
Swimming Performance of Four Carps on the Yangtze River for Fish Passage Design

Abstract
1. Introduction
2. Material and Methods
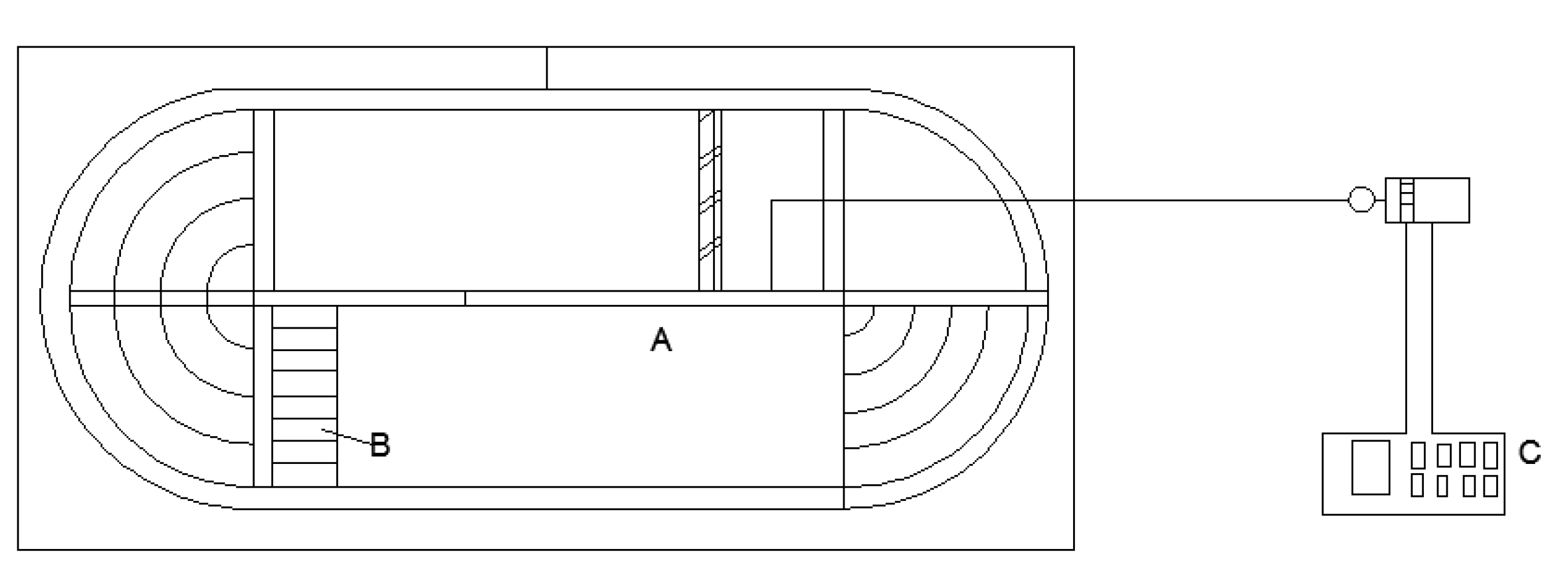
2.1. Experimental Setup and Fish
2.2. Test Methods
2.3. Statistical Analysis
3. Results
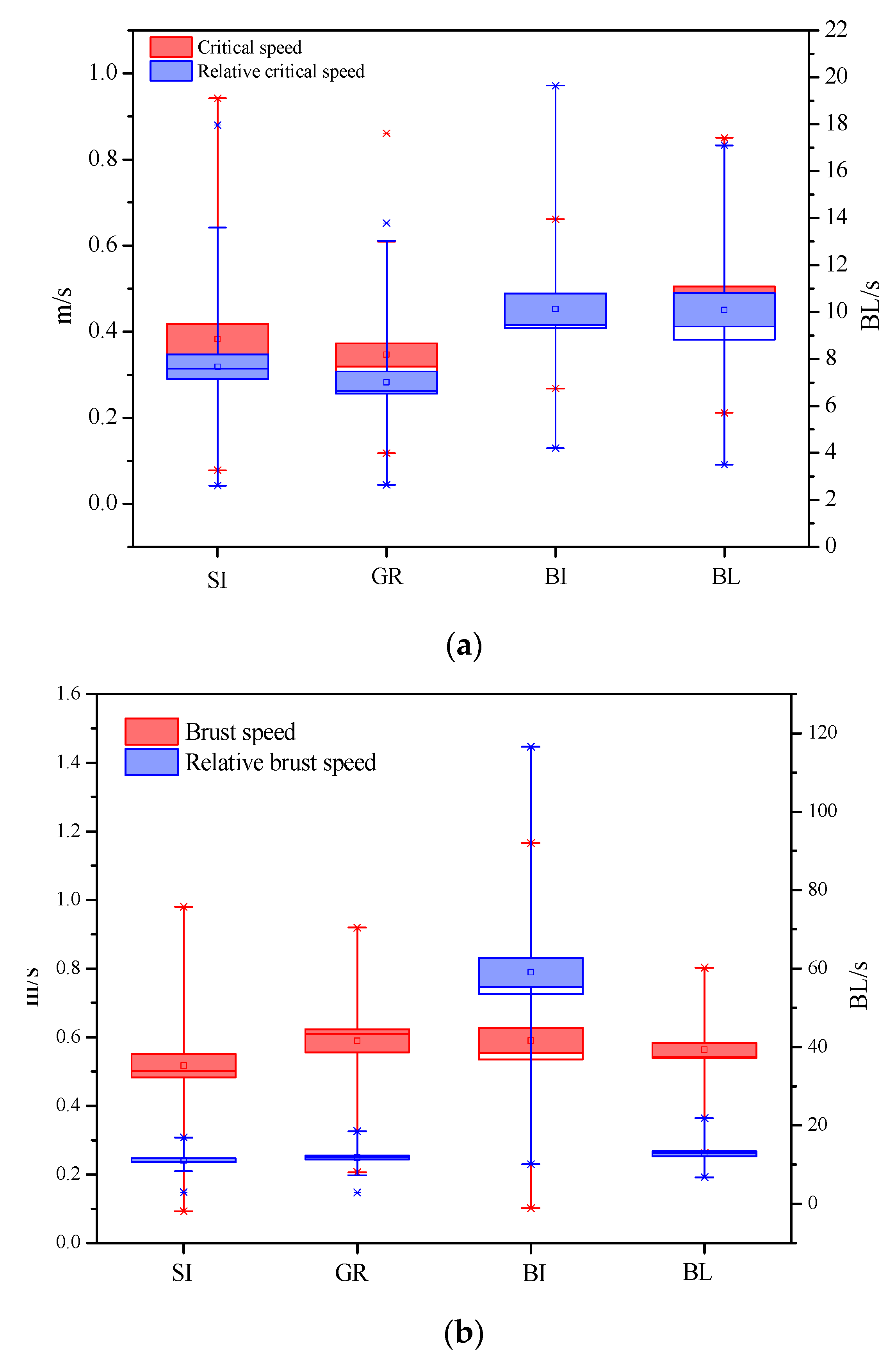
3.1. Critical Swimming Speed
3.2. Burst Swimming Speed
4. Discussion
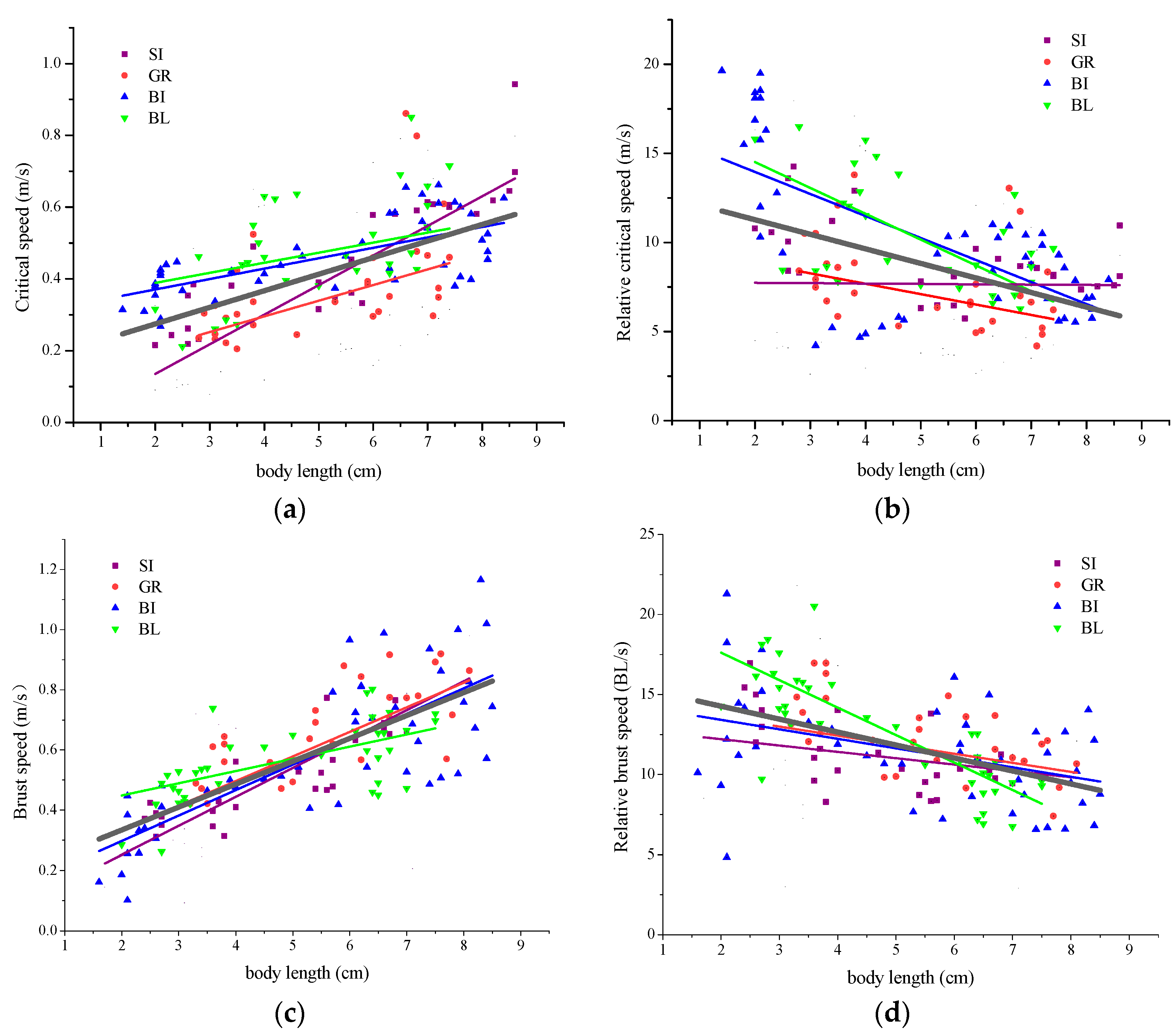
4.1. Relationship between Body Length and Swimming Capability
4.2. Comparison of Swimming Capability of Four Carps
4.3. The Relationship between Critical Swimming Speed and Burst Swimming Speed
4.4. Applications for Fish Passage
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, J.; Gao, Z.; Dai, H.; Shi, X. Effects of turbulence and velocity on the movement behaviour of bighead carp (Hypophthalmichthys nobilis) in an experimental vertical slot fishway. Ecol. Eng. 2019, 127, 363–374. [Google Scholar] [CrossRef]
- Newbold, L.R.; Shi, X.; Hou, Y.; Han, D.; Kemp, P.S. Swimming performance and behaviour of bighead carp (Hypophthalmichthys nobilis): Application to fish passage and exclusion criteria. Ecol. Eng. 2016, 95, 690–698. [Google Scholar] [CrossRef]
- Hu, M.; Hua, Q.; Zhou, H.; Wu, Z.; Wu, X. The effect of dams on the larval abundance and composition of four carp species in key river systems in China. Environ. Biol. Fishes 2015, 98, 1201–1205. [Google Scholar] [CrossRef]
- Katopodis, C.; Williams, J.G. The development of fish passage research in a historical context. Ecol. Eng. 2012, 48, 8–18. [Google Scholar] [CrossRef]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Cooke, S.J. The future of fish passage science, engineering, and practice. Fish Fish. 2018, 19, 340–362. [Google Scholar] [CrossRef]
