Sens. Environ. 2006, 102, 282–292. [Google Scholar] [CrossRef]
- Liu, J.; Meng, B.; Ge, J. Spatio-temporal dynamic changes of grassland NPP in Gannan prefecture, as determined by the CASA model. Acta Pratacult. Sin. 2019, 28, 19–32. [Google Scholar]
- Liu, Y.; Zhou, R.; Ren, H. Evaluating the dynamics of grassland net primary productivity in response to climate change in China. Glob. Ecol. Conserv. 2021, 28, e01574. [Google Scholar] [CrossRef]
- Yang, H.; Zhong, X.; Deng, S.; Xu, H. Assessment of the impact of LUCC on NPP and its influencing factors in the Yangtze River Basin, China. CATENA 2021, 206, 105542. [Google Scholar] [CrossRef]
- Bo, Y.; Li, X.; Liu, K.; Wang, S.; Zhang, H.; Gao, X.; Zhang, X. Three Decades of Gross Primary Production (GPP) in China: Variations, Trends, Attributions, and Prediction Inferred from Multiple Datasets and Time Series Modeling. Remote Sens. 2022, 14, 2564. [Google Scholar] [CrossRef]
- Friedl, M.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. MODIS Collection 5 global land cover: Algorithm refinements and characterization of new datasets. Remote Sens. Environ. 2010, 114, 168–182. [Google Scholar] [CrossRef]
- Zhu, W.; Pan, Y.; Zhang, J. Estimation of primary productivity of Chinese terrestrial vegetation on remote sensing. J. Plant Ecol. 2007, 31, 413–424. [Google Scholar]
- Ruimy, A.; Saugier, B.; Dedieu, G. Methodology for the estimation of terrestrial net primary production from remotely sensed data. J. Geophys. Res. Atmos. 1994, 99, 5263–5283. [Google Scholar] [CrossRef]
- Zhu, W.; Pan, Y.; He, H. Simulation of maximum light use efficiency for some typical vegetation types in China. Chin. Sci. Bull. 2006, 51, 457–463. [Google Scholar] [CrossRef]
- Wang, B.; Wang, J.; Yang, Y. Algorithm improvements for two important parameters of FPAR and maximum solar energy utilization efficiency. Acta Pratacult. Sin. 2013, 22, 220–228. [Google Scholar]
- Luo, T.; Pan, Y.; Ouyang, H. Leaf area index and net primary productivity along subtropical to alpine gradients in the Tibetan Plateau. Glob. Ecol. Biogeogr. 2004, 13, 345–358. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.; Nemani, R. Sensitivity of moderate resolution imaging spectroradiometer (MODIS) terrestrial primary production to the accuracy of meteorological reanalyses. J. Geophys. Res. Biogeosci. 2015, 111, 338–356. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Yang, X.; Liu, W.; Chen, F. Spatial distribution of carbon density in grassland vegetation of the Loess Plateau of China. Acta Ecol. Sin. 2012, 32, 0226–0237. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.; Liu, X.; Wang, X.; Li, S. Changes in vegetation coverage and its influencing factors across the Yellow River Basin during 2001-2020. J. Desert Res. 2021, 41, 205–212. [Google Scholar]
- Li, D.; Fan, J.; Wang, J. Variation characteristics of vegetation net primary productivity in Shaanxi Province based on MO17A. Chin. J. Ecol. 2001, 30, 2776–2782. [Google Scholar]
- Zhang, Z.; Chang, J. Spatial-temporal differentiation and eco-economic coordination of vegetation NPP in the Yellow River Basin from 2001 to 2018. J. Huazhong Agri. Univ. 2021, 40, 166–177. [Google Scholar]
- Yang, Z.; Tian, J.; Li, W.; Su, W.; Guo, R.; Liu, W. Spatio-temporal pattern and evolution trend of ecological environment quality in the Yellow River Basin. Acta Ecol. Sin. 2021, 41, 7627–7636. [Google Scholar]
- Lv, M.; Ma, Z.; Peng, S. Responses of terrestrial water cycle components to afforestation within and around the Yellow River basin. Atmos. Ocean. Sci. Lett. 2019, 12, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, J.; Liu, P.; Lu, G.; Li, Y. Climatic Changes Dominant Interannual Trend in Net Primary Productivity of Alpine Vulnerable Ecosystems. J. Resour. Ecol. 2019, 10, 379–388. [Google Scholar]