- Li, M.; Shi, X.; Jin, Z.; Ke, S.; Lin, C.; An, R.; Li, J.; Katopodis, C. Behaviour and ability of a cyprinid (Schizopygopsis younghusbandi) to cope with accelerating flows when migrating downstream. River Res. Appl. 2020, 1–12. [Google Scholar] [CrossRef]
- Brett, J. The respiratory metabolism and swimming performance of young sockeye salmon. Can. J. Fish Aquat. Sci. 1964, 21, 1183–1226. [Google Scholar] [CrossRef]
- Eliason, E.J.; Clark, T.D.; Hague, M.J.; Hanson, L.M.; Gallagher, Z.S.; Jeffries, K.M.; Gale, M.K.; Patterson, G.A.; Hinch, S.G.; Farrell, A.P. Differences in thermal tolerance among sockeye salmon populations. Science 2011, 332, 109–112. [Google Scholar] [CrossRef]
- Cano‑Barbacil, C.; Radinger, J.; Argudo, M.; Rubio‑Gracia, F.; Vila‑Gispert, A.; García‑Berthou, E. Key factors explaining critical swimming speed in freshwater fish: A review and statistical analysis for Iberian species. Sci. Rep. 2020, 10, 18947. [Google Scholar] [CrossRef]
- Cai, L.; Fang, M.; Tu, Z.; Liu, G.; Shi, X.; Huang, Y. Research progress on fish swimming characteristics related to fish migration. J. Wuhan Univ. 2013, 59, 363–368. [Google Scholar]
- Gong, L.; Wu, Y.; Bai, Y.; Mu, X. Experimental study on swimming ability and swimming behavior of juvenile grass carp. J. Chin. Inst. Water Resour. Hydropower Res. 2015, 13, 211–216. [Google Scholar] [CrossRef]
- Hou, Y.; Newbold, L.; Cai, L.; Wang, X.; Hu, W.; Qiao, Y. Swimming performance of juvenile Aristichthys nobilis under fixed velocity swimming tests. Chin. J. Ecol. 2016, 35, 1583–1588. [Google Scholar] [CrossRef]
- Li, H.; Cai, D.; Yang, P. Swimming ability and behavior of different sized silver carp. J. Hydroecol. 2016, 37, 88–92. [Google Scholar] [CrossRef]
- Peake, S.J.; Beamish, F.W.; McKinley, R.S.; Scruton, D.A.; Katopodis, C. Relating swimming performance of lake sturgeon, Acipenser fulvescens, to fishway desing. Can. J. Fish Aquat. Sci. 1997, 54, 1361–1366. [Google Scholar] [CrossRef]
- Deslauriers, D.; Kieffer, J.D. The influence of flume length and group size on swimming performance in shortnose sturgeon Acipenser brevirostrum. J. Fish Biol. 2011, 79, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Cai, L.; Wang, X.; Chen, X.; Zhu, D.; Johnson, D.; Shi, X. Swimming performance of 12 Schizothoracinae species from five rivers. J. Fish Biol. 2018, 92, 2022–2028. [Google Scholar] [CrossRef]
- Castro, M.A.; Santos, H.A.; Sampalo, A.C.; Pompeu, P.S. Swimming performance of the small characin Bryconamericus stramineus (Characiformes: Characidae). Zoologia 2010, 27, 939–944. [Google Scholar] [CrossRef]
- Smit, H.; Amelink-Kouutsataal, J.M.; Vijverberg, J.; von Vaupel-Klein, J.C. Oxygen consumption and efficiency of swimming goldfish. Comp. Biochem. Physiol. 1971, 39, 1–28. [Google Scholar] [CrossRef]
- Laborde, A.; González, A.; Sanhueza, C.; Arriagada, P.; Wilkes, M.; Habit, E.; Link, O. Hydropower Development, Riverine Connectivity, and Non-sport Fish Species: Criteria for Hydraulic Design of Fishways. River Res. Appl. 2016, 32, 1949–1957. [Google Scholar] [CrossRef]
- Poletto, J.B.; Cocherell, D.E.; Ho, N.; Cech, J.J.J.; Klimley, A.P.; Fangue, N.A. The effect of size on juvenile green sturgeon (Acipenser medirostris) behavior near water-diversion fish screens. Environ. Biol. Fish 2018, 101, 67–77. [Google Scholar] [CrossRef]
- Cai, L.; Katopodis, C.; Johnson, D.; Zhang, P.; Zhao, P. Case study: Targeting species and applying swimming performance data to fish lift design for the Huangdeng Dam on the upper Mekong River. Ecol. Eng. 2018, 122, 32–38. [Google Scholar] [CrossRef]