- Zhang, Z.; Chang, T.; Qiao, X.; Yang, Y.; Guo, J.; Zhang, H. Eco-Economic Coordination Analysis of the Yellow River Basin in China: Insights from Major Function-Oriented Zoning. Sustainability 2021, 13, 2715. [Google Scholar] [CrossRef]
- Jiang, C.; Wang, D.; Luo, S.; Li, D.; Zhang, L.; Gao, Y. Ecosystem Status Changes and Attribution in the Three-River Headwaters Region. Res. Environ. Sci. 2017, 30, 10–19. [Google Scholar]
- Yuan, L.; Jiang, W.; Shen, W. The spatio-temporal variations of vegetation cover in the Yellow River Basin from 2000 to 2010. Acta Ecol. Sin. 2013, 33, 7798–7806. [Google Scholar]
- Chang, T.; Zhang, Z.; Qiao, X.; Zhang, Y. Land use transformation and its eco-environment effects of ecological- production-living spaces in Yellow River Basin. Bull. Soi. Wat. Cons. 2021, 41, 268–275. [Google Scholar]
- Yang, S.; Liu, C.; Sun, R. The vegetation covers over last 20 years in Yellow River Basin. Acta Geogr. Sin. 2002, 57, 679–684. [Google Scholar]
- Cheng, Q.; Chen, Y.; Wang, M. Change of vegetation net primary productivity in Yellow River watersheds from 2001 to 2010 and its climatic driving factors analysis. Chin. Appl. Ecol. 2014, 25, 2811–2818. [Google Scholar]
- Wang, P.; Xie, D.; Zhou, Y.; Youhao, E.; Zhu, Q. Estimation of net primary productivity using a process-based model in Gansu Province, Northwest China. Environ. Earth Sci. 2014, 71, 647–658. [Google Scholar] [CrossRef]
- Chao, Q.; Yan, Z.; Sun, Y. A recent scientific understanding of climate change in China. China Popul. Resour. Environ. 2020, 30, 1–9. [Google Scholar]
- Wu, G.; Wang, W. Regionalization and Revegetation in the Agricultural and Pasturing Interlaced Zone of China. J. Desert Res. 2022, 22, 439–442. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Maximum LUE εmax (gC MJ–1) |
|---|---|
| Evergreen coniferous forest | 0.389 |
| Evergreen broad-leaved forests | 0.985 |
| Deciduous, coniferous forest | 0.485 |
| Deciduous broad-leaved forest | 0.692 |
| Coniferous and broad-leaved mixed forest | 0.475 |
| Evergreen, deciduous broad-leaved mixed forest | 0.768 |
| Grassland | 0.542 |
| Cultivated vegetation | 0.542 |
| Shrub | 0.429 |
| Other vegetation | 0.542 |
| Ecosystem Types | Area (km2) | NPP Mean (gC m–2 a–1) | NPP Max gC m–2 a–1) | NPP SD (gC m–2 a–1) | NPP Total Annual (TgC) |
|---|---|---|---|---|---|
| Forest | 105,778 | 401.7 | 1124 | 201.4 | 42.5 |
| Grassland | 378,984 | 276.0 | 1057 | 185.8 | 105 |
| Farmland | 213,267 | 303.9 | 1042 | 130.7 | 64.8 |
| Wetland | 7968 | 498.1 | 710 | 192.8 | 3.96 |
| Desert | 63,920 | 70.9 | 823 | 67.4 | 4.53 |
| Total | 283.4 | 1124 | 182.2 | 220 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, F.; Liu, Q.; Xu, Y. Estimation of Terrestrial Net Primary Productivity in the Yellow River Basin of China Using Light Use Efficiency Model. Sustainability 2022, 14, 7399. https://doi.org/10.3390/su14127399
Xiao F, Liu Q, Xu Y. Estimation of Terrestrial Net Primary Productivity in the Yellow River Basin of China Using Light Use Efficiency Model. Sustainability. 2022; 14(12):7399. https://doi.org/10.3390/su14127399
Chicago/Turabian StyleXiao, Fengjin, Qiufeng Liu, and Yuqing Xu. 2022. "Estimation of Terrestrial Net Primary Productivity in the Yellow River Basin of China Using Light Use Efficiency Model" Sustainability 14, no. 12: 7399. https://doi.org/10.3390/su14127399
APA StyleXiao, F., Liu, Q., & Xu, Y. (2022). Estimation of Terrestrial Net Primary Productivity in the Yellow River Basin of China Using Light Use Efficiency Model. Sustainability, 14(12), 7399. https://doi.org/10.3390/su14127399