- Cooke, S.J.; Cech, J.J.; Glassman, D.M.; Simard, J.; Louttit, S.; Lennox, R.J.; Cruz-Font, L.; O’Connor, C.M. Water resource development and sturgeon (Acipenseridae): State of the science and research gaps related to fish passage, entrainment, impingement and behavioural guidance. Rev. Fish Biol. Fish. 2020, 30, 7755. [Google Scholar] [CrossRef]
- Hammer, C. Fatigue and exercise tests with fish. Comp. Biochem. Physiol. Part A Physiol. 1995, 112, 1–20. [Google Scholar] [CrossRef]
- Cai, L.; Hou, Y.; Katopodis, C.; He, D.; Johnson, D.M.; Zhang, P. Rheotaxis and swimming performance of Perch-barbel (Percocypris pingi, Tchang, 1930) and application to design of fishway entrances. Ecol. Eng. 2019, 132, 102–108. [Google Scholar] [CrossRef]
- Farrell, A.P. Comparisons of swimming performance in rainbow trout using constant acceleration and critical swimming speed tests. J. Fish Biol. 2008, 72, 693–710. [Google Scholar] [CrossRef]
- Plaut, I. Critical swimming speed: Its ecological relevance. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 131, 41–50. [Google Scholar] [CrossRef]
- Xiong, F.; Wang, C.; Liu, D.; Mo, W.; Liu, W.; Shi, X. Comparative study of burst swimming speed of black carp, grass carp, silver carp and bighead carp from Songhua River. Ecol. Sci. 2014, 33, 339–343. [Google Scholar]
- Liu, H.; Wang, C.; Zhu, L.; Chen, M. Comparative study of critical swimming speed for juvenile silver and bighead carp. J. Hydroecol. 2016, 37, 63–69. [Google Scholar]
- Fu, X.; Fu, C.; Fu, S. Comparison of swimming ability among five freshwater fish species. Chin. J. Ecol. 2020, 39, 1629–1635. [Google Scholar] [CrossRef]
- Kern, P.; Cramp, R.L.; Gordos, M.A.; Waston, J.R.; Franklin, C.E. Measuring Ucrit and endurance: Equipment choice influences estimates of fish swimming performance. J. Fish Biol. 2018, 92, 237–247. [Google Scholar] [CrossRef]
- Ojanguren, A.F.; Brana, F. Effects of size and morphology on swimming performance in juvenile brown trout (Salmo trutta L). Ecol. Freshw. Fish 2003, 12, 241–246. [Google Scholar] [CrossRef]
- Norin, T.; Clark, T.D. Measurement and relevance of maximum metabolic rate in fishes. J. Fish Biol. 2016, 88, 122–151. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; He, X.; Cao, Z.; Fu, S. The trade-off between steady and unsteady swimming performance in six cyprinids at two temperatures. J. Therm. Biol. 2012, 37, 424–431. [Google Scholar] [CrossRef]
- Losos, J.B.; Walton, B.M.; Bennett, A.F. Trade-offs between sprinting and clinging ability in Kenyan chameleons. Funct. Ecol. 1993, 7, 281–286. [Google Scholar] [CrossRef]
- Langerhans, R.B. Trade-off between steady and unsteady swimming underlies predator-driven divergence in Gambusiaaffinis. J. Evol. Biol. 2009, 22, 1057–1075. [Google Scholar] [CrossRef] [PubMed]
- Lyon, J.P.; Ryan, T.J.; Scroggie, M.P. Effects of temperature on the fast-start swimming performance of an Australian freshwater fish. Ecol. Freshw. Fish 2008, 17, 184–188. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Swimming Index | Fish Species | No. of Tested Fish | Weight (g) | Body Length (cm) | Total Length (cm) |
|---|---|---|---|---|---|
| Burst swimming speed | BL | 32 | 4.72 ± 0.13 | 6.63 ± 1.12 | 7.17 ± 1.31 |
| GR | 36 | 5.25 ± 0.20 | 7.23 ± 1.05 | 7.86 ± 1.37 | |
| SI | 39 | 5.46 ± 0.15 | 7.36 ± 1.12 | 7.97 ± 1.33 | |
| BI | 46 | 6.30 ± 0.21 | 7.52 ± 0.91 | 8.12 ± 1.30 | |
| Critical swimming speed | BL | 41 | 4.53 ± 0.25 | 6.80 ± 1.04 | 7.46 ± 1.23 |
| GR | 36 | 5.57 ± 0.21 | 6.52 ± 1.17 | 7.55 ± 1.32 | |
| SI | 32 | 5.73 ± 0.19 | 6.98 ± 1.06 | 7.82 ± 1.22 | |
| BI | 46 | 6.23 ± 0.23 | 7.51 ± 1.14 | 8.42 ± 1.16 |
| Fish Species | Body Length of Fish | Critical Swimming Speed (m/s) | Relative Critical Swimming Speed (BL/s) |
|---|---|---|---|
| BL | 2–5 cm (n = 16) | 0.42 ± 0.33 | 11.20 ± 7.75 |
| 5–8 cm (n = 16) | 0.47 ± 0.33 | 7.09 ± 4.85 | |
| GR | 2–5 cm (n = 16) | 0.27 ± 0.20 | 8.02 ± 5.42 |
| 5–8 cm (n = 20) | 0.40 ± 0.35 | 6.14 ± 5.12 | |
| SI | 1–5 cm (n = 17) | 0.23 ± 0.21 | 8.54 ± 7.67 |
| 5–9 cm (n = 22) | 0.48 ± 0.11 | 7.13 ± 2.36 | |
| BI | 1–5 cm (n = 21) | 0.33 ± 0.21 | 15.68 ± 9.76 |
| 5–9 cm (n = 25) | 0.51 ± 0.31 | 7.10 ± 4.26 | |
| Fish Species | Body Length of Fish | Burst Swimming Speed (m/s) | Relative Burst Swimming Speed (BL/s) |
| BL | 2–5 cm (n = 21) | 0.52 ± 0.39 | 16.37 ± 8.47 |
| 5–8 cm (n = 20) | 0.57 ± 0.48 | 8.58 ± 6.29 | |
| GR | 2–5 cm (n = 19) | 0.48 ± 0.24 | 14.03 ± 7.45 |
| 5–9 cm (n = 17) | 0.76 ± 0.39 | 11.13 ± 7.23 | |
| SI | 1–5 cm (n = 17) | 0.38 ± 0.14 | 11.87 ± 3.51 |
| 5–9 cm (n = 15) | 0.69 ± 0.44 | 9.89 ± 5.89 | |
| BI | 1–5 cm (n = 17) | 0.34 ± 0.20 | 13.23 ± 8.24 |
| 5–9 cm (n = 29) | 0.69 ± 0.44 | 9.89 ± 5.89 |
| Speed Indexes | UCrit | UCrit’ |
| BL | R2 = 0.335, p < 0.05 | R2 = 0.323, p < 0.05 |
| GR | R2 = 0.325, p < 0.05 | R2 = 0.350, p < 0.05 |
| SI | R2 = 0.791, p < 0.05 | R2 = 0.316, p < 0.05 |
| BI | R2 = 0.461, p < 0.05 | R2 = 0.421, p < 0.05 |
| Speed Indexes | Uburst | Uburst’ |
| BL | R2 = 0.415, p <0.05 | R2 = 0.643, p < 0.05 |
| GR | R2 = 0.543, p < 0.05 | R2 = 0.382, p < 0.05 |
| SI | R2 = 0.722, p < 0.05 | R2 = 0.397, p < 0.05 |
| BI | R2 = 0.603, p < 0.05 | R2 = 0.63, p < 0.05 |
| Speed Index | Fish Species | Two Categories Lengths of Fish | p-Value | |
|---|---|---|---|---|
| Critical swimming speed for fish | BL | 2–5 cm (n = 16) | 5–8 cm (n = 16) | p < 0.05 |
| GR | 2–5 cm (n = 16) | 5–8 cm (n = 20) | p < 0.05 | |
| SI | 1–5 cm (n = 17) | 5–9 cm (n = 22) | p < 0.05 | |
| BI | 1–5 cm (n = 21) | 5–9 cm (n = 25) | p < 0.05 | |
| Burst swimming speed for fish | BL | 2–5 cm (n = 21) | 5–8 cm (n = 20) | p < 0.05 |
| GR | 2–5 cm (n = 19) | 5–9 cm (n = 17) | p < 0.05 | |
| SI | 1–5 cm (n = 17) | 5–9 cm (n = 15) | p < 0.05 | |
| BI | 1–5 cm (n = 17) | 5–9 cm (n = 29) | p < 0.05 | |
| Speed Index | Different Fish Species | p-Value |
|---|---|---|
| Critical swimming speed | BI and SI | p < 0.05 |
| BL and GR | p < 0.05 | |
| BI and BL | p > 0.05 | |
| BI and GR | p > 0.05 | |
| SI and GR | p > 0.05 | |
| SI and BL | p > 0.05 | |
| Burst swimming speed | BI and SI | p < 0.05 |
| BL and GR | p < 0.05 | |
| BI and BL | p > 0.05 | |
| BI and GR | p > 0.05 | |
| SI and GR | p > 0.05 | |
| SI and BL | p < 0.05 |
| Fish Species | Temperature (°C) | Body Length (cm) | Ucrit’ (BL/s) | Ucrit (m/s) | Uburst’ (BL/s) | Uburst (m/s) | References |
|---|---|---|---|---|---|---|---|
| BL | 25 ± 1 | 6.81 ± 0.05 | / | 0.45 ± 0.05 | / | / | [29] |
| 23 ± 0.8 | 6.80 ± 1.04 | 9.14 ± 4.82 | 0.44 ± 0.22 | / | / | In our study | |
| 21 ± 2 | 19.0 ± 6.09 | / | / | 5.67 ± 1.22 | / | [27] | |
| 23 ± 0.8 | 6.63 ± 1.12 | / | / | 8.46 ± 2.13 | 0.59 ± 0.14 | In our study | |
| GR | 28 ± 1 | 9.8 ± 0.80 | 8.88 ± 0.50 | 0.87 ± 0.10 | / | / | [11] |
| 28 ± 1 | 12.3 ± 0.80 | 7.71 ± 0.82 | 0.95 ± 0.05 | / | / | [11] | |
| 23 ± 0.8 | 6.52 ± 1.17 | 6.97 ± 2.83 | 0.34 ± 0.16 | / | / | In our study | |
| 21+2 | 21.57 ± 6.10 | / | / | 5.98 ± 1.14 | / | [27] | |
| 23 ± 0.8 | 7.23 ± 1.05 | / | / | 10.79 ± 2.24 | 0.71 ± 0.07 | In our study | |
| SI | 20 ± 1 | 10.11 ± 0.24 | 6.45 ± 0.27 | 0.66 ± 0.02 | / | / | [28] |
| 20 ± 1 | 16.90 ± 0.55 | 4.97 ± 0.14 | 0.84 ± 0.03 | / | / | [28] | |
| 30 ± 1 | 10.09 ± 0.53 | 6.69 ± 0.11 | 0.67 ± 0.03 | / | / | [13] | |
| 30 ± 1 | 11.84 ± 0.67 | 6.45 ± 0.08 | 0.76 ± 0.03 | / | / | [13] | |
| 23 ± 0.8 | 6.98 ± 1.06 | 10.49 ± 5.98 | 0.38 ± 0.14 | / | / | In our study | |
| 21 ± 2 | 20.70 ± 7.63 | / | / | 6.60 ± 1.62 | / | [27] | |
| 23 ± 0.8 | 7.36 ± 1.12 | / | / | 11.07 ± 1.16 | 0.73 ± 0.06 | In our study | |
| BI | 20 ± 1 | 10.78 ± 1.34 | 7.57 ± 0.62 | 0.81 ± 0.04 | / | / | [28] |
| 20 ± 1 | 15.39 ± 0.62 | 5.66 ± 0.23 | 0.87 ± 0.03 | / | / | [28] | |
| 20 ± 1 | 19.99 ± 1.58 | 4.60 ± 3.54 | 0.92 ± 0.04 | / | / | [28] | |
| 23 ± 0.8 | 7.51 ± 1.14 | 10.49 ± 5.98 | 0.38 ± 0.14 | / | / | In our study | |
| 21 ± 2 | 21.29 ± 7.01 | / | / | 5.05 ± 9.04 | / | [27] | |
| 23 ± 0.8 | 7.52 ± 0.91 | / | / | 9.91 ± 1.76 | 0.65 ± 0.11 | In our study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, J.; Li, H.; Guo, W.; Tan, H.; Ke, S.; Wang, J.; Shi, X. Swimming Performance of Four Carps on the Yangtze River for Fish Passage Design. Sustainability 2021, 13, 1575. https://doi.org/10.3390/su13031575
Tan J, Li H, Guo W, Tan H, Ke S, Wang J, Shi X. Swimming Performance of Four Carps on the Yangtze River for Fish Passage Design. Sustainability. 2021; 13(3):1575. https://doi.org/10.3390/su13031575
Chicago/Turabian StyleTan, Junjun, Hong Li, Wentao Guo, Honglin Tan, Senfan Ke, Jibao Wang, and Xiaotao Shi. 2021. "Swimming Performance of Four Carps on the Yangtze River for Fish Passage Design" Sustainability 13, no. 3: 1575. https://doi.org/10.3390/su13031575
APA StyleTan, J., Li, H., Guo, W., Tan, H., Ke, S., Wang, J., & Shi, X. (2021). Swimming Performance of Four Carps on the Yangtze River for Fish Passage Design. Sustainability, 13(3), 1575. https://doi.org/10.3390/su13031575